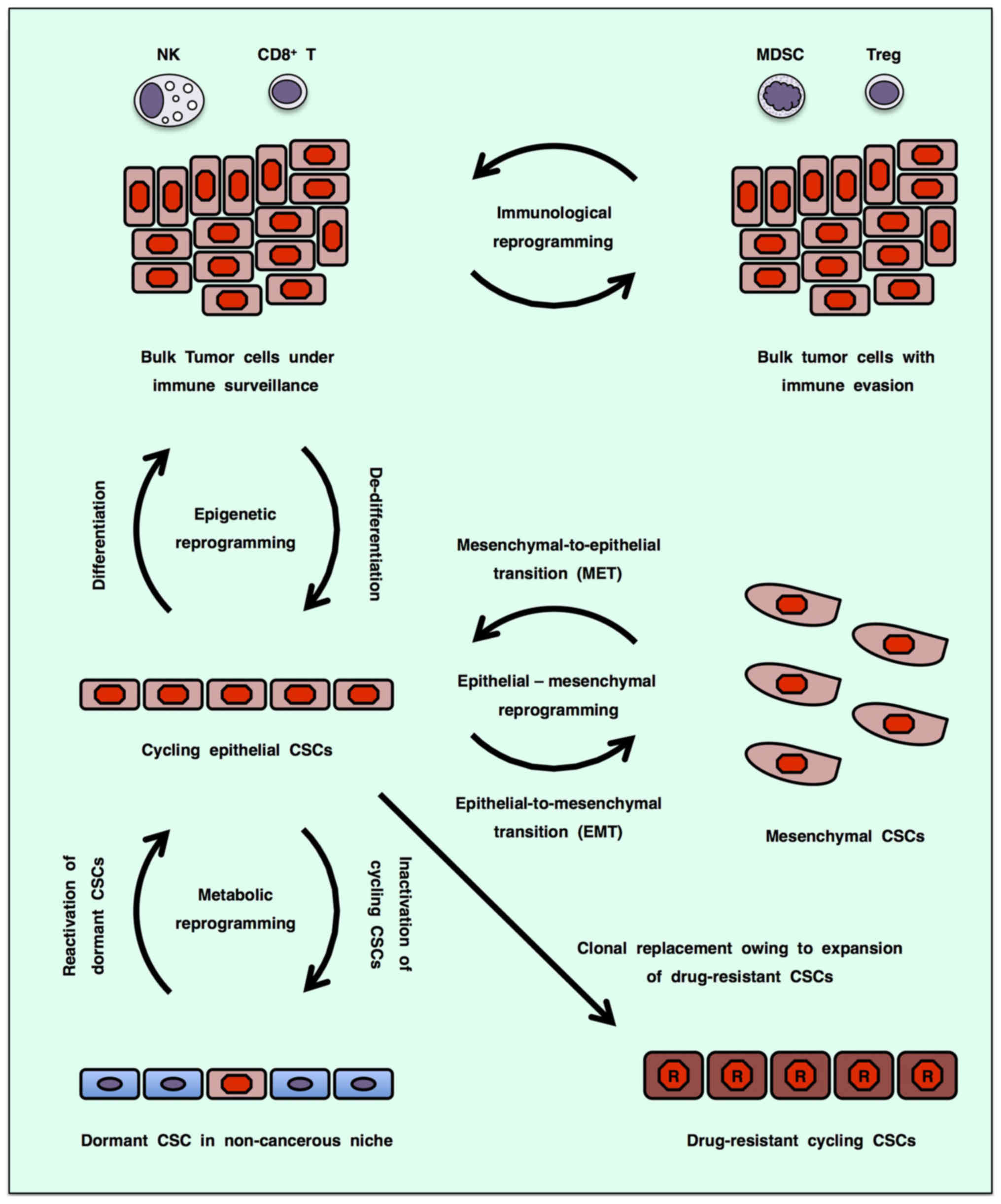
Cancer stem cells (CSCs), which show the potential
for self-renewal and differentiation, have been identified in a
variety of human cancers based on their tumor initiating potential
in vivo (1–3). Clonal expansion of a minor CSC
population with a drug-resistant mutation causes early recurrence,
whereas reactivation of dormant CSCs into cycling CSCs owing to
tumor plasticity leads to late relapse (4–6).
CSCs or bulk tumor cells undergo epigenetic reprogramming (7), epithelial-mesenchymal reprogramming
[epithelial-to-mesenchymal transition (EMT) and
mesenchymal-to-epithelial transition (MET)] (8,9),
immunological reprogramming (immunoediting) (10,11)
and metabolic reprogramming (12)
to adapt to the tumor microenvironment, which is collectively
defined here as 'omics reprogrammming' (Fig. 1). Since cycling CSCs that depend on
aerobic glycolysis converge into quiescent mesenchymal CSCs through
omics reprogramming to survive therapeutic insult for later
recurrence, CSC targeting is necessary to avoid relapse after
cancer therapy and improve the cost-effectiveness ratio of cancer
precision medicine.
CD44, CD133 (PROM1), EPCAM and LGR5 (GPR49) are
representative cell-surface markers of CSCs (2,13–16).
LGR5, encoding an R-spondin (RSPO) receptor, is a target gene of
the canonical WNT/β-catenin signaling cascade in quiescent as well
as cycling stem cells, whereas CD44 and CD133 are further
upregulated by WNT and RSPO signals in LGR5+ cycling
stem/progenitor cells (17–19).
EPCAM can potentiate the canonical WNT/β-catenin signaling cascade
through intra-membrane proteolysis and subsequent nuclear
translocation of its intracellular C-terminal domain (20). WNT signaling cascades cross-talk
with the FGF, Notch, Hedgehog and TGFβ/BMP signaling cascades to
constitute the stem cell signaling network, which regulates
expression of functional CSC markers (21–24).
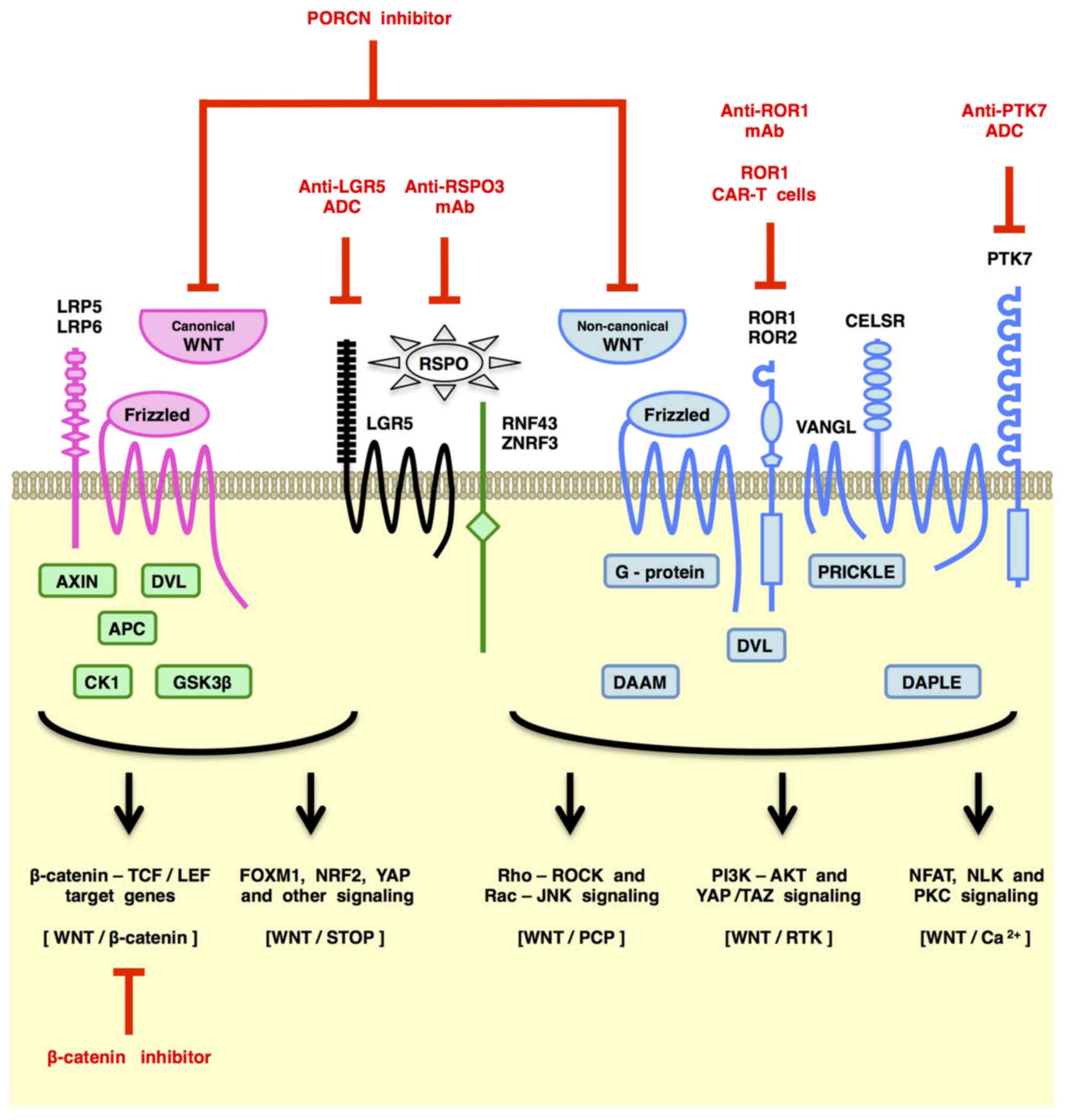
The WNT family proteins transduce signals through
the Frizzled (FZD) and LRP5/6 receptors to the WNT/β-catenin and
WNT/STOP (stabilization of proteins) signaling cascades (also known
as the canonical WNT signaling cascades) and through the FZD and/or
ROR1/ROR2/RYK receptors to the WNT/PCP (planar cell polarity),
WNT/RTK (receptor tyrosine kinase) and WNT/Ca2+
signaling cascades (also known as the non-canonical WNT signaling
cascades) (21,25–29).
