Introduction
Colorectal cancer (CRC) is a common malignant tumor
involving approximately 14 million new cases per year, and is the
cause of approximately 690,000 deaths worldwide each year (1). The incidence of CRC increases with
age; 79% of CRC patients are >60 years of age when diagnosed,
and 34% are >80 years of age when diagnosed (2). Furthermore, it is predicted that the
incidence of CRC in elderly patients will increase in the future.
The pathogenesis of CRC involves activation of an oncogene and
inactivation of a tumor suppressor gene (3), and the tumor suppressor gene
inactivation is closely related to epigenetic changes. These
epigenetic changes include DNA methylation, histone acetylation,
and chromatin remodeling (4). DNA
methylation plays an important role in the development and
progression of different cancers such as CRC. Increased evidence
has suggested that promoter methylation of tumor suppressor genes
can be used as a tumor marker in the early diagnosis of multiple
cancers (5,6). When CRC is diagnosed, it is often
already in an advanced stage, so it is important to identify
markers for early diagnosis of CRC.
Zinc finger proteins containing the Kruppel
associated box (KRAB-ZFPs) belong to the zinc finger protein
family, which is the largest family of transcription factors in
mammals (7,8). KRAB-ZFPs regulate cell proliferation,
differentiation, apoptosis, and formation of tumor cells, and play
an important role in biological evolution (9). KRAB-ZFPs are transcription factors
that bind to the promoter of target genes to inhibit or activate
their expression (10), and to
either inhibit or promote cancinogenesis. ZIC1, ZNF569 and
ZFP932 have been reported to be tumor suppressor genes
(11–13), and KLF5, SALL4 and
ZNFEB have been reported to be oncogenes (14–16).
ZNF545, located on the 19q13 chromosome, is a
member of the KRAB-ZFP family of tumor suppressor genes, and is
downregulated and frequently methylated in multiple types of tumors
(17–19). However, the effect of epigenetic
inactivation of ZNF545 in colorectal carcinogenesis and its
possible use as a tumor marker remain unclear. We therefore
characterized the methylation status and expression of
ZNF545 in CRC, together with its related functions and
mechanism of action.
Materials and methods
Cell lines and tumor samples
Five CRC cell lines (HT-29, SW480, HCT116, CaCo-2,
and LoVo) were used. The CRC cell lines, HT-29 and HCT116, were
provided by Professor Q. Tao at the Chinese University of Hong
Kong, and the SW480, CaCo-2, and LoVo cell lines were purchased
from the Chinese Academy of Sciences. The cell lines were cultured
with RPMI-1640 medium (Gibco-BRL, Karlsruhe, Germany) containing
10% fetal bovine serum (FBS) (ExCell Bio, Shanghai, China), and
cultured in a 5% CO2 incubator at 37°C. The CRC tissues
and paracarcinoma tissues, which were diagnosed by a pathologist,
were obtained from patients during surgery at the First Affiliated
Hospital of Chongqing Medical University (Chongqing, China). All
patients signed an informed consent form, and the research protocol
was approved by the Institutional Ethics Committee of the First
Affiliated Hospital of Chongqing Medical University.
RNA, DNA, and protein extraction
Total RNA was extracted separately from 32 paired
CRC tissues, surgical margin tissues, and five CRC cell lines using
TRIzol® reagent (Life Technologies, Carlsbad, CA, USA).
Genomic DNA was separately obtained from 24 CRC tissues, six normal
colorectal tissues, and the five CRC cell lines using the QIAamp
DNA Mini kit (Qiagen, Hilden, Germany) according to the
manufacturer's protocol. The concentration of DNA and RNA were
measured using a NanoDrop 2000 Spectrophotometer (Thermo
Scientific, Rockford, IL, USA) and stored at −80°C. The
experimental and control group of the HT-29 and SW480 cells were
lysed using a protein extraction reagent (Thermo Scientific) that
contained the protease inhibitor, phenylmethane sulfonyl fluoride,
and a phosphatase inhibitor cocktail (Sigma-Aldrich, St. Louis, MO,
USA), and the lysate was then homogenized using a Ultrasonic Cell
Grinder (Scientz, Ningbo). The supernatant was collected after
centrifugation, and the concentration of protein in the supernatant
was determined using the BCA protein kit (Thermo Scientific).
Semiquantitative polymerase chain
reaction (PCR) and real-time PCR
ZNF545 expression in CRC cells and tissues
was determined using semiquantitative PCR and quantitative PCR. The
RNA (1 µg) was reverse transcribed to 20 µg of cDNA
using the Reverse Transcription system (Promega, Madison, WI, USA).
For semiquantitative PCR, the ZNF545 gene was amplified
using GoTaq DNA polymerase (Promega) with 35 cycles, using GAPDH as
an internal control. The primer sequences are listed in Table I. Quantitative PCR was performed
with a SYBR mix green fluorescent reagent (Promega) using an ABI
7500 Real-Time PCR system (Applied Biosystems, Foster City, CA,
USA). β-actin was used as a control, and each sample was tested in
triplicate.
 | Table IThe primers used in this study. |
Table I
The primers used in this study.
| Primer | Sequences
(5′–3′) | Product sizes
(bp) | Annealing
temperature (°C) | Cycles |
|---|
| RT-PCR | ZNF545-F |
GAGCCTTGGAAAGTTGTGAG | 245 | 55 | 35 |
| q-PCR | ZNF545-R |
GGCATTTTCACACTACTGAAG |
| MSP | ZNF545-M1 |
TTTTTTTTAGGTTTTGTCGCGTC | 177 | 60 | 40 |
| ZNF545-M2 |
CTACTAAAAAAACCGAACGCG |
| ZNF545-U1 |
TTTTTTTTTAGGTTTTGTTGTGTT | 164 | 58 | 40 |
| ZNF545-U2 |
CCAAACACACTCACAAAATACA |
Methylation-specific PCR analyses of
ZNF545
As previously described (20), methylation-specific PCR (MSP) was
used to detect the ZNF545 promoter methylation status.
bisulfite-treated DNA (50 ng) was mixed with AmpliTaq Gold
polymerase (Applied Biosystems), mgCl2, and
deoxynucleotide triphosphates for the MSP amplification reaction.
The methylation-specific primers are listed in Table I. The PCR amplification was
performed for a total of 40 cycles with an annealing temperature of
60°C or 58°C for 30 sec. The final products were analyzed on a 2%
agarose gel, and then recorded using a Molecular Imager (Bio-Rad,
Hercules, CA, USA).
5-Aza-2′-deoxycytidine (Aza) and
trichostatin A (TSA) treatment
Aza, a DNA methyltransferase (DNMT) inhibitor, makes
DNMT inactivation through DNMT covalent bonding with thiol on
cysteine residues, causing reactivation genes silenced by promoter
methylation. TSA, a histone deacetylase inhibitor, plays an
important role in controlling the tightness of DNA around histone.
Combination treatment of Aza and TSA leads the synergistic
activation of methylated genes.
CRC cell lines, SW480 and HT-29, were cultured and
demethylated. The cells were treated with Aza (Sigma-Aldrich) in
the dark at a final concentration of 10 mM/l for 3 days or TSA at a
final concentration of 100 mM/l for 1 day, and further treated with
or without 100 mM/l TSA (Cayman Chemical, Ann Arbor, MI, USA) for
another 1 day.
The establishment of stable cell
lines
A ZNF545-expressing plasmid was provided by
Professor Q. Tao at the Chinese University of Hong Kong. After
SW480 and HT-29 cells were plated into 6-well plates, the cells
were transfected with pcDNA3.1-ZNF545 or pcDNA3.1 (+)
vectors using Lipofectamine® 2000 reagent (Invitrogen)
and cultured with serum-free RPMI-1640 medium. After 4–6 h, the
medium containing 10% FBS was changed to a selection medium
containing 400 µg/ml G418 (Invitrogen/Gibco) and incubated
for 48 h. Approximately 14 days later, the HT-29 and SW480 cells
were stably expressing pcDNA3.1 and pcDNA3.1-ZNF545.
