Introduction
Neuroblastoma is the most common extracranial solid
tumor in children and accounts for 15% of all childhood related
cancer deaths (1). Neuroblastoma
is clinically heterogeneous, with a subset of patients showing
spontaneous tumor regression and another subset of patients showing
aggressive progression. Despite intensive multimodal therapy which
also includes radiation, the survival rates of patients with
high-risk neuroblastoma remain poor (2). Resistance to these therapies and the
subsequent aggressive behavior of tumors can be attributed to
several factors which include increased migration, invasion and
angiogenesis (3). The role of
vascular endothelial growth factors (VEGFs) in angiogenesis has
been well characterized. Of the several members of the VEGF family,
VEGF-A is the best characterized and is considered to be the
fundamental mediator of pathologic angiogenesis (4). VEGF-A, aka VEGF is secreted by tumor
cells, binds with high affinity to vascular endothelial growth
factor receptor 2 (VEGFR2) on endothelial cells, initiates receptor
dimerization mediating its phosphorylation, which leads to
activation of downstream signaling cascades. This cascade includes
key players such as extracellular signal-regulated kinase 1 and 2
(ERK1/2), p38 kinase, and c-Jun N-terminal kinase/stress-activated
protein kinase (JNK/SAPK). These players belong to the family of
mitogen-activated protein kinases (MAPKs) that promote cell growth
and proliferation (5).
Therapeutic strategies for treatment of
neuroblastoma have evolved over the years and are generally based
on risk grade. The current treatment strategy for high-risk
neuroblastoma is multi-modal and is divided into three phases:
Induction phase, consolidation phase and the maintenance phase
(6). i) Induction phase includes
high dose chemotherapy using cisplatin and etoposide alternating
with vincristine, cyclophosphamide and doxorubicin (7). ii) Consolidation phase includes
surgical resection, myeloablative chemotherapy followed by
hematopoietic stem cell rescue/transplantation (HSCT), and further
followed by radiation. Radiation therapy is given to control local
metastatic sites when appropriate (8). iii) The maintenance phase is aimed at
targeting residual tumor cells with isotretinoin in combination
with immunotherapy such as anti-GD2 antibodies combined with
interleukin-2 and GM-CSF. This multimodality approach has been
shown to improve event-free survival (EFS) (9) in most cases, however, refractory
cases still remain.
Secreted protein acidic and rich in cysteine (SPARC,
also known as osteonectin; or basement-membrane-40, BM-40) belongs
to a group of non-structural components of the extra-cellular
matrix (ECM) that modulate interactions between cells and their
micro-environment. It is highly expressed in a variety of cell
types and is associated with tissue remodeling. The role of SPARC
in tumorigenesis appears to be dichotomous. In some types of cancer
such as breast (10,11), ovarian (12) and gastric cancers (13), high levels of SPARC expression has
been shown to correlate with disease progression and poor
prognosis. In other types of cancer such as prostate (14), acute myelogenous leukemia (15) and neuroblastoma (16), SPARC functions as a tumor
suppressor, making SPARC both a pro- and an anti-oncogenic
molecule. In neuroblastomas there is an inverse correlation between
SPARC expression and tumor stage (17). Studies have demonstrated that
tumorigenic neuroblastoma cell lines showed low or undetectable
levels of SPARC, while non-tumorigenic cells expressed high levels
of SPARC (18,19), making SPARC overexpression a viable
option for neuroblastoma treatment. In addition, SPARC has been
shown to have anti-angiogenic properties including the ability to
inhibit proliferation and migration of endothelial cells stimulated
by bFGF and VEGF (17). This study
was aimed at deciphering one of the mechanisms by which SPARC
over-expression mediates angiogenic suppression. Here we present
our findings that show combination of SPARC overexpression
accompanied with radiation reduces VEGF-A induced angiogenesis via
the VEGF-A targeting microRNA miR-410 in NB1691 and SK-N-BE(2)
neuroblastoma cells.
Materials and methods
Human neuroblastoma tissue array
Human neuroblastoma tissue arrays were obtained from
US Biomax, Inc. (Rockville, MD, USA). The array consisted of 61
cases of neuroblastoma cancer and normal tissues with stage and
grade information. Tissue arrays were processed for
immunohistochemistry as per previously described standard protocol
(20). Briefly, slides were
deparaffinized and endogenous peroxidases activity was blocked at
room temperature by 5–10 min incubation in 0.3%
H2O2 in PBS (pH 7.7). The slides were then
rinsed in PBS for 5 min, followed by antigen retrieval by heating
at 95°C in citrate buffer (0.01 mol/l sodium citrate buffer, pH
6.0) for 10 min. The tissues were blocked using normal goat serum
for 20 min at room temperature and incubated in primary antibody
(anti-VEGF or SPARC) for 60 min at room temperature. The slides
were then rinsed in PBS followed by the addition of a horseradish
peroxidase (HRP)-conjugated secondary antibody for 20 min at room
temperature. HRP substrate DAB (3,3′-diaminobenzidine
tetrahydrochloride) at 1 mg/ml in 50 mmol/l Tris, pH 7.2, and 0.3%
H2O2 was then added for the development of
DAB substrate followed by haematoxylin staining. Next, the slides
were dehydrated and mounted with coverslips.
Cell lines and culture conditions
Neuroblastoma cell line SK-N-BE(2) [obtained from
the American Type Culture Collection (ATCC; Manassas, VA, USA)] was
obtained, characterized and authenticated by the ATCC, low passage
cells were used for experiments in all the cases (less than 6
months of passages) and NB1691 (provided by Peter Houghton, St.
Jude Children's Research Hospital, Memphis, TN, USA) were
characterized and authenticated by the providers institution (less
than 6 months of passages) were cultured in Opti-MEM medium with 5%
fetal bovine serum (FBS) and 1% penicillin/streptomycin in a
humidified 5% atmospheric CO2 at 37°C. Endothelial cells
were cultured using the endothelial cell growth Medium-200 (Thermo
Fisher Scientific, Waltham, MA, USA) with 10 ng/ml EGF, 10 ng/ml
FGF, 10 ng/ml heparin, and 1 μg/ml of hydrocortisone, 2% FBS and 1%
penicillin/streptomycin in a humidified 5% CO2
atmosphere at 37°C with Calcein AM Dye (Thermo Fisher Scientific)
as per manufacturer's instructions.
SPARC overexpression
SK-N-BE(2) and NB1691 cells were transfected with
SPARC overexpression plasmid RC209964 (Origene Technologies, Inc.,
Rockville, MD, USA) as per standard protocols using Lipofectamine
with 10 μg of plasmid on 60% confluent plates. SPARC overexpression
was also validated in vitro using SPARC Adenovirus (Human)
from Applied Biological Materials Inc., (Richmond, BC, Canada) on
60% confluent plates at 50 MOI infection. Empty adenovirus was used
as control. SPARC expression was validated by western blot
analysis.
Antibodies
Antibodies were obtained from the following sources:
SPARC (Aviva Systems Biology Corp., San Diego, CA, USA), VEGF-A and
β-actin (Santa Cruz Biotechnology, Dallas, TX, USA).
Western blotting
SK-N-BE(2) and NB1691 cells were transfected with
SPARC overexpression plasmid. After 24 h, cells were treated with
or without 5 Gy of ionizing radiation and incubated for another 24
h. Cells were collected and total protein extracted by using M-PER
mammalian protein extraction reagent (Thermo Fisher Scientific,
Waltham, MA, USA) and protein concentrations measured using Pierce
660 nm protein assay reagent (Thermo Fisher Scientific). Equal
amounts of protein (10 μg/lane) were electrophoresed under reducing
conditions on 4–16% gradient polyacrylamide gels. After SDS-PAGE,
the separated proteins were transferred on to a polyvinylidene
difluoride membrane (Bio-Rad Laboratories, Hercules, CA, USA).
Membranes were blocked with TBS-T containing 5% non-fat skim milk
for 1 h. Subsequently, membranes were immunoprobed with primary
antibody and appropriate horseradish peroxidase-labelled secondary
antibody. Specific protein bands were visualized using enhanced
chemiluminescence detection reagents (Life Technologies, Carlsbad,
CA, USA). A similar protocol was used for total protein isolated
from subcutaneous tumors of nude mice described in section below
(see method for neuroblastoma subcutaneous tumor model).
In vitro angiogenesis assay
To determine in vitro angiogenic inhibition
by SPARC overexpression in neuroblastoma cells, we collected
conditioned media from controls or radiated NB1691 and SK-NB-E(2)
cells with or without SPARC overexpression as per standard
protocols. The collected conditioned media (100 μl) was used to
culture human endothelial cells over Matrigel. As a positive
control for VEGF-A inhibition, we used 50 μg/ml of bevacizumab a
recombinant humanized monoclonal antibody that targets VEGF-A in
control NB1691 or SK-N-BE(2) conditioned media to compare
angiogenic inhibition with SPARC overexpression. The cells were
monitored every hour for network formation and after 6 h
endothelial cell network was visualized at 490 nm excitation and
520 nm emission using an inverted fluorescent microscope.
Angiogenesis was quantified by determining the number of branch
points as per standard protocols.
Animals
BALB/c nude female mice aged 6–8 weeks were obtained
from Harlen Labs, Inc. (Indianapolis, IN, USA) and housed in
micro-isolation cages in groups of five animals in ventilated racks
at a constant temperature of 20–26°C and humidity of 30–70%. All
animal experiments were carried out after obtaining approval from
the Institutional Animal Care and Use Committee.
