Introduction
Gastric cancer is one of the most common cancers
worldwide, with more than 900,000 new cases and 700,000 deaths each
year (1). High-incidence areas
include Eastern Asia, Central and Eastern Europe and South America
(1). Current treatments to improve
survival involve surgical resection, supplemented with neoadjuvant
or adjuvant chemotherapy, with or without radiotherapy (2-4).
Although the incidence of gastric cancer is decreasing, the
prognosis is still among the poorest of all solid-organ tumors. The
5-year survival for stage IV disease rarely exceeds 5% and the
median overall survival is less than 1 year (5,6).
Fibroblast growth factors (FGFs) comprise a large
family of heparin-binding growth factors that regulate numerous
cellular and physiological processes including proliferation,
angiogenesis, invasion and migration (7). FGFs carry out their diverse functions
by binding and activating members of the FGF receptor (FGFR) family
of tyrosine kinase receptors (8).
FGF7, also known as keratinocyte growth factor is expressed
specifically in mesenchyme (7).
FGF7 exerts its effect in a paracrine manner and specifically
activates the receptor tyrosine kinase FGFR2IIIb (8,9).
FGF7 binding forces FGFR2 dimerization, and in turn rapidly
triggers several signal transduction pathways, including
RAS-mitogen-activated protein kinase (MAPK) and phosphoinositide
3-kinase (PI3K) Akt signaling pathways, which play fundamental
roles in tumor progression (10).
The coexpression of FGF7 and FGFR2 in gastric and pancreatic
cancers, as well as in lung adenocarcinomas, is associated with
poor prognosis (11–13). Paradoxically, decreased expression
of FGFR2 has been reported in several cancers (14–16).
These contrasting and context-dependent roles of FGFR2 underscore
the complexity of FGF7/FGFR2 signaling.
Thrombospondins are a family of homologous proteins
that regulate cellular phenotype and extracellular structure during
tissue genesis and remodeling (17). Thrombospondin 1 (THBS1) was the
first member to be identified, and it is widely known as an
endogenous inhibitor of angiogenesis (18). THBS1 is synthesized by many cell
types and can be secreted to the extracellular matrix. Numerous
cell assays attribute a role for THBS1 in cell invasion and
migration (19–21). However, conflicting results were
obtained in different cell types (22). In recent years, an increasing
number of studies have suggested THBS1 as a poor prognosis and
recurrence marker in various cancer types including glioma,
melanoma, as well as ovarian and pancreatic carcinomas (19,23–25).
Of note, THBS1 is a multi-modular and multi-functional protein that
exerts intricate and sometimes opposite effects on tumor
progression.
Based on the previous reports, we hypothesized that
FGF7/FGFR2 signal promotes invasion and migration in human gastric
cancer through regulation of THBS1. In the present study, we
investigated the mechanisms of THBS1 regulation by FGF7/FGFR2, and
we assessed the functional roles of FGF7/FGFR2/THBS1 in gastric
cancer invasion and migration in vitro and in
vivo.
Materials and methods
Tissue specimens
Fifty-three gastric cancer primary tissue samples
and adjacent matched non-tumor tissue samples were collected
between January 2014 and August 2014 after obtaining informed
consent from the patients under institutional review board-approved
protocols. None of the patients had received treatment prior to
enrolment in the present study. All diagnoses were
histopathologically confirmed. This study was approved by the
institutional research ethics committee of Tongji Hospital of
Tongji Medical College, Huazhong University of Science and
Technology (Wuhan, China).
Immunohistochemistry (IHC)
Formalin-fixed and paraffin-embedded tissue sections
(5 μm thick) were deparaffinized, rehydrated, blocked of
endogenous peroxidase and subjected to antigen retrieval. The
sections were incubated with 5% bovine serum albumin (BSA) at room
temperature for 30 min immediately followed by primary antibodies
(FGFR2, 11835, 1:50; Cell Signaling Technology, Inc., Danvers, MA,
USA; THBS1, TA325040, 1:100; Origene, Rockville, MD, USA) at 4°C
overnight. After washing in Tris-buffered saline (TBS), the slides
were reacted with peroxidase polymer-conjugated secondary antibody
and DAB following counterstaining with hematoxylin. Primary
antibodies were replaced by TBS as negative control. Each stained
section was evaluated by two senior pathologists blinded to the
clinical information, and conflicting cases were adjudicated by a
third pathologist. Five fields were randomly selected and observed
under a light microscope. The staining results were scored based on
the percentage of positive cells (0, <10% positive cells; 1,
10–30% positive cells; 2, 30–50% positive cells; and 3, >50%
positive cells), as well as on staining intensity (0 for negative
staining, 1 for weak staining, 2 for moderate staining and 3 for
strong staining). IHC total score was calculated as the product of
both scores, which ranged from 0 to 9. The cut-off was arbitrarily
defined as a score ≥4 represents high expression and a score <4
indicates low expression.
Cell lines and reagents
Human gastric cancer cell lines SGC7901, MKN28 and
NCI-N87, human colorectal cancer cell lines SW480, Caco2 and
HCT116, human lung adenocarcinoma cell line A549, human
nasopharyngeal carcinoma cell line CNE2, and human breast cancer
cell lines SKBR3 and MDA-MB-231 were acquired from Oncology
Laboratory of Tongji Hospital (Wuhan, China). Cells were maintained
in RPMI-1640 (Hyclone Laboratories, Inc., Logan, UT, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco, Waltham, MA,
USA) at 37°C under a humidified 5% CO2 atmosphere. FGF7
was purchased from PeproTech (Rocky Hill, NJ, USA). LY294002,
U0126, SP600125, SB203580 and RAD001 were obtained from Promoter
Biotechnology (Wuhan, China).
Invasion and migration assays
Cell migration and invasion assays were performed
using polycarbonate membrane Trans-well inserts (8 μm pore
size) in a 24-well format (Corning, Inc., Corning, NY, USA). For
the invasion assay, the insert membranes were precoated with
diluted Matrigel (BD Biosciences, San Jose, CA, USA). Cells
(1×105/well) in 0.2 ml of serum-free medium were seeded
onto the membranes of the upper chambers, which had been inserted
into the wells containing 10% FBS-supplemented medium. After 24 h,
the cells were fixed with 100% methanol and stained with 0.1%
crystal violet. Non-invaded cells were removed from the upper well
with cotton swabs. The Transwell migration assay was performed in a
similar manner but without Matrigel on the filter. Cells were
counted in three randomly chosen fields and photographed under an
inverted microscope (magnification, ×200).
shRNA-mediated FGFR2 knockdown
Lentivirus-FGFR2-RNAi vector and the corresponding
empty vector were obtained from Shanghai GeneChem, Co., Ltd.
(Shanghai, China). Two independent shRNA constructs (target
sequence: shRNA-1, 5′-CCCTGTTTGATAGAGTATA-3′; shRNA-2,
5′-GAGGCTACAAGGTACGAAA-3′) were designed and subcloned into the
lentivirus-FGFR2-RNAi vector to generate hU6-MCS-CMV-Puromycin,
which was confirmed by Sanger sequencing. The recombinant
lentiviruses were packaged and prepared at a final titer of more
than 3×108 transducing units/ml. SGC7901 cells were
cultured to 60% confluency in complete medium for 24 h before
transduction. Lentivirus-FGFR2-RNAi vectors or empty vectors were
added at a multiplicity of infection of 100 and supplemented with 5
μg/ml polybrene (Shanghai GeneChem). The supernatant was
changed after 12 h. After culturing for another 72 h, the stably
transfected cells were selected on 2 μg/ml puromycin
(Shanghai GeneChem) for 1 week. The expression level was examined
by quantitative reverse-transcription (qRT)-PCR and western
blotting at 1 week after selection.
Western blotting
Cultured cells were lysed in
radioimmunoprecipitation assay lysis buffer with protease inhibitor
cocktail (Roche Diagnostics, Indianapolis, IN, USA). Equal amounts
of protein (30 μg) solubilized in sample buffer were
separated by 10% sodium dodecyl sulfate polyacrylamide gel
electrophoresis and transferred onto polyvinylidene fluoride
membranes (Millipore, Billerica, MA, USA). The membranes were
blocked in TBS containing 0.1% Tween-20 (TBS-T) plus 5% non-fat
dried milk for 1 h at room temperature and probed with primary
antibodies at 4°C overnight. Primary antibodies were used at the
specified dilutions: anti-FGFR2 (11835, 1:1,000; Cell Signaling
Technology), anti-phospho-FGFR (3471, 1:1,000; Cell Signaling
Technology), anti-THBS1 (TA325040, 1:1,000; Origene) and
anti-β-actin (66009-1-lg, 1:5000; Proteintech, Chicago, IL, USA).