The cell proliferation and colony
formation assays
SW480 and HT-29 cells stably expressing ZNF545 or
pcDNA 3.1 were seeded into 96-well plates at 2,000 cells/well, and
the cell viability was measured using MTS (Promega) at 0, 24, 48
and 72 h using a Multiskan Spectrum™ (Tecan, Switzerland). The
stably expressing cells were counted and seeded into 6-cell plates
at 500 cells/well. Visible cell clones appeared in ~2 weeks, then
they were fixed with 4% paraformaldehyde for 30 min and then
stained with crystal violet solution for 30 min. Finally, the cells
were scanned using a CanoScan 8800F (Canon, Tokyo, Japan).
Flow cytometry of the cell cycle and
apoptosis
The cell cycle and apoptosis were measured using
flow cytometry as previously described (20). For the cell cycle measurements,
SW480 and HT-29 cells stably expressing ZNF545 or pcDNA 3.1
were digested and washed twice using precooled phosphate-buffered
saline (PBS), then fixed with ice-cold 70% ethanol, stored at 4°C
overnight, and then stained with propidium iodide (PI), followed by
analysis using CellQuest software (BD Biosciences, San Jose, CA,
USA). After the stably transfected cells were collected and washed
twice with PBS, they were stained with Annexin V-FITC/PI in the
dark, and the cell apoptosis was measured using CellQuest software,
followed by data analyses.
The analyses of wound healing
Stably ZNF545- or vector-transfected SW480 or HT-29
cells were seeded into 6-well plates, and when the cells reached
95% confluency, a sterile P-20 pipette was used to scratch wounds
across each well, followed by removal of cell debris using PBS. The
cells were then cultured with medium without FBS to minimize cell
proliferation. The images of the wound closure areas of SW480 cells
were observed at 0, 48, and 60 h, and the images of the wound
closure areas of HT-29 cells were observed at 0, 24, 48, and 84 h
using a light microscope (CTR4000; Leica, Germany).
Western blot analyses
A total of 40 mg of protein lysate was separated by
sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then
transferred to a polyvinylidene fluoride (PVDF) membrane (Bio-Rad).
The primary antibodies were against cleaved caspase-3 (12742; Cell
Signaling Technology, Danvers, MA, USA), cleaved poly(ADP-ribose)
polymerase (PARP) (9541; Cell Signaling Technology), BAX (5023;
Cell Signaling Technology), ZNF545 (sc-102235; Santa Cruz
Biotechnology, Santa Cruz, CA, USA), PI3K (ab125571; Abcam,
Cambridge, UK), phospho-PI3K (Abcam), Akt (4691; Cell Signaling
Technology), phospho-Akt (ser473) (4060; Cell Signaling
Technology), phospho-GSK3β (ser9) (9323; Cell Signaling
Technology), β-catenin (2677; Cell Signaling), active β-catenin
(4270; Cell Signaling), c-Myc (1472-1; Epitomic), cyclin D1 (2261;
Epitomics), β-actin (LK-ab008-100; Liankebio, China), and Flag
(F3165; Sigma-Aldrich).
In vivo tumorigenicity
Six groups of 4-week-old nude mice were used. Empty
vector, stably transfected SW480 cells (3×106 cells
suspended in 200 µl of PBS) were injected subcutaneously
into the left dorsal flank of nude mice, and
ZNF545-transfected SW480 cells (3×106 cells
suspended in 200 µl of PBS) were injected subcutaneously
into the right dorsal flank. After the tumor mass appeared, its
length and width were measured every 3 days using a microcaliper.
As previously described (23), the
tumor volume (mm3) was calculated using the equation:
volume = length × width2/2. The mice were euthanized and
the tumors were removed when the tumor length reached 1.5 cm. All
tumor tissues were weighed before fixation in 4% paraformaldehyde
and embedded in paraffin, followed by preparation of the sections
for immunohistochemical analyses. All experimental procedures were
approved by the Animal Ethics Committee of the Experimental Animal
Center of the Chongqing Medical University, Chongqing, China.
Immunohistochemical staining
The samples of excised tumor tissues from nude mice
were fixed in 4% paraformaldehyde before being embedded in
paraffin, followed by sectioning into 4-µm slices. After the
sections were dewaxed for 2 h in a 60°C incubator, dewaxing and
alcohol dehydration were performed in the following order: xylene
I, xylene II, xylene III, xylene IV, absolute ethyl alcohol, 95%
ethyl alcohol, 80% ethyl alcohol, and 70% ethyl alcohol. Antigen
retrieval was performed by boiling the slides in citrate buffer
solution for 20 min, followed by cooling at room temperature. The
sections were then incubated in 3% hydrogen peroxide for 20 min to
prevent endogenous peroxidase activity and washed three times with
PBS, then incubated in sheep serum for 20 min. The sections were
incubated at 4°C overnight with rabbit anti-PCNA monoclonal
antibody. The next day, the sections were warmed to room
temperature for 2 h, then washed three times with PBS and incubated
with rabbit secondary antibody for 20 min. The slices were washed
three times with PBS and incubated with horseradish
peroxidase-labeled streptomycin anti-biotin antibody for 20 min.
The sections were washed three times, followed by color development
with DAB. The sections were washed with water, and the cell nuclei
were dyed using hematoxylin, followed by observation using light
microscopy.
Statistical analysis
All statistical analyses were performed using SPSS
statistical software for Windows, version 16. Student's t-test was
used to compare the expression of ZNF545 in CRC tissues and
adjacent normal tissues. The four table Chi-square test or the
t-test were used to analyze categorical variables. A value of
p<0.05 was considered statistically significant.
Results
ZNF545 is downregulated in CRC tissues
and cell lines
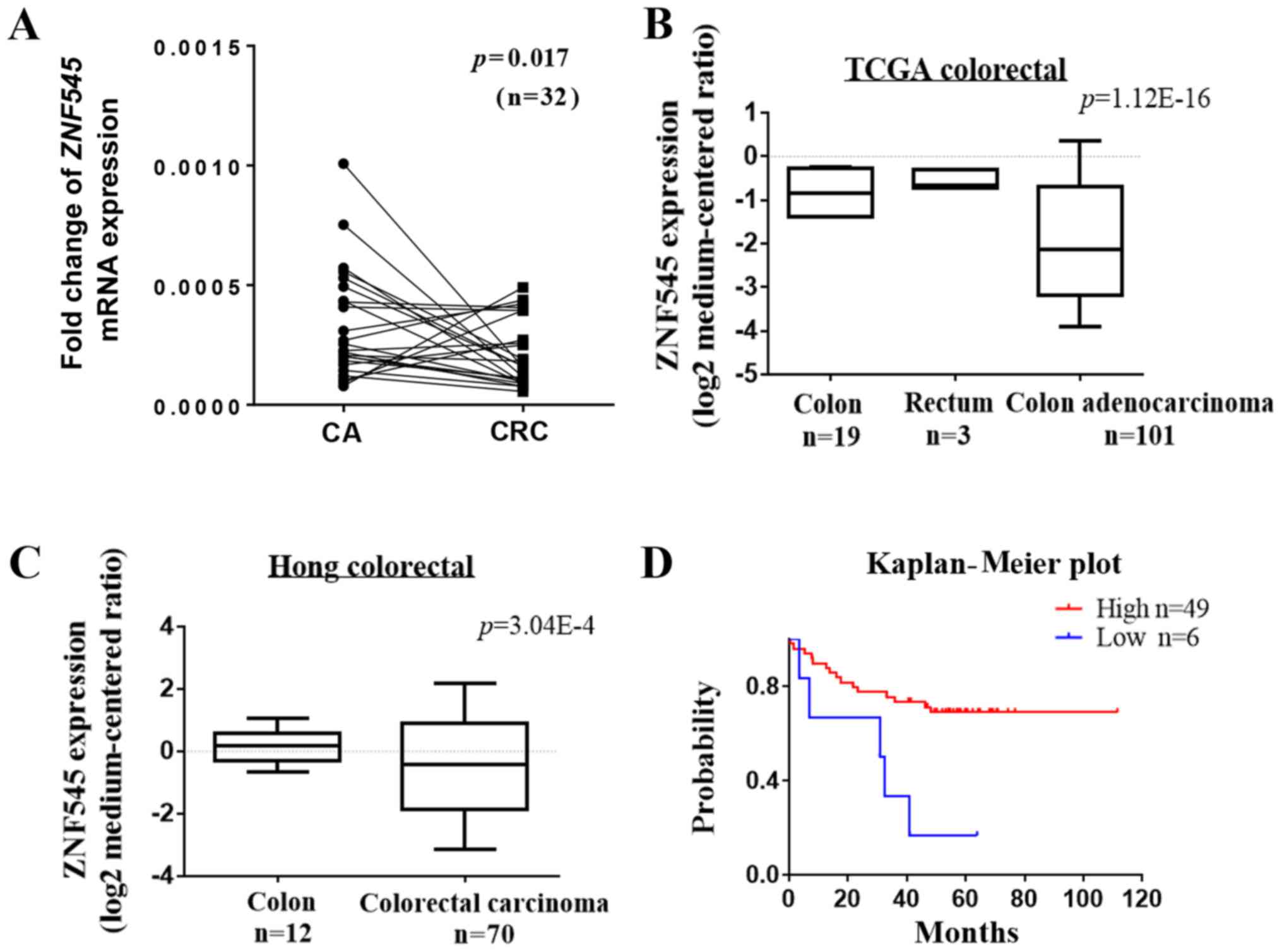
The messenger RNA (mRNA) expression of ZNF545 in CRC
and its corresponding adjacent tissues was detected by quantitative
PCR. The results showed that the expression of ZNF545 was
downregulated in tumor tissues compared with the adjacent tissue
(p<0.05, Fig. 1A). RT-PCR was
then used to examine the expression of ZNF545 in the HT-29,
SW480, HCT116, CoCo-2, and LoVo colorectal cell lines.