The canonical WNT/β-catenin signaling cascade is involved in
self-renewal of stem cells and proliferation or differentiation of
progenitor cells (30–33), whereas non-canonical WNT signaling
cascades are involved in maintenance of stem cells, directional
cell movement or inhibition of the canonical WNT signaling cascade
(34–37). Both canonical and non-canonical WNT
signaling cascades play key roles in the development and evolution
of CSCs.
By contrast, tumors consist of heterogeneous
populations of cancer cells and non-cancerous stromal/immune cells
(38,39). Intra-tumor heterogeneity of cancer
cells is caused by the evolution of CSCs based on epigenetic and
genetic alterations (40–42), as well as the differentiation of
CSCs into bulk tumor cells (1–3),
niche-like cancer supporting cells (43), endothelial-like cancer cells
(44) and fibroblast-like cancer
cells (45). On the other hand,
intra-tumor heterogeneity of non-cancerous stromal/immune cells is
orchestrated by and reciprocally orchestrates CSCs and their
descendants (39,45–47).
Interaction and co-evolution of CSCs and niche cells are driving
forces of cancer progression. Herein, canonical and non-canonical
WNT signaling in CSCs will be described, with a focus on the
heterogeneity of cancer and stromal/immune cells in the tumor
microenvironment; then, anti-CSC mono- and combination therapies
using WNT signaling-targeted therapeutics will be reviewed with
emphases on omics reprogramming and tumor plasticity.
Canonical WNT signaling through the FZD-LRP5/6
receptor complex leads to de-repression of β-catenin as well as
STOP-target proteins, such as ATOH1, CCND1 (Cyclin D1), FOXM1, MYC
(c-MYC), NRF2 (NFE2L2), PLK1, SMAD1/3/4, SNAI1 (Snail) and YAP/TAZ,
from proteasomal degradation induced by GSK-3β-dependent
phosphorylation and subsequent ubiquitylation (27–29,48)
(Fig. 2). β-catenin stabilization
and subsequent nuclear translocation leads to transcriptional
activation of β-catenin-TCF/LEF target genes, such as ATOH1,
CCND1, CD44, FGF20, JAG1, LGR5, MYC and SNAI1, although
transcriptional outputs of the WNT/β-catenin signaling cascade are
determined in a cellular context-dependent manner (e.g., epigenetic
status of target genes and activities of other transcriptional
regulators). ATOH1, CCND1, MYC and SNAI1 are upregulated
transcriptionally and post-translationally by the β-catenin and
STOP signaling cascades, respectively. Canonical WNT signals
control cell fate and function through transcriptional and
post-translational regulation of the omics network.
Canonical WNT signaling in CSCs is activated by
WNT2B, WNT3 and other canonical WNT ligands derived from cancerous
supporting cells or non-cancerous stromal cells (49–52),
as well as genetic alterations in the canonical WNT/β-catenin
signaling components, such as EIF3E-RSPO2 fusions, PTPRK-RSPO3
fusions, gain-of-function mutations in the CTNNB1
(β-catenin) gene and loss-of-function mutations in the APC,
AXIN1, AXIN2, RNF43 and ZNRF3 genes (29,53–55).
Canonical WNT signals increase the LGR5 receptor level on CSCs for
the maintenance of the canonical WNT responsive state but also
upregulate AXIN2, DKK1, NOTUM, RNF43 and ZNRF3 for negative
feedback regulation (18–21,29).
Loss-of-function mutations in the APC, AXIN2, RNF43 and
ZNRF3 genes release CSCs from the constraints of the
negative feedback regulation.
Canonical WNT signals can directly promote CSC
proliferation through upregulation of CCND1, FOXM1, MYC and YAP/TAZ
as described above. By contrast, canonical WNT signaling in CSCs
induces expression and secretion of growth factors, such as FGFs,
KIT ligand (KITLG or SCF) and VEGF (VEGFA), to fine-tune the tumor
microenvironment (18,21,29).
For example, MET (HGF receptor) is upregulated in human basal-like
breast cancers with TP53 mutations as well as mouse basal-like
breast tumors with compound gain-of-function Ctnnb1 mutation
and homozygous Tp53 deletion (56), and combined activation of the
canonical WNT/β-catenin and HGF/MET signaling cascades induces SHH
upregulation in mouse mammary CSCs and subsequent activation of
cancer-associate fibroblasts for the synergistic proliferation of
CSCs and cancer-associate fibroblasts (57).
Together, these findings indicate that canonical WNT
signaling is involved in the maintenance and expansion of CSCs
through direct effects on CSCs themselves and indirect effects via
CSC-stromal/immune interactions.
Non-canonical WNT signaling through FZD receptors
and/or ROR1/ROR2/RYK co-receptors activates the PCP, RTK or
Ca2+ signaling cascades (Fig. 2).
Non-canonical WNT/PCP signaling through FZD
receptors and Dishevelled (DVL) adaptor proteins regulates the
coordinated cellular orientation within an epithelial plane,
collective cell movements during gastrulation and neurulation
stages of embryogenesis and directional cell movement during
invasion and metastasis of cancer cells (58–62).
WNT/PCP signals are converted to actin cytoskeletal dynamics via
the small G-proteins RAC and RHO (Fig.
2), and then, RAC and RHO activate JNK-dependent transcription
and YAP/TAZ-dependent transcription, respectively (63–66).
WNT/PCP signaling regulates actin cytoskeletal dynamics,
directional cell movement and JNK- or YAP/TAZ-dependent
transcription.
Non-canonical WNT signaling through RTKs, such as
ROR1, ROR2 and RYK, activates the PI3K-AKT signaling cascade
(29,67–69).
ROR1 and ROR2, with the extracellular WNT-binding FZD-like domain,
are homologous to MUSK, NTRK1, NTRK2, NTRK3, DDR1 and DDR2 in their
cytoplasmic tyrosine kinase domain, whereas RYK with an
extracellular WNT-binding WIF domain is homologous to AXL, EGFR,
ERBB2, ERBB3, ERBB4, MET, MERTK, MST1R and TYRO3 in its cytoplasmic
tyrosine kinase domain (39,70–73).
ROR1 and ROR2 are atypical RTKs that are defective in intrinsic
tyrosine kinase activity for auto-phosphorylation; however, ROR1
and ROR2 can be tyrosine phosphorylated by other tyrosine kinases,
such as EGFR, ERBB3, MET and SRC, to activate the PI3K-AKT and YAP
signaling cascades (29,74–78)
(Fig. 2). WNT/RTK signaling is
involved in therapeutic resistance and recurrence of human cancers
in part through PI3K-AKT signaling activation.
Non-canonical WNT signaling in CSCs is activated by
WNT5A, WNT11 and other non-canonical WNT ligands (58) that are secreted from cancer cells
(86,87) or stromal/immune cells (88,89),
as well as genetic alterations that trans-activate
non-canonical WNT signaling cascades, such as E2A-PBX1
fusion and MET amplification (74–76).
Non-canonical WNT signaling through FZD7 activates the PI3K-AKT
signaling cascade as a result of Daple (CCDC88C)-mediated
dissociation of Gβγ from Gαi (90), whereas non-canonical WNT signaling
through ROR1 activates PI3K-AKT signaling cascade owing to ROR1
trans-phosphorylation by other tyrosine kinases, such as MET
and SRC (67,75). ROR1 is involved in HER3-Y1307
trans-phosphorylation and subsequent NSUN6-dependent
MST1-K59 methylation, which induces YAP/TAZ-dependent
transcriptional activation through LATS1/LATS2 inhibition (78). WNT/PCP signaling can also induce
Rho-mediated LATS1/LATS2 inhibition for transcriptional activation
of YAP/TAZ-target genes (91,92),
whereas non-canonical WNT signaling through FZD10 induces YAP/TAZ
activation through Gα13 (93).
Non-canonical WNT signaling promotes survival and therapeutic
resistance of CSCs through PI3K-AKT signaling activation and
YAP/TAZ-mediated transcriptional activation.
By contrast, invasion and metastasis are driven by
canonical WNT signaling cascades and non-canonical WNT signaling
cascades. For example, canonical WNT/β-catenin and WNT/STOP
signaling cascades synergistically upregulate SNAI1 to repress
epithelial genes, such as CDH1 (E-cadherin), for the
initiation of EMT of CSCs, and non-canonical WNT signals promote
invasion, survival and metastasis of CSCs or circulating tumor
cells (28,29,35,62,87).
Together, these findings clearly indicate that canonical
WNT/β-catenin signaling as well as other WNT signaling cascades are
critically involved in the malignant features of CSCs.
WNT signaling cascades are hot and cutting-edge
topics in the field of translational oncology and medicinal
chemistry (29,94–96).
Therapeutics directly targeting WNT signaling cascades are
classified into i) ligand/receptor-targeted drugs binding to
ligands or transmembrane proteins involved in WNT signaling, ii)
porcupine (PORCN) inhibitors abrogating WNT secretion and
FZD-dependent signaling, iii) tankyrase (TNKS) inhibitors
repressing WNT/β-catenin and WNT-independent signaling cascades and
iv) β-catenin inhibitors blocking TCF/LEF-dependent transcription
(Table I).
Human/humanized monoclonal antibody (mAb) drugs,
such as anti-FZD1/2/5/7/8 mAb (vantictumab/OMP-18R5) (97), anti-FZD5 mAb (IgG-2919) (52), anti-FZD10 antibody-drug conjugate
(ADC) (OTSA101-DTPA-90Y) (98), anti-LGR5 ADC (mAb-mc-vc-PAB-MMAE)
(99), anti-PTK7 ADC (PF-06647020)
(100), anti-ROR1 mAb
(cirmtuzumab/UC-961) (101) and
anti-RSPO3 mAb (rosmantuzumab/OMP-131R10) (102) have been developed as
large-molecule cancer therapeutics. ROR1 CAR-T cells (103) and WNT-trapping FZD8-Fc chimeric
protein (ipafricept/OMP-54F28) (104) are also classified as WNT
ligand/receptor-targeted drugs. Among this class of therapeutics,
cirmtuzumab, ipafricept, PF-06647020, rosmantuzumab and
vantictumab, which showed anti-CSC effects in preclinical model
experiments, are in clinical trials to treat cancer patients
(Table I).
