Introduction
Prostate cancer is one of the most frequently
diagnosed types of cancer and is the second leading cause of
cancer-related mortality in males (1,2). The
majority of prostate cancers begin in an androgen-dependent state,
and thus, hormonal therapy, including surgical castration and
medical/chemical castration has become the standard therapy.
However, following androgen deprivation therapy, the tumor
re-emerges in a more metastatic form that is no longer responsive
to this type of therapy and this cancer is then classified as
castration-resistant prostate cancer (CRPC) (3). At this stage, the cancer has become
incurable and the average survival time of patients is only 16–18
months (4). Moreover, therapeutic
strategies have had little effect on the progression of the
disease. Therefore, novel treatment strategies and new drug options
are urgently required for patients with CRPC.
Curcumin (diferuloylmethane), a polyphenol
phytochemical extracted from the plant, Curcuma longa, is
widely used in Southeast Asia, China and India in food preparation
and for medicinal purposes, such as health maintenance and cancer
prevention. Due to its anticancer and anti-angiogenic properties,
as well as its lack of toxicity and high affordability, curcumin
has been extensively studied as a chemotherapeutic agent (5–7). In
particular, curcumin has been shown to be cytotoxic towards both
the androgen-dependent prostate cancer cell line, LNCaP and the
androgen-independent cell lines, DU145 and PC3 (8–10).
However, various experimental studies and clinical trials have
shown that the systemic bioavailability of orally administered
curcumin is low. Serum concentrations often do not reach >0.1%
of intake and curumin exhibits rapid elimination in the feces, bile
and urine (11). In order to
overcome this limitation, different strategies, including the
design and synthesis of structural analogs have been explored. For
example, the novel synthetic curcumin analogue,
3,5-bis(2-fluorobenzylidene)-4-piperidone (EF24), has been shown to
exhibit improved anticancer properties in vitro and in
vivo compared to curcumin (12). It has been found that 10 µM
of EF24 is cytotoxic toward 70–80% of DU145 human prostate cancer
cells (12). Pyridine analogs of
curcumin with a tetrahydrothiopyrane-4-one linker, analogs FN1, FN2
and FN3, have also exhibited potent anticancer activities in PC3
cells showing a potent inhibitory effect on growth and a potent
stimulatory effect on apoptosis at low concentrations (≤1
µM) (13).
Previously, our group synthesized a series of
heterocyclic cyclohexanone analogs of curcumin and investigated
their inhibitory activity against nuclear factor (NF)-κB
trans-activation in non-adherent K562 leukemia cells, as well as
cytotoxicity toward ER-negative breast cancer cell lines (14). It has been found that curcumin
analogs exhibited potent cytotoxicity towards a variety of breast
cancer cell lines. In particular,
1-methyl-3,5-bis(4-pyridyl)methylidene-4-piperi-done (RL66) was
shown to exhibit EC50 values of 0.8, 0.5 and 0.6
µM, while
3,5-bis(3,4,5-trimethoxybenzylidene)-1-meth-ylpiperidine-4-one
(RL71) produced EC50 values of 0.3, 0.3 and 0.4
µM in MDA-MB-231, MDA-MB-468 and SKBr3 cells, respectively
(14). Moreover, both compounds
have been shown to significantly increase the number of cells
underoing apoptosis (15,16). Furthermore, both analogs have been
shown to decrease HER2/neu phosphorylation and increase p27 levels
in SKBr3 cells, while significantly decreasing AKT phosphorylation
and transiently increasing the stress kinases c-Jun N-terminal
kinase (JNK)1/2 and mitogen-activated protein kinase (MAPK) p38 in
MDA-MB-231 and MDA-MB-468 cells (15,16).
In the present study, we explored the cytotoxic effects and
anticancer mechanisms of a novel series of curcumin analogs towards
androgen-independent PC3 and DU145 prostate cancer cells. The
initial screening revealed that two curcumin analogs termed
1-isopropyl-3,5-bis(pyridin-3-ylmethylene)-4-piperidone (RL118) and
1-methyl-3,5-[(6-methoxynaphthalen-2-yl)methylene]-4-piperidone
(RL121) (Fig. 1) elicited potent
cytotoxicity towards the CRPC cell lines. Thus, these two compounds
were selected for further experimentation. The results demonstrated
that these two curcumin analogs induced cell cycle arrest and
apoptosis. Moreover, the results of western blot analysis
demonstrated that both analogs modulated the expression of key cell
signaling proteins involved in cell proliferation and cell death.
Thus, our findings provide evidence that RL118 and RL121 have
potent anticancer activity against CPRC cells, and both analogs
warrant further examination in vivo.
Materials and methods
Materials
The PC3 and DU145 prostate cancer cells and NIH/3T3
mouse embryonic fibroblasts were purchased from the American Type
Culture Collection (Manassas, VA, USA). PNT1A normal human prostate
epithelial cells were purchased from the European Collection of
Authenticated Cell Cultures which is operated by Public Health
England (Salisbury, UK). Dulbecco's modified Eagle's medium (DMEM)
nutrient mixture Ham's F-12, Eagle's Minimum Essential Medium
(EMEM), sulforhodamine B salt, propidium iodide (PI), ammonium
persulfate, horseradish peroxidase and β-tubulin (cat. no. T5293)
were purchased from Sigma-Aldrich (St. Louis, MO, USA). Epidermal
growth factor receptor (EGFR; cat. no. 2646), p-EGFR (cat. no.
4404), NF-κB (p65/RelA) (cat. no. 8242), cleaved caspase-3 (cat.
no. 9664), eukaryotic translation initiation factor 4E-binding
protein 1 (4E-BP1; cat. no. 9452), p-4E-BP1 (cat. no. 9455),
mammalian target of rapamycin (mTOR; cat. no. 2972) p-mTOR (cat.
no. 2971) primary antibodies were purchased from Cell Signaling
Technology (Danvers, MA, USA). AKT (cat. no. 559028) and p-AKT
(cat. no. 4058) primary antibodies were purchased from BD
Biosciences (Auckland, New Zealand). Goat anti-rabbit horseradish
peroxidase (cat. no. 401353) and goat anti-mouse horseradish
peroxidase (cat. no. 401253) secondary antibodies were purchased
from Merck (Billerica, MA, USA). Acrylamide, bisacrylamide, sodium
dodecylsulfate and PVDF membrane were purchased from Bio-Rad
Laboratories (Hercules, CA, USA). Complete Mini EDTA-free protease
inhibitor cocktail and Annexin-V-FLUOS were purchased from Roche
Diagnostics Corp. (Mannheim, Germany). RNAse A, bovine serum
albumin (BSA) and trypsin were purchased from Invitrogen (Auckland,
New Zealand). All synthetic curcumin analogs were synthesized by Dr
Bill Hawkins from the Department of Chemistry at the University of
Otago, Dunedin, New Zealand.