ZNF545 expression was downregulated or silenced in all cell
lines compared with the vector (Fig.
2A). In addition, analyses of an online microarray database
(www.oncomine.org, Compendia Bioscience, Ann
Arbor, MI, USA) showed that ZNF545 was downregulated in CRC
tissues but not in normal adult colorectal tissues (p<0.001;
Fig. 1B and C). Prognostic
analyses from the PrognoScan database (http://www.prognoscan.org/) showed that patients with
higher expression of ZNF545 mRNA had significantly increased
relapse-free survival compared with those with lower ZNF545
mRNA levels (Fig. 1D). Taken
together, these results suggested that ZNF545 was a cancer
suppressor gene.
The ZNF545 promoter was frequently
methylated during CRC
Aberrant promoter methylation is a common mechanism
involved in tumor suppressor gene silencing in cancers (21,22).
To identify whether the decreased expression of ZNF545 was
due to promoter methylation, we determined the methylation status
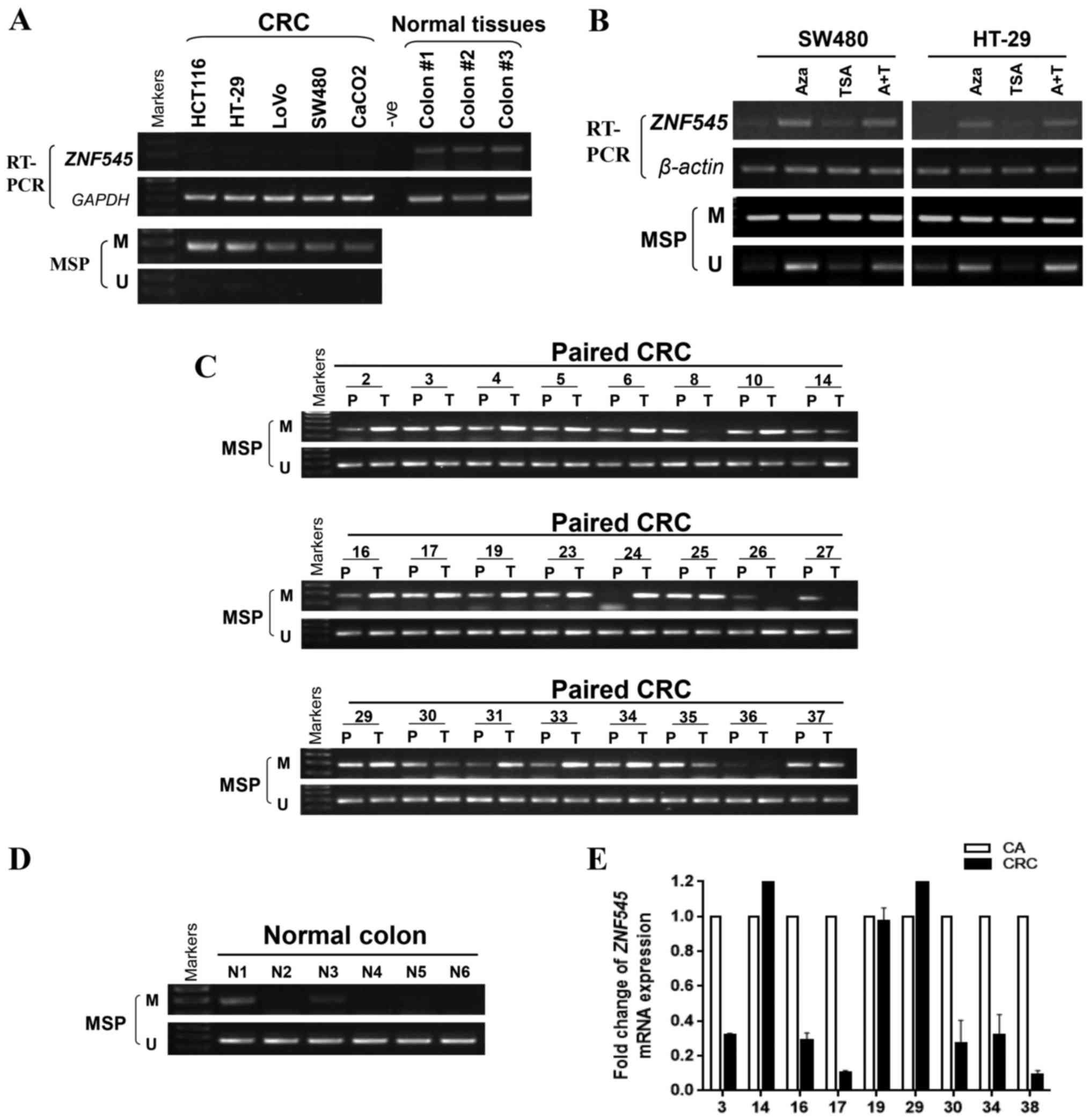
of the ZNF545 promoter in five CRC cell lines using MSP. The
results showed that ZNF545 was methylated in all cell lines
that were associated with silenced or reduced ZNF545
expression, while normal colorectal tissues expressed significantly
higher levels of ZNF545 mRNA (Fig. 2A). We then treated SW480 and HT-29
cells which have low-expression of ZNF545 with Aza, TSA and
combination, results showed that its expression was restored
together with increasing unmethylated alleles of the ZNF545
promoter (Fig. 2B). The results
suggested that promoter methylation is directly responsible for
ZNF545 downregulation in colorectal cancer cells.
To investigate whether ZNF545 promoter
methylation existed in CRC tissues, MSP was used to examine 32
primary colorectal carcinomas and six normal colorectal tissues.
The clinical information of colorectal carcinoma tissues are shown
in Table II. We observed that
ZNF545 methylation was detected in 28 out of 32 (87.5%) CRC
tissues, but in only two out of six (33.33%) normal colorectal
tissues (Fig. 2C and D and
Table III p=0.0116). Nine pairs
of primary CRC tissues and para-carcinoma tissues which were
randomly selected in the 32 CRC tissues, were also tested for
ZNF545 expression by qPCR. Most tumors have lower
ZNF545 mRNA expression compared with their paired adjacent
tissues (Fig. 2E), while
displaying a higher level of promoter methylation.
| Table IIThe clinical information of
colorectal carcinoma tissues. |
Table II
The clinical information of
colorectal carcinoma tissues.