PORCN inhibitors restrain PORCN-dependent
palmitoleoylation of WNT family ligands in the endoplasmic
reticulum, which obstructs WNT signaling through blockade of WNT
secretion as well as palmitoleoylated WNT-mediated oligomerization
of FZD receptors (105–108). ETC-159 (109), IWP-2 (110), WNT-C59 (111) and WNT974 (LGK974) (112) are small-molecule PORCN
inhibitors. A preclinical study of IWP-2 on organoids derived from
colorectal cancer patients revealed that PORNC inhibitors are
applicable for the treatment of cancers with RNF43 mutations
but not APC mutations (52). By
contrast, preclinical studies of WNT974 indicated that PORNC
inhibitors repress the survival and tumor initiating potential of
CSCs (43,112). ETC-159 and WNT974 are in clinical
trials for the treatment of cancer patients (Table I).
TNKS inhibitors repress TNKS-dependent
poly-ADP-ribosylation and subsequent degradation of negative
regulators of oncogenic signaling cascades, such as AXIN family
proteins, AMOT family proteins, PTEN and TERF1 (TRF1), which
results in inhibition of WNT/β-catenin signaling, repression of
YAP-dependent transcription, suppression of PI3K signaling and
telomere shortening, respectively (113–116). AZ1366 (117), G007-LK (118), JW55 (119), NVP-TNKS656 (120) and XAV939 (121) are representative TNKS inhibitors
that abrogate WNT/β-catenin signaling and tumorigenesis in
preclinical mouse model experiments. TNKS inhibitors show
synergistic antitumor effects with other therapeutics, such as an
AKT inhibitor (API2), EGFR inhibitors (gefitinib and erlotinib), a
MEK inhibitor (AZD6244), a PI3K inhibitor (BKM120) and irinotecan
(117,118,120,122–124). TNKS inhibitors are promising
candidates for CSC-targeted therapeutics; however, because of
diverse on-target effects, TNKS inhibitors stalled in their
preclinical stage.
β-catenin inhibitors block TCF/LEF-dependent
transcription through inhibition of protein-protein interactions
(PPI) between β-catenin and other transcriptional regulators
(29,125), promotion of β-catenin degradation
(126) or inhibition of β-catenin
kinases, such as TNIK (127–129). BC2059 (130), CGP049090 (131), CWP232228 (132), ICG-001 (133), LF3 (134), PKF115–584 (135), PRI-724 (136) and SAH-BCL9 (137) are small-molecule β-catenin PPI
inhibitors. MSAB is a small-molecule compound that binds to
β-catenin and promotes proteasomal degradation of β-catenin
(126). KY-05009 (128), mebendazole (129) and PF-794 (138) are TNIK inhibitors that repress
phosphorylation of TNIK substrates, such as TCF4, FMNL2, PRICKLE1,
SMAD1 and SMAD2, which leads to inhibition of
β-catenin-TCF/LEF-dependent transcription and a variety of cellular
processes. Among the β-catenin inhibitors mentioned above, BC2059,
CWP232228 and ICG-001 repress the expansion of CSCs. The β-catenin
inhibitors PRI-724 and mebendazole are in phase I/II clinical
trials for cancer patients (Table
I), whereas other β-catenin inhibitors are still in the
preclinical stage of drug development. β-catenin inhibitors are
challenging therapeutics for cancer patients.
Tyrosine kinase inhibitors are rational anticancer
therapeutics because tyrosine kinases with intrinsic enzyme
activities are aberrantly activated in cancer cells owing to
genetic alterations. Tyrosine kinase inhibitors have contributed to
the improved prognosis of cancer patients and are essential for
genome-based precision medicine; however, unavoidable drug
resistance or recurrence is a serious issue for cancer patients and
health care systems (4).
Activated tyrosine kinases, such as BCR-ABL fusion
kinase, EGFR-T790M mutant, FLT3 internal tandem duplication
(FLT3-ITD) mutant, KIT-D814V mutant and RET, promote β-catenin
phosphorylation at Y654 to release E-cadherin-bound β-catenin from
the adherens junction for its stabilization and subsequent nuclear
translocation (139–143). By contrast, canonical WNT signals
inhibit β-catenin phosphorylation at S33, S37, T41 and S45 to
release β-catenin from proteasomal degradation for its
stabilization and nuclear translocation (21,25,26,29).
Since canonical WNT signals and oncogenic tyrosine kinases converge
to β-catenin stabilization for the maintenance and expansion of
CSCs, canonical WNT signaling inhibitors can block CSC evasion of
tyrosine kinase inhibitors. For example, the porcupine inhibitor
WNT974 significantly reduced residual stem/progenitor cells of
chronic myeloid leukemia (CML) after treatment with the BCR-ABL
inhibitor nilotinib via blockade of WNT ligand secretion into the
bone marrow microenvironment (112); the β-catenin inhibitors ICG-001
and PRI-724 induced synergistic effects with the BCR-ABL inhibitors
imatinib and nilotinib, respectively, on CML stem/progenitor cells
(136,144); and the TNKS inhibitor AZ1366 and
EGFR inhibitor gefitinib showed synergistic effects on lung cancer
cells in vivo (124).
These preclinical studies indicate that combination therapies using
WNT signaling-targeted therapeutics and tyrosine kinase inhibitors
might be applicable for treatment of a subset of patients with
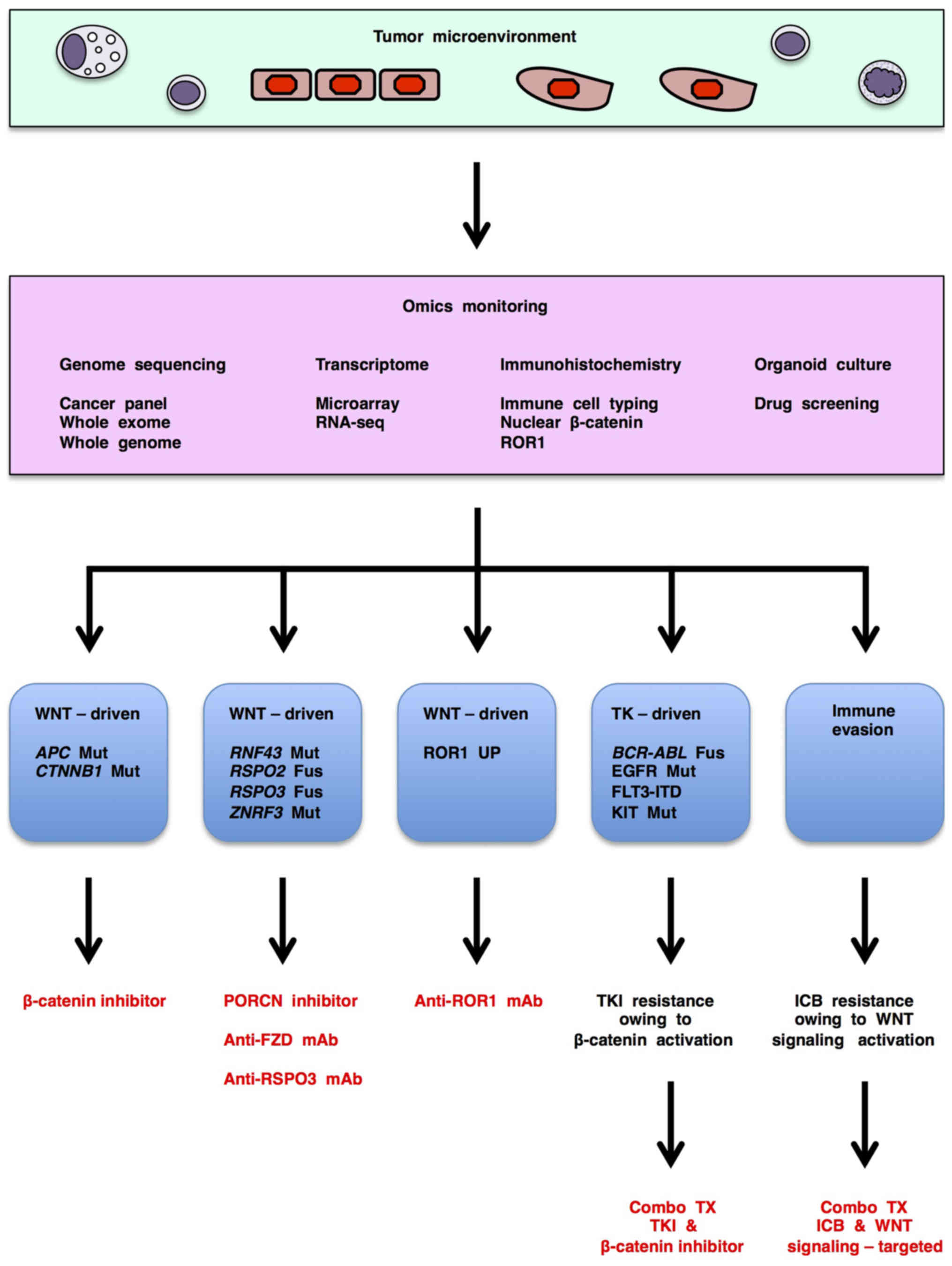
tyrosine kinase-driven cancers (Fig.
3).
Immune checkpoint blockers that abrogate
interactions of ligands and inhibitory receptors on CD8+
T cells are promising antitumor drugs in the clinic or clinical
trials (145–151). PD-L1 (CD274) is a representative
ligand for inhibitory immune signaling, whereas PD-1 (PDCD1) and
CTLA4 are representative receptors for inhibitory immune signaling.
Anti-PD-L1 mAbs (atezolizumab, avelumab and durvalumab), anti-PD-1
mAbs (nivolumab and pembrolizumab) and an anti-CTLA4 mAb
(ipilimumab) are approved for the treatment of patients with
melanoma or other types of solid tumors. Immune checkpoint blockers
result in significant therapeutic effects in a subset of patients;
however, the lack of benefits in other patients owing to primary or
acquired resistance to immune checkpoint blockers has resulted in a
cost-effectiveness issue (152–156).
