Introduction
Bladder cancer (BC) is the most common malignancy of
the urinary tract and is associated with significantly high
morbidity and mortality (1,2). BC
can be divided into three groups: non-muscle invasive BC (NMIBC),
muscle-invasive BC(MIBC) and metastatic BC. Approximately 20–25% of
patients were found to have MIBC and the remaining 75–80% of
patients had NMIBC at the first diagnosis (3). Transurethral resection of bladder
tumor (TRUBT) combined with intravesical chemotherapy is typically
the primary method for the treatment of NMIBC (4). Once BC invades muscle, radical
cystectomy should be performed to improve the treatment success
rate (5). Due to current surgical
limitations in radical cystectomy, up to 50% of MIBC patients will
eventually experience recurrence and progress to metastatic disease
(3). If metastatic BC presents,
systemic chemotherapy and PD-1/PD-L1 antibody based immunotherapy
are the primary choices to prolong the quantity and quality of
life, although the majority of patients may eventually die of their
disease (6,7). These statistics demonstrates the
urgent necessity for the discovery of new biomarkers to detect BC,
as well as the need to produce new pharmaceutical solutions for the
treatment of BC.
Recent data obtained from the ENCODE consortium
indicate that ~70% of the human genome is transcribed, generating a
vast range of non-coding RNAs (8).
Long non-coding RNAs (lncRNAs) are a class of RNA molecules defined
as transcripts longer than 200 nucleotides, although these
molecules lack protein-coding potential. lncRNAs are found in sense
or antisense orientation for protein-coding genes, within introns
of protein-coding genes or intergenic regions of the genome
(9). Though small non-coding RNAs
(<200 bp), particularly miRNAs (21-23 nucleotides), are well
characterized as post-transcriptional regulators of mRNAs and their
roles in cancer are well established (10,11),
lncRNAs are still under investigation and more information is
needed to understand the role of lncRNAs in cancer occurrence and
progression (12).
Several lncRNAs have been identified for their
relation to BC. For example, UCA1 was demonstrated to be highly
expressed in the BC tissues, and promote glutamine metabolism by
targeting miR-16 in human BC (13). ANRIL was reported to be
significantly overexpressed in BC tissues and regulated BC cell
proliferation and apoptosis both in vitro and in vivo
(14). GHET1 was demonstrated to
be significantly upregulated in BC tissues and promoted the
proliferation and invasion of BC cells (15). These results indicate that lncRNAs
can be a possible source of valuable information to better
understand the potential mechanisms underlying BC occurrence and
progression.
DUXAP10 is a newly found lncRNA, the expression of
which was significantly elevated in BC tissues compared to matched
normal bladder tissues (>8-fold changes) by lncRNA microarray
(16). However, its biological
functions in BC are largely unknown. To further investigate its
role in BC cells, we first detected the expression level of DUXAP10
in human normal bladder epithelial cell line SV-HUC-1 and different
BC cell lines. T24 and 5637 were then selected as a potential cell
model for further investigation. Specific siRNA oligos targeting
DUXAP10 (si-DUXAP10) were transfected to T24 and 5637 cells, and
cell proliferation, apoptosis and cell cycle changes were
observed.
Materials and methods
Cell culture
Human BC cell lines (5637, T24, E-j, TCCSUP,
UM-UC-3, RT4) and human normal bladder epithelial cell line
SV-HUC-1 were obtained from the Institute of Cell Research, Chinese
Academy of Sciences, Shanghai, China. The human BC cell lines and
normal bladder epithelial cell line were respectively cultured in
RPMI-1640 (5637, T24, E-j), MEM (TCCSUP, UM-UC-3), McCoy's 5A
Medium (RT4) (Gibco, Life Technologies, Carlsbad, CA, USA) and
F12-k medium (Gibco). All media were supplemented with 10% fetal
bovine serum (FBS; Gibco) and 1% penicillin-streptomycin (Gibco).
All cell lines were cultured in a humidified air atmosphere of 5%
CO2 at 37°C. Cell transfection was conducted by
Lipofectamine 2000.
Total RNA extraction and reverse
transcription
Total RNA from cells was extracted using TRIzol
reagent (Invitrogen, Karlsruhe, Germany) following the
manufacturer's instructions. Subsequently, total RNA was quantified
using a NanoDrop ND-2000 (Thermo Fisher Scientific, Wilmington, DE,
USA). cDNA was synthesized by reverse transcription (RT) using
random primers and the GoScript RT system (Promega, Madison, WI,
USA).
qRT-PCR detection of DUXAP10
The real-time quantitative reverse
transcription-polymerase chain reaction (qRT-PCR) was achieved by
using the GoTaq qPCR Master Mix (Promega) on a Mx3005P real-time
PCR system (Stratagene, La Jolla, CA, USA) following the
manufacturer's instructions. Primers for DUXAP10 and actin were
synthesized by Sangon Biotech (Shanghai, China). The primer
sequences used in the study are listed in Table I. The data were analyzed using the
∆Ct method. All results are expressed as the mean ± SEM of the
three independent experiments.
 | Table ISequences of primer and siRNA
(5′→3′). |
Table I
Sequences of primer and siRNA
(5′→3′).
| Name | Sense | Antisense |
|---|
| lncRNA-DUXAP10
primer |
GGCTGGAAGATTGCTTGAG |
GGTAGTGTGGTGCCTCCTGT |
| Actin primer |
CATGTACGTTGCTATCCAGGC |
CTCCTTAATGTCACGCACGAT |
| lncRNA-DUXAP10
siRNA |
GACUAUGCCUUCUGAAUAUTT |
AUAUUCAGAAGGCAUAGUCTT |
| Negative control
siRNA |
UUCUCCGAACGUGUCACGUTT |
ACGUGACACGUUCGGAGAATT |
Protein extraction and western blot
analysis
Western blot analysis was performed to analyze cell
cycle and apoptotic protein expression. The cells were collected
and lysed with cell lysis buffer for western blotting (Beyotime,
Haimen, China). The proteins (30 µg per lane) were separated on 12%
SDS-polyacrylamide gels and transferred onto polyvinylidene
fluoride (PVDF) membranes (Millipore, Billerica, MA, USA).