| No. | Age | Size (cm) | Sex (1M2F) | Grade | Lymph
metastasis | Distant
metastasis | Localization | T-stage | N-status | Phase | Pathological
type |
|---|
| CRC2 | 42 | 3,2 | 2 | 3 | 0 | 0 | Middle, down | 3 | N0 | IIA | Ad |
| CRC3 | 78 | 2 | 2 | 2 | 1 | 0 | Middle | 3 | N1 | IIIB | Ad |
| CRC4 | 65 | 2 | 1 | 2 | 0 | 0 | Left | 3 | N1c | IIIB | Ad |
| CRC5 | 63 | 3 | 2 | 3 | 1 | 0 | Left | 4b | N1 | IIIC | Ad |
| CRC6 | 82 | 3,2 | 1 | 2 | 0 | 0 | Right, down | 3 | N0 | IIA | Ad |
| CRC8 | 72 | 2 | 1 | 2–3 | 0 | 0 | Middle | 3 | N0 | IIA | Ad |
| CRC10 | 76 | 2 | 2 | 2 | 0 | 0 | Right | 3 | N0 | IIA | Ad |
| CRC14 | 67 | 2 | 1 | 2–3 | 0 | 0 | Left | 1 | N0 | I | Ad |
| CRC16 | 76 | 2 | 1 | 2 | 0 | 0 | Left | 3 | N0 | IIA | Ad |
| CRC17 | 79 | 2 | 2 | 2 | 4 | 0 | Left | 3 | N2a | IIIB | Ad |
| CRC19 | 55 | 2 | 1 | 2 | 0 | 0 | Left | 3 | N0 | IIA | Ad |
| CRC23 | 76 | 2 | 2 | 2 | 2 | 0 | Left | 3 | N1 | IIIB | Ad |
| CRC24 | 44 | 2 | 2 | X | 0 | 0 | Left | 3 | N0 | IIA | Ad |
| CRC25 | 73 | 2 | 2 | 2 | 0 | 0 | Left | 3 | N0 | IIA | Ad |
| CRC26 | 62 | 3.2 | 1 | 2–3 | 7 | 0 | Left | 3 | N2b | IIIC | Ad |
| CRC27 | 66 | 2 | 1 | X | 1 | 0 | Left | 4b | N1 | IIIC | MN |
| CRC29 | 65 | 2 | 2 | 2 | 5 | 0 | Right | 3 | N2a | IIIB | Ad |
| CRC30 | 78 | 3 | 1 | 2 | 1 | 0 | Left | 3 | N1 | IIIB | Ad |
| CRC31 | 66 | 2 | 1 | 2 | 0 | 1 | Left | 3 | N1c | IVA | Ad |
| CRC33 | 78 | 2 | 2 | 2 | 0 | 0 | Right | 3 | N1 | IIIB | Ad |
| CRC34 | 70 | 2 | 1 | 2–3 | 0 | 0 | Right | 2 | N0 | I | Ad |
| CRC35 | 59 | 2 | 2 | 2 | 0 | 0 | Left | 3 | N0 | IIA | Ad |
| CRC36 | 45 | 2 | 1 | 2 | 0 | 0 | Left | 3 | N0 | IIA | Ad |
| CRC37 | 67 | 2 | 2 | 2 | 0 | 0 | Right | 3 | N0 | IIA | Ad |
| CRC38 | 76 | 2 | 2 | 2 | 1 | 0 | Right | 3 | N1 | IIIB | Ad |
| CRC39 | 64 | 2 | 1 | 2 | 0 | 0 | Left | 3 | N0 | IIA | Ad |
| CRC40 | 43 | 3 | 1 | 2 | 5 | 0 | Right | 3 | N2a | IIIB | Ad |
| CRC41 | 72 | 3 | 1 | 2 | 0 | 0 | Right | 3 | N0 | IIA | Ad |
| CRC42 | 68 | 2 | 1 | 2 | 0 | 0 | Left | 2 | N0 | I | Ad |
| CRC43 | 57 | 3 | 1 | 2 | 0 | 0 | Right | 3 | N0 | IIA | Ad |
| CRC44 | 80 | 2,2,2 | 1 | 2 | 2 | 0 | Left, down | 3 | N1 | IIIB | Ad |
| CRC45 | 54 | 2 | 2 | 2 | 0 | 0 | Right | 3 | N0 | IIA | Ad |
| Table IIIMethylation status of the
ZNF545 promoter in colorectal cancer. |
Table III
Methylation status of the
ZNF545 promoter in colorectal cancer.
| Samples | ZNF545
promoter
| Frequency of
methylation |
|---|
| Methylation | Unmethylation |
|---|
| CRC (n=32) | 28 | 4 | 87.5% |
| CN (n=6) | 2 | 4 | 33.3% |
Overexpression of ZNF545 inhibits cell
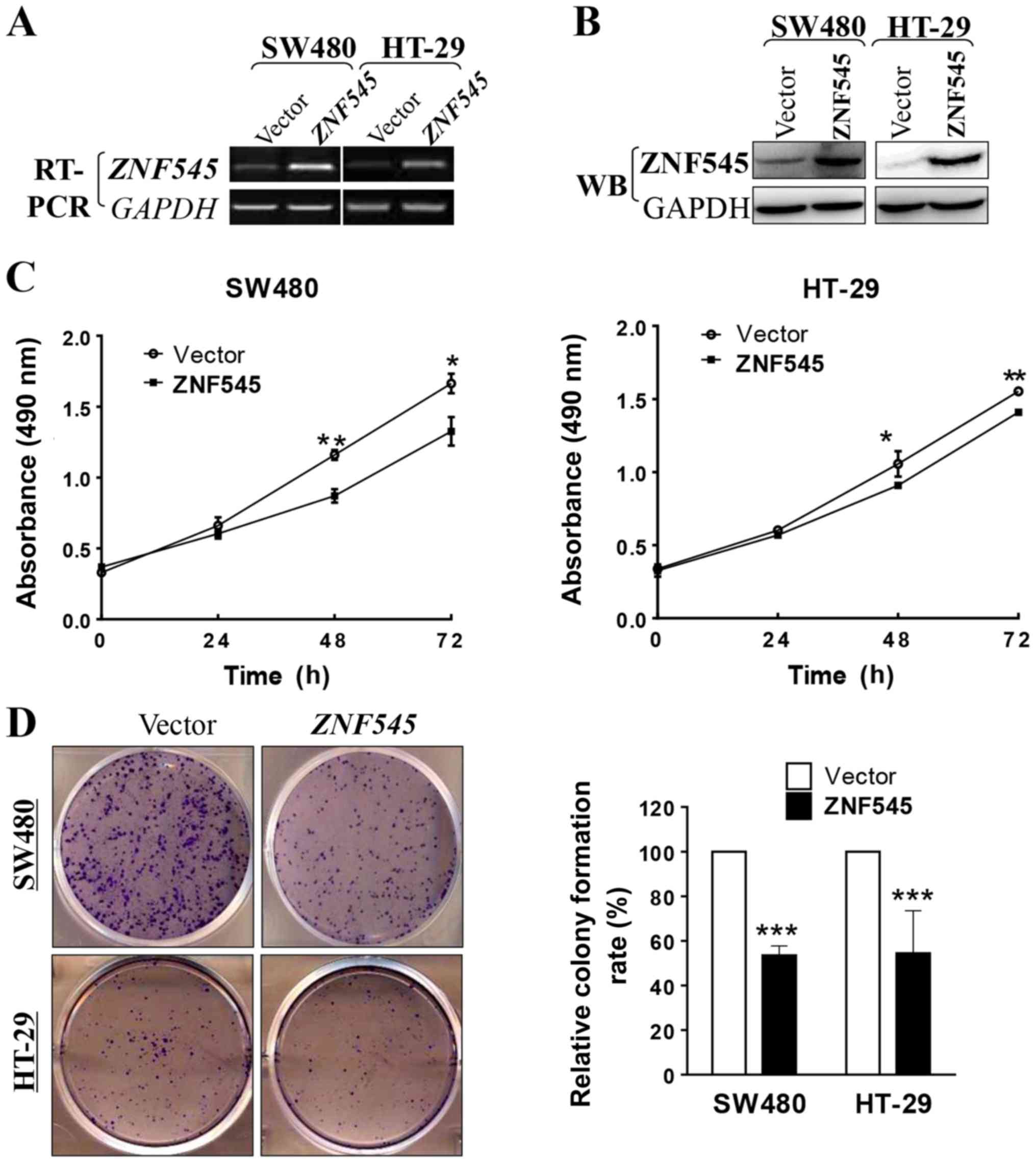
proliferation and clonogenicity of CRC cells in vitro
To determine whether ZNF545 is a functional
tumor suppressor gene in CRC we used the MTS assay and the colony
formation assay to measure the growth-inhibiting effects of
overexpressed ZNF545 in SW480 and HT-29 cells. The
transfection efficiencies of our ZNF545 constructs were
confirmed by RT-PCR and western blotting of tumor cell lines and
stable cell lines that were selected, respectively (Fig. 3A and B). The MTS cell proliferation
assay showed that the cell viability of the
ZNF545-overexpressing cells significantly decreased at 48
and 72 h compared with cells transfected with the empty vector
(p<0.05; Fig. 3C). The colony
formation assay showed that the colony number of cells transfected
with ZNF545 was reduced by ~45% when compared with that
transfected with the empty vector in SW480 and HT-29 cells
(p<0.001; Fig. 3D), indicating
that expression of ZNF545 decreased the cloning efficiency.