The membranes were washed 3 times for 10–15 min each in TBS-T and
incubated with horseradish peroxidase-conjugated goat anti-rabbit
or anti-mouse secondary antibodies (SA00001-2, SA00001-1, 1:5,000;
Proteintech) for 1 h at room temperature. Proteins were visualized
using an ECL detection kit (Thermo Fisher Scientific, Waltham, MA,
USA).
qRT-PCR
Total RNA was extracted using TRIzol reagent
(TransGen Biotech, Beijing, China), and reverse transcription was
carried out using the RevertAid First Strand cDNA Synthesis kit
(Thermo Fisher Scientific) according to the manufacturer's
instructions. For qPCR analysis, aliquots of cDNA were amplified
using the Fast SYBR-Green Master Mix (Thermo Fisher Scientific) on
an ABI Prism 7900 Sequence Detector. The threshold cycle (Ct) was
measured during the exponential amplification phase, and the
amplification plots were analyzed using SDS 2.1 software (Applied
Biosystems, Waltham, MA, USA). The PCR program consisted of an
initial denaturation cycle (10 min at 95°C) followed by 40 cycles
of denaturation (15 sec at 95°C) and annealing and elongation (60
sec at 60°C). A melting curve analysis was added at the end of the
program. The following primers were used: FGFR2, forward,
5′-GGAAAGTGTGGTCCCATCTGA-3′ and reverse
5′-TCCAGGTGGTACGTGTGATTG-3′; THBS1, forward,
5′-GCTCCAGCTCTACCAGTGTC-3′ and reverse, 5′-TCAGTCACTTGCGGATGCT-5′;
GAPDH, forward, 5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse
5′-GGCTGTTGTCATACTTCTCATGG-3′. GAPDH was used as an endogenous
control. The relative gene expression levels were calculated using
the comparative Ct (ΔΔCt) method, where the relative expression is
calculated as 2−ΔΔCt.
Enzyme-linked immunosorbent assay
(ELISA)
Serum-starved cells were treated with or without
FGF7. At 12, 24 and 48 h, conditioned media were collected and
ELISA for human THBS1 was conducted according to the manufacturer's
protocol (Origene). THBS1 concentration was determined from a
standard curve.
siRNA-mediated RNAi
siRNAs (THBS1, siRNA-1 5′-CGU UGGUGAUGUAACAGAATT-3′,
siRNA-2 5′-GAGUGGACCUCCUGUUCUATT-3′, siRNA-3
5′-GCGUGAAGUGUACUAGCUATT-3′) were synthesized at Shanghai GeneChem.
A non-targeting siRNA (Shanghai GeneChem) was used as a negative
control. Cells were transiently transfected with siRNA using
Lipofectamine 2000 (Invitrogen, Waltham, MA, USA) according to the
manufacturer's protocol. Briefly, siRNAs were diluted in serum-free
media and mixed with Lipofectamine 2000. The mixture was left at
room temperature for 10 min. Cells were washed twice with
phosphate-buffered saline (PBS) and then incubated with
Lipofectamine-siRNA complexes in a humidified incubator at 37°C
with 5% CO2 for 48 h. Transfection efficiency was
detected by qRT-PCR and western blotting.
Statistical analysis
Experiments were repeated at least three times. All
data are expressed as the mean ± standard deviation (SD). Means
were compared using Student's t-test. The Chi-square test or
Fisher's exact test was used to analyze the relation between FGFR2