In vivo angiogenesis assay
The in vivo angiogenic assay was done as
previously described with minor modifications (20). SK-N-BE(2) or NB1691 cells
(1×106) treated with SPARC overexpressing plasmid alone
or with radiation were loaded into a diffusing chamber. A 2-cm long
incision was made horizontally along the edge of the dorsal air sac
of the mice and the chambers were placed underneath the skin. The
mice were sacrificed 10 days later; and carefully skinned around
the implanted chambers. The skin folds covering the chambers were
photographed under visible light, and tumor induced vasculature
quantified.
Neuroblastoma subcutaneous tumor
model
The subcutaneous tumor model was done as previously
described by us with minor modifications (21). SK-N-BE(2) and NB1691 cells were
implanted subcutaneously into nude mice on day 0 (1×105
cells). After 25 days, animals developed detectable tumors. Mice
were separated into 4 groups of 5 animals per group. The animals
that lost ≥20% of body weight or had trouble ambulating, feeding or
grooming were sacrificed. On day 28 and day 32, two groups of
animals received 150 μg/animal per day intratumoral injection of
SPARC plasmid DNA given at three different locations on the tumor
(plasmid concentration of 1.5 μg/μl in PBS, 50 μg per site 120°
apart) at a depth of 3–5 mm using a 30-gauge insulin syringe. The
overall cumulative dose was 300 μg/animal. Two groups were left as
controls. On day 30, one group each of SPARC treated and control
animals were exposed to 5 Gy ionizing radiation. Tumor growth was
followed, till day 45, after which animals were sacrificed. Tumors
were harvested, imaged followed by, protein and total RNA isolation
by standard protocols. Tumors were fixed in buffered formaldehyde
and paraffin sectioned for immunohistochemical analysis of SPARC
and VEGF-A. In vivo tumor size measurement: Subcutaneous
tumors volumes were measured using the following formula: π/6 (L ×
W2) where L is the length and W is the width.
Immunohistochemistry
Excised tumors were fixed in 10% buffered formalin
and embedded in paraffin. Tissue sections (5 μm thick) collected on
slides were deparaffinized and endogenous peroxidases activity was
blocked at room temperature by 5–10 min of incubation in 0.3%
H2O2 in PBS (pH 7.7). The slides were then
rinsed in PBS for 5 min, followed by antigen retrieval by heating
to 95°C in citrate buffer (0.0 mol/l sodium citrate buffer, pH 6.0)
for 10 min. The tissues were blocked using normal goat serum for 20
min at room temperature and incubated in primary antibody
(anti-VEGF or SPARC) for 60 min at room temperature. The slides
were then rinsed in PBS followed by the addition of a horseradish
peroxidase (HRP)-conjugated secondary antibody for 20 min at room
temperature. HRP substrate DAB (3,3′-diaminobenzidine
tetrahydrochloride) at 1 mg/ml in 50 mmol/l Tris, pH 7.2, and 0.3%
H2O2 was then added for the development of
DAB substrate followed by haematoxylin staining. Next, the slides
were dehydrated and mounted with coverslips. Negative control
slides were obtained by treating with non-specific IgG. Sections
were mounted and analyzed using an inverted microscope. The
intensity of VEGF-A or SPARC expression per unit area was measured
in arbitrary pixel units using ImageJ software.
Real-time PCR
Total RNA was extracted from the tissue samples
using mirVana™ miRNA isolation kit (Ambion-Life Technologies,
Carlsbad, CA, USA). cDNA was synthesized from RNA using miScript II
RT kit (Qiagen, Valencia, CA, USA) according to the manufacturer's
instructions. Real-time PCR was performed using miScript SYBR-Green
PCR kit in an automated thermal cycler (Bio-Rad Laboratories).
miR-410 and SNORD68 primers were obtained from Qiagen. SNORD68 was
used as an internal control for normalization. The expression
levels of pro-angiogenic factors VEGF-A, -B, -C, FGF2, PDGF, ANGPT1
and ANGPT2 (Table I) were measured
as per standard protocol and normalized to HPRT. Relative gene
expression was calculated by the 2−ΔCt method.
 | Table IPCR primers. |
Table I
PCR primers.
| Primer name | Sequence
(5′-3′) |
|---|
| VEGF-A-F |
AGGAGGAGGGCAGAATCATCA |
| VEGF-A-R |
CTCGATTGGATGGCAGTAGCT |
| VEGFB-F |
GAGATGTCCCTGGAAGAACACA |
| VEGFB-R |
GAGTGGGATGGGTGATGTCAG |
| VEGFC-F |
GGCTGGCAACATAACAGAGAA |
| VEGFC-R |
CCCCACATCTATACACACCTCC |
| FGF2-F |
AGAAGAGCGACCCTCACATCA |
| FGF2-R |
CGGTTAGCACACACTCCTTTG |
| ANGPT1-F |
AGAACCTTCAAGGCTTGGTTAC |
| ANGPT1-R |
GGTGGTAGCTCTGTTTAATTGCT |
| ANGPT2-F |
CTCGAATACGATGACTCGGTG |
| ANGPT2-R |
TCATTAGCCACTGAGTGTTGTTT |
| PDGF-F |
GCAAGACCAGGACGGTCATTT |
| PDGF-R |
GGCACTTGACACTGCTCGT |
| HPRT1-F |
TGACACTGGCAAAACAATGCA |
| HPRT1-R |
GGTCCTTTTCACCAGCAAGCT |
Inhibition of miR-410 expression
MicroRNA transfection reagent, miR-410 mimic
(AAUAUAACACAGAUGGCCUGU) and miR-410 inhibitors
(UUAUAUUGUGUCUACCGGACA) were obtained from Sigma-Aldrich (St.
Louis, MO, USA). SK-N-BE(2) and NB1691 were transfected as per the
manufacturer's instructions and conditioned media from these cells
were used in CAM assay or for ELISA determination of VEGF-A protein
levels. To determine the effectiveness of miR-410 inhibition,
miR-410 inhibitor treated cells were assayed for the expression
levels of miR-410 using RT-PCR as per standard protocols and
normalized to SNORD68 expression.
Chicken chorioallantoic membrane (CAM)
assay
Fertilized white leghorn chicken eggs were obtained
from Charles River Laboratories (Wilmington, MA, USA) and incubated
at 37°C with 60% humidity. On day 7, a small window was made on the
egg shell under aseptic conditions. Sterile filter paper disks
(Whatman, 1–2 mm size) were placed on top of the CAM. Conditioned
medium collected from SK-N-BE(2) and NB1691 cells were added on top
of the filter disks, gap sealed with porous adhesive tape and the
eggs were returned to the incubator. On day 12, CAM surrounding the
filter disk was carefully removed, placed on glass slides and
visualized for angiogenesis using a stereomicroscope. Angiogenesis
was quantified by measuring the length of newly formed capillaries
by using imageJ software.
VEGF-A ELISA
Protein levels of VEGF-A were determined from the
conditioned media of SK-N-BE(2) and NB1691 cells overexpressed for
SPARC with or without miR-410 inhibitor using the VEGF-A ELISA kit
from Thermo Fisher Scientific as per manufacturer's protocols.
Statistical analysis
All data are expressed as mean ± SD. Statistical
analysis was performed using Student's t-test. A P-value of ≤0.05
was considered statistically significant. All experiments were
performed in triplicate, or as indicated.
Results
Expression of VEGF-A in human
neuroblastoma tissues is inversely proportional to the expression
of SPARC
Human neuroblastoma tissue array from US Biomax,
Inc., consisting of 61 neuroblastoma cases, was processed for
immunohistochemistry for the expression of VEGF-A and SPARC as
previously described (20). From
the tissue array, we observed that the expression of SPARC was
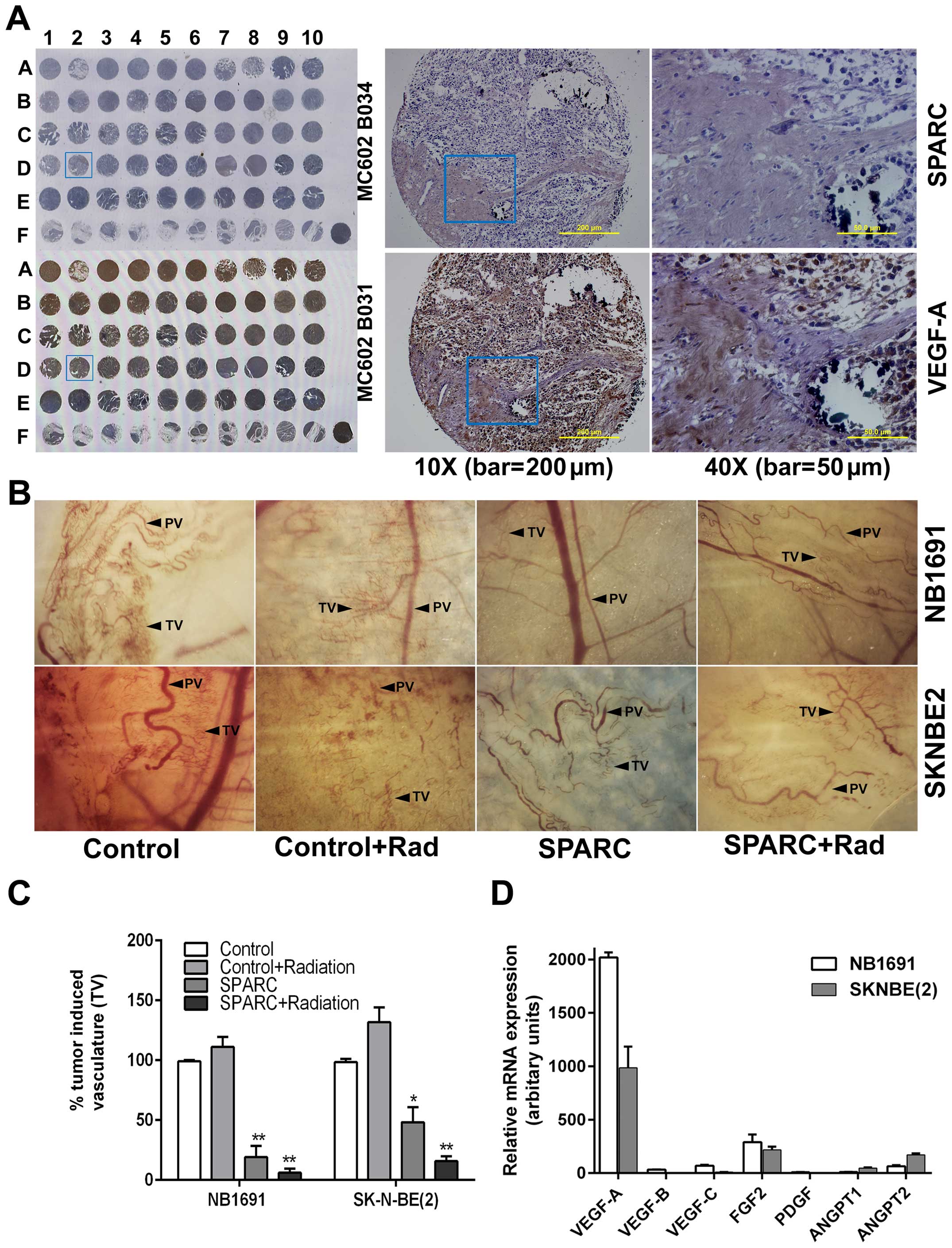
always inversely correlated with the expression of VEGF-A (Fig. 1A). Further semi-quantification of
expression of SPARC and VEGF-A in the human tissue array reveled
that higher expression of SPARC correlated with retroperitoneal
tumors rather than with tumors in the abdominal cavity,
mediastinum, or adrenal gland (data not shown).