Synthesis of curcumin derivatives
1-isopropyl-3,5-bis[(pyridine-3-yl)methylene]piperidin-4-one
(RL118) was synthesized as follows: To a solution of
1-isopropylpiperidin-4-one (1.4 g, 10 mmol) and
pyridine-3-carboxaldehyde (2.14 g, 20 mmol) in methanol (10 ml) was
added a solution of sodium methoxide in methanol (5 M, 1.5 ml) and
the mixture was stirred for 18 h. The resultant yellow crystalline
solid was removed by filtration then recrystallized from methanol
to yield pure
1-isopropyl-3,5-bis[(pyridine-3-yl)methylene]piper-idin-4-one
as a yellow crystalline solid (1.45 g, 45%). HRMS (+ve ESI)
calculated for
C20H21N3ONa+: 342.1577
m/z. [MNa+] found 342.1567 m/z.
1-methyl-3,5-bis[(6-methoxy-2-naphthyl)
methylene]piperidin-4-one (RL121) was synthesized as follows: To a
solution of the N-methyl-4-piperidone (0.50 ml, 0.46 g, 4.05
mmol) and 6-methoxy-2-naphthaldehyde (1.51 g, 8.10 mmol) in
methanol (10 ml) was added a solution of sodium methoxide in
methanol (5 M, 1.0 ml) and the mixture was stirred for 16 h. The
resultant solid was collected via filtration to afford the title
compound (1.35 g, 74%) as a light brown crystalline solid.
1H NMR (400 MHz, CDCl3), δ (ppm): 8.05, bs,
2H; 7.81–7.76, m, 6H; 7.48, dd, 2H, J=8.2, 1.5 Hz; 7.20, dd, 2H,
J=8.4, 2.5 Hz), 7.15, d, 2H, J=2.5 Hz; 4.02, bs, 2H; 3.07, m, 2H;
3.95, s, 6H; 2.55, s, 3H. HRMS-ESI calculated for
C30H28NO3+ [M+H]+:
449.1991; found: 449.1996.
Cell maintenance
The PC3 prostate cancer cells were cultured in
DMEM/Ham's F-12 supplemented with 10% FBS, 1% L-glutamine, 1%
penicillin/streptomycin and the DU145 cells were maintained in MEM
supplemented with 10% fetal bovine serum (FBS), 1% L-glutamine, 1%
penicillin/streptomycin. The cells were kept in a humidified
incubator at 37°C with 5% CO2.
Cytotoxicity
The prostate cancer cells, PC3 and DU145, and the
normal human prostate epithelial cells, PNT1A, were seeded in
96-well plates at a rate of 4,000 and 6,000 cells per well,
respectively and allowed to adhere to the plate for 24 h at 37°C.
The cells were then treated with curcumin analogs with a range of
concentrations (0.1–10 µM) of each compound for 72 h. The
vehicle control cells were treated with dimethyl sulfoxide (DMSO)
(0.1%). The cell number in each well was determined by the
sulforhodamine B (SRB) assay (14). Cytotoxicity time course studies
were conducted by treating the PC3 and DU145 cells with compounds
RL118 or RL121 at 0.5, 0.75, 1, 1.5 or 2 µM for 12–72 h.
Data are expressed as the number of viable cells determined from 3
independent experiments conducted in triplicate.
Cell cycle analysis
The PC3 (1.0×105 cells per well) and
DU145 (2.0×105 cells per well) cells were seeded in
6-well plates in 2 ml of complete medium and allowed to adhere to
the plate for 24 h. The cells were then treated with RL118 (1
µM for PC3 cells and 2 µM for DU145 cells) or RL121
(1 µM for both cell lines) using 0.1% DMSO as a control for
24 and 48 h. The cells were then harvested, washed with
phosphate-buffered saline (PBS) and fixed in 70% ethanol. Following
rehydration with PBS, the cells were stained with PI in the dark at
4°C as previously described (17).
The samples were analyzed via flow cytometry using a BD FACSCalibur
flow cytometer (BD Biosciences, Auckland, New Zealand). Data were
acquired and analyzed using FlowJo software. Data are expressed as
the mean proportion of cells in each phase ± SEM determined from 3
independent experiments conducted in triplicate.
Induction of apoptosis
The percentage of PC3 and DU145 cells undergoing
apoptosis following treatment with RL118 and RL121 was determined
by flow cytometry. The PC3 cells (1.0×105 cells per
well) and DU145 cells (2.0×105 cells per well) were
seeded in 6-well plates with 2 ml of complete medium and allowed to
adhere to the plate for 24 h. The cells were then treated with
RL118 (1 µM for the PC3 cells and 2 µM for the DU145
cells) or RL121 (1 µM for both cell lines) for 24 and 48 h.
The vehicle control cells were treated with 0.1% DMSO. Apoptosis
was assessed using Annexin-V-FLUOS/PI (Roche Diagnostics Corp.)
staining, as previously described (18). The samples were analyzed via flow
cytometry using a BD FACSCalibur flow cytometer. Data were acquired
and analyzed using FlowJo software. Values are expressed as the
number of apoptotic cells as a % of the total number of cells ± SEM
determined from 3 independent experiments conducted in
triplicate.
Western blot analysis
The PC3 cells (1.0×106 cells per dish)
and DU145 cells (2.0×106 cells per dish) were seeded in
10-cm cell culture dishes in 10 ml of complete medium and allowed
to adhere to the plate for 24 h. The cells were then treated with
RL118 (1 µM for the PC3 cells and 2 µM for the DU145
cells), RL121 (1 µM for both cell lines) and the vehicle
control (0.1% DMSO) for 24 and 48 h. At the end of treatment, whole
cell lysates were prepared and the protein concentration of the
lysates was determined using the bicinchoninic acid (BCA) assay as
previously described (17). Cell
lysates were resolved on a SDS-PAGE gel (1 µg of protein per
lane) and transferred onto a PVDF membrane. Protein levels were
analyzed with the desired primary antibodies, followed by
horseradish peroxidase-conjugated secondary antibodies (Merck).