Immunoblotting of the membranes was performed using the following
primary antibodies: 1:1,000 rabbit anti-human phospho-Akt (Ser473),
Akt, P13K-p110a, phospho-mTOR, m-TOR, PTEN, cleaved caspase-3,
cleaved caspase-9, Bcl-xL, Bad, CDK4, cyclin E, p27 (Cell
Signaling, Danvers, MA, USA) and 1:1,000 mouse anti-human β-actin
(4A Biotech, Beijing, China). The signals were revealed after
incubation with the recommended secondary antibodies (1:4,000 goat
anti-rabbit IgG HRP-conjugated) (Transgen Biotech, Beijing, China)
using a Bio-Rad ChemiDOX XRS+ Imaging system (Bio-Rad Laboratories,
Inc., CA, USA).
RNA interference
siRNA oligos targeting DUXAP10 (si-DUXAP10) and
non-targeting siRNAs (negative control, si-NC) were designed and
synthesized from GenePharma (Shanghai, China). Three siRNAs were
designed in our pre-experiment (DUXAP10-home-957, DUXAP10-home-773
and DUXAP10-home-1080) and the transfection effect of
DUXAP10-home-957 was found to be the best. The sequences of the
si-DUXAP10 and the si-NC used are listed in Table I. The sequences of the two other
siRNAs are listed in Table II.
The cells were cultured in 6 well-plates, and were maintained in
reduced serum Opti-MEM medium (Gibco). siRNA transfections were
performed with 120 nM siRNA and 2 µl (per well) Lipofectamine 2000
(Gibco) following the manufacturer's instructions. After
transfection, the medium was removed and the cells were cultured in
RPMI-1640 and F12-k medium supplemented with 10% fetal bovine
serum.
| Table IISequences of siRNA (5′→3′). |
Table II
Sequences of siRNA (5′→3′).
| Name | Sense | Antisense |
|---|
| DUXAP10-home-773 |
GAGAAUGGGUCUAAAAGGAATT |
UUCCUUUAGACCCAUUCUCTT |
|
DUXAP10-home-1080 |
CCAUAUCCUGGUAAGGCUUTT |
AAGCCUUACCAUAUGGTT |
Cell proliferation
For the cell proliferation assay, cells were plated
in E-Plate 96 (ACEA Biosciences, San Diego, CA, USA), using a
Real-Time Cell Analyzer (RTCA) (ACEA Biosciences) according to the
manufacturer's instructions and the cell number was calculated by
TC10 Automated Cell Counter (Bio-Rad Laboratories, Inc.).
Cell cycle and apoptosis
The cells were washed with PBS and fixed in 75%
ice-cold ethanol at −20°C overnight. After rehydrating with
ice-cold PBS, the cells were stained with cell cycle staining
solution (MultiSciences Co., Ltd., Beijing, China) and analyzed by
flow cytometry, on a FACSCalibur flow cytometer (BD Biosciences)
using CellQuest Pro software. The cells were washed in PBS and
stained in Annexin V/PI apoptosis kit (MultiSciences Co., Ltd.) and
the rate of cell apoptosis was also analyzed by flow cytometry on a
FACSCalibur flow cytometer.
Statistical analysis
The data are presented as the mean ± SEM The
differences between groups were evaluated with two-tailed Student's
t-tests using SPSS Statistics 20.0 software (IBM, Armonk, NY, USA).
p<0.05 was considered to be statistically significant.