and THBS1 expression and other clinicopathological parameters.
Additionally, correlation analysis was used to determine the
correlation between FGFR2 and THBS1 expression. There was a
positive correlation at the correlation coefficience of r>0.
P<0.05 was considered statistically significant.
Results
Expression of FGFR2 and THBS1 is
increased in tumor tissues, positively correlated and correlated
with clinicopathological factors
We determined the protein expression of FGFR2 and
THBS1 in paired tumor and adjacent normal tissues of 53 patients
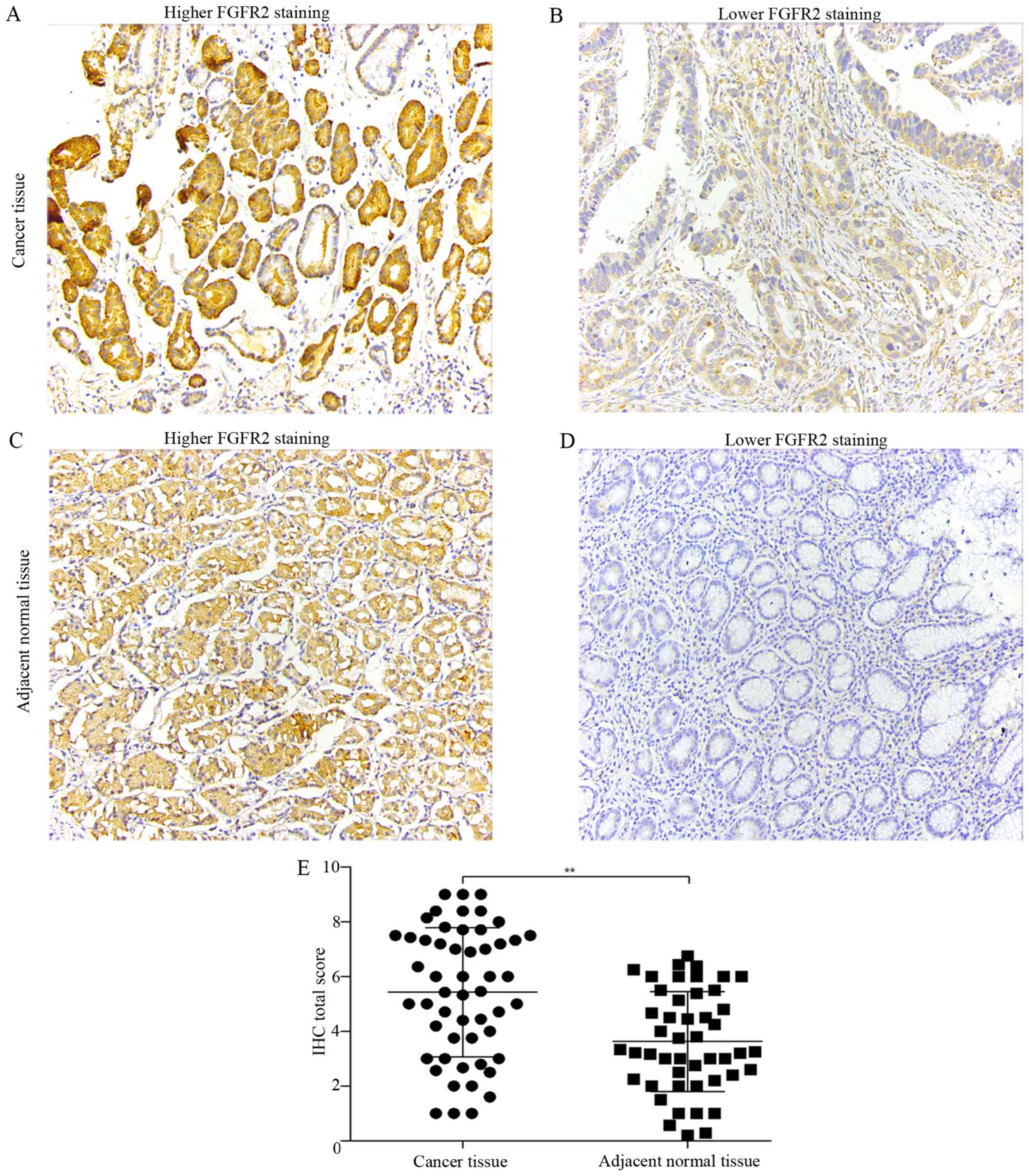
undergoing curative resection by IHC. FGFR2 was mainly expressed in
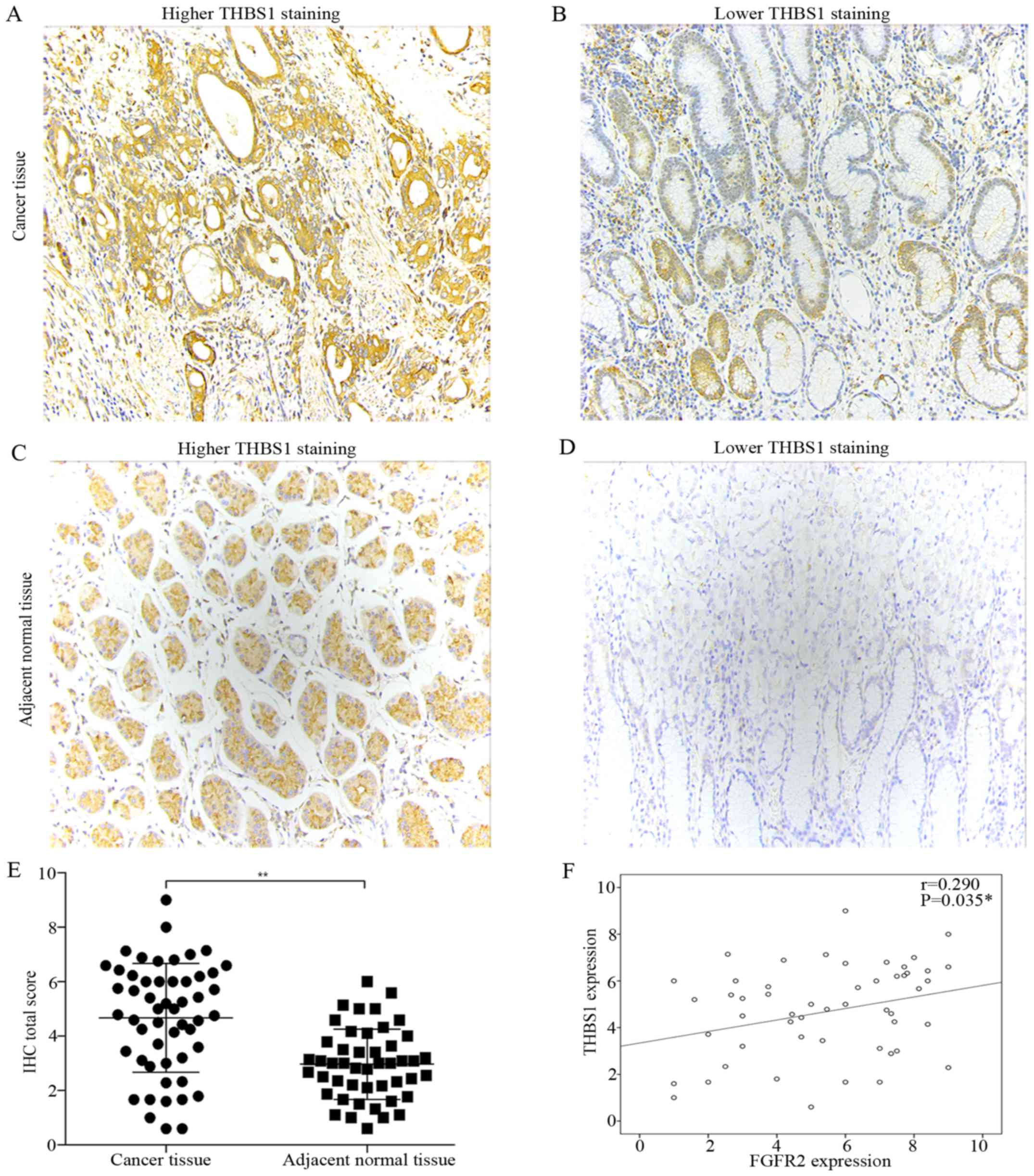
the cytoplasm (Fig. 1A and C),
while THBS1 was expressed in the cytoplasm and extracellular matrix
(Fig. 2A and C). Compared with
adjacent normal tissues, tumor tissues had significantly elevated
expression of FGFR2 and THBS1 (Figs.
1E and 2E).
Next, we classified the cancer tissues into high and
low-expression groups according to the criteria described in
Materials and methods. With the aim to identify the parameters
affecting processes involving FGFR2 and THBS1 in gastric cancer, we
analyzed the correlations between FGFR2 and THBS1 expression and
those with clinicopathological factors using the Chi-square test or
Fisher's exact test. Correlation analysis suggested a positive
correlation between FGFR2 and THBS1 expression (r=0.29, P=0.035)
(Fig. 2F). The median age at the
time of diagnosis was 53 years (range, 27–81 years). Patients with
high FGFR2 expression had deeper tumor depth (P=0.021) and were in
a later clinical stage (P=0.036) than patients with low FGFR2
expression (Table I). However,
high expression of THBS1 was correlated with well and moderate
tumor differentiation (P=0.003) (Table II). These findings indicated that
high FGFR2 expression may promote the invasion and migration in
gastric cancer, and THBS1 is a multifunctional protein that exerts
important roles on tumor progression.
 | Table ICorrelation of FGFR2 expression with
clinicopathological factors. |
Table I
Correlation of FGFR2 expression with
clinicopathological factors.
| Parameters | No. | Low expression | High
expression | P-value |
|---|
| Age (years) |
| <53 | 27 | 8 | 19 | 0.827 |
| ≥53 | 26 | 7 | 19 | |
| Gender |
| Male | 37 | 10 | 27 | 0.754 |
| Female | 16 | 5 | 11 | |
| Tumor size
(cm) |
| ≤4 | 29 | 10 | 19 | 0.272 |
| >4 | 24 | 5 | 19 | |
|
Differentiation |
| Poor | 31 | 8 | 23 | 0.632 |
| Well+moderate | 22 | 7 | 15 | |
| Tumor depth |
| T1+T2+T3 | 16 | 8 | 8 | 0.022a |
| T4 | 37 | 7 | 30 | |
| Lymph node |
| N0+N1 | 28 | 9 | 19 | 0.511 |
| N2+N3 | 25 | 6 | 19 | |
| Distant
metastasis |
| M0 | 53 | 15 | 38 | – |
| M1 | 0 | 0 | 0 | |
| Clinical stage |
| I+II | 20 | 9 | 11 | 0.036a |
| III+IV | 33 | 6 | 27 | |
| Table IICorrelation of THBS1 expression with
clinicopathological factors. |
Table II
Correlation of THBS1 expression with
clinicopathological factors.