Digital chemiluminescence images were acquired using a VersaDoc
(Bio-Rad Laboratories) imaging system and All-PRO Imaging system
and quantified using Quantity One software (Bio-Rad
Laboratories).
Statistical analysis
When time was a factor, data were analyzed with a
two-way ANOVA coupled with a Bonferroni post-hoc test, where
p<0.05 was required for a statistically significant difference.
When time was not a factor, data were analyzed with a one-way ANOVA
coupled with Bonferroni post-hoc test, where p<0.05 was required
for a statistically significant difference.
Results
In total, 9 different curcumin analogs namely,
RL109, RL110, RL114, RL115, RL116, RL117, RL118, RL120 and RL121
were screened for their cytotoxicity towards the PC3 cells. A total
of 5 curcumin analogs elicited EC50 values within the
range of 0.5–100 µM, while RL120 was unable to elicit an
EC50 value over the range of concentrations tested
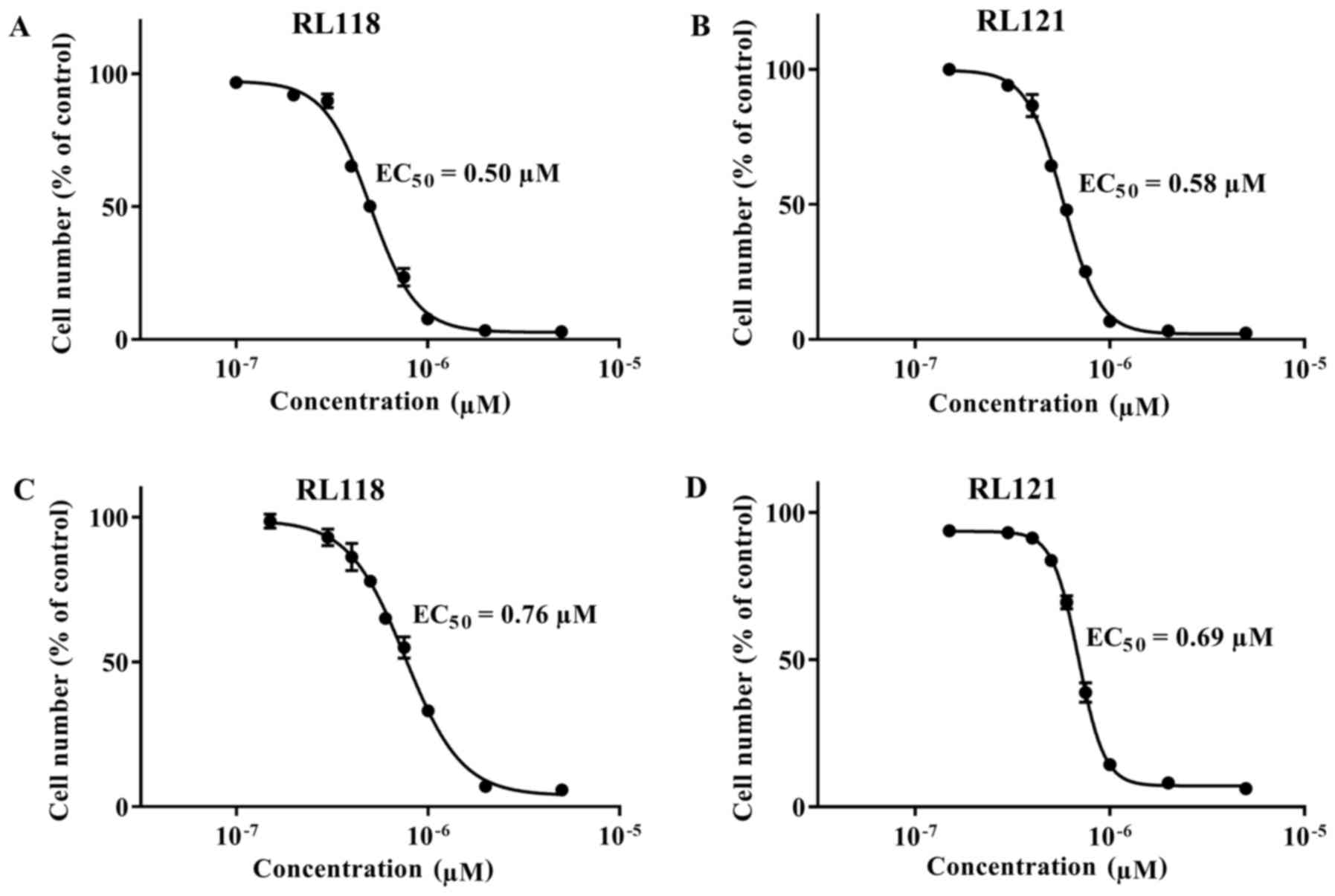
(Table I). Since RL118 and RL121
were the most potent compounds towards the PC3 cells (Fig. 2A and B), they were also examined in
the DU145 cells, where they exhibited equivalent cytotoxicity
(Fig. 2C and D). To determine the