Overexpression of SPARC combined with
radiation suppresses angiogenesis
The role of SPARC in angiogenesis has been
demonstrated by us and other researchers (22,23).
The addition of radiation treatment was included because radiation
induced vascular injury can promote the development of leaky
vasculature that can contribute to local hypoxic regions promoting
an angiogenic response (24–26).
Therefore, targeting angiogenesis accompanied with radiation would
greatly impact the success of the consolidation therapy phase
(6). As we have observed that
VEGF-A is a significant player in angiogenesis in neuroblastomas
and that expression of SPARC is always inversely correlated with
the expression of VEGF-A (Fig.
1A), overexpression of SPARC could be a strategy for
controlling radiation induced angiogenesis that is mediated by
VEGF-A. To determine whether overexpression of SPARC suppressed
angiogenesis, we performed the dorsal skin fold angiogenesis assay
using nude mice as previously described (20). We observed that control mice showed
increased tumor induced vasculature (TV) which was clearly
differentiated from pre-existing vasculature (PV). Neuroblastoma
tumor that overexpressed SPARC showed a significantly reduced
amount of tumor induced vasculature (20%) in both NB1691 cells
(P<0.001) and in SK-N-BE(2) (50%) cells (P<0.05). Addition of
radiation treatment along with SPARC further suppressed tumor cell
induced vasculature by 94% in NB1691 cells (P<0.001) and by 85%
in SK-N-BE(2) cells (P<0.001) (Fig.
1B and C). Notably, addition of radiation alone, did increase
tumor cell induced vasculature to 110% (P=0.07) in NB1691 and to
128% (P=0.01) in SK-N-BE(2) of controls in the dorsal sac chamber
nude mouse models.
Both SK-N-BE(2) and NB1691 neuroblastoma
cells overexpress VEGF-A
Based on the results from the in vivo
angiogenic assay, we attempted to validate the involvement of
VEGF-A as a significant angiogenic contributor in SK-N-BE(2) and
NB1691 neuroblastoma cell lines. We performed RT-PCR to determine
the expression levels of known pro-angiogenic molecules VEGF-A, -B,
-C, FGF2, PDGF, ANGPT1 and ANGPT2 (Table I) (27) and observed that expression of
VEGF-A was significantly higher than the expression levels of
VEGF-B, -C, FGF2, PDGF, ANGPT1 or ANGPT2 in both SK-N-BE(2) and
NB1691 cells (Fig. 1D). Since the
expression of VEGF-A is the prominent contributor of angiogenesis;
we further investigated whether the suppression of angiogenesis by
SPARC is mediated by modulation of VEGF-A expression.
SPARC overexpression suppresses tumor
growth and VEGF-A expression
As we determined that VEGF-A is a major contributor
to angiogenesis in both SK-N-BE(2) and NB1691 cells (Fig. 1D), and in continuation with this
finding and our previous findings (22), we attempted to determine whether
SPARC expression can suppress tumor growth and angiogenesis by
modulating the expression of VEGF-A. We performed
immunohistochemical analysis for VEGF-A and SPARC on neuroblastoma
subcutaneous tumors developed in mice from SK-N-BE(2) or NB1691
neuroblastoma cells (Fig. 2A and
B). As described, SK-N-BE(2) and NB1691 cells were implanted
subcutaneously into nude mice followed by intratumoral injection of
SPARC plasmid DNA (150 μg/animal) and/or 5 Gy ionizing radiation.
Tumors were harvested after 45 days, and we observed that mice
radiated with 5 Gy showed slightly larger tumors when compared to
non-radiated controls. SPARC overexpression plasmid treated mice
showed decreased tumor size in NB1691 tumors but not so much in
SK-N-BE(2) tumors. Furthermore, mice treated with both radiation
and SPARC overexpression showed the greatest decrease in tumor
growth with two of the five mice showing undetectable tumors in
both NB1691 and SK-N-BE(2) implanted mice (Fig. 2B and C). Furthermore, we observed
that the expression of SPARC and VEGF-A were inversely correlated
(Fig. 2D) and was similar to the
human tissue array expression of SPARC and VEGF-A (Fig. 1A), where we observed that the
expression of SPARC and VEGF-A was inversely correlated. Graphical
representation of VEGF-A expression obtained by determining the
pixel intensity of HRP-DAB reaction showed that SPARC
overexpression strongly inhibited VEGF-A expression in both
SK-N-BE(2) (P<0.05) and NB1691 (P<0.05) neuroblastoma
subcutaneous tumors (Fig. 3A).
Addition of radiation further reduced VEGF-A levels in SK-N-BE(2)
(P<0.001) and NB1691 (P<0.001) derived tumors (Fig. 3A). Furthermore, mice injected with
SPARC overexpressing plasmids did show increased expression of
SPARC (P<0.001) (Fig. 3B). To
validate the immunohistochemistry data, proteins were extracted
from these tumors and expression levels of SPARC and VEGF-A
determined by western blot analysis (Fig. 3C). This also correlated well with
the human tissue array data (Fig.
1A) and the immunohistochemistry data (Fig. 2D). To further validate these
findings, we performed western blots for VEGF-A and SPARC on NB1691
and SK-N-BE(2) cells infected with adenovirus expressing SPARC at
50 MOI with and without radiation. We observed that upregulation of
SPARC resulted in significant reduction of VEGF-A expression in
both NB1691 and SK-N-BE(2) cells (Fig.
3D). To determine whether the suppression of angiogenesis was
similar to bevacizumab, we treated human endothelial cells with
conditioned media from NB1691 or SK-N-BE(2) cells with or without
SPARC overexpression and with or without radiation. We used
bevacizumab, a known VEGF-A inhibitor as a positive control. We
observed that angiogenic inhibition by overexpression of SPARC was
comparable to cells treated with bevacizumab both with and without
radiation (Fig. 3E and F).