Together, these results showed that ZNF545 inhibited cell
proliferation in CRC.
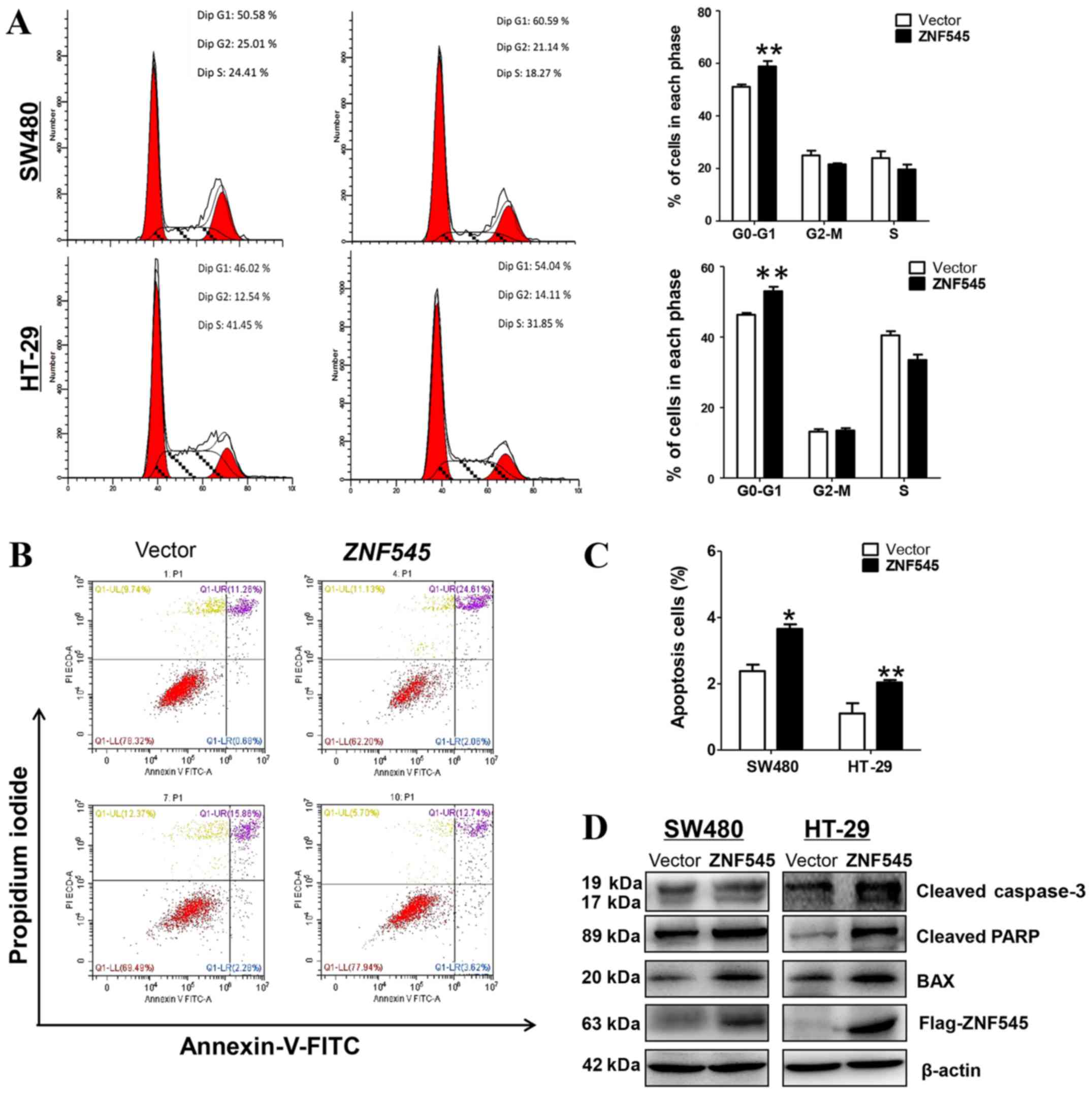
Overexpression of ZNF545 induces cell
cycle arrest and apoptosis of CRC cells
Further studies characterized the effects of ZNF545
on apoptosis and the cell cycle of CRC cells. Fig. 4A shows that the stably-transfected
SW480 and HT-29 cells showed a significantly increased number of
cells accumulating in the G0/G1 phase when compared with control
cells (p<0.01), suggesting that the inhibition of cell
proliferation by ZNF545 was likely mediated by a cell cycle
arrest at the G0/G1 interface. In addition, flow cytometry showed
that the percentage of apoptotic Annexin V-PI-positive cells
increased with the overexpression of ZNF545 (Fig. 4B and C).
We also determined the apoptosis of CRC cell lines
by western blot analyses of Bax, the active form of caspase-3, and
PARP. Fig. 4D shows the increased
expression of Bax, cleaved-caspase-3, and cleaved-PARP in both cell
lines when compared with cells transfected with the control vector.
Together, these results suggested that ZNF545 inhibited cell
proliferation by apoptosis and by mediating the cell cycle arrest
at the G0/G1 interface.
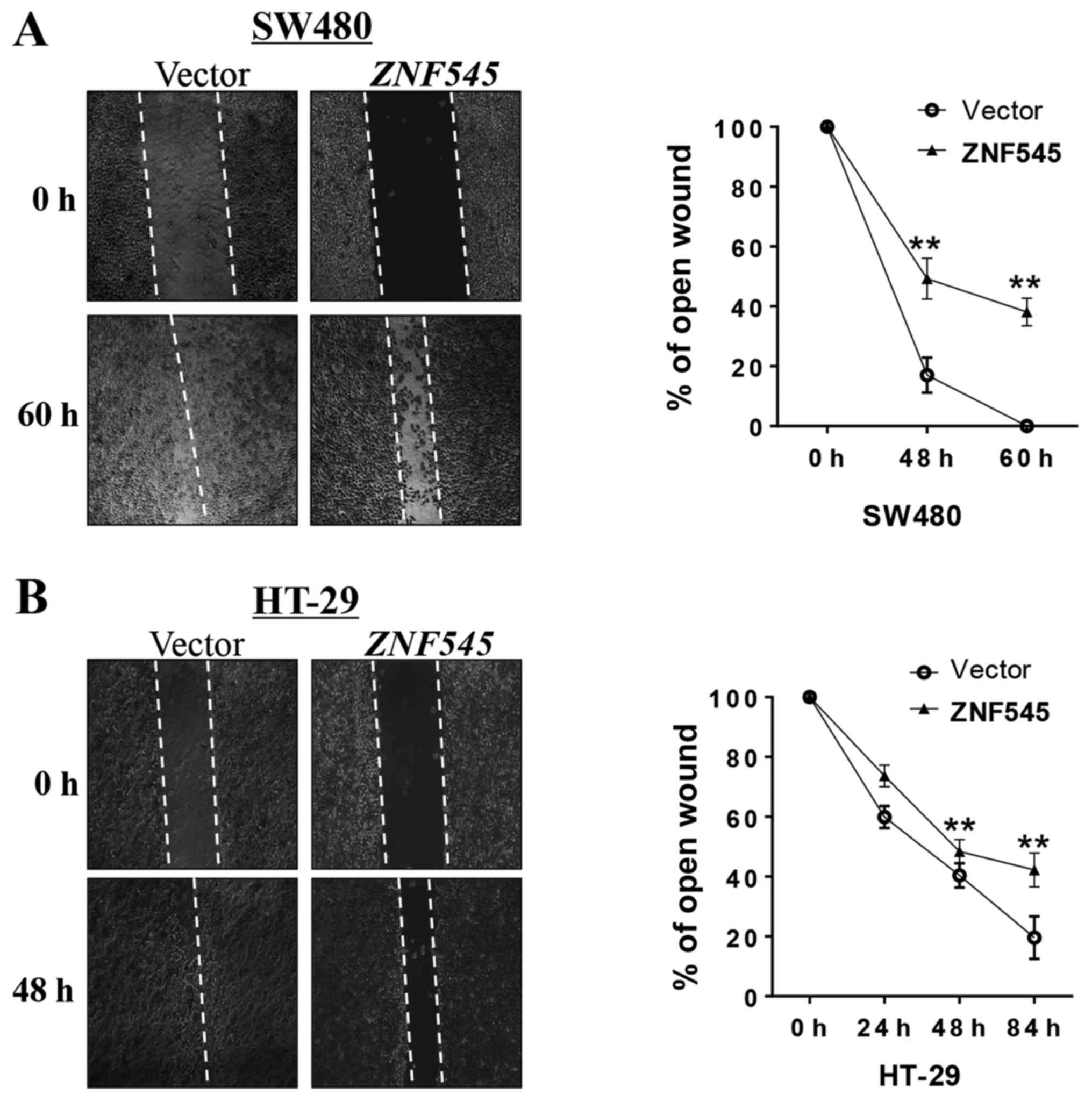
ZNF545 inhibits CRC cell migration
A wound-healing assay was performed to test the
effect of ZNF545 on CRC cell migration. Cells expressing
ZNF545 spread along the wound edges significantly more
slowly than the control cells (Fig.