| Parameters | No. | Low expression | High
expression | P-value |
|---|
| Age (years) |
| <53 | 27 | 10 | 17 | 0.430 |
| ≥53 | 26 | 7 | 19 | |
| Gender |
| Male | 37 | 9 | 28 | 0.066 |
| Female | 16 | 8 | 8 | |
| Tumor size
(cm) |
| ≤4 | 29 | 7 | 22 | 0.174 |
| >4 | 24 | 10 | 14 | |
|
Differentiation |
| Poor | 31 | 15 | 16 | 0.003a |
| Well+Moderate | 22 | 2 | 20 | |
| Tumor depth |
| T1+T2+T3 | 16 | 3 | 13 | 0.172 |
| T4 | 37 | 14 | 23 | |
| Lymph node |
| N0+N1 | 28 | 7 | 21 | 0.243 |
| N2+N3 | 25 | 10 | 15 | |
| Distant
metastasis |
| M0 | 53 | 17 | 36 | – |
| M1 | 0 | 0 | 0 | |
| Clinical stage |
| I+II | 20 | 5 | 15 | 0.39 |
| III+IV | 33 | 12 | 21 | |
FGF7/FGFR2 promotes invasion and
migration in human gastric cancer cells
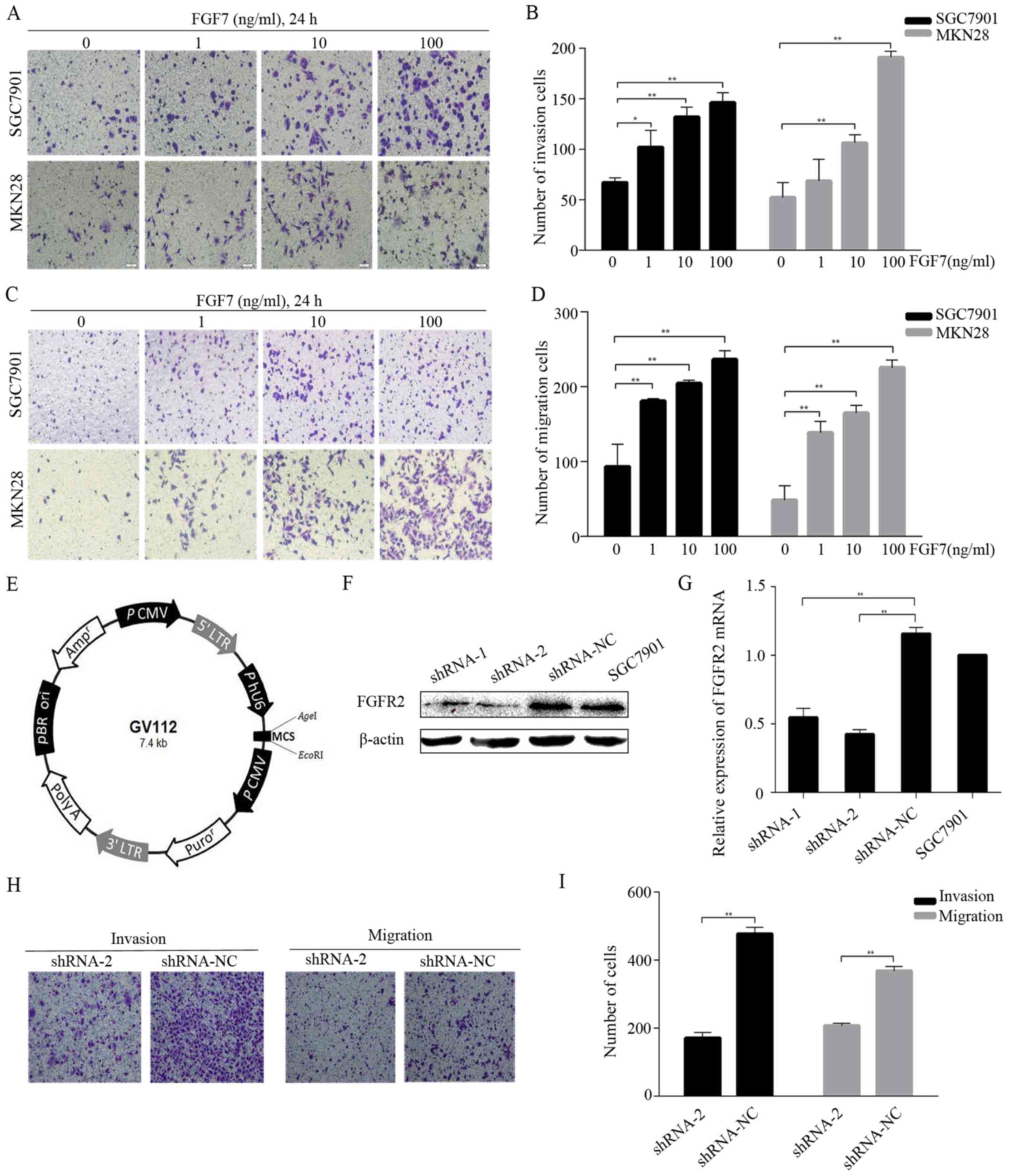
To mimic paracrine regulation and to check whether
exogenous FGF7 would promote invasion and migration in
vitro, we conducted invasion and migration assays using SGC7901
and MKN28 cells and different concentrations (1, 10 and 100 ng/ml)
of FGF7. The treatment with FGF7 caused a dose-dependent increase
in the number of invading (Fig. 3A and
B) and migrating (Fig. 3C and
D) cells. To study FGF7 function further, we knocked down FGFR2
expression in SGC7901 cells via lentivirus-mediated shRNA
transduction. Two independent shRNAs (shRNA-1 and shRNA-2) and a
negative control shRNA (shRNA-NC) were subcloned into the vector
GV112 (Fig. 3E). After lentiviral
transduction was stabilized, FGFR2 expression was verified both at
the protein and the mRNA level. Western blotting showed that the
protein expression in the shRNA-2 cells was significantly
suppressed (Fig. 3F) and the mRNA
level in the shRNA-2 cells was >50% lower than shRNA-NC cells
(Fig. 3G). The shRNA-1 was not as
effective as shRNA-2 in the knockdown of FGFR2. FGFR2 protein and
mRNA expression was similar between shRNA-NC and normal SGC7901
cells (Fig. 3F and G). FGF7
treatment (10 ng/ml for 24 h) elevated the invasive and migratory
abilities of shRNA-NC cells compared to shRNA-2 cells (Fig. 3H and I). Thus, FGF7/FGFR2 is
involved in the invasion and migration of human gastric cancer
cells.
THBS1 is required for the effect of
FGF7/FGFR2 on invasion and migration in gastric cancer cells
Next, we investigated the relation of FGF7/FGFR2 and
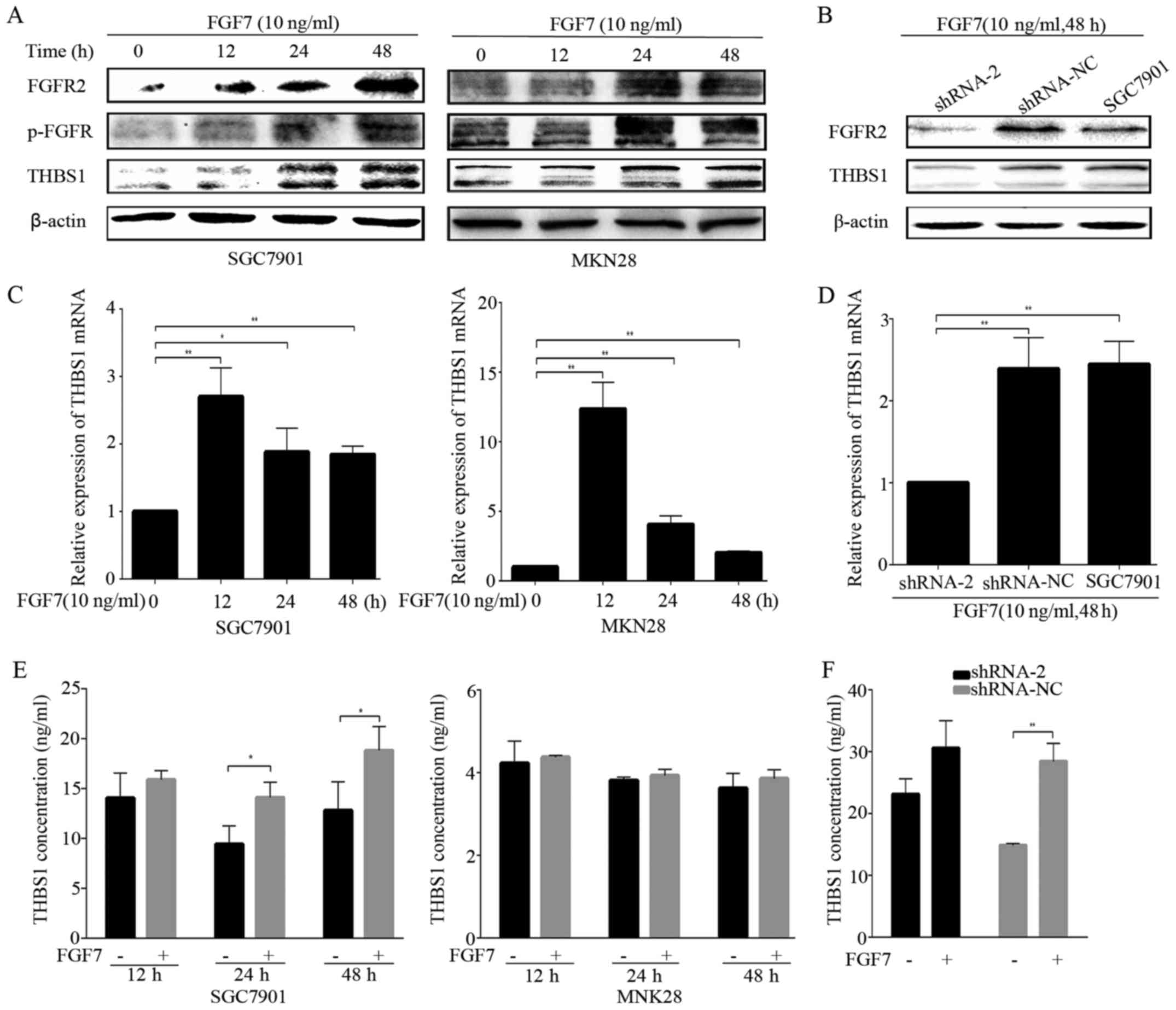
THBS1 in vitro. FGF7 treatment elevated the expression of
FGFR2, p-FGFR and THBS1 in both SGC7901 and MKN28 gastric cancer
cells in a time-dependent manner (Fig.