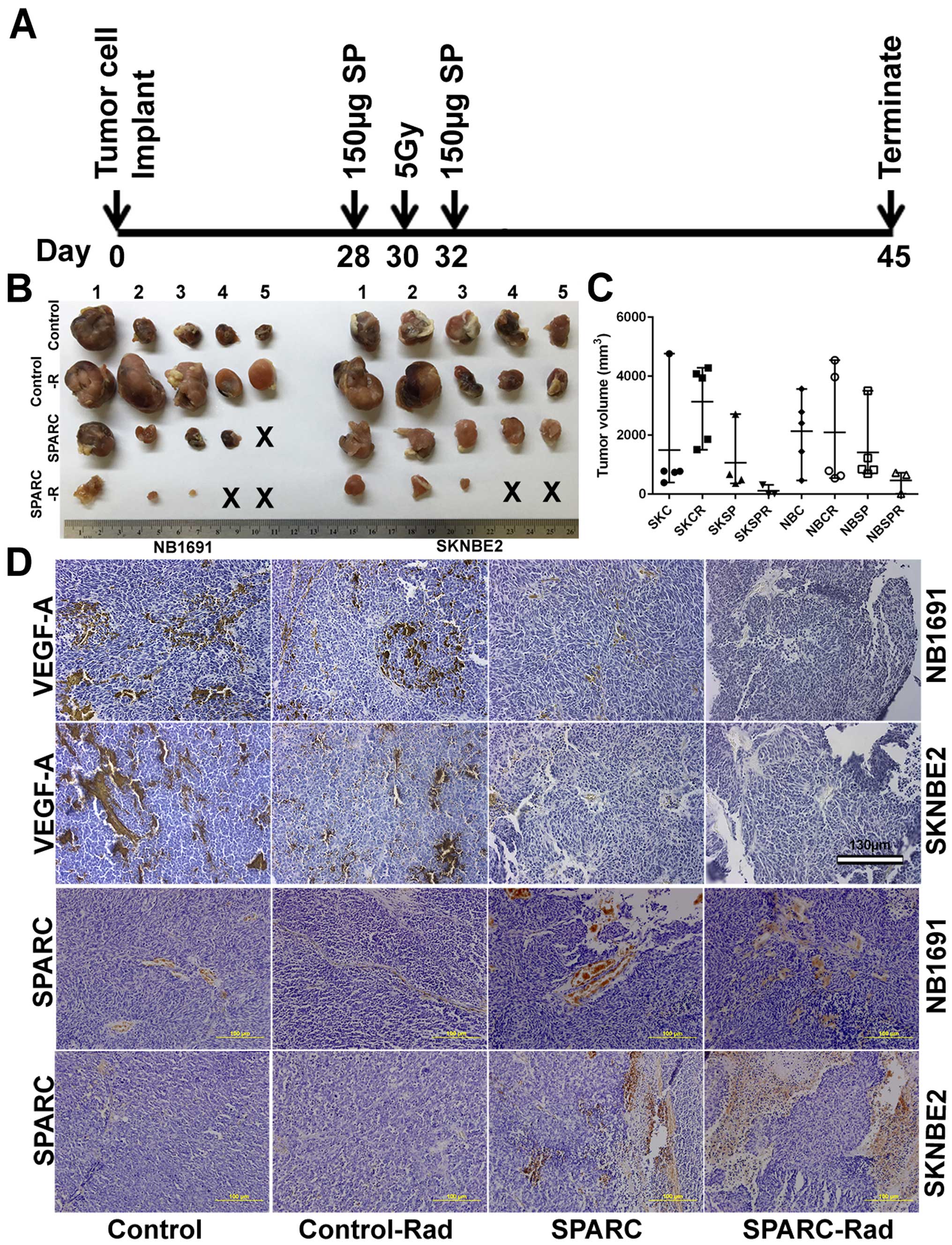 | Figure 2Overexpression of SPARC and low-dose
radiation in NB1691 and SK-N-BE(2) subcutaneous tumors suppresses
tumor growth. Time line showing neuroblastoma tumor implantation
and SPARC treatment with or without radiation in nude mice (A), 45
days after tumor development, tumors were harvested and volumes
were measured (B). (C) (SKC, SK-N-BE(2) control; SKCR, SK-N-BE(2) +
5 Gy radiation; SKSP, SK-N-BE(2) + SPARC overexpression; SKSPR,
SK-N-BE(2) + SPARC overexpression + 5 Gy radiation; NBC, NB1691
control; NBCR, NB1691 + 5 Gy radiation; NBSP, NB1691 + SPARC
overexpression; NBSPR, NB1691 + SPARC overexpression + 5 Gy
radiation). (D) After formalin fixation the tumors were processed
for paraffin sectioning and VEGF-A and SPARC expression levels were
visualized by immunohistochemistry. |
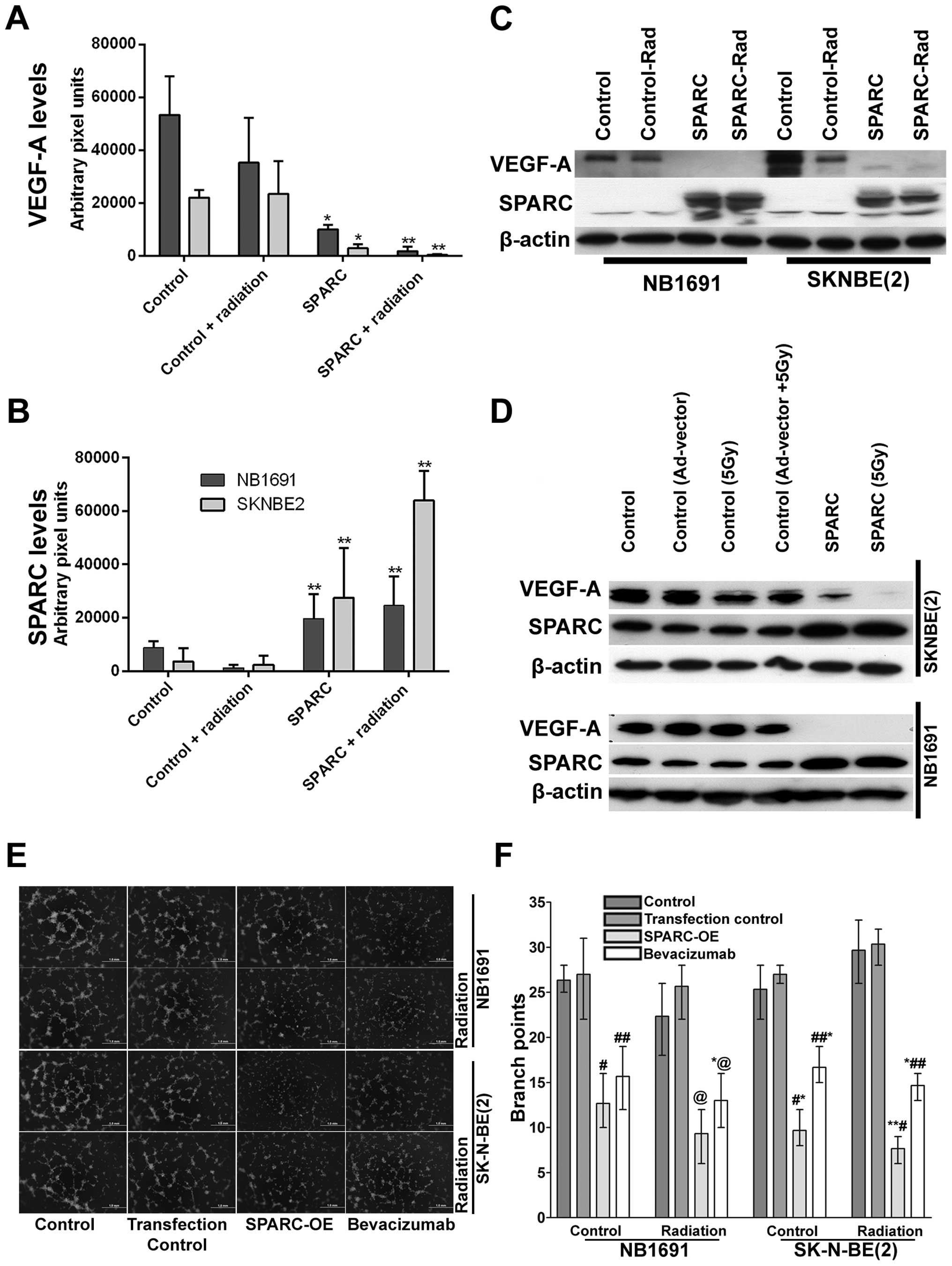 | Figure 3Overexpression of SPARC and low-dose
radiation suppresses NB1691 and SK-N-BE(2) VEGF-A expression. SPARC
and VEGF-A expression from paraffin sections of NB1691 and
SK-N-BE(2) tumors were quantified by ImageJ and represented as
arbitrary pixel units. SPARC overexpression resulted in significant
reduction of VEGF-A expression, radiation further decreased SPARC
induced reduction of VEGF-A (control vs. SPARC and control vs.
SPARC + radiation, *P<0.05, **P<0.001)
(A). Semi-quantitative analysis showing increase of SPARC
expression when treated with SPARC overexpressing plasmid (control
vs. SPARC and control vs. SPARC + radiation
**P<0.001) (B). Western blotting for VEGF-A and SPARC
in NB1691 and SK-N-BE(2): subcutaneous tumors treated with SPARC
overexpression plasmid with or without radiation showed decrease in
levels of VEGF-A (C). Similarly, increased SPARC expression along
with radiation greatly reduced the expression of VEGF-A in both
SK-N-BE(2) and NB1691 cell lines (D). In vitro angiogenic
inhibition by SPARC overexpression was compared to bevacizumab
using endothelial cells cultured over Matrigel in NB1691 and
SK-N-BE(2) conditioned media as indicated (E). Quantative analysis
for angiogenic inhibition revealed that SPARC overexpression (OE)
was comparable to bevacizumab treatments in both NB1691 (control
vs. SPARC-OE #P=0.05, control vs. bevacizumab
##P=0.06, control vs. SPARC-OE radiated
@P=0.02 and control vs. bevacizumab radiated
*,@P=0.03) and SK-N-BE(2) (control vs. SPARC-OE
*P=0.005, control vs. bevacizumab
##,*P=0.004, control vs. SPARC-OE radiated
**,#P=0.004 and control vs. bevacizumab radiated
*,##P=0.0002) cells (F). |
SPARC regulates VEGF-A mediated
angiogenesis through miR-410
VEGF-A is a major contributor of angiogenesis and is
a recognized therapeutic target (28–30).