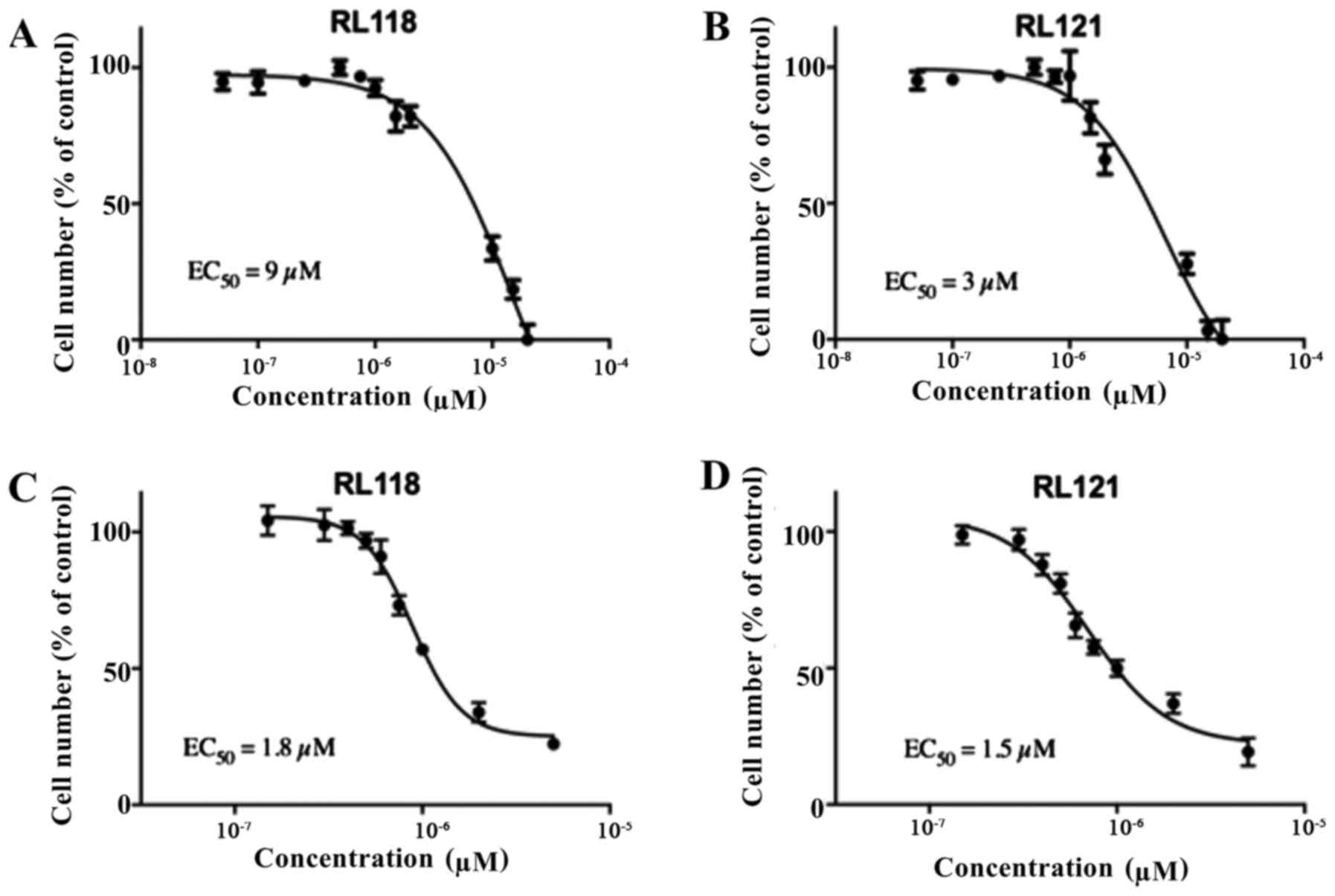
selectivity of RL118 and RL121 towards cancer cells, the effects of
these compounds on PNT1A non-cancerous prostate cells and NIH/3T3
mouse embryonic fibroblasts were examined. The results revealed
that RL118 had EC50 values of 9 µM in the NIH/3T3
cells and 1.8 µM in the PNT1A cells (Fig. 3). For RL121, the EC50
values were 3 µM in the NIH/3T3 cells and 1.5 µM in
the PNT1A cells (Fig. 3). Thus,
the two compounds were 3- to 18-fold less potent towards
non-cancerous cells compared to the PC3 cells. In order to further
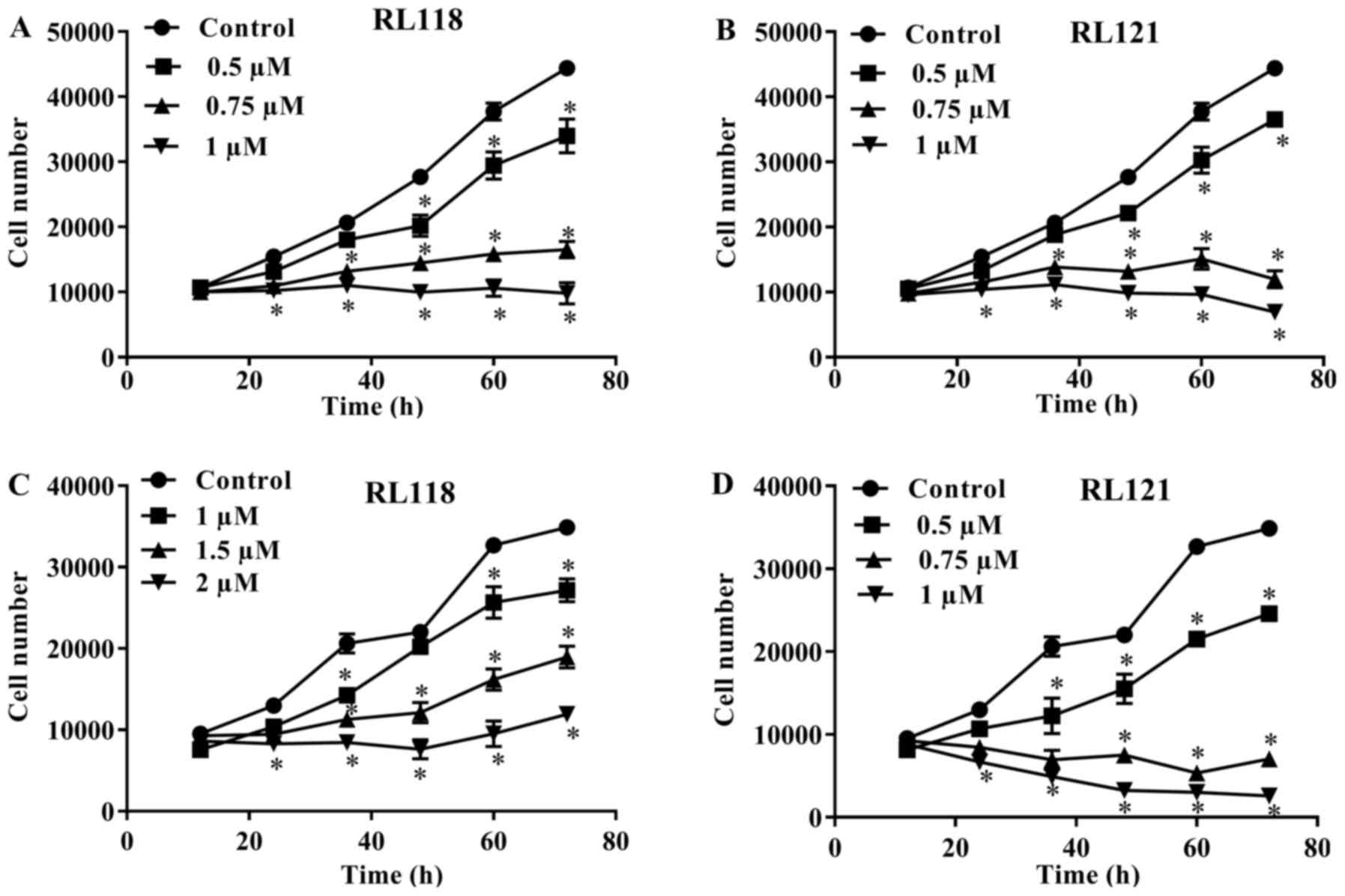
investigate the cytotoxicity of the two curcumin analogs, the PC3
and DU145 cells were treated with various concentrations of RL118
and RL121 over a time-course. Time- and concentration-dependent
effects were observed for both cell lines. Treatment of the PC3 and
DU145 cells with both analogs delayed the growth of cells beginning
from 12 h, and this differed significantly compared to the control
after 48 h (Fig. 4). The DU145
cells were more resistant to the effects of RL118 as higher
concentrations were required to delay cell growth and no
concentration was cytotoxic (Fig.