5), suggesting that ZNF545 suppressed the migration of
SW480 and HT-29 cells.
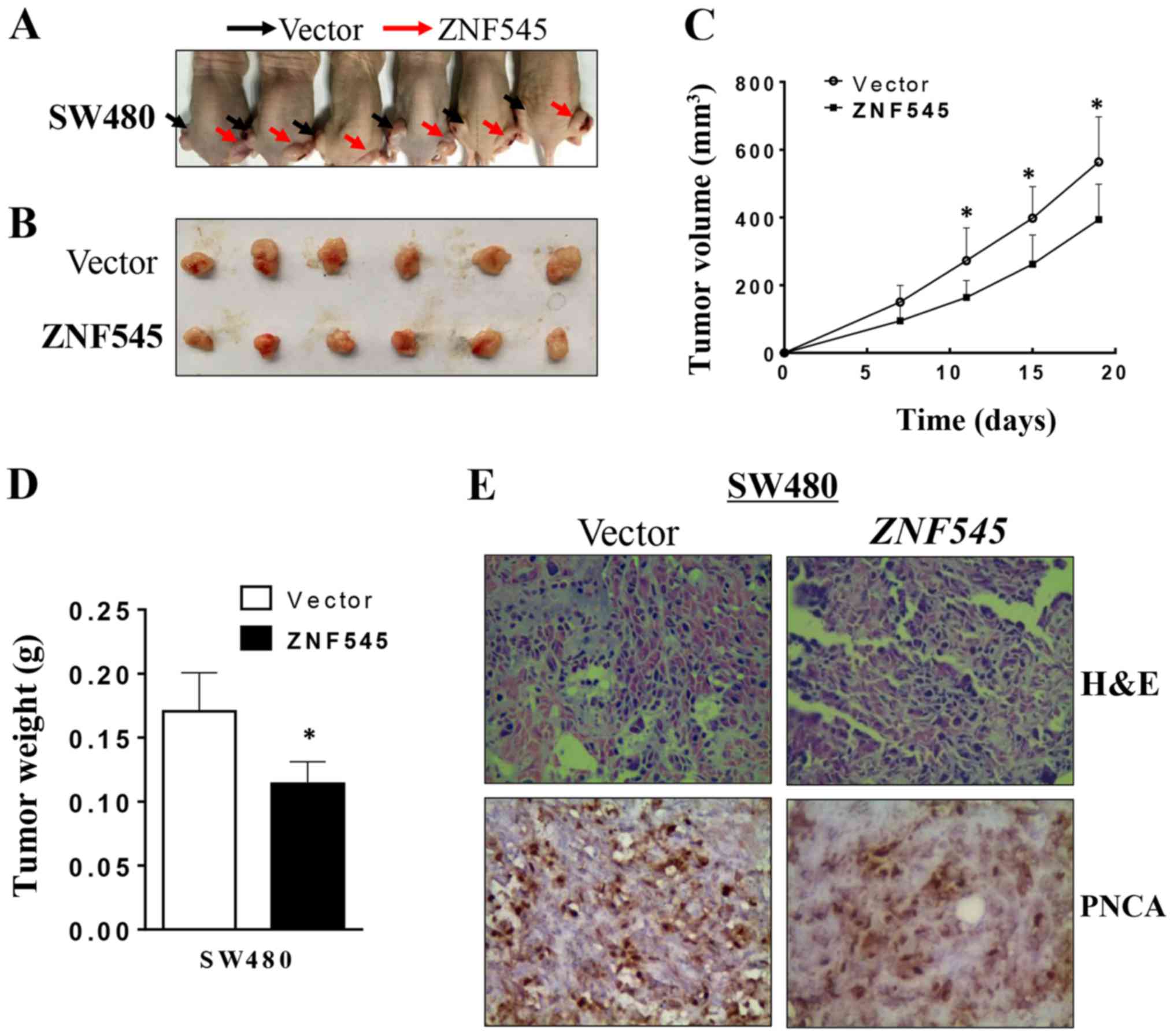
ZNF545 inhibits the growth of tumor
xenografts in nude mice
We also determined if ZNF545 could suppress
the growth of CRC cells in vivo. SW480 cells stably
transfected with ZNF545 or an empty vector formed tumors in
nude mice (Fig. 6A). The
subcutaneous tumor growth curve of SW480 cells stably-transfected
with ZNF545 or with empty vector showed a significant
difference (p<0.05; Fig. 6B),
and the tumor volumes from the ZNF545-treated animals were
significantly smaller than that of the control group (p<0.05;
Fig. 6C). Moreover, the mean
weight of the tumors formed by ZNF545-transfected cells was
significantly lower than that of tumors formed from the empty
vector control cells (p<0.05; Fig.
6D), suggesting that ZNF545 inhibited the growth of CRC
cells in vivo. After using PCNA to stain the tissue sections
from nude mice, immunohistochemical analyses showed that cell
proliferation in the tumors from empty vector-treated cells was
greater than that of tumors from the ZNF545-treated animals
(Fig. 6E).