Our data from miRanda analysis presented shows that miR-410 is a
significant modulator of VEGF-A expression (Table II). To validate this in a
biological setting, we investigated the relevance of miR-410 as an
angiogenic regulator using a chicken chorioallantoic membrane (CAM)
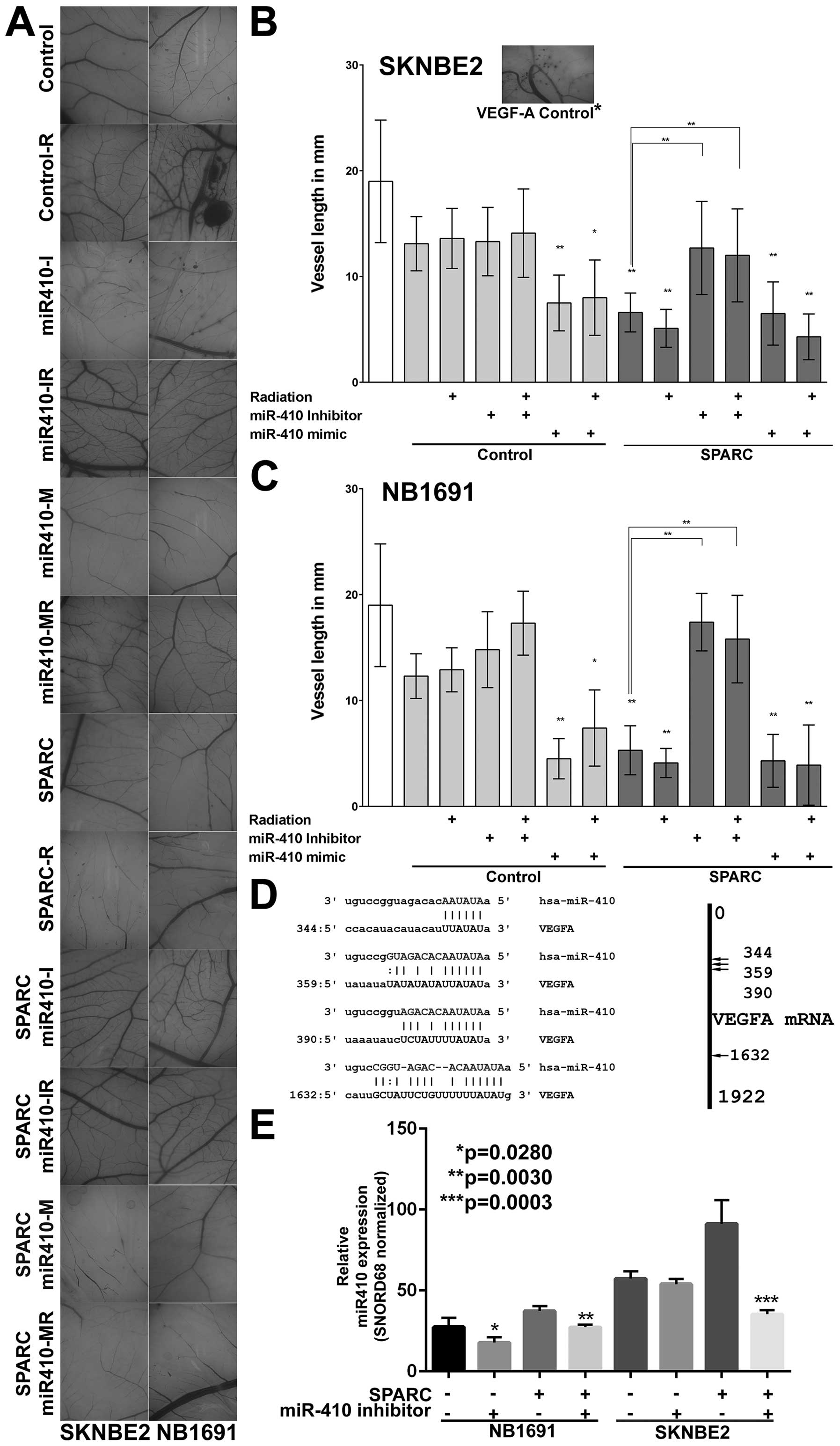
assay (Fig. 4A), which is an
accepted model for in vivo angiogenesis (31). We observed that NB1691 and
SK-N-BE(2) cells treated with SPARC reduced angiogenesis when
compared to controls in both NB1691 (P<0.001) and SK-N-BE(2)
cells (P<0.001) (Fig. 4B and
C). Interestingly, we also found that miR-410 mimic (AAUAUAACACAGAU
GGCCUGU) alone showed the same angiogenic inhibitory effect in both
NB1691 (P<0.001) and SK-N-BE(2) cells (P<0.001) (Fig. 4B and C). To study if this
inhibition of angiogenesis is specifically controlled by miR-410,
we inhibited miR-410 using a specific inhibitor (UUAUAUUGUGUCU
ACCGGACA) in SPARC overexpressing cells. Inhibition of miR-410
reversed SPARC induced reduction of angiogenesis in both NB1691
(P<0.001) and SK-N-BE(2) cells (P<0.001) (Fig. 4B and C). This reversal of
angiogenic vessel formation was also consistent in the presence of
radiation. These findings clearly show that SPARC downregulates
VEGF-A mediated angiogenesis specifically by upregulating the
expression of miR-410. The P-values represent change in vessel
length in 5 replicates. Similar to the in vivo dorsal skin
assay where we observed that radiation increased tumor cell induced
vasculature, in the CAM assay also we observed that radiation
treated cells induced an angiogenic response greater than
non-radiated cells. This radiation induced angiogenesis in the CAM
ex vivo model was, however, not as pronounced as seen in the
in vivo model.
| Table IIThe miRNAs targeting VEGF-A. |
Table II
The miRNAs targeting VEGF-A.
| MicroRNA | Position | No. of sites |
|---|
| hsa-miR-410 | 344, 359, 390,
1632 | 4 |
| hsa-miR-590-3p | 1375 | 1 |
| hsa-miR-374a | 355 | 1 |
| hsa-miR-429 | 1286 | 1 |
| hsa-miR-185 | 1396, 1686 | 2 |
| hsa-miR-383 | 1900 | 1 |
| hsa-miR-361-5p | 1604 | 1 |
| hsa-miR-186 | 410 | 1 |
| hsa-miR-300 | 1366, 1553,
1879 | 3 |
| hsa-miR-381 | 1366, 1553,
1883 | 3 |
| hsa-miR-424 | 262 | 1 |
| hsa-miR-29a | 1730 | 1 |
| hsa-miR-29b | 1729 | 1 |
| hsa-miR-29c | 1730 | 1 |
| hsa-miR-497 | 263 | 1 |
| hsa-miR-15b | 262 | 1 |
| hsa-miR-374b | 357 | 1 |
| hsa-miR-15a | 260 | 1 |
| hsa-miR-200b | 1284 | 1 |
| hsa-miR-200c | 1283 | 1 |
| hsa-miR-16 | 261 | 1 |
| hsa-miR-195 | 264 | 1 |
| hsa-miR-299-3p | 329 | 1 |
| hsa-miR-494 | 1382 | 1 |
| hsa-miR-140-5p | 1053 | 1 |
| hsa-miR-205 | 138 | 1 |
| hsa-miR-134 | 1412 | 1 |
| hsa-miR-340 | 342, 388, 1635 | 3 |
| hsa-miR-495 | 1708 | 1 |
| hsa-miR-1 | 1496 | 1 |
| hsa-miR-206 | 1496 | 1 |
| hsa-miR-613 | 1497 | 1 |
| hsa-miR-339-5p | 518, 1150 | 2 |
| hsa-miR-329 | 1562 | 1 |
| hsa-miR-362-3p | 1562 | 1 |
| hsa-miR-141 | 1197 | 1 |
| hsa-miR-203 | 1305 | 1 |
| hsa-miR-382 | 1550 | 1 |
| hsa-miR-20a | 164 | 1 |
| hsa-miR-20b | 164 | 1 |
| hsa-miR-93 | 162 | 1 |
| hsa-miR-106a | 164 | 1 |
| hsa-miR-106b | 166 | 1 |
| hsa-miR-17 | 164 | 1 |
| hsa-miR-519d | 164 | 1 |
| hsa-miR-144 | 1735 | 1 |
| hsa-miR-505 | 1096 | 1 |
| hsa-miR-503 | 261 | 1 |
| hsa-miR-543 | 435 | 1 |
| hsa-miR-103 | 260, 800 | 2 |
| hsa-miR-107 | 260, 800 | 2 |
| hsa-miR-23a | 1597 | 1 |
| hsa-miR-23b | 1597 | 1 |
| hsa-miR-486-5p | 717 | 1 |
| hsa-miR-342-3p | 1088 | 1 |
|
hsa-miR-125a-5p | 4 | 1 |
| hsa-miR-125b | 11 | 1 |
| hsa-miR-150 | 484 | 1 |
|
hsa-miR-199b-5p | 451 | 1 |
| hsa-miR-200a | 1197 | 1 |
|
hsa-miR-199a-5p | 454 | 1 |
| hsa-miR-101 | 1733 | 1 |
| hsa-miR-373 | 163 | 1 |
| hsa-miR-520e | 163 | 1 |
|
hsa-miR-125a-3p | 873 | 1 |
| hsa-miR-302a | 161 | 1 |
| hsa-miR-302b | 161 | 1 |
| hsa-miR-302c | 157 | 1 |
| hsa-miR-372 | 158 | 1 |
|
hsa-miR-520a-3p | 161 | 1 |
| hsa-miR-302e | 167 | 1 |
SPARC overexpression upregulates miR-410
(a microRNA that targets VEGF-A)
To test whether SPARC regulates VEGF-A expression
via microRNAs, we first performed a target prediction analysis
using the miRanda algorithm (32)
and determined that miR-410 can be a potential mediator for VEGF-A
suppression. Of several miRNAs that had potential target regions in
the VEGF-A mRNA, miR-410 targeted 4 specific regions of VEGF-A mRNA
(Fig. 4D). To study if SPARC was
associated with miR-410 expression, we measured miR-410 levels in
tumor samples collected from mice treated with SPARC overexpressing
plasmid. Our results show that SPARC overexpression significantly
increased miR-410 levels in NB1691 cells (P<0.05) and SK-N-BE(2)
cells (P<0.001) and addition of miR-410 inhibitor showed
suppression of SPARC induced miR-410 expression in both SK-N-BE(2)
and NB1691 cells (Fig. 4E).