Canonical WNT signaling activation in melanoma
induces immune evasion through CCL4 repression and immunological
reprogramming into non-T cell-infiltrated melanoma (11). Since melanoma-derived WNT5A
promotes β-catenin signaling activation and subsequent IDO
upregulation in dendritic cells to induce immune evasion through
accumulation of regulatory T (Treg) cells, combination
immunotherapy using the porcupine inhibitor WNT-C59 and anti-CTLA4
mAb showed synergistic anti-melanoma effects in vivo
(157). By contrast, WNT5A
and ROR2 are relatively frequently upregulated in
pretreatment tumors of melanoma patients that do not respond to
PD-1 immune checkpoint blockade (158), which suggests involvement of
non-canonical WNT signaling in resistance to immune checkpoint
blockers. Since DKK1-dependent canonical WNT signaling inhibition
or putative reciprocal non-canonical WNT signaling activation in
tumor microenvironment induces immune evasion through accumulation
of myeloid-derived suppressor cells (MDSCs) and depletion of T
cells (159), combination therapy
using anti-DKK1 mAb (BHQ880 or DKN-01) (160,161) and immune checkpoint blockers
might show synergistic antitumor effects in vivo. WNT
signaling-targeted therapeutics might be applicable for combination
immunotherapy for cancer patients (Fig. 3); however, context-dependent
effects of WNT signaling on immunity (4) should be kept in mind.
WNT-related human cancers are classified into three
major subtypes based on signaling aberrations associated with
therapeutic choices (Fig. 3):
APC/CTNNB1-altered cancers with WNT/β-catenin signaling activation
that can be treated with β-catenin inhibitors;
RNF43/ZNRF3/RSPO2/RSPO3-altered cancers with WNT/β-catenin and
other WNT signaling activation that can be treated with PORCN
inhibitors, anti-FZD mAb or anti-RSPO3 mAb; and ROR1-upregulated
cancers with WNT/PCP and WNT/RTK signaling activation that can be
treated with anti-ROR1 mAb, anti-ROR1 × CD3 bispecific antibody and
ROR1 chimeric antigen receptor-modified T (CAR-T) cells (29). Genome sequencing, transcriptomic
and/or immunohistochemical tests are necessary for the detection
and subtyping of WNT signaling-driven cancers and subsequent
determination of appropriate WNT signaling-targeted therapeutics
(Fig. 3).
WNT signaling-targeted therapeutics are also
applicable for combination therapies with tyrosine kinase
inhibitors or immune checkpoint blockers as mentioned above
(Fig. 3). Since resistance to
tyrosine kinase inhibitors occur owing to multiple mechanisms, such
as acquired drug-resistant mutations in targeted tyrosine kinases,
EMT, activation of other tyrosine kinase signaling cascades to
bypass targeted tyrosine kinases (4,162)
and activation of WNT/β-catenin signaling cascade (Fig. 3), genomic, transcriptomic and/or
immunohistochemical monitoring during tyrosine kinase inhibitor
treatment is also necessary to identify a subset of patients for
combination therapy with tyrosine kinase inhibitor and WNT
signaling-targeted therapeutics. By contrast, because WNT signaling
in the tumor microenvironment orchestrates antitumor immunity and
immune tolerance in a context-dependent manner, immune monitoring
is necessary to choose the appropriate WNT signaling-targeted
therapeutics for cancer patients with immune evasion (Fig. 3).
Investigational genome medicine platforms based on
nucleotide sequencing of transcribed regions are applicable for
determination of targeted therapeutics only in 10–24% of cancer
patients (163,164). Since alterations in
non-transcribed regulatory regions also drive human carcinogenesis,
whole-genome sequencing rather than whole- or partial-exome
sequencing is preferable to improve the precision of genome-based
medicine (4,165). In addition, organoid culture is a
cutting-edge technology in the fields of oncology and stem cell
biology (166–168), and organoid-based tests are also
used for selecting targeted therapeutics (163,166). However, because
tumor-stromal/immune interactions are not recapitulated in
patient-derived organoid models, immunological monitoring in the
tumor microenvironment is also necessary to improve genome-based
medicine.
Together, these findings indicate that 'omics
monitoring', including genome sequencing, transcriptomic,
immunohistochemical and organoid-based tests, before and during
treatment is necessary to choose and fine-tune WNT
signaling-targeted therapeutics for the treatment of cancer
patients (Fig. 3).
Cancer stem cells (CSCs) are part of the tumor
microenvironment and survive host defense or therapeutic insult
through omics reprogramming. Aberrant WNT signaling activation in
human cancers promotes CSC survival, bulk-tumor expansion and
invasion/metastasis. Anti-FZD mAb, anti-ROR1 mAb, anti-RSPO3 mAb,
PORCN inhibitors and β-catenin inhibitors are representative WNT
signaling-targeted therapeutics in clinical trials or preclinical
studies. WNT signaling-targeted therapeutics are applicable for
combination therapy with tyrosine kinase inhibitors or immune
checkpoint blockers. Omics monitoring is necessary for therapeutic
optimization of WNT signaling-targeted therapy.
This study was financially supported in part by a
grant-in-aid for the Knowledgebase Project from M. Katoh's
Fund.
|
1
|
Visvader JE and Lindeman GJ: Cancer stem
cells in solid tumours: Accumulating evidence and unresolved
questions. Nat Rev Cancer. 8:755–768. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Medema JP: Cancer stem cells: The
challenges ahead. Nat Cell Biol. 15:338–344. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Abbaszadegan MR, Bagheri V, Razavi MS,
Momtazi AA, Sahebkar A and Gholamin M: Isolation, identification,
and characterization of cancer stem cells: A review. J Cell
Physiol. 232:2008–2018. 2017. View Article : Google Scholar
|
|
4
|
Katoh M: Therapeutics targeting FGF
signaling network in human diseases. Trends Pharmacol Sci.
37:1081–1096. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
de Sousa e Melo F, Kurtova AV, Harnoss JM,
Kljavin N, Hoeck JD, Hung J, Anderson JE, Storm EE, Modrusan Z,
Koeppen H, et al: A distinct role for Lgr5(+) stem cells in primary
and metastatic colon cancer. Nature. 543:676–680. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Koury J, Zhong L and Hao J: Targeting
signaling pathways in cancer stem cells for cancer treatment. Stem
Cells Int. 2017:29258692017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McDonald OG, Li X, Saunders T,
Tryggvadottir R, Mentch SJ, Warmoes MO, Word AE, Carrer A, Salz TH,
Natsume S, et al: Epigenomic reprogramming during pancreatic cancer
progression links anabolic glucose metabolism to distant
metastasis. Nat Genet. 49:367–376. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tam WL and Weinberg RA: The epigenetics of
epithelial-mesenchymal plasticity in cancer. Nat Med. 19:1438–1449.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gonzalez DM and Medici D: Signaling
mechanisms of the epithelial-mesenchymal transition. Sci Signal.
7:re82014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schreiber RD, Old LJ and Smyth MJ: Cancer
immunoediting: Integrating immunity's roles in cancer suppression
and promotion. Science. 331:1565–1570. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Spranger S, Bao R and Gajewski TF:
Melanoma-intrinsic β-catenin signalling prevents anti-tumour
immunity. Nature. 523:231–235. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cairns RA, Harris IS and Mak TW:
Regulation of cancer cell metabolism. Nat Rev Cancer. 11:85–95.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
O'Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110. 2007.
View Article : Google Scholar
|
|
14
|
Yamashita T, Ji J, Budhu A, Forgues M,
Yang W, Wang HY, Jia H, Ye Q, Qin LX, Wauthier E, et al:
EpCAM-positive hepatocellular carcinoma cells are tumor-initiating
cells with stem/progenitor cell features. Gastroenterology.
136:1012–1024. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Todaro M, Gaggianesi M, Catalano V,
Benfante A, Iovino F, Biffoni M, Apuzzo T, Sperduti I, Volpe S,
Cocorullo G, et al: CD44v6 is a marker of constitutive and
reprogrammed cancer stem cells driving colon cancer metastasis.
Cell Stem Cell. 14:342–356. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hirsch D, Barker N, McNeil N, Hu Y, Camps
J, McKinnon K, Clevers H, Ried T and Gaiser T: LGR5 positivity
defines stem-like cells in colorectal cancer. Carcinogenesis.
35:849–858. 2014. View Article : Google Scholar :
|
|
17
|
Van der Flier LG, Sabates-Bellver J, Oving
I, Haegebarth A, De Palo M, Anti M, Van Gijn ME, Suijkerbuijk S,
Van de Wetering M, Marra G, et al: The intestinal Wnt/TCF
signature. Gastroenterology. 132:628–632. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yan KS, Janda CY, Chang J, Zheng GXY,
Larkin KA, Luca VC, Chia LA, Mah AT, Han A, Terry JM, et al:
Non-equivalence of Wnt and R-spondin ligands during Lgr5(+)
intestinal stem-cell self-renewal. Nature. 545:238–242. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hilkens J, Timmer NC, Boer M, Ikink GJ,
Schewe M, Sacchetti A, Koppens MAJ, Song JY and Bakker ERM: RSPO3
expands intestinal stem cell and niche compartments and drives
tumorigenesis. Gut. 66:1095–1105. 2017. View Article : Google Scholar :
|
|
20
|
Mani SK, Zhang H, Diab A, Pascuzzi PE,
Lefrançois L, Fares N, Bancel B, Merle P and Andrisani O:
EpCAM-regulated intra-membrane proteolysis induces a cancer stem
cell-like gene signature in hepatitis B virus-infected hepatocytes.
J Hepatol. 65:888–898. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Katoh M and Katoh M: WNT signaling pathway
and stem cell signaling network. Clin Cancer Res. 13:4042–4045.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ranganathan P, Weaver KL and Capobianco
AJ: Notch signalling in solid tumours: A little bit of everything
but not all the time. Nat Rev Cancer. 11:338–351. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Katoh M and Nakagama H: FGF receptors:
Cancer biology and therapeutics. Med Res Rev. 34:280–300. 2014.