Results
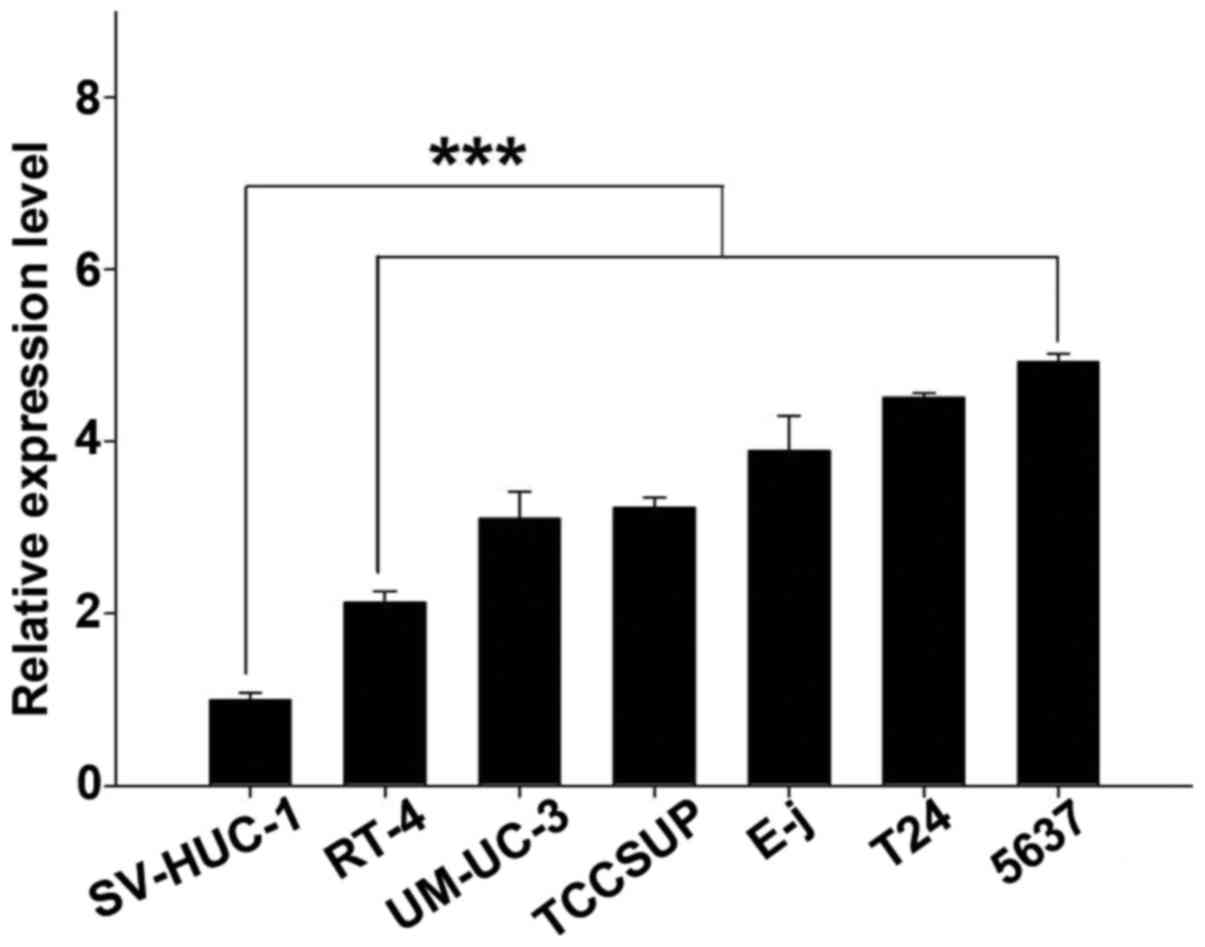
DUXAP10 is overexpressed in BC cells
The relative expression level of DUXAP10 was
evaluated in BC cells T24, 5637, E-j, TCCSUP, UM-UC-3, RT4, and
normal urothelial cell SV-HUC-1. Comparing with SV-HUC-1, the
expression of DUXAP10 is significantly upregulated in 5637
(493.0%), T24 (451.3%), E-j (389.3%), TCCSUP (323.5%), UM-UC-3
(310.9%) and RT4 (213.1%) cells (p<0.01) (Fig. 1), which provides evidence that
DUXAP10 may play a role in BC.
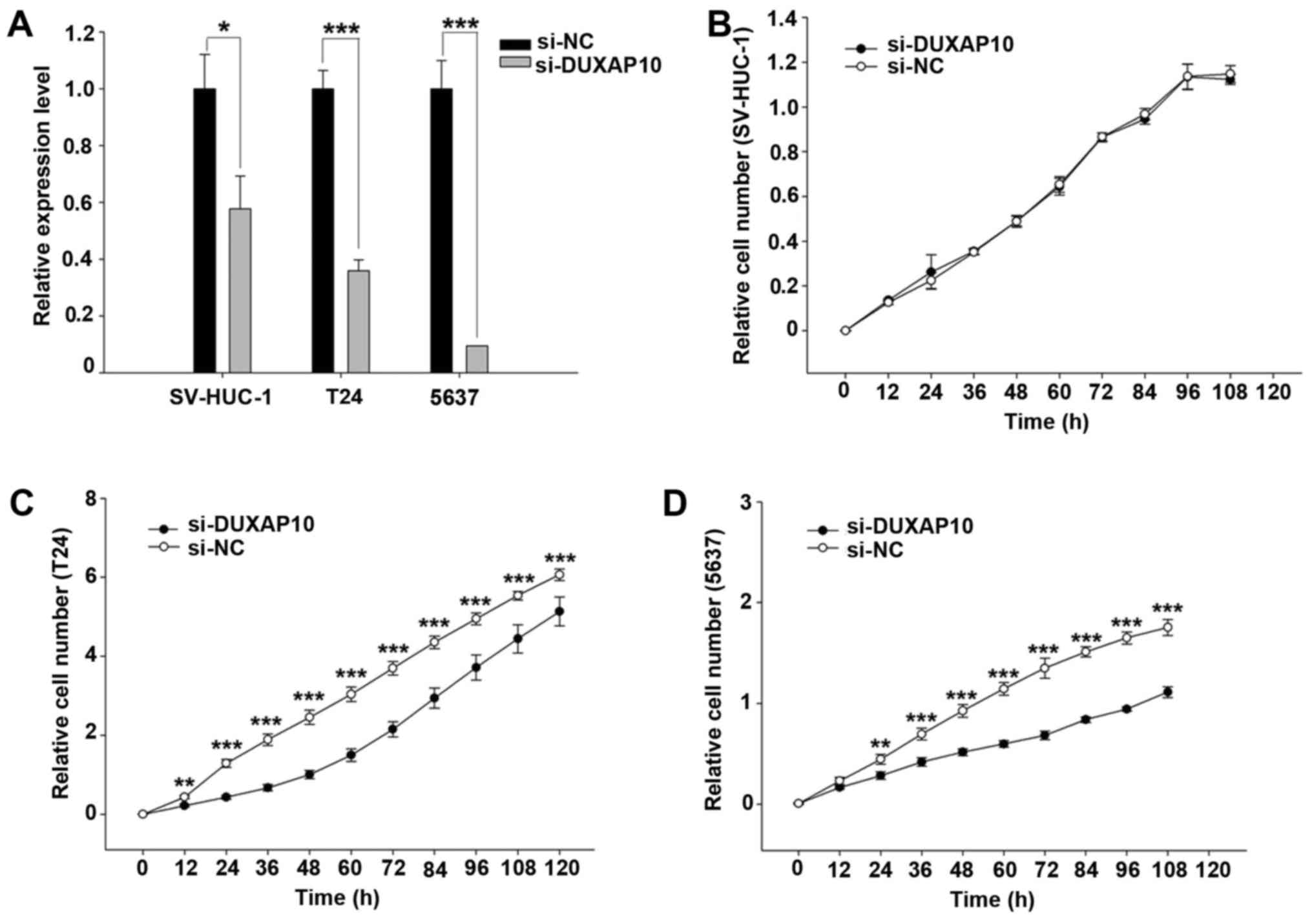
Knockdown of DUXAP10 inhibits cell
proliferation in human BC cells
To study the biological functions of DUXAP10 in BC
cells, we designed a siRNA to knock down DUXAP10 expression. As T24
and 5637 have highly expressed DUXAP10, we chose both T24 and 5637
as cell models for further investigation. After transfection with