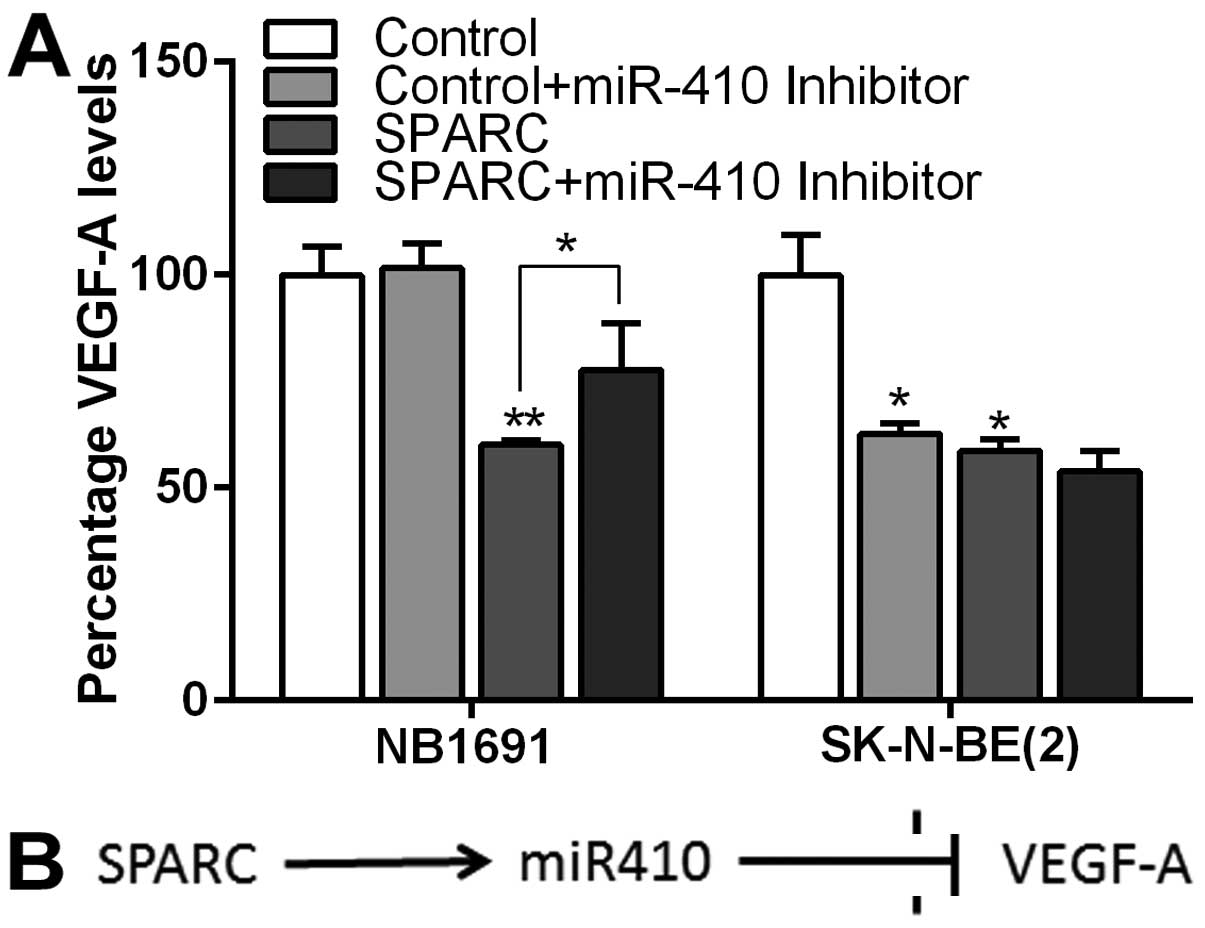
SPARC-induced downregulation of VEGF-A is
mediated by miR-410
To further validate our findings, we looked for
secreted levels of VEGF-A using ELISA in conditioned media used for
the CAM (Fig. 5A). We observed
that SPARC overexpression alone decreased VEGF-A protein levels by
40% in NB1691 cells (P<0.001) and 42% in SK-N-BE(2) cells
(P<0.05) (Fig. 5A). Addition of
miR-410 inhibitor to SPARC overexpressed cells significantly
rescued VEGF-A to 77% of controls (P=0.05) in NB1691, whereas in
SK-N-BE(2) cells no such reversal was observed; P-value represents
change in VEGF-A protein levels in culture media compared to
controls in 3 replicates.
Discussion
Angiogenesis is the process by which new blood
vessels develop from pre-existing vasculature. This process of
angiogenesis provides an efficient way for tumor cells to leave
their primary site and enter the blood stream resulting in
metastasis. Angiogenesis is a crucial factor for cancer progression
in neuroblastoma (33–35). Neuroblastoma (NB) is the most
common extracranial solid cancer which originates from developing
cells of sympathetic nervous system in children. The invasive and
hyper-vascular nature of the high-risk neuroblastoma makes it
highly metastatic and refractory to therapy (36). Current therapy for high-risk
neuroblastoma is multimodal and includes chemotherapy, surgery,
radiation therapy, hematopoietic stem cell transplantation and
immunotherapy. It is evident from various studies that radiation
intensifies angiogenesis by upregulating angiogenic agents
(37), making radiation therapy
somewhat counterproductive. Targeting this radiation induced
amplification of angiogenesis may be a better therapeutic addition
for the treatment of NB. We have previously demonstrated that SPARC
expression inversely correlated with angiogenesis (22), indicating that SPARC expression can
be used as a modulator of angiogenesis. SPARC is known to be both a
pro- and an anti-oncogenic marker. In neuroblastomas, however,
there is an inverse correlation between SPARC expression levels and
the stage of tumors (36). There
also appears to a link between angiogenesis and SPARC expression
levels as seen from our studies. Notably, the expression of SPARC
is found to be low in neuroblastoma tumors (19) making SPARC a possible
anti-angiogenic molecule for therapeutic intervention. The goal of
this study was to demonstrate that SPARC overexpression can be a
possible option for the suppression of angiogenesis in
neuroblastomas. Our results here have demonstrated that SPARC
overexpression suppresses angiogenesis. Therefore, to pursue this
goal, we first used an in vivo mouse model to confirm the
effect of SPARC overexpression on angiogenesis. We observed that
the overexpression of SPARC suppressed formation of new blood
vessels confirming its antiangiogenic effect. This result was more
pronounced when SPARC transfection was combined with ionizing
radiation. SPARC overexpression also suppressed the moderate
increase in vasculature seen after radiation. Our results are in
agreement with the study by Chleski and colleagues who showed that
SPARC protein secreted by schwann cells surrounding neuroblastoma
tumors inhibits angiogenesis (18). They also showed that full length
SPARC protein and a SPARC peptide were able to inhibit angiogenesis
and block growth in preclinical models of neuroblastoma tumors
(23). Though the present study
showed that radiation increased angiogenesis moderately, the
results were not statistically significant. We also observed that
radiated tumors showed increased tumor size, but this was not
consistent in all radiation treated mice. Previously other
researchers have demonstrated that low doses of radiation can
promote tumor growth, metastasis and also enhance angiogenesis
(38), therefore, suppression of
this low-dose radiation induced angiogenesis can be therapeutically
very useful. We see that SPARC suppresses angiogenesis, but until
now no clear mechanism has been proposed for the possible
antiangiogenic effect of SPARC.
Angiogenesis is mediated by several factors such as
the vascular endothelial growth factor (VEGF), the fibroblast
growth factor (FGF), epidermal growth factor (EGF), the
angiopoietins (Ang-1 and Ang-2) and the platelet derived growth
factor (PDGF) (34). Of these,
VEGF is the most common, best studied and as seen in the present
study the most overexpressed angiogenic factor in neuroblastoma
cells and tissues. This in vivo study showed that SPARC
overexpression with or without radiation suppressed expression of
VEGF-A. This result is also supported by in vitro
experiments in which western blot data on neuroblastoma cell lines
NB1691 and SK-N-BE(2) showed reduced expression of VEGF-A protein
levels when SPARC was overexpressed. Our previous study has shown
that SPARC overexpression downregulates VEGF-A in neuroblastoma
cells by inhibiting Stat3 phosphorylation partially mediated by
notch signaling (22). Though this
pathway somewhat defines mechanistically the downregulation of
VEGF-A, it fails to explain the precise molecular players directly
involved in inhibiting VEGF-A mediated angiogenesis. A recent study
reported that VEGF-A has a possible anti-necrotic effect (39), indicating that VEGF-A under certain
conditions can behave as a protective agent for the promotion of
cell growth. It was reported that SPARC overexpression induces
endothelial cell apoptosis and inhibited angiogenesis (22). Though the observations are valid,
the mechanism detailed is circuitous and derivative of the
observations. Similar observations were made in gastric cancer
where, overexpression of SPARC was shown to inhibit VEGF-mediated
angiogenesis (40), but no clear
mechanistic pathway was elucidated in that study. In the present
study, we see that miR-410 targets the expression of VEGF-A. We
observed that among the 71 miRNAs targeting VEGF-A, miR-410 had a
significantly higher number of target sites (4 target sites). A
preliminary microarray analysis showed that the miR-410 was
differentially expressed in SPARC overexpressed NB1691 cells (data
not shown). This was further validated using miR-410 mimics and
inhibitors. Similar to the results in our study, miR-410 has been
shown to be a tumor suppressor miRNA in controlling angiogenesis in
other studies (41), moreover, in
pancreatic cancer, overexpression of miR-410 suppressed cell
growth, migration and angiogenesis by blocking VEGF and ERK
signaling pathways (42). Another
study done in mice employed the use of eye drops containing miR-410
to effectively treat retinal neovascularization (RNV) by blocking
VEGF mediated angiogenesis (41).