4C).
 | Table IEC50 values of curcumin
analogs in PC3 cells. |
Table I
EC50 values of curcumin
analogs in PC3 cells.
| Curcumin
analogs | EC50
(µM)a |
|---|
| RL109 | >100 |
| RL110 | 2.66±0.48 |
| RL114 | 3.09±0.30 |
| RL115 | 6.70±8.52 |
| RL116 | 12.88±10 |
| RL118 | 0.50±0.05 |
| RL117 | 1.09±0.13 |
| RL120 | NDb |
| RL121 | 0.53±0.03 |
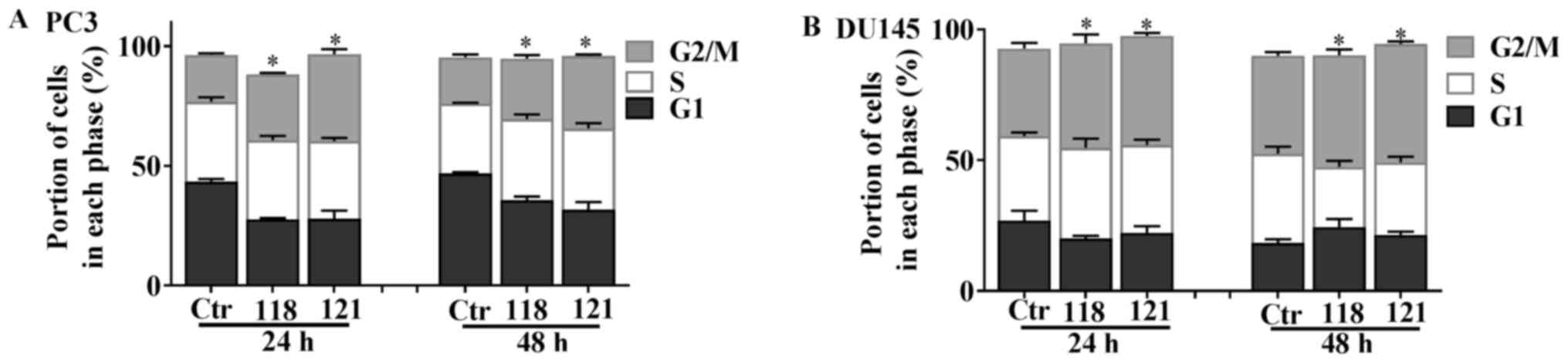
Cell cycle analysis by flow cytometry was then
performed in order to examine whether the cytotoxicity of RL118 and
RL121 was driven by cell cycle arrest. The results revealed that
both RL118 and RL121 resulted in a cell line-, time- and
dose-dependent cell cyce arrest. Both analogs significantly induced
cell cycle arrest in the G2/M phase in the PC3 and DU145 cell
lines. Specifically, both RL118 and RL121 led to G2/M phase arrest
at 24 and 48 h in the PC3 cells at 1 µM. Specifically, RL121
induced a significant 86% increase at 24 h and a 57% increase at 48
h, while RL118 caused a significant increase by 42% at 24 h and a
30% increase at 48 h in the number of PC3 cells in G2/M phase
arrest compared to the controls (Fig.
5A). Treatment of the DU145 cells with 2 µM of RL118 and
1 µM of RL121 exhibited a differential cell cycle effect at
both time-points (Fig. 5B). RL118
induced a significant increase in the number of cells in the G2/M