4A). THBS1 mRNA expression was also elevated after FGF7
treatment in both gastric cancer cells (Fig. 4C). Based on this observation, we
sought to confirm the potential relation between FGF7/FGFR2 and
THBS1, using the FGFR2 knockdown SGC7901 cells. The shRNA-2 hairpin
potently inhibited THBS1 upon treatment with FGF7 (10 ng/ml for 48
h), while shRNA-NC did not decrease the THBS1 protein and mRNA
level (Fig. 4B and D). It has been
previously noted that THBS1 in the extracellular matrix modulates
tumor progress and metastasis, therefore, we evaluted THBS1
secretion by ELISA. SGC7901 and MKN28 cells were exposed to FGF7
(10 ng/ml) and incubated for 12, 24 or 48 h. The level of
FGF7-enhanced THBS1 secretion increased significantly after 24 h of
stimulation in SGC7901 cells, while MKN28 cells showed a slight,
but statistically insignificant increase in secretion (Fig. 4E). THBS1 in the conditioned media
of shRNA-NC transfected SGC7901 cells was significantly increased
after treatment with FGF7, while no significant alternation was
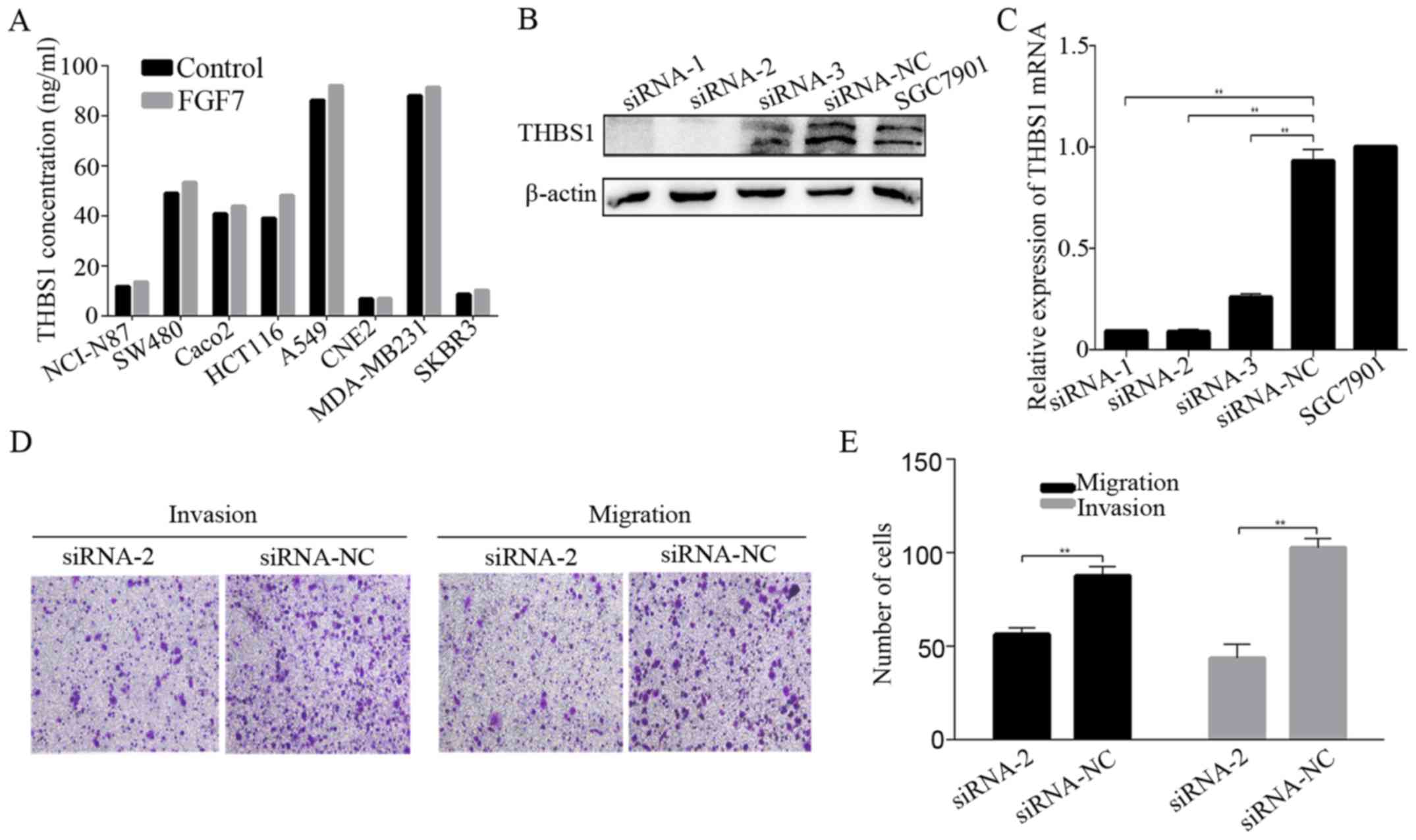
observed in FGFR2 shRNA-2 transfected cells (Fig. 4F). Similarly, FGF7 enhanced the
secretion of THBS1 in most other cell lines derived from cancers of
various locations except CNE2 (Fig.
5A).
To define the role of THBS1 in FGF7-induced invasion
and migration, we knocked down THBS1 expression in SGC7901 cells
using siRNA and Lipofectamine 2000. The knockdown proved succesful
(Fig. 5B and C). Invasion and
migration assays revealed that knowdown of THBS1 inhibited
FGF7-induced invasion and migration (Fig. 5D and E), indicating that THBS1 is
required for the effect of FGF7 on invasion and migration in
gastric cancer cells.
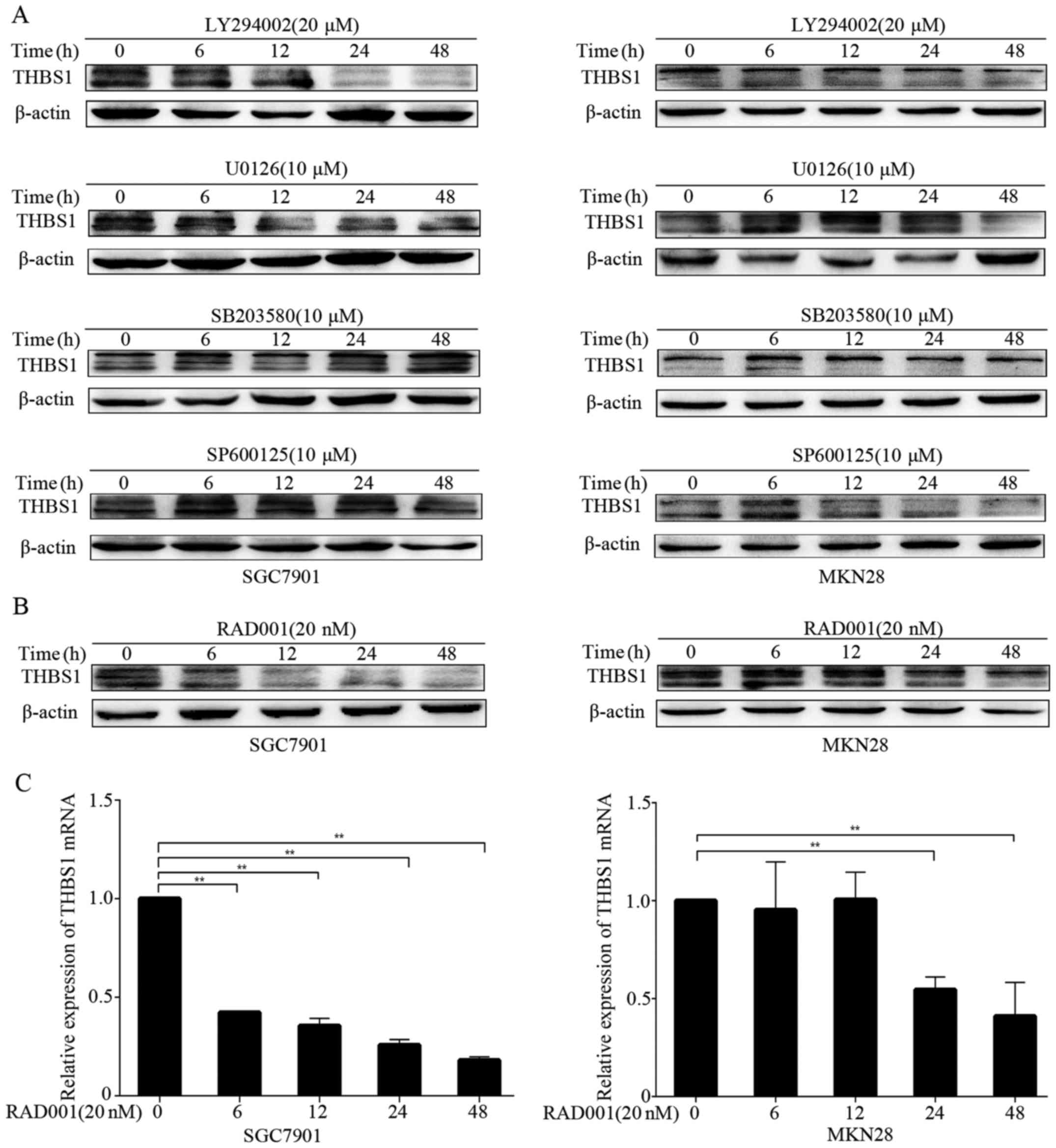
FGF7 regulates THBS1 through
PI3K/Akt/mTOR pathway in vitro
Because FGF7/FGFR2 upregulated THBS1, we aimed to
elucidate the underlying molecular mechanisms by using several
small-molecular inhibitors, including LY294002 (PI3K inhibitor),
U0126 (MEK inhibitor), SB203580 (p38 MAPK inhibitor) and SP600125
(JNK inhibitor). After pretreatment of the cells with FGF7, we
examined the level of THBS1 in the presence of small-molecular
inhibitors by western blotting at different time-points. We
observed inhibition of THBS1 after treatment with LY294002 in both
SGC7901 and MKN28 cells (Fig. 6A).