View Article : Google Scholar
|
|
24
|
Lamb R, Bonuccelli G, Ozsvári B,
Peiris-Pagès M, Fiorillo M, Smith DL, Bevilacqua G, Mazzanti CM,
McDonnell LA, Naccarato AG, et al: Mitochondrial mass, a new
metabolic biomarker for stem-like cancer cells: Understanding
WNT/FGF-driven anabolic signaling. Oncotarget. 6:30453–30471. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Niehrs C: The complex world of WNT
receptor signalling. Nat Rev Mol Cell Biol. 13:767–779. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Holland JD, Klaus A, Garratt AN and
Birchmeier W: Wnt signaling in stem and cancer stem cells. Curr
Opin Cell Biol. 25:254–264. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rada P, Rojo AI, Offergeld A, Feng GJ,
Velasco-Martín JP, González-Sancho JM, Valverde ÁM, Dale T,
Regadera J and Cuadrado A: WNT-3A regulates an Axin1/NRF2 complex
that regulates antioxidant metabolism in hepatocytes. Antioxid
Redox Signal. 22:555–571. 2015. View Article : Google Scholar :
|
|
28
|
Acebron SP and Niehrs C:
β-catenin-independent roles of Wnt/LRP6 signaling. Trends Cell
Biol. 26:956–967. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Katoh M and Katoh M: Molecular genetics
and targeted therapy of WNT-related human diseases (Review). Int J
Mol Med. 40:587–606. 2017.PubMed/NCBI
|
|
30
|
Lui JH, Hansen DV and Kriegstein AR:
Development and evolution of the human neocortex. Cell. 146:18–36.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Barker N: Adult intestinal stem cells:
Critical drivers of epithelial homeostasis and regeneration. Nat
Rev Mol Cell Biol. 15:19–33. 2014. View Article : Google Scholar
|
|
32
|
Van Camp JK, Beckers S, Zegers D and Van
Hul W: Wnt signaling and the control of human stem cell fate. Stem
Cell Rev. 10:207–229. 2014. View Article : Google Scholar
|
|
33
|
Yang K, Wang X, Zhang H, Wang Z, Nan G, Li
Y, Zhang F, Mohammed MK, Haydon RC, Luu HH, et al: The evolving
roles of canonical WNT signaling in stem cells and tumorigenesis:
Implications in targeted cancer therapies. Lab Invest. 96:116–136.
2016. View Article : Google Scholar :
|
|
34
|
Qin L, Yin YT, Zheng FJ, Peng LX, Yang CF,
Bao YN, Liang YY, Li XJ, Xiang YQ, Sun R, et al: WNT5A promotes
stemness characteristics in nasopharyngeal carcinoma cells leading
to metastasis and tumorigenesis. Oncotarget. 6:10239–10252. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Webster MR, Kugel CH III and Weeraratna
AT: The Wnts of change: How Wnts regulate phenotype switching in
melanoma. Biochim Biophys Acta. 1856:244–251. 2015.PubMed/NCBI
|
|
36
|
Kumawat K and Gosens R: WNT-5A: Signaling
and functions in health and disease. Cell Mol Life Sci. 73:567–587.
2016. View Article : Google Scholar :
|
|
37
|
Wang MT, Holderfield M, Galeas J,
Delrosario R, To MD, Balmain A and McCormick F: K-Ras promotes
tumorigenicity through suppression of non-canonical Wnt signaling.
Cell. 163:1237–1251. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Plaks V, Kong N and Werb Z: The cancer
stem cell niche: How essential is the niche in regulating stemness
of tumor cells? Cell Stem Cell. 16:225–238. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Katoh M: FGFR inhibitors: Effects on
cancer cells, tumor microenvironment and whole-body homeostasis
(Review). Int J Mol Med. 38:3–15. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bolli N, Avet-Loiseau H, Wedge DC, Van Loo
P, Alexandrov LB, Martincorena I, Dawson KJ, Iorio F, Nik-Zainal S,
Bignell GR, et al: Heterogeneity of genomic evolution and
mutational profiles in multiple myeloma. Nat Commun. 5:29972014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li S, Garrett-Bakelman FE, Chung SS,
Sanders MA, Hricik T, Rapaport F, Patel J, Dillon R, Vijay P, Brown
AL, et al: Distinct evolution and dynamics of epigenetic and
genetic heterogeneity in acute myeloid leukemia. Nat Med.
22:792–799. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Abbosh C, Birkbak NJ, Wilson GA,
Jamal-Hanjani M, Constantin T, Salari R, Le Quesne J, Moore DA,
Veeriah S, Rosenthal R; TRACERx consortium; et al: PEACE
consortium: Phylogenetic ctDNA analysis depicts early-stage lung
cancer evolution. Nature. 545:446–451. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tammela T, Sanchez-Rivera FJ, Cetinbas NM,
Wu K, Joshi NS, Helenius K, Park Y, Azimi R, Kerper NR, Wesselhoeft
RA, et al: A Wnt-producing niche drives proliferative potential and
progression in lung adenocarcinoma. Nature. 545:355–359. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Weis SM and Cheresh DA: Tumor
angiogenesis: Molecular pathways and therapeutic targets. Nat Med.
17:1359–1370. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mao Y, Keller ET, Garfield DH, Shen K and
Wang J: Stromal cells in tumor microenvironment and breast cancer.
Cancer Metastasis Rev. 32:303–315. 2013. View Article : Google Scholar
|
|
46
|
Son B, Lee S, Youn H, Kim E, Kim W and
Youn B: The role of tumor microenvironment in therapeutic
resistance. Oncotarget. 8:3933–3945. 2017.
|
|
47
|
Anderson KG, Stromnes IM and Greenberg PD:
Obstacles posed by the tumor microenvironment to T cell activity: A
case for synergistic therapies. Cancer Cell. 31:311–325. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang Z, Liu P, Inuzuka H and Wei W: Roles
of F-box proteins in cancer. Nat Rev Cancer. 14:233–247. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Katoh M, Hirai M, Sugimura T and Terada M:
Cloning, expression and chromosomal localization of Wnt-13, a novel
member of the Wnt gene family. Oncogene. 13:873–876.
1996.PubMed/NCBI
|
|
50
|
Katoh M, Kirikoshi H, Terasaki H and
Shiokawa K: WNT2B2 mRNA, up-regulated in primary gastric cancer, is
a positive regulator of the WNT-β-catenin-TCF signaling pathway.
Biochem Biophys Res Commun. 289:1093–1098. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jiang H, Li F, He C, Wang X, Li Q and Gao
H: Expression of Gli1 and Wnt2B correlates with progression and
clinical outcome of pancreatic cancer. Int J Clin Exp Pathol.
7:4531–4538. 2014.PubMed/NCBI
|
|
52
|
Steinhart Z, Pavlovic Z, Chandrashekhar M,
Hart T, Wang X, Zhang X, Robitaille M, Brown KR, Jaksani S,
Overmeer R, et al: Genome-wide CRISPR screens reveal a Wnt-FZD5
signaling circuit as a druggable vulnerability of RNF43-mutant
pancreatic tumors. Nat Med. 23:60–68. 2017. View Article : Google Scholar
|
|
53
|
Seshagiri S, Stawiski EW, Durinck S,
Modrusan Z, Storm EE, Conboy CB, Chaudhuri S, Guan Y, Janakiraman
V, Jaiswal BS, et al: Recurrent R-spondin fusions in colon cancer.
Nature. 488:660–664. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kinzler KW and Vogelstein B: Lessons from
hereditary colorectal cancer. Cell. 87:159–170. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Mazzoni SM and Fearon ER: AXIN1 and AXIN2
variants in gastrointestinal cancers. Cancer Lett. 355:1–8. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Chiche A, Moumen M, Romagnoli M, Petit V,
Lasla H, Jézéquel P, de la Grange P, Jonkers J, Deugnier MA,
Glukhova MA, et al: p53 deficiency induces cancer stem cell pool
expansion in a mouse model of triple-negative breast tumors.
Oncogene. 36:2355–2365. 2017. View Article : Google Scholar
|
|
57
|
Valenti G, Quinn HM, Heynen GJJE, Lan L,
Holland JD, Vogel R, Wulf-Goldenberg A and Birchmeier W: Cancer
stem cells regulate cancer-associated fibroblasts via activation of
Hedgehog signaling in mammary gland tumors. Cancer Res.
77:2134–2147. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Katoh M: WNT/PCP signaling pathway and
human cancer (Review). Oncol Rep. 14:1583–1588. 2005.PubMed/NCBI
|
|
59
|
Yang Y and Mlodzik M: Wnt-Frizzled/planar
cell polarity signaling: Cellular orientation by facing the wind
(Wnt). Annu Rev Cell Dev Biol. 31:623–646. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Minegishi K, Hashimoto M, Ajima R, Takaoka
K, Shinohara K, Ikawa Y, Nishimura H, McMahon AP, Willert K, Okada
Y, et al: A Wnt5 activity asymmetry and intercellular signaling via
PCP proteins polarize node cells for left-right symmetry breaking.
Dev Cell. 40:439–452.e4. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Wu J and Mlodzik M: Wnt/PCP instructions
for cilia in left-right asymmetry. Dev Cell. 40:423–424. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang W, Runkle KB, Terkowski SM, Ekaireb
RI and Witze ES: Protein depalmitoylation is induced by Wnt5a and
promotes polarized cell behavior. J Biol Chem. 290:15707–15716.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Nishimura T, Honda H and Takeichi M:
Planar cell polarity links axes of spatial dynamics in neural-tube
closure. Cell. 149:1084–1097. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
De Marco P, Merello E, Piatelli G, Cama A,
Kibar Z and Capra V: Planar cell polarity gene mutations contribute
to the etiology of human neural tube defects in our population.