phase by 18% at 24 h and 14% at 48 h, compared to the controls.
RL121 caused a significant increase in the number of cells in the
G2/M phase by 24 and 21% at 24 and 48 h, respectively, compared to
the controls.
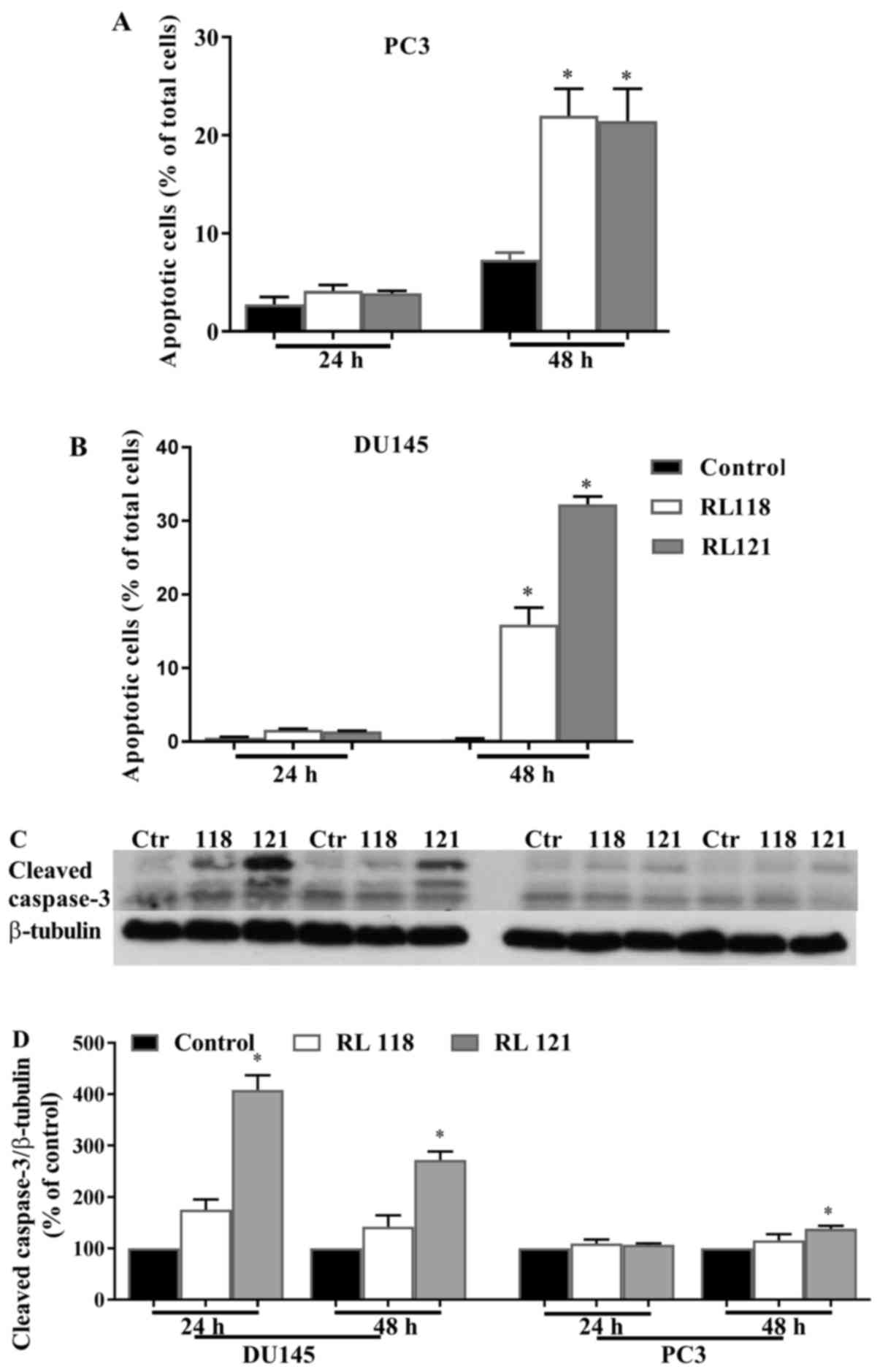
The ability of RL118 and RL121 to elicit the
apoptotic cell death of CRPC cells was then examined. The results
revealed that both RL118 and RL121 induced the apoptosis of the PC3
and DU145 cells in a time- and cell line-dependent manner.
Specifically, RL118 and RL121 signficantly increased the number of
apoptotic cells in both cell lines compared to the controls after
48 h (Fig. 6). Moreover, this
effect was more potent in the DU145 cells, as >30% of the total
number of cells was apoptotic following treatment with RL121 (1
µM) (Fig. 6B), compared to
20% of the PC3 cells. Apoptosis was also analyzed by quantifying
the changes in the levels of cleaved caspase-3 by western blot
analysis. Significantly elevated protein levels of cleaved
caspase-3 were only elicited by RL121 (1 µM) and this effect
was strongest in the DU145 cells after 24 h of treatment (Fig. 6C and D). Since the activation of
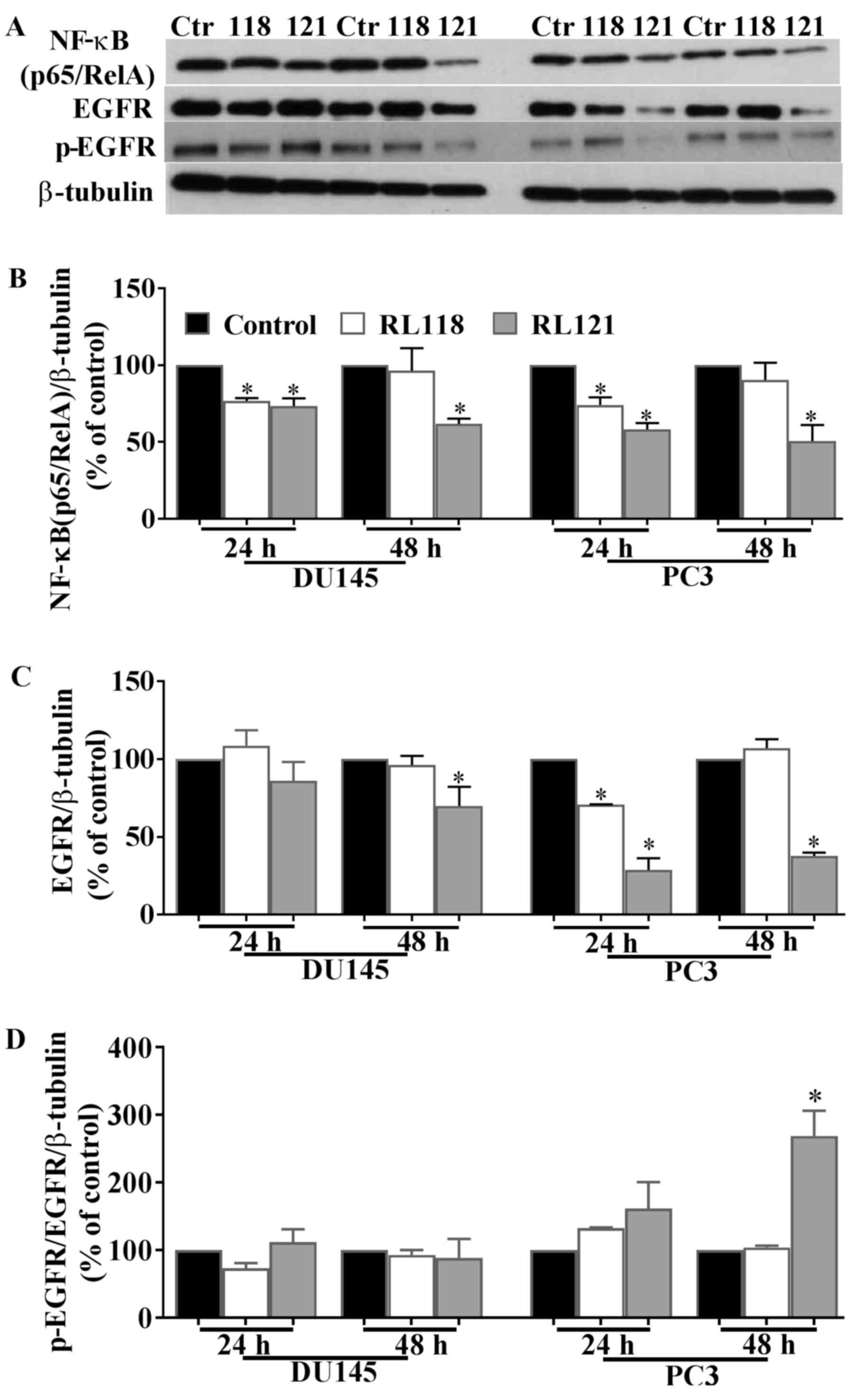
NF-κB leads to cell survival and proliferation, we examined the
effects of the two compounds on the expression of this protein.