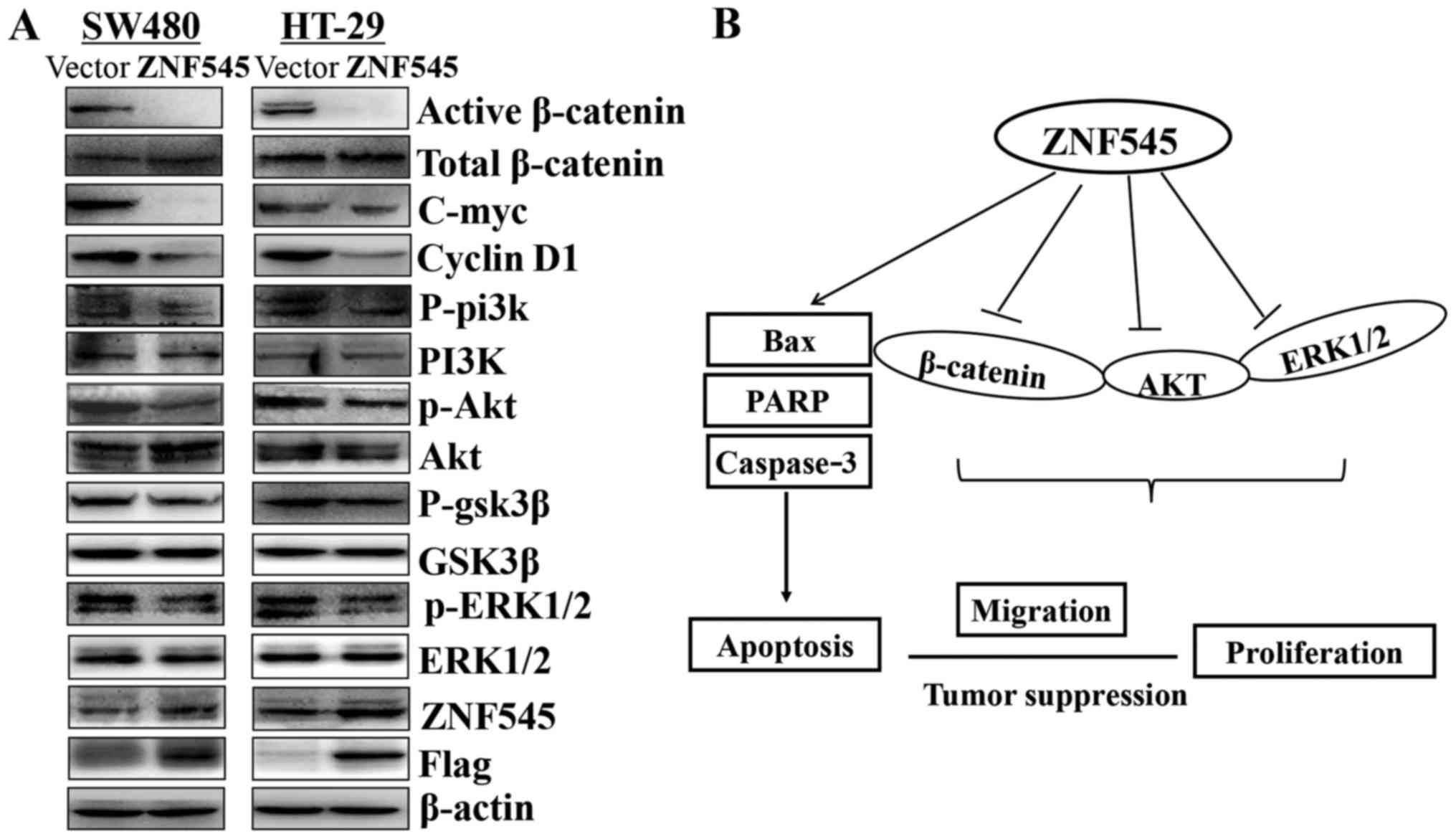
The tumor suppressive property of ZNF545
was mediated by the Wnt/β-catenin, PI3K/AKT, and MAPK/ERK signaling
pathways
Multiple zinc finger family members have been
reported to interact the β-catenin activity, and we investigated
whether ZNF545 functions as a tumor suppressor through
Wnt/β-catenin signaling pathway. The results showed that the
expression of active-catenin was downregulated. Moreover, we
characterized the effects of ZNF545 on the PI3K/Akt and ERK
signaling pathways. The results showed that phospho-AKT and
phospho-ERK1/2 were downregulated, indicating that the two pathways
also participated in the carcinogenic process. GSK3β is a
downstream target gene of the PI3K/AKT pathway and ZNF545
resulted in a profound reduction of GSK3β (Fig. 7A).
Discussion
Increasing evidence has shown that ZNF545
plays an important role in the development of various cancers.
ZNF545 is abnormally expressed, and acts as a suppressor
gene in some tumors. For example, ZNF545 was poorly
expressed in breast cancer (19)
and gastric cancer (23), and
suppressed breast cancer and gastric cancer proliferation.
Furthermore, the Oncomine database showed that ZNF545 was
significantly downregulated in multiple types of CRC tissues when
compared to normal adult colorectal tissues. In the present study,
we found that the expression of ZNF545 was frequently
downregulated in CRC cells, as well as in primary colorectal tumor
tissues when compared with surgical margin tissues, suggesting that
ZNF545 may be a tumor suppressor in CRC.
It has been reported that the decreased expression
of ZNF545 is related to its methylation, and it is generally
accepted that promoter methylation is a major mechanism involved in
gene changes such as gene silencing or gene downregulation that
have been found in human malignant tumors (21). Using MSP and a demethylation
treatment, we found that the reduced expression of ZNF545
was associated with methylation of the promoter, suggesting that
DNA methylation is a regulatory mechanism of ZNF545
inactivation in CRC.
It has been reported that the methylation of
ZNF545 can be used as a prognostic marker for early stage
hepatocellular carcinoma (HCC) after thermal ablation (24), and low expression of ZNF545
was determined positively correlated with the poor prognosis in CRC
patients through analyzing the PrognoScan database. However, there
was an insufficient number of clinical samples of CRC in our study,
so we could not determine the possible relationship between
ZNF545 methylation and the pathological features and
survival prognoses of CRCs. Further analyses of more clinical
samples is therefore necessary to determine if ZNF545 is
indeed a prognostic marker for CRC.
The biological functions of ZNF545 in CRC
were also studied by in vitro and in vivo
overexpression of ZNF545. Previous studies reported that
expression of ZNF545 inhibited clone formation and induced
apoptosis in HCT116 CRC cells (18). The results of the present study
showed that ZNF545 affected apoptosis and G0/G1 cell cycle
arrest to inhibit cell proliferation and clone formation in
vitro. Furthermore, using the wound healing assay, we showed
that overexpression of ZNF545 markedly inhibited cell
migration. The inhibition of tumor growth by overexpression of
ZNF545 was also tested in vivo, by monitoring the
formation of tumor xenografts in nude mice. Flow cytometry and
western blot analyses were used to show the induction of apoptosis
by expression of ZNF545 in CRC cells. Flow cytometry showed
that expression of ZNF545 resulted in early apoptosis in CRC
cells, and western blot analyses showed increased expression of
Bax, cleaved caspase-3, and cleaved PARP. Taken together, the
results suggested that ZNF545 functions as a tumor
suppressor in CRC.
The members of KRAB-ZFPs family have N-terminal KRAB
domain and C-terminal C2H2 domain, and the C2H2 domain can function
as structural motif to bind to the promoter of DNA or RNA. Previous
studies have reported that many ZFPs influence cell biological
function through Wnt/β-catenin pathway, such as ZNF488, ZNF191,
KLF4 and ZIC2. ZNF488 acts as an oncogene via the Wnt/β-catenin
pathway to induce EMT in nasopharyngeal carcinoma (25). ZNF191 can directly bind to the
β-catenin promoter and invoke the expression of β-catenin to
promote cell proliferation in hepatoma cell lines (26). Recent report showed that the KLF4
binds to the β-catenin through its C-terminus which contains three
zinc-finger domains, inhibits Wnt/β-catenin signaling pathway and
suppresses cell proliferation in colorectal cancer (27). The zinc finger domain of ZIC2 is
required for the interaction with TCF4, and ZIC2 can modulate Wnt
pathway in 293T cells (28).
Whether ZNF545 performs a similar mechanism that acts on
Wnt/β-catenin pathway in colorectal cancer remains unclear.
Wnt signaling plays an important role in the
formation and maintenance of human intestinal epithelium (29). The deregulated Wnt signaling is an
early event in colon carcinogenesis (30). Our data showed that re-expression
of ZNF545 inhibited the activated β-catenin and its
downstream target genes in colorectal cancer cell lines.
It has been reported that the PI3K/AKT and MAPK/ERK
pathways can promote the biological function of proliferation, and
migration in multiple cancer cells (31–33).
AKT is a key component in the PI3K pathway, where it suppresses
cell survival and growth (34).
Our results showed that when compared to the empty vector group,
the expression level of phosphorylated AKT and phosphorylated
GSK-3β were decreased in the ZNF545 group. The total amount
of ERK1/2 protein was unchanged in the ZNF545- and
vector-transfected cells, but phosphorylated ERK1/2 levels
decreased in the ZNF545-transfected cells. Thus, ZNF545 may inhibit
the PI3K/Akt/GSK3β and MAPK/ERK pathways to reduce the cell
proliferation and migration ability in colorectal cancer cells.
Identification of the target gene of ZNF545 (RNA sequence or
Chip sequence) will therefore assist in identifying its role in the
progression of CRC.
In conclusion, these results suggested that
ZNF545 functions as a tumor suppressor in CRC, and that its
inactivation results from promoter methylation. ZNF545 may
be an epigenetic biomarker for colorectal tumors. Overall,
ZNF545 can regulate cell proliferation, apoptosis, and
metastasis by affecting the Wnt/β-catenin, PI3K/AKT, and MAPK/ERK
signaling pathways.
Acknowledgments
The authors thank Professor Qian Tao (the Chinese
University of Hong Kong, Hong Kong, China) for generously providing
CRC cell lines, plasmid and primers, technical assistance, and help
with the whole design of the experiment. This study was supported
by National Natural Science Foundation of China (no. 31420103915)
and special research funds from The Chinese University of Hong
Kong. This study was also supported by National Key Clinical
Specialties Construction Program of China (no. [2013] 544).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics. 2012.CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics. 2016.CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Grady WM and Carethers JM: Genomic and
epigenetic instability in colorectal cancer pathogenesis.