Birth Defects Res A Clin Mol Teratol. 100:633–641. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Gödde NJ, Pearson HB, Smith LK and Humbert
PO: Dissecting the role of polarity regulators in cancer through
the use of mouse models. Exp Cell Res. 328:249–257. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Johnson R and Halder G: The two faces of
Hippo: Targeting the Hippo pathway for regenerative medicine and
cancer treatment. Nat Rev Drug Discov. 13:63–79. 2014. View Article : Google Scholar :
|
|
67
|
Zhang S, Chen L, Cui B, Chuang HY, Yu J,
Wang-Rodriguez J, Tang L, Chen G, Basak GW and Kipps TJ: ROR1 is
expressed in human breast cancer and associated with enhanced
tumor-cell growth. PLoS One. 7:e311272012. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Anastas JN, Kulikauskas RM, Tamir T, Rizos
H, Long GV, von Euw EM, Yang PT, Chen HW, Haydu L, Toroni RA, et
al: WNT5A enhances resistance of melanoma cells to targeted BRAF
inhibitors. J Clin Invest. 124:2877–2890. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Yu J, Chen L, Cui B, Widhopf GF II, Shen
Z, Wu R, Zhang L, Zhang S, Briggs SP and Kipps TJ: Wnt5a induces
ROR1/ROR2 heterooligomerization to enhance leukemia chemotaxis and
proliferation. J Clin Invest. 126:585–598. 2016. View Article : Google Scholar :
|
|
70
|
Green JL, Kuntz SG and Sternberg PW: Ror
receptor tyrosine kinases: Orphans no more. Trends Cell Biol.
18:536–544. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Lu W, Yamamoto V, Ortega B and Baltimore
D: Mammalian Ryk is a Wnt coreceptor required for stimulation of
neurite outgrowth. Cell. 119:97–108. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Petrova IM, Malessy MJ, Verhaagen J,
Fradkin LG and Noordermeer JN: Wnt signaling through the Ror
receptor in the nervous system. Mol Neurobiol. 49:303–315. 2014.
View Article : Google Scholar
|
|
73
|
Debebe Z and Rathmell WK: Ror2 as a
therapeutic target in cancer. Pharmacol Ther. 150:143–148. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Bicocca VT, Chang BH, Masouleh BK, Muschen
M, Loriaux MM, Druker BJ and Tyner JW: Crosstalk between ROR1 and
the Pre-B cell receptor promotes survival of t(1;19) acute
lymphoblastic leukemia. Cancer Cell. 22:656–667. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Hojjat-Farsangi M, Moshfegh A,
Daneshmanesh AH, Khan AS, Mikaelsson E, Osterborg A and Mellstedt
H: The receptor tyrosine kinase ROR1 - an oncofetal antigen for
targeted cancer therapy. Semin Cancer Biol. 29:21–31. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Gentile A, Lazzari L, Benvenuti S,
Trusolino L and Comoglio PM: The ROR1 pseudokinase diversifies
signaling outputs in MET-addicted cancer cells. Int J Cancer.
135:2305–2316. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Yamaguchi T, Lu C, Ida L, Yanagisawa K,
Usukura J, Cheng J, Hotta N, Shimada Y, Isomura H, Suzuki M, et al:
ROR1 sustains caveolae and survival signalling as a scaffold of
cavin-1 and caveolin-1. Nat Commun. 7:100602016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Li C, Wang S, Xing Z, Lin A, Liang K, Song
J, Hu Q, Yao J, Chen Z, Park PK, et al: A ROR1-HER3-lncRNA
signalling axis modulates the Hippo-YAP pathway to regulate bone
metastasis. Nat Cell Biol. 19:106–119. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Dijksterhuis JP, Petersen J and Schulte G:
WNT/Frizzled signalling: receptor-ligand selectivity with focus on
FZD-G protein signalling and its physiological relevance: IUPHAR
Review 3. Br J Pharmacol. 171:1195–1209. 2014. View Article : Google Scholar :
|
|
80
|
Zhan T, Rindtorff N and Boutros M: Wnt
signaling in cancer. Oncogene. 36:1461–1473. 2017. View Article : Google Scholar :
|
|
81
|
Gong B, Shen W, Xiao W, Meng Y, Meng A and
Jia S: The Sec14-like phosphatidylinositol transfer proteins
Sec14l3/SEC14L2 act as GTPase proteins to mediate Wnt/Ca(2+)
signaling. eLife. 6:e263622017. View Article : Google Scholar
|
|
82
|
Kim S, Nie H, Nesin V, Tran U, Outeda P,
Bai CX, Keeling J, Maskey D, Watnick T, Wessely O, et al: The
polycystin complex mediates Wnt/Ca(2+) signalling. Nat Cell Biol.
18:752–764. 2016.PubMed/NCBI
|
|
83
|
Ishitani T, Kishida S, Hyodo-Miura J, Ueno
N, Yasuda J, Waterman M, Shibuya H, Moon RT, Ninomiya-Tsuji J and
Matsumoto K: The TAK1-NLK mitogen-activated protein kinase cascade
functions in the Wnt-5a/Ca(2+) pathway to antagonize Wnt/β-catenin
signaling. Mol Cell Biol. 23:131–139. 2003. View Article : Google Scholar :
|
|
84
|
Zhang M, Hagenmueller M, Riffel JH,
Kreusser MM, Bernhold E, Fan J, Katus HA, Backs J and Hardt SE:
Calcium/calmodulin-dependent protein kinase II couples Wnt
signaling with histone deacetylase 4 and mediates
dishevelled-induced cardiomyopathy. Hypertension. 65:335–344. 2015.
View Article : Google Scholar
|
|
85
|
Scholz B, Korn C, Wojtarowicz J, Mogler C,
Augustin I, Boutros M, Niehrs C and Augustin HG: Endothelial RSPO3
controls vascular stability and pruning through non-canonical
WNT/Ca(2+)/NFAT signaling. Dev Cell. 36:79–93. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Wang W, Snyder N, Worth AJ, Blair IA and
Witze ES: Regulation of lipid synthesis by the RNA helicase Mov10
controls Wnt5a production. Oncogenesis. 4:e1542015. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Miyamoto DT, Zheng Y, Wittner BS, Lee RJ,
Zhu H, Broderick KT, Desai R, Fox DB, Brannigan BW, Trautwein J, et
al: RNA-Seq of single prostate CTCs implicates noncanonical Wnt
signaling in antiandrogen resistance. Science. 349:1351–1356. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Blumenthal A, Ehlers S, Lauber J, Buer J,
Lange C, Goldmann T, Heine H, Brandt E and Reiling N: The Wingless
homolog WNT5A and its receptor Frizzled-5 regulate inflammatory
responses of human mononuclear cells induced by microbial
stimulation. Blood. 108:965–973. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Wang L, Steele I, Kumar JD, Dimaline R,
Jithesh PV, Tiszlavicz L, Reisz Z, Dockray GJ and Varro A: Distinct
miRNA profiles in normal and gastric cancer myofibroblasts and
significance in Wnt signaling. Am J Physiol Gastrointest Liver
Physiol. 310:G696–G704. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Aznar N, Midde KK, Dunkel Y, Lopez-Sanchez
I, Pavlova Y, Marivin A, Barbazán J, Murray F, Nitsche U, Janssen
KP, et al: Daple is a novel non-receptor GEF required for trimeric
G protein activation in Wnt signaling. eLife. 4:e070912015.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yu FX, Zhao B and Guan KL: Hippo pathway
in organ size control, tissue homeostasis, and cancer. Cell.
163:811–828. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Hansen CG, Moroishi T and Guan KL: YAP and
TAZ: A nexus for Hippo signaling and beyond. Trends Cell Biol.
25:499–513. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Hot B, Valnohova J, Arthofer E, Simon K,
Shin J, Uhlén M, Kostenis E, Mulder J and Schulte G: FZD10-Gα13
signalling axis points to a role of FZD10 in CNS angiogenesis. Cell
Signal. 32:93–103. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Takebe N, Miele L, Harris PJ, Jeong W,
Bando H, Kahn M, Yang SX and Ivy SP: Targeting Notch, Hedgehog, and
Wnt pathways in cancer stem cells: Clinical update. Nat Rev Clin
Oncol. 12:445–464. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Kahn M: Wnt signaling in stem cells and
tumor stem cells. Semin Reprod Med. 33:317–325. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Tai D, Wells K, Arcaroli J, Vanderbilt C,
Aisner DL, Messersmith WA and Lieu CH: Targeting the WNT signaling
pathway in cancer therapeutics. Oncologist. 20:1189–1198. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Gurney A, Axelrod F, Bond CJ, Cain J,
Chartier C, Donigan L, Fischer M, Chaudhari A, Ji M, Kapoun AM, et
al: Wnt pathway inhibition via the targeting of Frizzled receptors
results in decreased growth and tumorigenicity of human tumors.
Proc Natl Acad Sci USA. 109:11717–11722. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Nielsen TO, Poulin NM and Ladanyi M:
Synovial sarcoma: Recent discoveries as a roadmap to new avenues
for therapy. Cancer Discov. 5:124–134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Gong X, Azhdarinia A, Ghosh SC, Xiong W,
An Z, Liu Q and Carmon KS: LGR5-targeted antibody-drug conjugate
eradicates gastrointestinal tumors and prevents recurrence. Mol
Cancer Ther. 15:1580–1590. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Damelin M, Bankovich A, Bernstein J, Lucas
J, Chen L, Williams S, Park A, Aguilar J, Ernstoff E, Charati M, et
al: A PTK7-targeted antibody-drug conjugate reduces tumorinitiating
cells and induces sustained tumor regressions. Sci Transl Med.
9:pii: eaag2611. 2017. View Article : Google Scholar
|
|
101
|
Zhang S, Cui B, Lai H, Liu G, Ghia EM,
Widhopf GF II, Zhang Z, Wu CC, Chen L, Wu R, et al: Ovarian cancer
stem cells express ROR1, which can be targeted for
anti-cancer-stem-cell therapy. Proc Natl Acad Sci USA.