However, other inhibitors did not decrease THBS1 expression in
either cell line. These results suggested that FGF7/FGFR2 modulated
THBS1 mainly through the PI3K pathway. Furthermore, we examined
whether mTOR participates in the modulation. As shown in Fig. 6B and C, the mTOR inhibitor RAD001
significiantly inhibited THBS1 protein and mRNA expression in
SGC7901 and MKN28 cells. Thus, FGF7 upregulates THBS1 through the
PI3K/Akt/mTOR pathway in vitro.
Discussion
Overexpression of FGFR2 has been reported in various
cancers including lung, pancreatic and breast cancers (26–28),
and FGFR2 has been suggested as a novel therapeutic target
(29,30). While increased expression of FGFR2
in gastric cancer has been reported (31,32),
its role is not clear. To elucidate its role in gastric cancer, we
examined FGFR2 expression in human gastric cancer tissues. FGFR2
was generally overexpressed in cancer vs. adjacent normal tissues.
Especially, FGFR2 was highly expressed in T4 or stage III and IV
patients, and its increased expression tended to correlate with
invasion and migration. The 5-year survival rates for stage I and
II patients exceeded 60%, while the rates for stage III and IV
patients rarely exceeded 50% (33). Despite many advances in diagnosis
and treatment of this disease, the prognosis for gastric cancer
remains poor, especially in more advanced stages. Therefore, we
evaluated two groups of clinical stage of gastric cancer together
(stage I together with stage II and stage III together with stage
IV). Similarly, FGFR2 expression significantly associated with
tumor depth and survival of patients in esophagogastric junction
adenocarcinoma (34). The study by
Nakazawa et al (35)
suggested that gastric fibroblast expressed FGF7, whereas gastric
cancer cells did not and FGFR2 was expressed in gastric cancer
cells, while not expressed in fibroblast, indicating that FGF7
stimulated proliferation of gastric cancer cells in a paracrine
manner. In gastric cancer cell lines, we found that administration
of FGF7, the ligand of FGFR2, induced cell invasion and migration,
while FGFR2 shRNA-2- transfected cells treated with FGF7 showed a
decrease in invasion and migration. These lines of evidence suggest
that FGF7/FGFR2 plays an important role in invasion and migration
of gastric cancer. These data are consistent with previous
reporting of FGFR2 as a candidate therapeutic target (36).
THBS1 is a multifunctional protein that exerts a
variety of biological activities. Immunohistochemimcal staining for
THBS1 was detected both in the cytoplasm of the gastric cancer
cells and the stromal tissue. It has been reported that THBS1 was
expressed in the cytoplasm of the tumor cells of the thyroid,
breast and colorectal carcinoma (37–39).
Whereas, there were some reports indicating that THBS1 only
expressed in the stromal tissues of certain cancers including
bladder, pancreas and prostate cancer (40–42).
Lin et al (43) revealed
that THBS1 was mainly located in stromal myofibroblasts in gastric
carcinoma tissues. While Nakao et al (44) found positive staining for THBS1 in
the cytoplasm of the gastric cancer cells. Weak staining of THBS1
was occasionally detected in the stromal tissues in the study by
Zhang et al (45).
Therefore, pattern of THBS1 expression in cancer may depend on
organs or histological types, differences in the populations
studied and the antibodies used in the studies. Although the role
of THBS1 in angiogenesis is well documented, its role in tumor
metastasis is only just emerging. THBS1 expression and secretion
was elevated in melanoma cells and was associated with
epithelial-to-mesenchymal transition (20). A study showed that THBS1 was
inversely correlated with the degree of invasion in papillary
thyroid carcinoma (37), while
other studies reported that THBS1 was a potent stimulator of cell
migration and invasion in prostate, colon and breast cancers
(46–48). In addition, increased primary tumor
growth and decreased metastases were observed in THBS1-knockdown
animal model of breast cancer (49). Thus, the role of THBS1 in cancer
cell invasion and migration have yielded mixed results.
Our data demonstrated a marked overexpression of
THBS1 in gastric cancer tissues when compared to adjacent normal
tissues. Nevertheless, THBS1 was correlated with well and moderate
tumor differentiation and this is clearly a less aggressive tumor
biology. THBS1 in tumor microenvironment modulates angiogenesis,
adhesion, proliferation, invasion, migration and immunity. The
pleiotropic nature of THBS1 depends on the environment conditions,
and the presence of its different receptors may have different,
even opposite, effects on cell behavior and biological process.
This result was a comprehensive effect of THBS1 and needed more
patient samples and survival analysis. Thus, the effect of THBS1 on
gastric cancer progression needs further investigation.
Researchers have found that there exists a
relationship between FGF family and THBS1. In the study by Yu et
al (50), they demonstrated
that THBS1-derived molecules inhibited FGF2-stimulated
angiogenesis. Taraboletti et al (51) concluded the FGF2-binding sequence
of THBS1 served as a template for the development of non-peptide
inhibitors of angiogenesis. Expression of THBS1 is upregulated in
response to various cytokines and growth factors including
platelet-derived growth factor, epidermal growth factor but
downregulated in response to interleukin-1β, tumor necrosis factor
κ, interleukin-6 and bFGF (52,53).
During prostate cancer progression, production of THBS1 was
downregulated, while FGF2 was rised (54). FGF8-activated signaling pathways
mediated THBS1 repression in breast cancer cells (55). Based on these observations, the
exact role of THBS1, in particular after FGF7 binding of FGFR2 in
gastric cancer, remains unclear. The present study demonstrated
that THBS1 expression was positively correlated with FGFR2
expression. Moreover, FGF7 promoted THBS1 expression in
vitro, and enhanced the secretion of THBS1 in most cell lines
derived from cancers of various locations except MKN28 and CNE2,
which needs further investigation. Knowdown of THBS1 inhibited
FGF7-stimulated increase in invasion and migration. Together, these
findings suggested that FGF7/FGFR2 may promote gastric cancer cell
invasion and migration through a THBS1-mediated pathway.
FGF binding to their FGFRs activates multiple
transduction pathways. Thus, we next screened which signaling
pathway is involved in FGF7/FGFR2-driven regulation of THBS1
expression. The key pathways in FGF signaling are the RAS-MAP
kinase pathway which includes ERK1/2, p38 and JNK kinase and the
PI3K-Akt pathway (56). Using a
series of inhibitors, we were able to demonstrate that
FGF7/FGFR2-mediated upregulation of THBS1 may occur through the
PI3K/Akt/mTOR pathway. Further studies are warranted to elucidate
the detailed mechanism.
Activation of FGFR2 signal pathway is related to
disease progression in gastric cancer, thus, FGFR2-targeted therapy
is considered promising. Unfortunately, the multi-targeted tyrosine
kinase inhibitors (TKIs) with FGFR inhibitory activity has not been
demonstrated superior to standard chemotherapy in most phase III
clinical trials. FGF7/FGFR2-related signaling cascades play
important roles in gastric cancer progression through diverse
mechnisms, thus, FGFR2 regulates different pathways in different
gastric cancer cells. The identification of biomarkers of
FGFR2-based therapy can help to choose the right patient population
in personalized therapy. The combination of FGFR2 inhibitors and
other agents may deliver better responses to gastric cancer
patients (57). Here, we
hypothesize that FGFR2 inhibitors combined with THBS1 inhibitors or
mTOR inhibitors may benefit gastric cancer patients. Continued
basic research is needed to gain more insights into the FGFR2
signal pathway in tumor progression.