Gastroenterology. 135:1079–1099. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jones PA and Baylin SB: The epigenomics of
cancer. Cell. 128:683–692. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dehan P, Kustermans G, Guenin S, Horion J,
Boniver J and Delvenne P: DNA methylation and cancer diagnosis: New
methods and applications. Expert Rev Mol Diagn. 9:651–657. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shivapurkar N and Gazdar AF: DNA
methylation based biomarkers in non-invasive cancer screening. Curr
Mol Med. 10:123–132. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Urrutia R: KRAB-containing zinc-finger
repressor proteins. Genome Biol. 4:2312003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lupo A, Cesaro E, Montano G, Zurlo D, Izzo
P and Costanzo P: KRAB-zinc finger proteins: A repressor family
displaying multiple biological functions. Curr Genomics.
14:268–278. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huntley S, Baggott DM, Hamilton AT,
Tran-Gyamfi M, Yang S, Kim J, Gordon L, Branscomb E and Stubbs L: A
comprehensive catalog of human KRAB-associated zinc finger genes:
Insights into the evolutionary history of a large family of
transcriptional repressors. Genome Res. 16:669–677. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cowger JJ, Zhao Q, Isovic M and Torchia J:
Biochemical characterization of the zinc-finger protein 217
transcriptional repressor complex: Identification of a ZNF217
consensus recognition sequence. Oncogene. 26:3378–3386. 2007.
View Article : Google Scholar
|
|
11
|
Qiang W, Zhao Y, Yang Q, Liu W, Guan H, Lv
S, Ji M, Shi B and Hou P: ZIC1 is a putative tumor suppressor in
thyroid cancer by modulating major signaling pathways and
transcription factor FOXO3a. J Clin Endocrinol Metab.
99:E1163–E1172. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang X, Yuan W, Huang W, Bai Y, Deng Y,
Zhu C, Liang P, Li Y, Du X, Liu M, et al: ZNF569, a novel
KRAB-containing zinc finger protein, suppresses MAPK signaling
pathway. Biochem Biophys Res Commun. 346:621–628. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang GJ, He Z and Ma L: ZFP932 suppresses
cellular Hedgehog response and Patched1 transcription. Vitam Horm.
88:309–332. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li A, Jiao Y, Yong KJ, Wang F, Gao C, Yan
B, Srivastava S, Lim GS, Tang P, Yang H, et al: SALL4 is a new
target in endometrial cancer. Oncogene. 34:63–72. 2015. View Article : Google Scholar
|
|
15
|
Tune CE, Pilon M, Saiki Y and Dosch HM:
Sustained expression of the novel EBV-induced zinc finger gene,
ZNFEB, is critical for the transition of B lymphocyte activation to
oncogenic growth transformation. J Immunol. 168:680–688. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nakaya T, Ogawa S, Manabe I, Tanaka M,
Sanada M, Sato T, Taketo MM, Nakao K, Clevers H, Fukayama M, et al:
KLF5 regulates the integrity and oncogenicity of intestinal stem
cells. Cancer Res. 74:2882–2891. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang S, Cheng Y, Du W, Lu L, Zhou L, Wang
H, Kang W, Li X, Tao Q, Sung JJ, et al: Zinc-finger protein 545 is
a novel tumour suppressor that acts by inhibiting ribosomal RNA
transcription in gastric cancer. Gut. 62:833–841. 2013. View Article : Google Scholar
|
|
18
|
Cheng Y, Liang P, Geng H, Wang Z, Li L,
Cheng SH, Ying J, Su X, Ng KM, Ng MH, et al: A novel 19q13
nucleolar zinc finger protein suppresses tumor cell growth through
inhibiting ribosome biogenesis and inducing apoptosis but is
frequently silenced in multiple carcinomas. Mol Cancer Res.
10:925–936. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xiao Y, Xiang T, Luo X, Li C, Li Q, Peng
W, Li L, Li S, Wang Z, Tang L, et al: Zinc-finger protein 545
inhibits cell proliferation as a tumor suppressor through inducing
apoptosis and is disrupted by promoter methylation in breast
cancer. PLoS One. 9:e1109902014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Y, Li J, Cui Y, Li T, Ng KM, Geng H,
Li H, Shu XS, Li H, Liu W, et al: CMTM3, located at the critical
tumor suppressor locus 16q22.1, is silenced by CpG methylation in
carcinomas and inhibits tumor cell growth through inducing
apoptosis. Cancer Res. 69:5194–5201. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jones PA and Baylin SB: The fundamental
role of epigenetic events in cancer. Nat Rev Genet. 3:415–428.
2002.PubMed/NCBI
|
|
22
|
Herman JG and Baylin SB: Gene silencing in
cancer in association with promoter hypermethylation. N Engl J Med.
349:2042–2054. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Deng J, Liang H, Ying G, Dong Q, Zhang R,
Yu J, Fan D and Hao X: Poor survival is associated with the
methylated degree of zinc-finger protein 545 (ZNF545) DNA promoter
in gastric cancer. Oncotarget. 6:4482–4495. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yu J, Li X, Tao Q, Yu XL, Cheng ZG, Han
ZY, Guo M and Liang P: Hypermethylation of ZNF545 is associated
with poor prognosis in patients with early-stage hepatocellular
carcinoma after thermal ablation. Gut. 64:1836–1837. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zong D, Yin L, Zhong Q, Guo WJ, Xu JH,
Jiang N, Lin ZR, Li MZ, Han P, Xu L, et al: ZNF488 enhances the
invasion and tumorigenesis in nasopharyngeal carcinoma via the Wnt
signaling pathway involving epithelial mesenchymal transition.
Cancer Res Treat. 48:334–344. 2016. View Article : Google Scholar :
|
|
26
|
Liu G, Jiang S, Wang C, Jiang W, Liu Z,
Liu C, Saiyin H, Yang X, Shen S, Jiang D, et al: Zinc finger
transcription factor 191, directly binding to β-catenin promoter,
promotes cell proliferation of hepatocellular carcinoma.
Hepatology. 55:1830–1839. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang W, Chen X, Kato Y, Evans PM, Yuan S,
Yang J, Rychahou PG, Yang VW, He X, Evers BM, et al: Novel cross
talk of Kruppel-like factor 4 and beta-catenin regulates normal
intestinal homeostasis and tumor repression. Mol Cell Biol.
26:2055–2064. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pourebrahim R, Houtmeyers R, Ghogomu S,
Janssens S, Thelie A, Tran HT, Langenberg T, Vleminckx K,
Bellefroid E, Cassiman JJ, et al: Transcription factor Zic2
inhibits Wnt/β-catenin protein signaling. J Biol Chem.
286:37732–37740. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Doerks T, Copley RR, Schultz J, Ponting CP
and Bork P: Systematic identification of novel protein domain
families associated with nuclear functions. Genome Res. 12:47–56.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Morin PJ, Sparks AB, Korinek V, Barker N,
Clevers H, Vogelstein B and Kinzler KW: Activation of
beta-catenin-Tcf signaling in colon cancer by mutations in
beta-catenin or APC. Science. 275:1787–1790. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Datta SR, Brunet A and Greenberg ME:
Cellular survival: A play in three Akts. Genes Dev. 13:2905–2927.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qing H, Gong W, Che Y, Wang X, Peng L,
Liang Y, Wang W, Deng Q, Zhang H and Jiang B: PAK1-dependent MAPK
pathway activation is required for colorectal cancer cell
proliferation. Tumour Biol. 33:985–994. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Levidou G, Saetta AA, Gigelou F, Karlou M,
Papanastasiou P, Stamatelli A, Kavantzas N, Michalopoulos NV,
Agrogiannis G, Patsouris E, et al: ERK/pERK expression and B-raf
mutations in colon adenocarcinomas: Correlation with
clinicopathological characteristics. World J Surg Oncol. 10:472012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Khor TO, Gul YA, Ithnin H and Seow HF:
Positive correlation between overexpression of phospho-BAD with
phosphorylated Akt at serine 473 but not threonine 308 in
colorectal carcinoma. Cancer Lett. 210:139–150. 2004. View Article : Google Scholar : PubMed/NCBI
|