si-DUXAP10, the expression of DUXAP10 is significantly reduced
comparing to transfection with si-NC (p<0.01) (Fig. 2A).
Inhibition of cell proliferation was observed in T24
and 5637 cells after transfection of si-DUXAP10 at 12 and 24 h,
respectively (p<0.01) (Fig. 2C and
D). Moreover, knockdown of DUXAP10 had no effect on cell
proliferation in the SV-HUC-1 cells (Fig. 2B). These results indicate that
DUXAP10 is associated with tumor cell proliferation.
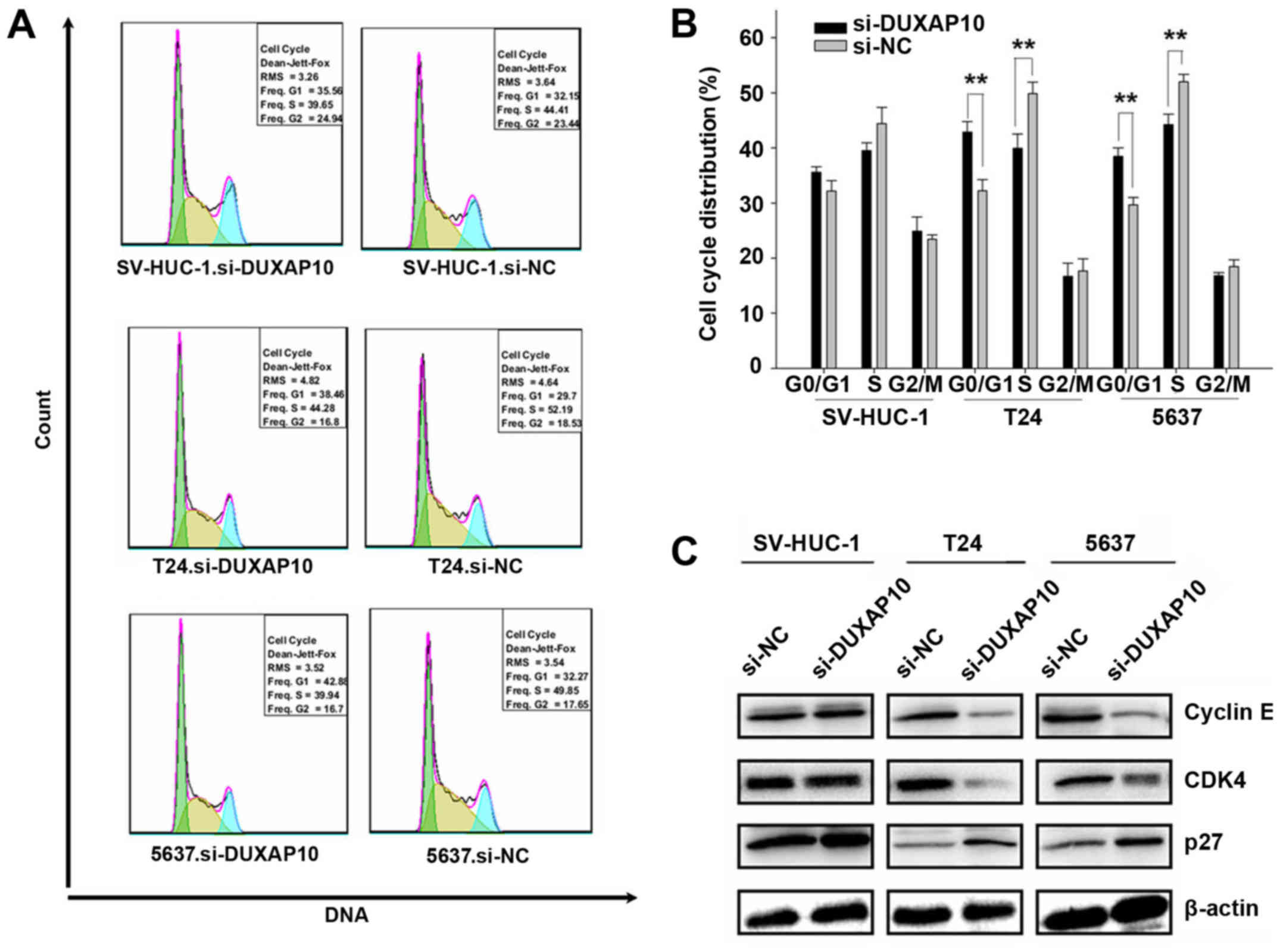
The effect of DUXAP10 knockdown on cell
cycle in human BC cells
Knockdown of DUXAP10 significantly affected cell
proliferation, so we decided to investigate the effects of DUXAP10
on the cell cycle. Flow cytometry demonstrated that DUXAP10
downregulation induced cell cycle arrest in G0/G1 in 5637 and T24
cells (p<0.01) (Fig. 3A and B).