RL121 (1 µM) significantly decreased NF-κB (p65/RelA)
expression by 28 and 42% in the DU145 and PC3 cells, respectively
after 24 h and this effect continued for 48 h (Fig. 7A and B). A similar effect was
observed with RL118 except that after 48 h, no significant change
in the expression of NF-κB (p65/RelA) was observed in either cell
line.
To account for these differences between RL118 and
RL121, we examined changes in the protein expression of EGFR. The
results revealed that the PC3 cells were the most susceptible to
the effects of RL121, as EGFR expression was decreased by 71 and
62% after 24 and 48 h, respectively, while the ratio of p-EGFR/EGFR
was significantly increased (Fig.
7). Of note, in the DU145 cells, this relative increase in
p-EGFR did not occur and this is likely due to the fact that DU145
cells express phosphatase and tensin homolog (PTEN), whereas PC3
cells lack this protein (19).
Further analysis of the impact that changes in PTEN expression may
have on these cells were then conducted by examining changes in the
levels of AKT, mTOR and 4E-BP1. Since no changes in EGFR expression
were observed after 24 h in the DU145 cells, further investigations
focused on treatment after 48 h in this cell line, while
experiments with PC3 cells utilized both the 24 and 48 h
time-points.
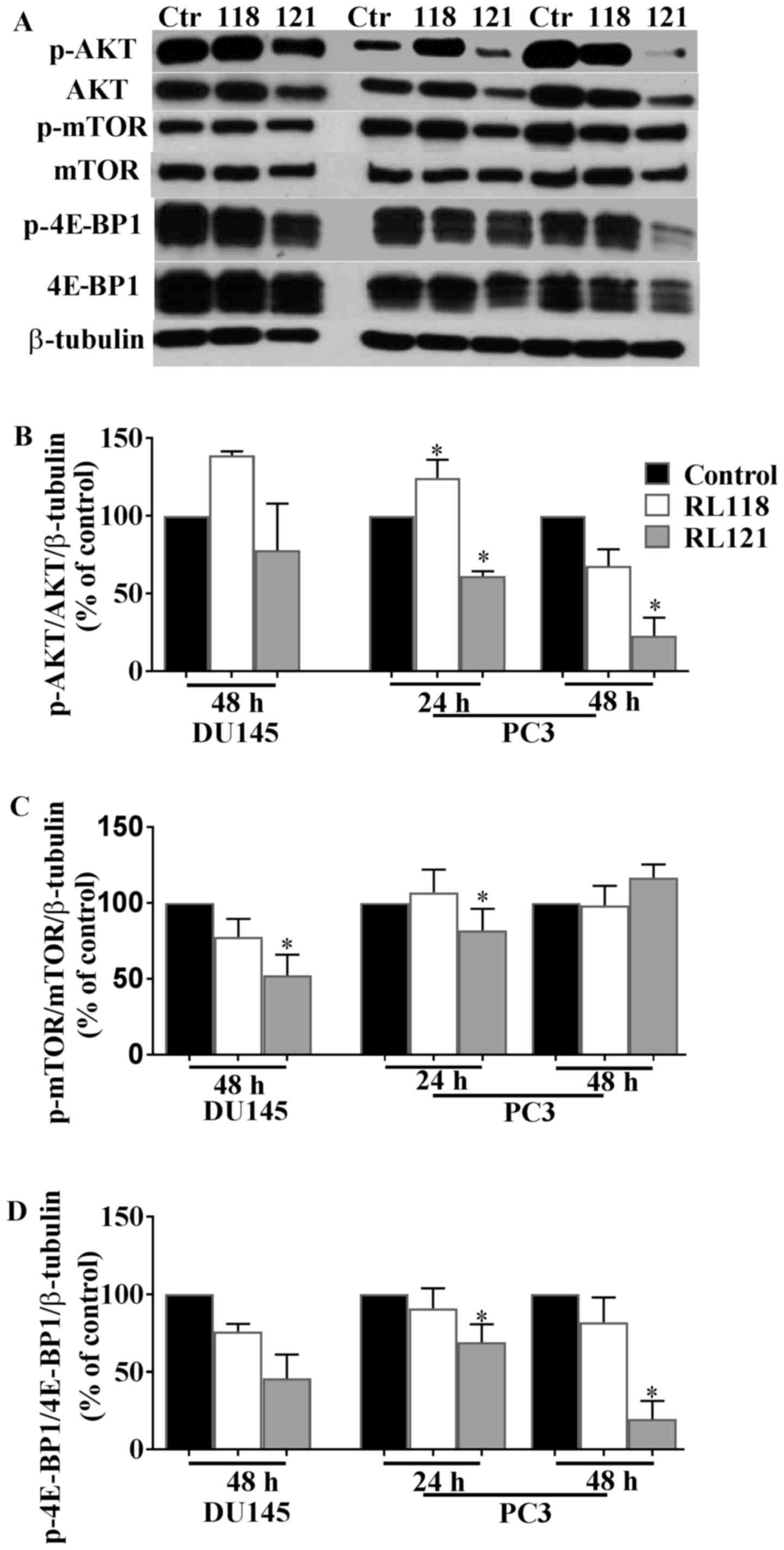
An important cell signaling hub downstream of the
EGFR is AKT and thus this was examined as the next mechainstic
protein. The results revealed that the PC3 cells were the most
susceptible to the effects of RL121, as this compound directly
inhibited AKT beginning at 24 h (Fig.
8A and B). Specifically, the ratio of p-AKT/AKT significantly
decreased by 39 and 73% compared to the controls at 24 and 28 h,
respectively (Fig. 8A and B). The
DU145 cells were unresponsive to treatment and RL118 was unable to
significantly inhibit AKT regardless of the cell line examined. AKT
activation leads to the phosphorylation of mTOR, which subsequently
phosphorylates 4E-BP1. Therefore, western blot analysis was carried
out to examine the effects of RL118 and RL121 on the expression of
mTOR, p-mTOR, 4E-BP1 and phosphorylated 4E-BP1 in the PC3 and DU145
cell lines. Of note, mTOR protein expression was unaltered in the
PC3 cells in contrast to their prior high susceptibility to AKT.