The present study has limited sample size and lack
of stage IV patients. Further investigations, such as survival
analysis, the expression of FGF7 in gastric cancer patients and
relevant animal experiments, are necessary to clarify the role of
FGF7/FGFR2/THBS1 signal in gastric cancer progression.
In conclusion, our results demonstrated that
FGF7/FGFR2 signal promotes invasion and migration in human gastric
cancer through the upregulation of thrombospondin 1. To the best of
our knowledge, this is the first study to characterize the
relationship of FGF7/FGFR2 and THBS1 in gastric cancer, which is
significantly associated with poor prognosis by promoting invasion
and migration. Our findings support further study of the
FGF7/FGFG2/THBS1 as a potential target for gastric cancer.
Abbreviations:
|
FGF
|
fibroblast growth factor
|
|
FGFR
|
fibroblast growth factor receptor
|
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grants nos. 81372664 and
81372434).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sakuramoto S, Sasako M, Yamaguchi T,
Kinoshita T, Fujii M, Nashimoto A, Furukawa H, Nakajima T, Ohashi
Y, Imamura H, et al ACTS-GC Group: Adjuvant chemotherapy for
gastric cancer with S-1, an oral fluoropyrimidine. N Engl J Med.
357:1810–1820. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Macdonald JS, Smalley SR, Benedetti J,
Hundahl SA, Estes NC, Stemmermann GN, Haller DG, Ajani JA,
Gunderson LL, Jessup JM, et al: Chemoradiotherapy after surgery
compared with surgery alone for adenocarcinoma of the stomach or
gastroesophageal junction. N Engl J Med. 345:725–730. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cunningham D, Allum WH, Stenning SP,
Thompson JN, Van de Velde CJ, Nicolson M, Scarffe JH, Lofts FJ,
Falk SJ, Iveson TJ, et al MAGIC Trial Participants: Perioperative
chemotherapy versus surgery alone for resectable gastroesophageal
cancer. N Engl J Med. 355:11–20. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Power DG, Kelsen DP and Shah MA: Advanced
gastric cancer–slow but steady progress. Cancer Treat Rev.
36:384–392. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thrumurthy SG, Chaudry MA, Chau I and
Allum W: Does surgery have a role in managing incurable gastric
cancer? Nat Rev Clin Oncol. 12:676–682. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Beenken A and Mohammadi M: The FGF family:
Biology, pathophysiology and therapy. Nat Rev Drug Discov.
8:235–253. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Knights V and Cook SJ: De-regulated FGF
receptors as therapeutic targets in cancer. Pharmacol Ther.
125:105–117. 2010. View Article : Google Scholar
|
|
9
|
Finch PW, Rubin JS, Miki T, Ron D and
Aaronson SA: Human KGF is FGF-related with properties of a
paracrine effector of epithelial cell growth. Science. 245:752–755.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Grose R and Dickson C: Fibroblast growth
factor signaling in tumorigenesis. Cytokine Growth Factor Rev.
16:179–186. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yamayoshi T, Nagayasu T, Matsumoto K, Abo
T, Hishikawa Y and Koji T: Expression of keratinocyte growth
factor/fibroblast growth factor-7 and its receptor in human lung
cancer: Correlation with tumour proliferative activity and patient
prognosis. J Pathol. 204:110–118. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Toyokawa T, Yashiro M and Hirakawa K:
Co-expression of keratinocyte growth factor and K-sam is an
independent prognostic factor in gastric carcinoma. Oncol Rep.
21:875–880. 2009.PubMed/NCBI
|
|
13
|
Cho K, Ishiwata T, Uchida E, Nakazawa N,
Korc M, Naito Z and Tajiri T: Enhanced expression of keratinocyte
growth factor and its receptor correlates with venous invasion in
pancreatic cancer. Am J Pathol. 170:1964–1974. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Wang H, Toratani S, Sato JD, Kan
M, McKeehan WL and Okamoto T: Growth inhibition by keratinocyte
growth factor receptor of human salivary adenocarcinoma cells
through induction of differentiation and apoptosis. Proc Natl Acad
Sci USA. 98:11336–11340. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ricol D, Cappellen D, El Marjou A,
Gil-Diez-de-Medina S, Girault JM, Yoshida T, Ferry G, Tucker G,
Poupon MF, Chopin D, et al: Tumour suppressive properties of
fibroblast growth factor receptor 2-IIIb in human bladder cancer.
Oncogene. 18:7234–7243. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Diez de Medina SG, Chopin D, El Marjou A,
Delouvée A, LaRochelle WJ, Hoznek A, Abbou C, Aaronson SA, Thiery
JP and Radvanyi F: Decreased expression of keratinocyte growth
factor receptor in a subset of human transitional cell bladder
carcinomas. Oncogene. 14:323–330. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Carlson CB, Lawler J and Mosher DF:
Structures of thrombospondins. Cell Mol Life Sci. 65:672–686. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Baenziger NL, Brodie GN and Majerus PW: A
thrombin-sensitive protein of human platelet membranes. Proc Natl
Acad Sci USA. 68:240–243. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Borsotti P, Ghilardi C, Ostano P, Silini
A, Dossi R, Pinessi D, Foglieni C, Scatolini M, Lacal PM, Ferrari
R, et al: Thrombospondin-1 is part of a Slug-independent motility
and metastatic program in cutaneous melanoma, in association with
VEGFR-1 and FGF-2. Pigment Cell Melanoma Res. 28:73–81. 2015.
View Article : Google Scholar
|
|
20
|
Jayachandran A, Anaka M, Prithviraj P,
Hudson C, McKeown SJ, Lo PH, Vella LJ, Goding CR, Cebon J and
Behren A: Thrombospondin 1 promotes an aggressive phenotype through
epithelial-to-mesenchymal transition in human melanoma. Oncotarget.
5:5782–5797. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pal SK, Nguyen CT, Morita KI, Miki Y,
Kayamori K, Yamaguchi A and Sakamoto K: THBS1 is induced by TGFB1
in the cancer stroma and promotes invasion of oral squamous cell
carcinoma. J Oral Pathol Med. 5:730–739. 2016. View Article : Google Scholar
|
|
22
|
Roberts DD: Regulation of tumor growth and
metastasis by thrombospondin-1. FASEB J. 10:1183–1191.
1996.PubMed/NCBI
|
|
23
|
Nie S, Lo A, Wu J, Zhu J, Tan Z, Simeone
DM, Anderson MA, Shedden KA, Ruffin MT and Lubman DM: Glycoprotein
biomarker panel for pancreatic cancer discovered by quantitative
proteomics analysis. J Proteome Res. 13:1873–1884. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lyu T, Jia N, Wang J, Yan X, Yu Y, Lu Z,
Bast RC Jr, Hua K and Feng W: Expression and epigenetic regulation
of angiogenesis-related factors during dormancy and recurrent
growth of ovarian carcinoma. Epigenetics. 8:1330–1346. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Perez-Janices N, Blanco-Luquin I, Tuñón
MT, Barba-Ramos E, Ibáñez B, Zazpe-Cenoz I, Martinez-Aguillo M,
Hernandez B, Martínez-Lopez E, Fernández AF, et al: EPB41L3, TSP-1
and RASSF2 as new clinically relevant prognostic biomarkers in
diffuse gliomas. Oncotarget. 6:368–380. 2015.PubMed/NCBI
|
|
26
|
Nomura S, Yoshitomi H, Takano S, Shida T,
Kobayashi S, Ohtsuka M, Kimura F, Shimizu H, Yoshidome H, Kato A,
et al: FGF10/FGFR2 signal induces cell migration and invasion in
pancreatic cancer. Br J Cancer. 99:305–313. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Behrens C, Lin HY, Lee JJ, Raso MG, Hong
WK, Wistuba II and Lotan R: Immunohistochemical expression of basic
fibroblast growth factor and fibroblast growth factor receptors 1
and 2 in the pathogenesis of lung cancer. Clin Cancer Res.
14:6014–6022. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bane AL, Pinnaduwage D, Colby S, Reedijk
M, Egan SE, Bull SB, O'Malley FP and Andrulis IL: Expression
profiling of familial breast cancers demonstrates higher expression
of FGFR2 in BRCA2-associated tumors. Breast Cancer Res Treat.