These results indicated that the inhibitory effect on T24 and 5637
cell proliferation after knockdown of DUXAP10 may be taken through
restraining the G0/G1 to S phase transition.
According to the results obtained from the cell
cycle changes, we examined the effects of DUXAP10 on the expression
levels of cyclin E, CDK4 and p27. As shown in Fig. 3C, the p27 protein level increased,
while cyclin E and CDK4 protein level decreased by the knockdown of
DUXAP10 in T24 and 5637 cells. These results suggest that the G0/G1
cell cycle arrest induced by knockdown of DUXAP10 was related to
the downregulation of cyclin E and CDK4 and upregulation of the p27
protein.
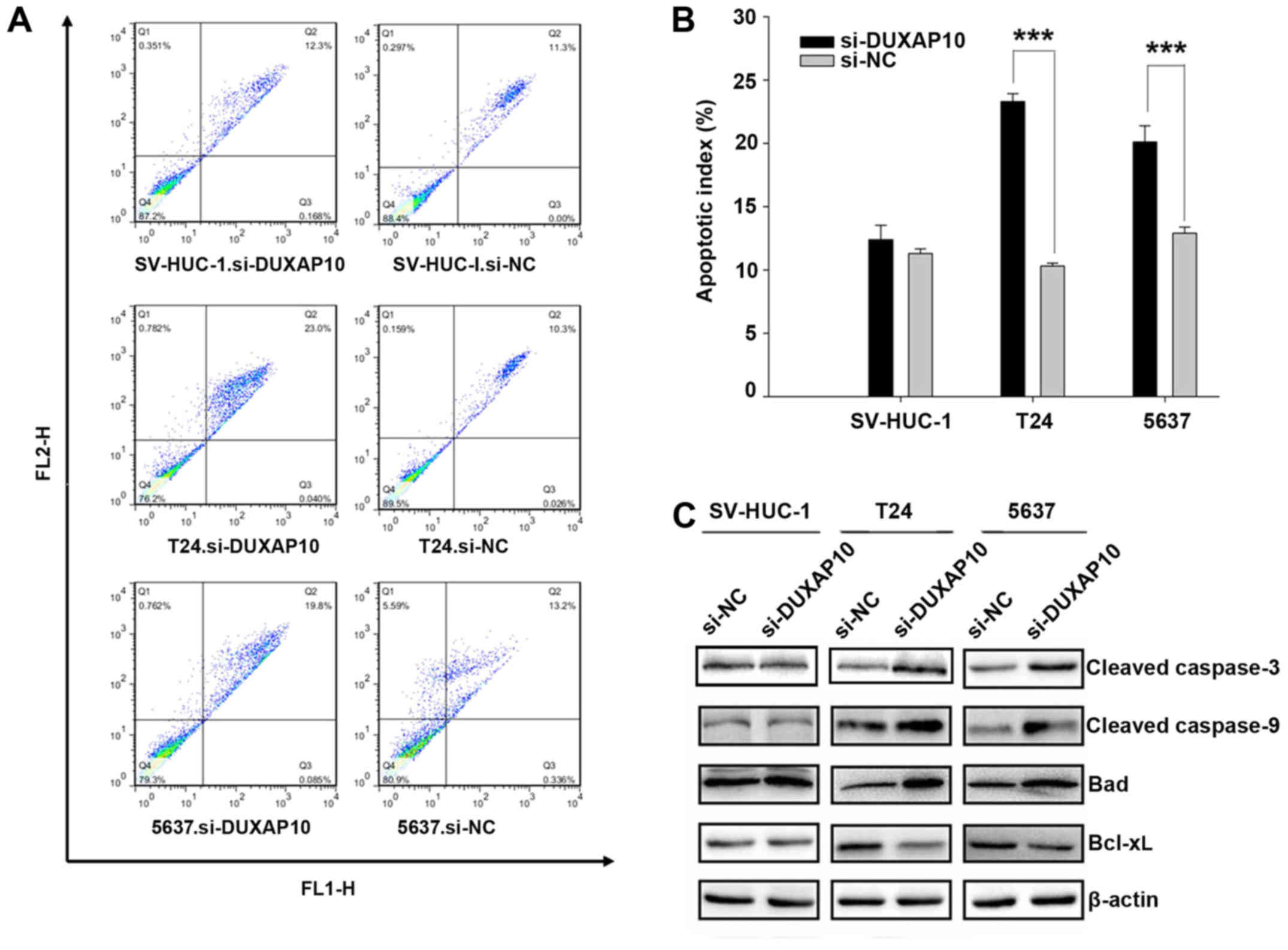
Knockdown of DUXAP10 promotes apoptosis
in human BC cells
To further study the biological function of DUXAP10,
we tested whether knockdown of DUXAP10 increases apoptosis in T24
and 5637 cells. The proportion of apoptotic cells after cell
transfection of si-DUXAP10 was higher compared with cells
transfection of si-NC (23.0 vs 10.3% and 20.1 vs 12.9%,
respectively) in T24 and 5637 cells (p<0.01) (Fig. 4A and B). These results indicate
that knockdown of DUXAP10 promotes cell apoptosis.