A recent study showed miR-410 overexpression resulted in better
suppression of VEGF expression than other VEGF targeting miRNAs
(43). Furthermore, studies
confirm that miR-410 directly binds to VEGF 3′-UTR region (43). This finding strengthens our results
and validate that miR-410 is a vital suppressor of angiogenesis and
is modulated by SPARC. Notably, miR-410 is also predicted to target
TNFSF11/RANKL among others. TNFSF11/RANKL (tumor necrosis factor
ligand superfamily member 11/receptor activator of nuclear factor
kappa-B ligand) is known to affect the immune system and controls
bone regeneration and remodeling (44). It is known that bone marrow
infiltrated neuroblastoma is considered an adverse prognostic
factor; with >50% of patients showing bone marrow infiltration
metastasis at diagnosis (45).
Another molecule that is strongly predicted to be a target of
miR-410 is NIPBL. NIPBL (Nipped-B-like protein), also known as
delangin or SCC2 homolog is a marker for poor prognosis and
promotes chemotherapy resistance (46). As we have previously discussed,
metastatic neuroblastomas show little to no expression of SPARC and
SPARC overexpression is known to induce expression of miR-410 by a
yet unknown mechanism. Therefore, the overexpression of SPARC can
suppress the expression of both TNFSF11/RANKL and NIPBL via miR-410
and may be a mechanism by which SPARC overexpressed cells show
decreased proliferation (16).
Though the present study is focused on VEGF-A mediated
angiogenesis, the implication of miR-410 expression are beyond the
scope of this study.
Notably, we observed that SPARC overexpression
reduced secreted VEGF-A levels and this effect was reversed by
adding miR-410 inhibitor in NB1691 cells and not in SK-N-BE(2)
cells. Though SPARC reduced VEGF-A levels in SK-N-BE(2) cells,
blocking miR-410 using an anti-miR-410 failed to rescue VEGF-A
levels. This may be due to SK-N-BE(2) being a p53 mutant cell line
and NB1691 being p53 wild-type cell line, but the mechanism is
still unclear. Interestingly, a study has shown that wild-type p53
alters VEGF expression by binding to its promoter (47). The molecular mechanisms involved in
SPARC regulated angiogenesis in tumors with deregulated p53 needs
to be further elucidated. Furthermore, another recent study showed
that p53 possesses dsRNA exonuclease activity (48), since the use of miR-410 inhibitors
involves the formation of dsRNA for its functioning, it is very
likely that the cytoplasmic mt-p53 of SK-N-BE(2) actively degrades
this miR-410 inhibitory complex with greater efficiency than the
wt-p53 of NB1691 cells. This correlates with the observation that
miR-410 inhibitor treated SK-N-BE(2) showed reduced rescue of
angiogenesis when compared to NB1691 cells. Functions of both
micro-RNAs and siRNAs involve similar mechanisms, and mammalian
cells are known to show varying degree of resistance to this RNAi
mechanism (49), this may also
explain why NB1691 and SK-N-BE(2) show variation in VEGF-A rescue.
Furthermore, the addition of miR-410 inhibitor to SPARC
overexpressed cells did show reversal of miR-410 in both NB1691 and
SK-N-BE(2) cells, this inhibitory effect was more pronounced in
SK-N-BE(2) cells than in NB1691 cells. This observation may be due
to the fact that even though SPARC overexpression increases the
expression of miR-410, the addition of miR-410 inhibitor may render
miR-410 functionally unavailable due to compliment sequence
binding.
MicroRNAs still hold a lot of potential for
therapeutic applicability. We see that the control of angiogenesis
by miRNAs is well documented; miRNAs have been shown to control
both pro- and anti-angiogenic pathways. In ovarian cancer, miR-497
has been shown to block VEGF signaling and this results in
suppression of angiogenesis by inhibition of PI3K/AKT and ERK
signaling pathways (50).
Moreover, miR-503 was shown to inhibit angiogenesis by
simultaneously inhibiting VEGF-A and FGF in hepatocellular
carcinoma (HCC) (51). In
colorectal cancer, miR-126 was shown to be epigenetically silenced
leading to tumor invasion and angiogenesis (52). Upregulation of miR-126 resulted in
reduced VEGF expression, leading to suppression of angiogenesis.
Upregulation of miR-107 in glioma cells have also been shown to
inhibit VEGF expression (53). To
the best of our knowledge, this study is the first to show the
possible mechanistic role of SPARC in controlling angiogenesis via
miR-410 in high-risk neuroblastomas.
The present study shows that VEGF-mediated
angiogenesis can be attenuated either by increasing SPARC
expression or by directly upregulating miR-410. In the subset of
patients with high-risk neuroblastoma who are treated with
radiation that show increased vascularization, SPARC and/or miR-410
can be considered as a valuable add-on therapeutic option.
Acknowledgements
This present study was supported by the NCI grant
R01CA147792 to C.S.G. The authors wish to thank Angela Daniels for
help with animal experimentation.
Abbreviations:
|
SPARC
|
secreted protein acidic and rich in
cysteine
|
|
VEGF
|
vascular endothelial growth factor
|
|
CAM
|
chorioallantoic-membrane
|
References
|
1
|
Maris JM, Hogarty MD, Bagatell R and Cohn
SL: Neuroblastoma. Lancet. 369:2106–2120. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pinto NR, Applebaum MA, Volchenboum SL,
Matthay KK, London WB, Ambros PF, Nakagawara A, Berthold F,
Schleiermacher G, Park JR, et al: Advances in risk classification
and treatment strategies for neuroblastoma. J Clin Oncol.
33:3008–3017. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yu DM, Huynh T, Truong AM, Haber M and
Norris MD: ABC transporters and neuroblastoma. Adv Cancer Res.
125:139–170. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Scaldaferri F, Vetrano S, Sans M, Arena V,
Straface G, Stigliano E, Repici A, Sturm A, Malesci A, Panes J, et
al: VEGF-A links angiogenesis and inflammation in inflammatory
bowel disease pathogenesis. Gastroenterology. 136:585–95. e52009.
View Article : Google Scholar
|
|
5
|
Kowshik J, Giri H, Kishore TK, Kesavan R,
Vankudavath RN, Reddy GB, Dixit M and Nagini S: Ellagic acid
inhibits VEGF/VEGFR2, PI3K/Akt and MAPK signaling cascades in the
hamster cheek pouch carcinogenesis model. Anticancer Agents Med
Chem. 14:1249–1260. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maris JM: Recent advances in
neuroblastoma. N Engl J Med. 362:2202–2211. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kushner BH, LaQuaglia MP, Bonilla MA,
Lindsley K, Rosenfield N, Yeh S, Eddy J, Gerald WL, Heller G and
Cheung NK: Highly effective induction therapy for stage 4
neuroblastoma in children over 1 year of age. J Clin Oncol.
12:2607–2613. 1994.PubMed/NCBI
|
|
8
|
De Ioris MA, Crocoli A, Contoli B,
Garganese MC, Natali G, Tomà P, Jenkner A, Boldrini R, De Pasquale
MD, Milano GM, et al: Local control in metastatic neuroblastoma in
children over 1 year of age. BMC Cancer. 15:792015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu AL, Gilman AL, Ozkaynak MF, London WB,
Kreissman SG, Chen HX, Smith M, Anderson B, Villablanca JG, Matthay
KK, et al; Children's Oncology Group. Anti-GD2 antibody with
GM-CSF, interleukin-2, and isotretinoin for neuroblastoma. N Engl J
Med. 363:1324–1334. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jones C, Mackay A, Grigoriadis A, Cossu A,
Reis-Filho JS, Fulford L, Dexter T, Davies S, Bulmer K, Ford E, et
al: Expression profiling of purified normal human luminal and
myoepithelial breast cells: Identification of novel prognostic
markers for breast cancer. Cancer Res. 64:3037–3045. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lien HC, Hsiao YH, Lin YS, Yao YT, Juan
HF, Kuo WH, Hung MC, Chang KJ and Hsieh FJ: Molecular signatures of
metaplastic carcinoma of the breast by large-scale transcriptional
profiling: Identification of genes potentially related to
epithelial-mesenchymal transition. Oncogene. 26:7859–7871. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen J, Wang M, Xi B, Xue J, He D, Zhang J
and Zhao Y: SPARC is a key regulator of proliferation, apoptosis
and invasion in human ovarian cancer. PLoS One. 7:e424132012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sato T, Oshima T, Yamamoto N, Yamada T,
Hasegawa S, Yukawa N, Numata K, Kunisaki C, Tanaka K, Shiozawa M,
et al: Clinical significance of SPARC gene expression in patients
with gastric cancer. J Surg Oncol. 108:364–368. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shin M, Mizokami A, Kim J, Ofude M, Konaka
H, Kadono Y, Kitagawa Y, Miwa S, Kumaki M, Keller ET, et al:
Exogenous SPARC suppresses proliferation and migration of prostate
cancer by interacting with integrin β1. Prostate. 73:1159–1170.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
DiMartino JF, Lacayo NJ, Varadi M, Li L,
Saraiya C, Ravindranath Y, Yu R, Sikic BI, Raimondi SC and Dahl GV:
Low or absent SPARC expression in acute myeloid leukemia with MLL
rearrangements is associated with sensitivity to growth inhibition
by exogenous SPARC protein. Leukemia. 20:426–432. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bhoopathi P, Gorantla B, Sailaja GS, Gondi
CS, Gujrati M, Klopfenstein JD and Rao JS: SPARC overexpression
inhibits cell proliferation in neuroblastoma and is partly mediated
by tumor suppressor protein PTEN and AKT. PLoS One. 7:e360932012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chlenski A, Liu S, Guerrero LJ, Yang Q,
Tian Y, Salwen HR, Zage P and Cohn SL: SPARC expression is
associated with impaired tumor growth, inhibited angiogenesis and
changes in the extracellular matrix. Int J Cancer. 118:310–316.