111:17266–17271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Storm EE, Durinck S, de Sousa e Melo F,
Tremayne J, Kljavin N, Tan C, Ye X, Chiu C, Pham T, Hongo JA, et
al: Targeting PTPRK-RSPO3 colon tumours promotes differentiation
and loss of stem-cell function. Nature. 529:97–100. 2016.
View Article : Google Scholar
|
|
103
|
Berger C, Sommermeyer D, Hudecek M, Berger
M, Balakrishnan A, Paszkiewicz PJ, Kosasih PL, Rader C and Riddell
SR: Safety of targeting ROR1 in primates with chimeric antigen
receptor-modified T cells. Cancer Immunol Res. 3:206–216. 2015.
View Article : Google Scholar :
|
|
104
|
Le PN, McDermott JD and Jimeno A:
Targeting the Wnt pathway in human cancers: Therapeutic targeting
with a focus on OMP-54F28. Pharmacol Ther. 146:1–11. 2015.
View Article : Google Scholar
|
|
105
|
Cheng Y, Phoon YP, Jin X, Chong SY, Ip JC,
Wong BW and Lung ML: Wnt-C59 arrests stemness and suppresses growth
of nasopharyngeal carcinoma in mice by inhibiting the Wnt pathway
in the tumor microenvironment. Oncotarget. 6:14428–14439. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Poulsen A, Ho SY, Wang W, Alam J, Jeyaraj
DA, Ang SH, Tan ES, Lin GR, Cheong VW, Ke Z, et al: Pharmacophore
model for Wnt/Porcupine inhibitors and its use in drug design. J
Chem Inf Model. 55:1435–1448. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Langton PF, Kakugawa S and Vincent JP:
Making, exporting, and modulating Wnts. Trends Cell Biol.
26:756–765. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
DeBruine ZJ, Ke J, Harikumar KG, Gu X,
Borowsky P, Williams BO, Xu W, Miller LJ, Xu HE and Melcher K:
Wnt5a promotes Frizzled-4 signalosome assembly by stabilizing
cysteine-rich domain dimerization. Genes Dev. 31:916–926. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Madan B, Ke Z, Harmston N, Ho SY, Frois
AO, Alam J, Jeyaraj DA, Pendharkar V, Ghosh K, Virshup IH, et al:
Wnt addiction of genetically defined cancers reversed by PORCN
inhibition. Oncogene. 35:2197–2207. 2016. View Article : Google Scholar
|
|
110
|
Chen B, Dodge ME, Tang W, Lu J, Ma Z, Fan
CW, Wei S, Hao W, Kilgore J, Williams NS, et al: Small
molecule-mediated disruption of Wnt-dependent signaling in tissue
regeneration and cancer. Nat Chem Biol. 5:100–107. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Proffitt KD, Madan B, Ke Z, Pendharkar V,
Ding L, Lee MA, Hannoush RN and Virshup DM: Pharmacological
inhibition of the Wnt acyltransferase PORCN prevents growth of
WNT-driven mammary cancer. Cancer Res. 73:502–507. 2013. View Article : Google Scholar
|
|
112
|
Agarwal P, Zhang B, Ho Y, Cook A, Li L,
Mikhail FM, Wang Y, McLaughlin ME and Bhatia R: Enhanced targeting
of CML stem and progenitor cells by inhibition of porcupine
acyltrans-ferase in combination with TKI. Blood. 129:1008–1020.
2017. View Article : Google Scholar
|
|
113
|
Liu C and Yu X: ADP-ribosyltransferases
and poly ADP-ribosylation. Curr Protein Pept Sci. 16:491–501. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Kulak O, Chen H, Holohan B, Wu X, He H,
Borek D, Otwinowski Z, Yamaguchi K, Garofalo LA, Ma Z, et al:
Disruption of Wnt/β-catenin signaling and telomeric shortening are
inextricable consequences of tankyrase inhibition in human cells.
Mol Cell Biol. 35:2425–2435. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Wang W, Li N, Li X, Tran MK, Han X and
Chen J: Tankyrase inhibitors target YAP by stabilizing Angiomotin
family proteins. Cell Rep. 13:524–532. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Nathubhai A, Haikarainen T, Koivunen J,
Murthy S, Koumanov F, Lloyd MD, Holman GD, Pihlajaniemi T, Tosh D,
Lehtiö L, et al: Highly potent and isoform selective dual site
binding Tankyrase/Wnt signaling inhibitors that increase cellular
glucose uptake and have antiproliferative activity. J Med Chem.
60:814–820. 2017. View Article : Google Scholar
|
|
117
|
Quackenbush KS, Bagby S, Tai WM,
Messersmith WA, Schreiber A, Greene J, Kim J, Wang G, Purkey A,
Pitts TM, et al: The novel tankyrase inhibitor (AZ1366) enhances
irinotecan activity in tumors that exhibit elevated tankyrase and
irinotecan resistance. Oncotarget. 7:28273–28285. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Lau T, Chan E, Callow M, Waaler J, Boggs
J, Blake RA, Magnuson S, Sambrone A, Schutten M, Firestein R, et
al: A novel tankyrase small-molecule inhibitor suppresses APC
mutation-driven colorectal tumor growth. Cancer Res. 73:3132–3144.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Waaler J, Machon O, Tumova L, Dinh H,
Korinek V, Wilson SR, Paulsen JE, Pedersen NM, Eide TJ, Machonova
O, et al: A novel tankyrase inhibitor decreases canonical Wnt
signaling in colon carcinoma cells and reduces tumor growth in
conditional APC mutant mice. Cancer Res. 72:2822–2832. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Arqués O, Chicote I, Puig I, Tenbaum SP,
Argilés G, Dienstmann R, Fernández N, Caratù G, Matito J,
Silberschmidt D, et al: Tankyrase inhibition blocks Wnt/β-catenin
pathway and reverts resistance to PI3K and AKT inhibitors in the
treatment of colorectal cancer. Clin Cancer Res. 22:644–656. 2016.
View Article : Google Scholar
|
|
121
|
Huang SM, Mishina YM, Liu S, Cheung A,
Stegmeier F, Michaud GA, Charlat O, Wiellette E, Zhang Y, Wiessner
S, et al: Tankyrase inhibition stabilizes axin and antagonizes Wnt
signalling. Nature. 461:614–620. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Schoumacher M, Hurov KE, Lehár J,
Yan-Neale Y, Mishina Y, Sonkin D, Korn JM, Flemming D, Jones MD,
Antonakos B, et al: Inhibiting Tankyrases sensitizes KRAS-mutant
cancer cells to MEK inhibitors via FGFR2 feedback signaling. Cancer
Res. 74:3294–3305. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Wang H, Lu B, Castillo J, Zhang Y, Yang Z,
McAllister G, Lindeman A, Reece-Hoyes J, Tallarico J, Russ C, et
al: Tankyrase inhibitor sensitizes lung cancer cells to endothelial
growth factor receptor (EGFR) inhibition via stabilizing
angiomotins and inhibiting YAP signaling. J Biol Chem.
291:15256–15266. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Scarborough HA, Helfrich BA, Casás-Selves
M, Schuller AG, Grosskurth SE, Kim J, Tan AC, Chan DC, Zhang Z,
Zaberezhnyy V, et al: AZ1366: An inhibitor of tankyrase and the
canonical Wnt pathway that limits the persistence of non-small cell
lung cancer cells following EGFR inhibition. Clin Cancer Res.
23:1531–1541. 2017. View Article : Google Scholar
|
|
125
|
Pelay-Gimeno M, Glas A, Koch O and
Grossmann TN: Structure-based design of inhibitors of
protein-protein interactions: Mimicking peptide binding epitopes.
Angew Chem Int Ed Engl. 54:8896–8927. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Hwang SY, Deng X, Byun S, Lee C, Lee SJ,
Suh H, Zhang J, Kang Q, Zhang T, Westover KD, et al: Direct
targeting of β-catenin by a small molecule stimulates proteasomal
degradation and suppresses oncogenic Wnt/β-catenin signaling. Cell
Rep. 16:28–36. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Mahmoudi T, Li VS, Ng SS, Taouatas N,
Vries RG, Mohammed S, Heck AJ and Clevers H: The kinase TNIK is an
essential activator of Wnt target genes. EMBO J. 28:3329–3340.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Lee Y, Jung JI, Park KY, Kim SA and Kim J:
Synergistic inhibition effect of TNIK inhibitor KY-05009 and
receptor tyrosine kinase inhibitor dovitinib on IL-6-induced
proliferation and Wnt signaling pathway in human multiple myeloma
cells. Oncotarget. 8:41091–41101. 2017.PubMed/NCBI
|
|
129
|
Tan Z, Chen L and Zhang S: Comprehensive
modeling and discovery of mebendazole as a novel TRAF2- and
NCK-interacting kinase inhibitor. Sci Rep. 6:335342016. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Fiskus W, Sharma S, Saha S, Shah B,
Devaraj SG, Sun B, Horrigan S, Leveque C, Zu Y, Iyer S, et al:
Pre-clinical efficacy of combined therapy with novel β-catenin
antagonist BC2059 and histone deacetylase inhibitor against AML
cells. Leukemia. 29:1267–1278. 2015. View Article : Google Scholar
|
|
131
|
Trautmann M, Sievers E, Aretz S, Kindler
D, Michels S, Friedrichs N, Renner M, Kirfel J, Steiner S, Huss S,
et al: SS18-SSX fusion protein-induced Wnt/β-catenin signaling is a
therapeutic target in synovial sarcoma. Oncogene. 33:5006–5016.
2014. View Article : Google Scholar
|
|
132
|
Kim JY, Lee HY, Park KK, Choi YK, Nam JS
and Hong IS: CWP232228 targets liver cancer stem cells through
Wnt/β-catenin signaling: A novel therapeutic approach for liver
cancer treatment. Oncotarget. 7:20395–20409. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Nagaraj AB, Joseph P, Kovalenko O, Singh
S, Armstrong A, Redline R, Resnick K, Zanotti K, Waggoner S and
DiFeo A: Critical role of Wnt/β-catenin signaling in driving
epithelial ovarian cancer platinum resistance. Oncotarget.