We also performed western blotting to detect caspase
activation (Fig. 4C). After
knockdown of DUXAP10 increased levels of cleaved caspase-3 and -9
expression were detected in the T24 and 5637 cells (Fig. 4C). In order to further test whether
DUXAP10 induced apoptosis through the activation of the
mitochondrial apoptotic pathway, we detected two major proteins
involved in this pathway. As shown in Fig. 4C, DUXAP10 knockdown led to an
increase in the expression of the pro-apoptotic protein Bad and a
decrease in the expression of anti-apoptotic protein Bcl-xL in T24
and 5637 cells. These results indicate that the inhibition of
DUXAP10 induces apoptosis through the mitochondrial apoptotic
pathway.
Knockdown of DUXAP10 regulates the
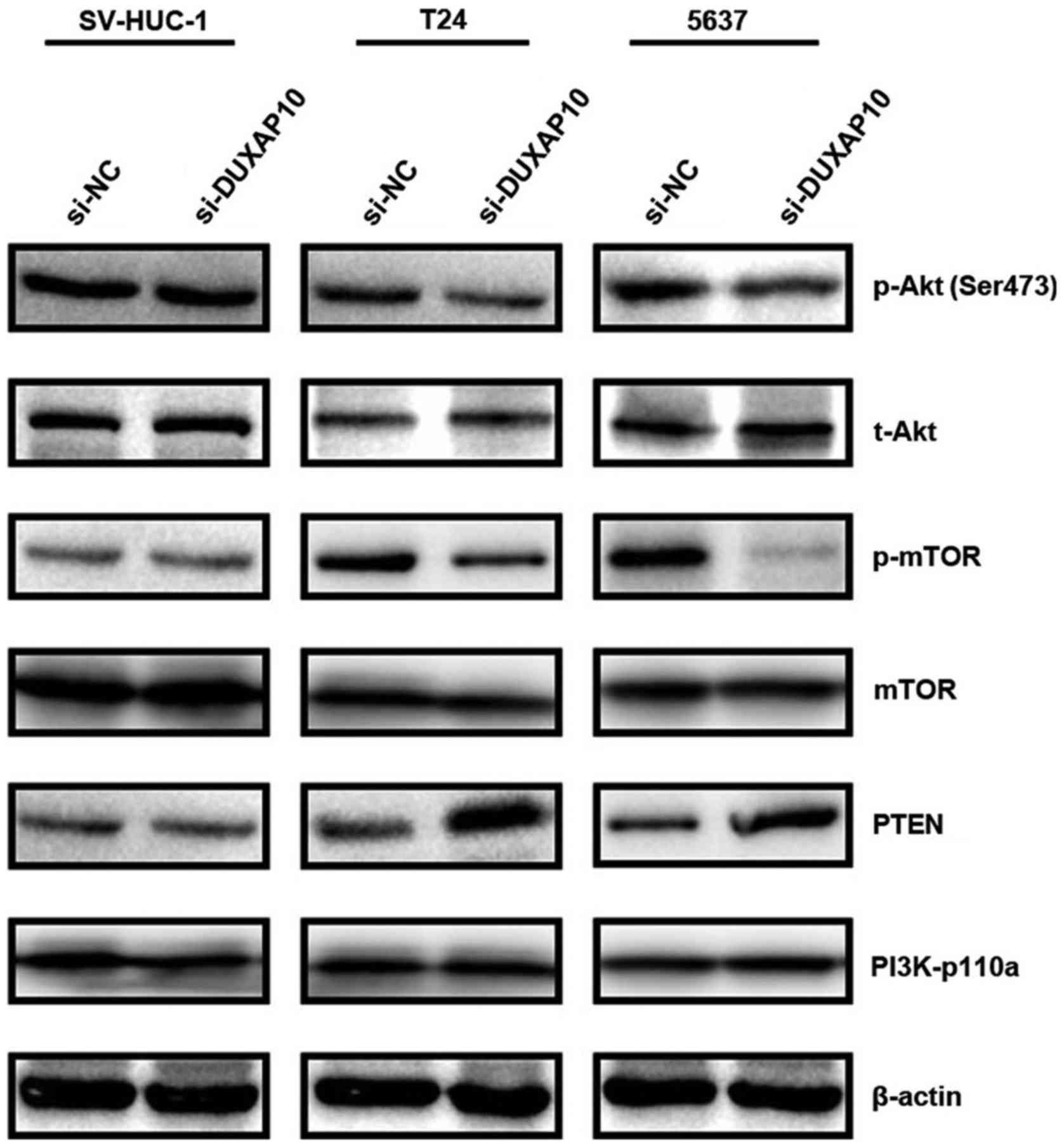
PI3K/Akt/mTOR signaling pathway in BC cells
To explore the potential mechanisms involved in
apoptosis and cell cycle arrest, we examined the effects of DUXAP10
on P13K/Akt/mTOR signaling pathways that contributes to the
development of BC. As shown in Fig.
5, the expression of PTEN was increased after knockdown of
DUXAP10 in T24 and 5637 cells. Then the phosphorylation levels of
Akt (Ser473) and mTOR were decreased by knockdown of DUXAP10 in T24
and 5637 cells. These results suggest that knockdown of DUXAP10
potentially exerts its antitumor function through interfering with
the PI3K/AKT/mTOR signaling pathway.