2006. View Article : Google Scholar
|
|
18
|
Chlenski A, Liu S, Crawford SE, Volpert
OV, DeVries GH, Evangelista A, Yang Q, Salwen HR, Farrer R, Bray J,
et al: SPARC is a key Schwannian-derived inhibitor controlling
neuroblastoma tumor angiogenesis. Cancer Res. 62:7357–7363.
2002.PubMed/NCBI
|
|
19
|
Feng J and Tang L: SPARC in tumor
pathophysiology and as a potential therapeutic target. Curr Pharm
Des. 20:6182–6190. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gorantla B, Asuthkar S, Rao JS, Patel J
and Gondi CS: Suppression of the uPAR-uPA system retards
angiogenesis, invasion, and in vivo tumor development in pancreatic
cancer cells. Mol Cancer Res. 9:377–389. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gondi CS, Lakka SS, Yanamandra N, Siddique
K, Dinh DH, Olivero WC, Gujrati M and Rao JS: Expression of
antisense uPAR and antisense uPA from a bicistronic adenoviral
construct inhibits glioma cell invasion, tumor growth, and
angiogenesis. Oncogene. 22:5967–5975. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gorantla B, Bhoopathi P, Chetty C,
Gogineni VR, Sailaja GS, Gondi CS and Rao JS: Notch signaling
regulates tumor-induced angiogenesis in SPARC-overexpressed
neuroblastoma. Angiogenesis. 16:85–100. 2013. View Article : Google Scholar
|
|
23
|
Chlenski A, Guerrero LJ, Peddinti R, Spitz
JA, Leonhardt PT, Yang Q, Tian Y, Salwen HR and Cohn SL:
Anti-angiogenic SPARC peptides inhibit progression of neuroblastoma
tumors. Mol Cancer. 9:1382010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liao D and Johnson RS: Hypoxia: A key
regulator of angiogenesis in cancer. Cancer Metastasis Rev.
26:281–290. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mabjeesh NJ and Amir S: Hypoxia-inducible
factor (HIF) in human tumorigenesis. Histol Histopathol.
22:559–572. 2007.PubMed/NCBI
|
|
26
|
Liu Y, Kudo K, Abe Y, Aoki M, Hu DL,
Kijima H and Nakane A: Hypoxia expression in radiation-induced late
rectal injury. J Radiat Res (Tokyo). 49:261–268. 2008. View Article : Google Scholar
|
|
27
|
Simons M: Integrative signaling in
angiogenesis. Mol Cell Biochem. 264:99–102. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang W, Ran S, Sambade M, Huang X and
Thorpe PE: A monoclonal antibody that blocks VEGF binding to VEGFR2
(KDR/Flk-1) inhibits vascular expression of Flk-1 and tumor growth
in an orthotopic human breast cancer model. Angiogenesis. 5:35–44.
2002. View Article : Google Scholar
|
|
29
|
Hong DS, Garrido-Laguna I, Ekmekcioglu S,
Falchook GS, Naing A, Wheler JJ, Fu S, Moulder SL, Piha-Paul S,
Tsimberidou AM, et al: Dual inhibition of the vascular endothelial
growth factor pathway: A phase 1 trial evaluating bevacizumab and
AZD2171 (cediranib) in patients with advanced solid tumors. Cancer.
120:2164–2173. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Waldner MJ and Neurath MF: Targeting the
VEGF signaling pathway in cancer therapy. Expert Opin Ther Targets.
16:5–13. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Staton CA, Reed MW and Brown NJ: A
critical analysis of current in vitro and in vivo angiogenesis
assays. Int J Exp Pathol. 90:195–221. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
John B, Enright AJ, Aravin A, Tuschl T,
Sander C and Marks DS: Human MicroRNA targets. PLoS Biol.
2:e3632004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ribatti D: Anti-angiogenesis in
neuroblastoma. Crit Rev Oncol Hematol. 86:212–221. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Roy CS, Karmakar S, Banik NL and Ray SK:
Targeting angiogenesis for controlling neuroblastoma. J Oncol.
2012:Article ID 782020. 2012. View Article : Google Scholar
|
|
35
|
Chlenski A, Liu S and Cohn SL: The
regulation of angiogenesis in neuroblastoma. Cancer Lett.
197:47–52. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ribatti D, Marimpietri D, Pastorino F,
Brignole C, Nico B, Vacca A and Ponzoni M: Angiogenesis in
neuroblastoma. Ann NY Acad Sci. 1028:133–142. 2004. View Article : Google Scholar
|
|
37
|
Gorski DH1, Beckett MA, Jaskowiak NT,
Calvin DP, Mauceri HJ, Salloum RM, Seetharam S, Koons A, Hari DM,
Kufe DW, et al: Blockage of the vascular endothelial growth factor
stress response increases the antitumor effects of ionizing
radiation. Cancer Res. 59:3374–3378. 1999.PubMed/NCBI
|
|
38
|
Sofia Vala I1, Martins LR, Imaizumi N,
Nunes RJ, Rino J, Kuonen F, Carvalho LM, Rüegg C, Grillo IM, Barata
JT, et al: Low doses of ionizing radiation promote tumor growth and
metastasis by enhancing angiogenesis. PLoS One. 5:e112222010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
El Ghazi F, Desfeux A, Brasse-Lagnel C,
Roux C, Lesueur C, Mazur D, Remy-Jouet I, Richard V, Jégou S,
Laudenbach V, et al: NO-dependent protective effect of VEGF against
excitotoxicity on layer VI of the developing cerebral cortex.
Neurobiol Dis. 45:871–886. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang JL, Chen GW, Liu YC, Wang PY, Wang
X, Wan YL, Zhu J, Gao HQ, Yin J, Wang W, et al: Secreted protein
acidic and rich in cysteine (SPARC) suppresses angiogenesis by
down-regulating the expression of VEGF and MMP-7 in gastric cancer.
PLoS One. 7:e446182012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen N, Wang J, Hu Y, Cui B, Li W, Xu G,
Liu L and Liu S: MicroRNA-410 reduces the expression of vascular
endothelial growth factor and inhibits oxygen-induced retinal
neovascularization. PLoS One. 9:e956652014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Guo R, Gu J, Zhang Z, Wang Y and Gu C:
MicroRNA-410 functions as a tumor suppressor by targeting
angiotensin II type 1 receptor in pancreatic cancer. IUBMB Life.
67:42–53. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhao D, Jia P, Wang W and Zhang G:
VEGF-mediated suppression of cell proliferation and invasion by
miR-410 in osteosarcoma. Mol Cell Biochem. 400:87–95. 2015.
View Article : Google Scholar
|
|
44
|
Hanada R, Hanada T, Sigl V, Schramek D and
Penninger JM: RANKL/RANK-beyond bones. J Mol Med (Berl).
89:647–656. 2011. View Article : Google Scholar
|
|
45
|
Morandi F, Corrias MV and Pistoia V:
Evaluation of bone marrow as a metastatic site of human
neuroblastoma. Ann NY Acad Sci. 1335:23–31. 2015. View Article : Google Scholar
|
|
46
|
Xu W, Ying Y, Shan L, Feng J, Zhang S, Gao
Y, Xu X, Yao Y, Zhu C and Mao W: Enhanced expression of cohesin
loading factor NIPBL confers poor prognosis and chemotherapy
resistance in non-small cell lung cancer. J Transl Med. 13:1532015.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Farhang Ghahremani M, Goossens S, Nittner
D, Bisteau X, Bartunkova S, Zwolinska A, Hulpiau P, Haigh K,
Haenebalcke L, Drogat B, et al: p53 promotes VEGF expression and
angiogenesis in the absence of an intact p21-Rb pathway. Cell Death
Differ. 20:888–897. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Grinberg S, Teiblum G, Rahav G and
Bakhanashvili M: p53 in cytoplasm exerts 3′→5′ exonuclease activity
with dsRNA. Cell Cycle. 9:2442–2455. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zheng ZM, Tang S and Tao M: Development of
resistance to RNAi in mammalian cells. Ann NY Acad Sci.
1058:105–118. 2005. View Article : Google Scholar
|
|
50
|
Wang W, Ren F, Wu Q, Jiang D, Li H and Shi
H: MicroRNA-497 suppresses angiogenesis by targeting vascular
endothelial growth factor A through the PI3K/AKT and MAPK/ERK
pathways in ovarian cancer. Oncol Rep. 32:2127–2133.
2014.PubMed/NCBI
|
|
51
|
Zhou B, Ma R, Si W, Li S, Xu Y, Tu X and
Wang Q: MicroRNA-503 targets FGF2 and VEGFA and inhibits tumor
angiogenesis and growth. Cancer Lett. 333:159–169. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang Y, Wang X, Xu B, Wang B, Wang Z,
Liang Y, Zhou J, Hu J and Jiang B: Epigenetic silencing of miR-126
contributes to tumor invasion and angiogenesis in colorectal
cancer. Oncol Rep. 30:1976–1984. 2013.PubMed/NCBI
|
|
53
|
Chen L, Li ZY, Xu SY, Zhang XJ, Zhang Y,
Luo K and Li WP: Upregulation of miR-107 inhibits glioma
angiogenesis and VEGF expression. Cell Mol Neurobiol. 36:113–120.
2015. View Article : Google Scholar : PubMed/NCBI
|