117:183–191. 2009. View Article : Google Scholar :
|
|
29
|
Byron SA, Gartside MG, Wellens CL, Mallon
MA, Keenan JB, Powell MA, Goodfellow PJ and Pollock PM: Inhibition
of activated fibroblast growth factor receptor 2 in endometrial
cancer cells induces cell death despite PTEN abrogation. Cancer
Res. 68:6902–6907. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Byron SA and Pollock PM: FGFR2 as a
molecular target in endometrial cancer. Future Oncol. 5:27–32.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Han N, Kim MA, Lee HS and Kim WH:
Evaluation of fibroblast growth factor receptor 2 expression,
heterogeneity and clinical significance in gastric cancer.
Pathobiology. 82:269–279. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ahn S, Lee J, Hong M, Kim ST, Park SH,
Choi MG, Lee JH, Sohn TS, Bae JM, Kim S, et al: FGFR2 in gastric
cancer: protein overexpression predicts gene amplification and high
H-index predicts poor survival. Mod Pathol. 29:1095–1103. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang J, Zhou Y, Jiang K, Shen Z, Ye Y and
Wang S: Evaluation of the seventh AJCC TNM staging system for
gastric cancer: a meta-analysis of cohort studies. Tumour Biol.
35:8525–8532. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tokunaga R, Imamura Y, Nakamura K,
Ishimoto T, Nakagawa S, Miyake K, Nakaji Y, Tsuda Y, Iwatsuki M,
Baba Y, et al: Fibroblast growth factor receptor 2 expression, but
not its genetic amplification, is associated with tumor growth and
worse survival in esophagogastric junction adenocarcinoma.
Oncotarget. 7:19748–19761. 2016.PubMed/NCBI
|
|
35
|
Nakazawa K, Yashiro M and Hirakawa K:
Keratinocyte growth factor produced by gastric fibroblasts
specifically stimulates proliferation of cancer cells from
scirrhous gastric carcinoma. Cancer Res. 63:8848–8852.
2003.PubMed/NCBI
|
|
36
|
Sommer A, Kopitz C, Schatz CA, Nising CF,
Mahlert C, Lerchen HG, Stelte-Ludwig B, Hammer S, Greven S,
Schuhmacher J, et al: Preclinical efficacy of the auristatin-based
antibody-drug conjugate BAY 1187982 for the treatment of
FGFR2-positive solid tumors. Cancer Res. 76:6331–6339. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tanaka K, Sonoo H, Kurebayashi J, Nomura
T, Ohkubo S, Yamamoto Y and Yamamoto S: Inhibition of infiltration
and angiogenesis by thrombospondin-1 in papillary thyroid
carcinoma. Clin Cancer Res. 8:1125–1131. 2002.PubMed/NCBI
|
|
38
|
Maeda K, Nishiguchi Y, Yashiro M, Yamada
S, Onoda N, Sawada T, Kang SM and Hirakawa K: Expression of
vascular endothelial growth factor and thrombospondin-1 in
colorectal carcinoma. Int J Mol Med. 5:373–378. 2000.PubMed/NCBI
|
|
39
|
Clezardin P, Frappart L, Clerget M,
Pechoux C and Delmas PD: Expression of thrombospondin (TSP1) and
its receptors (CD36 and CD51) in normal, hyperplastic, and
neoplastic human breast. Cancer Res. 53:1421–1430. 1993.PubMed/NCBI
|
|
40
|
Kasper HU, Ebert M, Malfertheiner P,
Roessner A, Kirkpatrick CJ and Wolf HK: Expression of
thrombospondin-1 in pancreatic carcinoma: correlation with
microvessel density. Virchows Archiv. 438:116–120. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Grossfeld GD, Ginsberg DA, Stein JP,
Bochner BH, Esrig D, Groshen S, Dunn M, Nichols PW, Taylor CR,
Skinner DG, et al: Thrombospondin-1 expression in bladder cancer:
Association with p53 alterations, tumor angiogenesis, and tumor
progression. J Natl Cancer Inst. 89:219–227. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kwak C, Jin RJ, Lee C, Park MS and Lee SE:
Thrombospondin-1, vascular endothelial growth factor expression and
their relationship with p53 status in prostate cancer and benign
prostatic hyperplasia. BJU Int. 89:303–309. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lin XD, Chen SQ, Qi YL, Zhu JW, Tang Y and
Lin JY: Overexpression of thrombospondin-1 in stromal
myofibroblasts is associated with tumor growth and nodal metastasis
in gastric carcinoma. J Surg Oncol. 106:94–100. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nakao T, Kurita N, Komatsu M, Yoshikawa K,
Iwata T, Utsunomiya T and Shimada M: Expression of thrombospondin-1
and Ski are prognostic factors in advanced gastric cancer. Int J
Clin Oncol. 16:145–152. 2011. View Article : Google Scholar
|
|
45
|
Zhang J, Ito R, Oue N, Zhu X, Kitadai Y,
Yoshida K, Nakayama H and Yasui W: Expression of thrombospondin-1
is correlated with microvessel density in gastric carcinoma.
Virchows Archiv. 442:563–568. 2003.PubMed/NCBI
|
|
46
|
Radziwon-Balicka A, Santos-Martinez MJ,
Corbalan JJ, O'Sullivan S, Treumann A, Gilmer JF, Radomski MW and
Medina C: Mechanisms of platelet-stimulated colon cancer invasion:
Role of clusterin and thrombospondin 1 in regulation of the
P38MAPK-MMP-9 pathway. Carcinogenesis. 35:324–332. 2014. View Article : Google Scholar
|
|
47
|
Firlej V, Mathieu JR, Gilbert C, Lemonnier
L, Nakhlé J, Gallou-Kabani C, Guarmit B, Morin A, Prevarskaya N,
Delongchamps NB, et al: Thrombospondin-1 triggers cell migration
and development of advanced prostate tumors. Cancer Res.
71:7649–7658. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang TN, Qian X, Granick MS, Solomon MP,
Rothman VL, Berger DH and Tuszynski GP: Thrombospondin-1 (TSP-1)
promotes the invasive properties of human breast cancer. J Surg
Res. 63:39–43. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yee KO, Connolly CM, Duquette M,
Kazerounian S, Washington R and Lawler J: The effect of
thrombospondin-1 on breast cancer metastasis. Breast Cancer Res
Treat. 114:85–96. 2009. View Article : Google Scholar :
|
|
50
|
Yu H, Tyrrell D, Cashel J, Guo NH, Vogel
T, Sipes JM, Lam L, Fillit HM, Hartman J, et al: Specificities of
heparin-binding sites from the amino-terminus and type 1 repeats of
thrombospondin-1. Arch Biochem Biophys. 374:13–23. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Taraboletti G, Rusnati M, Ragona L and
Colombo G: Targeting tumor angiogenesis with TSP-1-based compounds:
Rational design of antiangiogenic mimetics of endogenous
inhibitors. Oncotarget. 1:662–673. 2010. View Article : Google Scholar
|
|
52
|
Morandi V, Cherradi SE, Lambert S,
Fauvel-Lafève F, Legrand YJ and Legrand C: Proinflammatory
cytokines (interleukin-1 beta and tumor necrosis factor-alpha) down
regulate synthesis and secretion of thrombospondin by human
endothelial cells. J Cell Physiol. 160:367–377. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Loganadane LD, Berge N, Legrand C and
Fauvel-Lafève F: Endothelial cell proliferation regulated by
cytokines modulates thrombospondin-1 secretion into the
subendothelium. Cytokine. 9:740–746. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Doll JA, Reiher FK, Crawford SE, Pins MR,
Campbell SC and Bouck NP: Thrombospondin-1, vascular endothelial
growth factor and fibroblast growth factor-2 are key functional
regulators of angiogenesis in the prostate. Prostate. 49:293–305.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tarkkonen K, Ruohola J and Härkönen P:
Fibroblast growth factor 8 induced downregulation of thrombospondin
1 is mediated by the MEK/ERK and PI3K pathways in breast cancer
cells. Growth Factors. 28:256–267. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Dailey L, Ambrosetti D, Mansukhani A and
Basilico C: Mechanisms underlying differential responses to FGF
signaling. Cytokine Growth Factor Rev. 16:233–247. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Hong L, Han Y, Liu J and Brain L:
Fibroblast growth factor receptor 2: A therapeutic target in
gastric cancer. Expert Rev Gastroenterol Hepatol. 7:759–765. 2013.
View Article : Google Scholar : PubMed/NCBI
|