Discussion
Increasing studies are focusing on the effects of
lncRNAs in the diagnosis and treatment of cancers. With the
development of high-throughput DNA sequencing and array-based
technologies, various classes of lncRNAs have been identified
(16). Currently, lncRNAs have
been observed to regulate complex cellular behaviors and many of
them are implicated in the development and progression of cancer
(17). Dysregulation of lncRNAs,
such as HOTAIR, H19, MALAT-1, and PCA3, have been reported as a
primary feature of several human cancers including lung cancer,
breast cancer, colorectal cancer and prostate cancer (18). Thus, lncRNAs may reveal novel
mechanisms of tumorigenesis and progression, as well as present new
targets for the purposes of cancer therapy (19).
In the present study, we found that the expression
of DUXAP10 in the human BC cell lines 5637, T24, E-j, TCCSUP,
UM-UC-3, and RT4 was significantly higher compared to the human
normal bladder cell line SV-HUC-1 (Fig. 1). This result suggested that
DUXAP10 may be associated with BC. Therefore, we designed siRNA
oligos specifically targeting DUXAP10 to knock down DUXAP10 in BC
cells T24, 5637 (the expression of DUXAP10 was higher in the
selected cells than the others) and normal urothelial cells
(Fig. 2A).
After knockdown of the expression of DUXAP10, we
found that cell proliferation in BC cells T24 (12 h after
trans-fection) and 5637 (24 h after transfection) was inhibited but
had no significant effect on the SV-HUC-1 cells (Fig. 2B–D). Furthermore, apoptosis was
enhanced in T24 and 5637 after the transfection of si-DUXAP10
(Fig. 4A and B). DUXAP10 knockdown
increased the distribution of G0/G1 and decreased the distribution
of S phase in T24 and 5637 (Fig. 3A
and B). The results indicate that DUXAP10 is able to accelerate
proliferation through promoting the G0/G1 to S phase transition and
to depress apoptosis in BC cells T24 and 5637.
Alterations in the regulation of the cell cycle
progression and apoptosis are frequent events in human cancers. The
G1 cyclin-CDK complex cyclin E-CDK4 is required for S phase entry
(20). CDK inhibitors (CKIs) such
as p27KIP1 binds to cyclin-CDK complexes and renders
them inactive, resulting in the inhibition of cell cycle
progression (21). Our study
showed that the cyclin E and CDK4 levels dropped following
knockdown of DUXAP10. However, p27 levels were elevated. These
results suggest that DUXAP10 is associated with cell cycle
progression via downregulation of cyclin E and CDK4 while with
upregulation of p27 in human BC cells.
Apoptosis is a complex process regulated by a
balance of pro-apoptotic and anti-apoptotic genes (22,23).
Our results demonstrated that knockdown of DUXAP10 enhances
expression of cleaved caspase-9 and cleaved caspase-3. The
pro-apoptotic protein Bad was increased, while the anti-apoptotic
protein Bcl-xL was decreased. These results suggest that knockdown
of DUXAP10 induces apoptosis via dysregulation of Bad and Bcl-xL
proteins.
To explore the potential mechanisms underlying the
effects of DUXAP10, we evaluated the effect of DUXAP10 on the
cellular signaling pathways involved in the inhibition of apoptosis
and regulation of cell cycle arrest. Akt and mTOR are two
downstream effectors of PI3K and involved in several cellular
activities (24). Akt is activated
by the phosphorylation of Thr308 and Ser473 key regulatory sites
(25). The PI3K/Akt/mTOR pathway
is a critical pathway in cancer and activation of this pathway is
associated with tumor growth and therapeutic resistance (24,26).
In addition, PTEN is a lipid phosphatase that directly resists the
activity of PI3K (27). In our
study, western blot analysis showed that the phosphorylation of Akt
(Ser473) and phosphorylation of mTOR were decreased by knockdown of
DUXAP10 in both T24 and 5637 cells. Moreover, we observed that the
expression of PTEN was increased after knockdown of DUXAP10
(Fig. 5). Therefore, our data
confirmed that DUXAP10 inhibited BC cell proliferation and induced
cell apoptosis via PI3K/Akt/mTOR signaling pathway.
To the best of our knowledge, this is the first
study focusing on the analysis of DUXAP10 expression and its
biological functions in BC cell lines. We found that DUXAP10 was
overexpressed in the BC cell lines 5637, T24, E-j, TCCSUP, UM-UC-3
and RT4 when compared to normal bladder cells (Fig. 1). Knockdown of DUXAP10 inhibited
cell proliferation and enhanced cell apoptosis in T24 and 5637
cells. Therefore, we can conclude that DUXAP10 plays a role in BC
and we will investigate expression level of DUXAP10 in clinical
samples and its relationship with clinical pathology and prognosis
in our further study.
In conclusion, knockdown of DUXAP10 inhibits BC cell
proliferation and increases apoptosis. This finding contributes to
a better understanding of the importance of the dysregulated
lncRNAs in BC progression, which provides a rationale for the
potential development of lncRNA-based approaches for the treatment
of BC.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant nos. 81272828 for Q.M., 31501113
for R.Y. and 81402099 for Y.L.), Zhejiang Provincial Foundation for
Medical and Health Sciences (grant nos. 2016KYB263 and 2014KYB355
for Q.M. and 2017KY576 to J.F.C) and Natural Science Foundation of
Ningbo (2016A610163 for X-Y.L).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Clark PE, Spiess PE, Agarwal N, Bangs R,
Boorjian SA, Buyyounouski MK, Efstathiou JA, Flaig TW, Friedlander
T, Greenberg RE, et al: NCCN Guidelines Insights: Bladder Cancer,
Version 2.2016. J Natl Compr Cancer Netw. 14:1213–1224. 2016.