6:23720–23734. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Fang L, Zhu Q, Neuenschwander M, Specker
E, Wulf-Goldenberg A, Weis WI, von Kries JP and Birchmeier W: A
small-molecule antagonist of the β-catenin/TCF4 interaction blocks
the self-renewal of cancer stem cells and suppresses tumorigenesis.
Cancer Res. 76:891–901. 2016. View Article : Google Scholar
|
|
135
|
Sukhdeo K, Mani M, Zhang Y, Dutta J, Yasui
H, Rooney MD, Carrasco DE, Zheng M, He H, Tai YT, et al: Targeting
the β-catenin/TCF transcriptional complex in the treatment of
multiple myeloma. Proc Natl Acad Sci USA. 104:7516–7521. 2007.
View Article : Google Scholar
|
|
136
|
Zhou H, Mak PY, Mu H, Mak DH, Zeng Z,
Cortes J, Liu Q, Andreeff M and Carter BZ: Combined inhibition of
β-catenin and Bcr-Abl synergistically targets tyrosine kinase
inhibitor-resistant blast crisis chronic myeloid leukemia blasts
and progenitors in vitro and in vivo. Leukemia. Apr 18–2017.Epub
ahead of print. View Article : Google Scholar
|
|
137
|
Takada K, Zhu D, Bird GH, Sukhdeo K, Zhao
JJ, Mani M, Lemieux M, Carrasco DE, Ryan J, Horst D, et al:
Targeted disruption of the BCL9/β-catenin complex inhibits
oncogenic Wnt signaling. Sci Transl Med. 4:148ra1172012. View Article : Google Scholar
|
|
138
|
Wang Q, Amato SP, Rubitski DM, Hayward MM,
Kormos BL, Verhoest PR, Xu L, Brandon NJ and Ehlers MD:
Identification of phosphorylation consensus sequences and
endogenous neuronal substrates of the psychiatric risk kinase TNIK.
J Pharmacol Exp Ther. 356:410–423. 2016. View Article : Google Scholar
|
|
139
|
Coluccia AM, Vacca A, Duñach M, Mologni L,
Redaelli S, Bustos VH, Benati D, Pinna LA and Gambacorti-Passerini
C: Bcr-Abl stabilizes β-catenin in chronic myeloid leukemia through
its tyrosine phosphorylation. EMBO J. 26:1456–1466. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Nakayama S, Sng N, Carretero J, Welner R,
Hayashi Y, Yamamoto M, Tan AJ, Yamaguchi N, Yasuda H, Li D, et al:
β-catenin contributes to lung tumor development induced by EGFR
mutations. Cancer Res. 74:5891–5902. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Kajiguchi T, Katsumi A, Tanizaki R, Kiyoi
H and Naoe T: Y654 of β-catenin is essential for FLT3/ITD-related
tyrosine phosphorylation and nuclear localization of β-catenin. Eur
J Haematol. 88:314–320. 2012. View Article : Google Scholar
|
|
142
|
Jin B, Ding K and Pan J: Ponatinib induces
apoptosis in imatinib-resistant human mast cells by
dephosphorylating mutant D816V KIT and silencing β-catenin
signaling. Mol Cancer Ther. 13:1217–1230. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Fernández-Sánchez ME, Barbier S, Whitehead
J, Béalle G, Michel A, Latorre-Ossa H, Rey C, Fouassier L, Claperon
A, Brullé L, et al: Mechanical induction of the tumorigenic
β-catenin pathway by tumour growth pressure. Nature. 523:92–95.
2015. View Article : Google Scholar
|
|
144
|
Zhao Y, Masiello D, McMillian M, Nguyen C,
Wu Y, Melendez E, Smbatyan G, Kida A, He Y, Teo JL, et al:
CBP/catenin antagonist safely eliminates drug-resistant
leukemiainitiating cells. Oncogene. 35:3705–3717. 2016. View Article : Google Scholar
|
|
145
|
Smyth MJ, Ngiow SF, Ribas A and Teng MW:
Combination cancer immunotherapies tailored to the tumour
microenvironment. Nat Rev Clin Oncol. 13:143–158. 2016. View Article : Google Scholar
|
|
146
|
Palucka AK and Coussens LM: The basis of
oncoimmunology. Cell. 164:1233–1247. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Zarour HM: Reversing T-cell dysfunction
and exhaustion in cancer. Clin Cancer Res. 22:1856–1864. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Chen DS and Mellman I: Elements of cancer
immunity and the cancer-immune set point. Nature. 541:321–330.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Inman BA, Longo TA, Ramalingam S and
Harrison MR: Atezolizumab: A PD-L1-blocking antibody for bladder
cancer. Clin Cancer Res. 23:1886–1890. 2017. View Article : Google Scholar
|
|
150
|
Kim ES: Avelumab: First global approval.
Drugs. 77:929–937. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Syed YY: Durvalumab: First global
approval. Drugs. 77:1369–1376. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Zaretsky JM, Garcia-Diaz A, Shin DS,
Escuin-Ordinas H, Hugo W, Hu-Lieskovan S, Torrejon DY,
Abril-Rodriguez G, Sandoval S, Barthly L, et al: Mutations
associated with acquired resistance to PD-1 blockade in melanoma. N
Engl J Med. 375:819–829. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Shin DS, Zaretsky JM, Escuin-Ordinas H,
Garcia-Diaz A, Hu-Lieskovan S, Kalbasi A, Grasso CS, Hugo W,
Sandoval S, Torrejon DY, et al: Primary resistance to PD-1 blockade
mediated by JAK1/2 mutations. Cancer Discov. 7:188–201. 2017.
View Article : Google Scholar
|
|
154
|
Anagnostou V, Smith KN, Forde PM, Niknafs
N, Bhattacharya R, White J, Zhang T, Adleff V, Phallen J, Wali N,
et al: Evolution of neoantigen landscape during immune checkpoint
blockade in non-small cell lung cancer. Cancer Discov. 7:264–276.
2017. View Article : Google Scholar
|
|
155
|
Huang AC, Postow MA, Orlowski RJ, Mick R,
Bengsch B, Manne S, Xu W, Harmon S, Giles JR, Wenz B, et al: T-cell
invigoration to tumour burden ratio associated with anti-PD-1
response. Nature. 545:60–65. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Manguso RT, Pope HW, Zimmer MD, Brown FD,
Yates KB, Miller BC, Collins NB, Bi K, LaFleur MW, Juneja VR, et
al: In vivo CRISPR screening identifies Ptpn2 as a cancer
immunotherapy target. Nature. 547:413–418. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Holtzhausen A, Zhao F, Evans KS, Tsutsui
M, Orabona C, Tyler DS and Hanks BA: Melanoma-derived Wnt5a
promotes local dendritic-cell expression of IDO and
immunotolerance: Opportunities for pharmacologic enhancement of
immunotherapy. Cancer Immunol Res. 3:1082–1095. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Hugo W, Zaretsky JM, Sun L, Song C, Moreno
BH, Hu-Lieskovan S, Berent-Maoz B, Pang J, Chmielowski B, Cherry G,
et al: Genomic and transcriptomic features of response to anti-PD-1
therapy in metastatic melanoma. Cell. 165:35–44. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
D'Amico L, Mahajan S, Capietto AH, Yang Z,
Zamani A, Ricci B, Bumpass DB, Meyer M, Su X, Wang-Gillam A, et al:
Dickkopf-related protein 1 (Dkk1) regulates the accumulation and
function of myeloid derived suppressor cells in cancer. J Exp Med.
213:827–840. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Fulciniti M, Tassone P, Hideshima T,
Vallet S, Nanjappa P, Ettenberg SA, Shen Z, Patel N, Tai YT,
Chauhan D, et al: Anti-DKK1 mAb (BHQ880) as a potential therapeutic
agent for multiple myeloma. Blood. 114:371–379. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Bendell JC, Murphy JE, Mahalingam D,
Halmos B, Sirard CA, Landau SB and Ryan DP: A Phase 1 study of
DKN-01, an anti-DKK1 antibody, in combination with paclitaxel in
patients with DKK1 relapsed or refractory esophageal cancer or
gastro-esophageal junction tumors. J Clin Oncol. 34(Suppl 4): pp.
S1112016, http://ascopubs.org/doi/abs/10.1200/jco.2016.34.4_suppl.111.
View Article : Google Scholar
|
|
162
|
Camidge DR, Pao W and Sequist LV: Acquired
resistance to TKIs in solid tumours: Learning from lung cancer. Nat
Rev Clin Oncol. 11:473–481. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Pauli C, Hopkins BD, Prandi D, Shaw R,
Fedrizzi T, Sboner A, Sailer V, Augello M, Puca L, Rosati R, et al:
Personalized in vitro and in vivo cancer models to guide precision
medicine. Cancer Discov. 7:462–477. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Massard C, Michiels S, Ferté C, Le Deley
MC, Lacroix L, Hollebecque A, Verlingue L, Ileana E, Rosellini S,
Ammari S, et al: High-throughput genomics and clinical outcome in
hard-to-treat advanced cancers: Results of the MOSCATO 01 trial.
Cancer Discov. 7:586–595. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Rheinbay E, Parasuraman P, Grimsby J, Tiao
G, Engreitz JM, Kim J, Lawrence MS, Taylor-Weiner A,
Rodriguez-Cuevas S, Rosenberg M, et al: Recurrent and functional
regulatory mutations in breast cancer. Nature. 547:55–60. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
166
|
van de Wetering M, Francies HE, Francis
JM, Bounova G, Iorio F, Pronk A, van Houdt W, van Gorp J,
Taylor-Weiner A, Kester L, et al: Prospective derivation of a
living organoid biobank of colorectal cancer patients. Cell.
161:933–945. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Merker SR, Weitz J and Stange DE:
Gastrointestinal organoids: How they gut it out. Dev Biol.
420:239–250. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Zhang L, Adileh M, Martin ML, Klingler S,
White J, Ma X, Howe LR, Brown AM and Kolesnick R: Establishing
estrogen-responsive mouse mammary organoids from single Lgr5(+)
cells. Cell Signal. 29:41–51. 2017. View Article : Google Scholar
|