View Article : Google Scholar
|
|
4
|
Babjuk M, Burger M, Zigeuner R, Shariat
SF, van Rhijn BW, Compérat E, Sylvester RJ, Kaasinen E, Böhle A,
Palou Redorta J, et al European Association of Urology: EAU
guidelines on non-muscle-invasive urothelial carcinoma of the
bladder: Update 2013. Eur Urol. 64:639–653. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Abufaraj M, Gust K, Moschini M, Foerster
B, Soria F, Mathieu R and Shariat SF: Management of muscle
invasive, locally advanced and metastatic urothelial carcinoma of
the bladder: A literature review with emphasis on the role of
surgery. Transl Androl Urol. 5:735–744. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Witjes JA, Compérat E, Cowan NC, De Santis
M, Gakis G, Lebret T, Ribal MJ, Van der Heijden AG and Sherif A: AU
guidelines on muscle-invasive and metastatic bladder cancer:
summary of the 2013 guidelines. Eur Urol. 65:778–792. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bidnur S, Savdie R and Black PC:
Inhibiting immune checkpoints for the treatment of bladder cancer.
Bladder Cancer. 2:15–25. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Djebali S, Davis CA, Merkel A, Dobin A,
Lassmann T, Mortazavi A, Tanzer A, Lagarde J, Lin W, Schlesinger F,
et al: Landscape of transcription in human cells. Nature.
489:101–108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kong YW, Ferland-McCollough D, Jackson TJ
and Bushell M: microRNAs in cancer management. Lancet Oncol.
13:e249–e258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Enokida H, Yoshino H, Matsushita R and
Nakagawa M: The role of microRNAs in bladder cancer. Investig Clin
Urol. 57(Suppl 1): S60–S76. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen J, Miao Z, Xue B, Shan Y, Weng G and
Shen B: Long non-coding RNAs in urologic malignancies: Functional
roles and clinical translation. J Cancer. 7:1842–1855. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li HJ, Li X, Pang H, Pan JJ, Xie XJ and
Chen W: Long non-coding RNA UCA1 promotes glutamine metabolism by
targeting miR-16 in human bladder cancer. Jpn J Clin Oncol.
45:1055–1063. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu H, Li X, Song Y, Zhang P, Xiao Y and
Xing Y: Long non-coding RNA ANRIL is up-regulated in bladder cancer
and regulates bladder cancer cell proliferation and apoptosis
through the intrinsic pathway. Biochem Biophys Res Commun.
467:223–228. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li LJ, Zhu JL, Bao WS, Chen DK, Huang WW
and Weng ZL: Long noncoding RNA GHET1 promotes the development of
bladder cancer. Int J Clin Exp Pathol. 7:7196–7205. 2014.PubMed/NCBI
|
|
16
|
Zhu YP, Bian XJ, Ye DW, Yao XD, Zhang SL,
Dai B, Zhang HL and Shen YJ: Long noncoding RNA expression
signatures of bladder cancer revealed by microarray. Oncol Lett.
7:1197–1202. 2014.PubMed/NCBI
|
|
17
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bartonicek N, Maag JL and Dinger ME: Long
noncoding RNAs in cancer: Mechanisms of action and technological
advancements. Mol Cancer. 15:432016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jiang C, Li X, Zhao H and Liu H: Long
non-coding RNAs: Potential new biomarkers for predicting tumor
invasion and metastasis. Mol Cancer. 15:622016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sheppard KE and McArthur GA: The
cell-cycle regulator CDK4: An emerging therapeutic target in
melanoma. Clin Cancer Res. 19:5320–5328. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bertoli C, Skotheim JM and de Bruin RA:
Control of cell cycle transcription during G1 and S phases. Nat Rev
Mol Cell Biol. 14:518–528. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Um HD: Bcl-2 family proteins as regulators
of cancer cell invasion and metastasis: A review focusing on
mitochondrial respiration and reactive oxygen species. Oncotarget.
7:5193–5203. 2016. View Article : Google Scholar :
|
|
23
|
Gibson CJ and Davids MS: BCL-2 antagonism
to target the intrinsic mitochondrial pathway of apoptosis. Clin
Cancer Res. 21:5021–5029. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Guo H, German P, Bai S, Barnes S, Guo W,
Qi X, Lou H, Liang J, Jonasch E, Mills GB, et al: The PI3K/AKT
pathway and renal cell carcinoma. J Genet Genomics. 42:343–353.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nitulescu GM, Margina D, Juzenas P, Peng
Q, Olaru OT, Saloustros E, Fenga C, Spandidos DA, Libra M and
Tsatsakis AM: Akt inhibitors in cancer treatment: The long journey
from drug discovery to clinical use (Review). Int J Oncol.
48:869–885. 2016.
|
|
26
|
Lee JJ, Loh K and Yap YS: PI3K/Akt/mTOR
inhibitors in breast cancer. Cancer Biol Med. 12:342–354. 2015.
|
|
27
|
Engelman JA: Targeting PI3K signalling in
cancer: Opportunities, challenges and limitations. Nat Rev Cancer.
9:550–562. 2009. View
Article : Google Scholar : PubMed/NCBI
|