The only significant decrease in the ratio of p-mTOR/mTOR was
observed in the DU145 cells following treatment with RL121 for 48
h, where a decrease of 48% compared to the controls was observed
(Fig. 8A and C). This change
correlated with a 55% decrease in the ratio of p4E-BP1/4E-BP1 in
the DU145 cells compared to the controls following 48 h of
treatment with RL121 (Fig. 7A and
D). The PC3 cells exhibited the most significant decrease in
the ratio of p4E-BP1/4E-BP1, where RL121 significantly decreased
the expression of p-4E-BP1/4E-BP1 by 81% compared to the controls
after 48 h. Overall, of the two curcumin derivatives, RL121
produced a stronger and more consistent effect on cell
signaling-related proteins in the PC3 cells.
Discussion
In this study, in order to identify potent novel
potential drugs for the treatment of CRPC, 9 different curcumin
analogs were evaluated for their cytotoxicity towards the PC3
cells. While RL118 and RL121 exhibited the most potent cytotoxicity
towards the PC3 cells, these two analogs exhibited a slightly lower
potency in the DU145 cells. This may be due in part to the fact
that PC3 cells have a higher metastatic potential than DU145 cells.
Additioanlly, time-course experiments revealed that RL121 was
cytotoxic, while RL118 was cytostatic. It has been reported that
curcumin analogs containing a cyclohexanone or
N-methylpiperidone core exhibit enhanced potency compared to
other curcumin analogs in PC3 and DU145 cells (20). Indeed, we have previously found
that 2,6-bis(pyridin-4-ylmethylene)-cyclohexanone (RL91), which
contains a cyclohexanone core, exhibited high potency in PC3 and
DU145 cells with EC50 values of 2.4 and 2.1 µM,
respectively (21). However, RL118
and RL121 were more potent than RL91 in both cell lines. This may
be due to the fact that both RL118 and RL121 incorporate an
N-methylpiperidone core. In our previous study, on the
structure-activity relationship of curcumin analogs, the
N-methylpiperidone analogs
3,5-bis(pyridine-4-yl)-1-methylpiperidin-4-one (RL66) and
3,5-bis(3,4,5-trimethoxybenzylidene)-1-methylpiperidin-4-one (RL71)
exhibited potent cytotoxicity with EC50 values of 0.3
and 0.8 µM in the MDA-MB-231 cells, respectively (14). While in the same cell line, the
EC50 values of cyclohexanone containing analogs were
>1 µM (14). RL118 and
RL121 also elicited more potent cytotoxicity in CRPC cells compared
to the amono-carbonyl containing derivative, WZ35 (22) and the mono-ketone derivatives
FLLL11 and FLLL12 (23).
RL121 was a more potent inducer of G2/M-phase arrest
in both cell lines than RL118. G2/M phase cell cycle arrest is a
common response following curcumin and many of its analogs
(15,16,21–24)
and this arrest often is not the driver of apoptosis. It does
however, lead to a sustained and potent apoptotoic effect (12,13).
This was also shown by RL121, as >25% of the cell population of
both PC3 and DU145 cells was apoptotic by 48 h. Apoptosis, in
contrast to necrosis, is characterized as active and programmed
cell death with distinct morphological changes via energy-dependent
biochemical mechanisms (25). The
loss or suppression of apoptosis correlates with the progression of
prostate cancer; therefore, the induction of apoptosis has been
considered an effective therapeutic approach for the treatment of
prostate tumors (26). Not
surprisingly, RL118 and RL121 were both more effective at inducing
apoptosis than other curcumin derivatives, such as WZ35 (22), compound 17 (27), EF24 (12), A2 and A4 (28), as all required concentrations
between 2.5–20 µM in order to significantly increase the
number of apoptotic CRPC cells or significantly increase the
activity of caspase-3.
As an important class of transcriptional regulators,
NF-κB plays a pivotal role in the regulation of cell growth,
apoptosis, angiogenesis and metastasis (29–31).
The overexpression of NF-κB has been observed in prostate cancer
cell lines and clinically (32–34).
Activation of NF-κB in the nucleus of prostate cancer cells was
associated with chemoresistance, PSA recurrence and metastatic
spread of prostate cancer (35–37).
Furthermore, the inhibition of NF-κB activity is required to induce
the apoptosis of both PC3 and DU145 cells (38). Moreover, a recent study showed that
the activation of NF-κB signaling in prostate cancer cells
increased the expression of osteoclastogenic genes which
subsequently resulted in bone metastases formation (39). Therefore, targeting NF-κB may prove
to be a potent therapeutic target for CRPC. A previous study
revealed that transfected PC3 cells in which NF-κB activity was
blocked, produced slow-growing tumors with a low metastatic
potential (40). RL121 was more
consistent at inhibiting NF-κB (p65/RelA) protein expression in
CRPC cells and again was more potent that other analogs, such as
EF24 (41) and EF31 (42), both of which contain the same
piperidone core as RL118 and RL121.
EGFR plays a key role in the proliferation,
migration and invasion of tumor cells in prostate tumor progression
(43). In particular, EGFR is
frequently overexpressed in prostate cancer and is associated with
a poor prognosis (44). For
example, the frequency of EGFR expression increased from 41% in
patients with localized disease to 100% in patients with
castrate-resistant metastatic prostate cancer (45). Previously, combination experiments
with curcumin (25 µM) and β-phenylethyl isothiocyanate
(PEITC) (10 µM) significantly suppressed p-EGFR (Y1068)
protein by 86% of the control in PC3 cells (46). In this study, treatment with RL121
for 48 h significantly inhibited EGFR expression, but increased the
ratio of p-EGFR/EGFR in the PC3 cells, but not the DU145 cells.
This may be due to the fact that PC3 cells are PTEN-negative. PTEN
controls endocytic trafficking of EGFR by promoting late endosome
maturation and is required for the transition of ligand-bound EGFR
from early to late endosomes. The depletion of PTEN delays EGFR
trafficking and degradation (47).
The slower kinetics of receptor degradation will likely affect the
rates of EGFR and p-EGFR differentially as p-EGFR cycling is
activated upon ligand binding which directs the internalization of
these receptors to early endosome to the multivesicular body and
then on to the lysosome. When PTEN is present, the accumulation of
EGFRs on the limiting membrane of the endosomes is sufficient to
produce EGFR-dependent apoptosis (48). The deletion of PTEN delays this
process and may uncouple it from apoptosis-induced EGFR
degradation. Hence, this is likely the reason for the relative
increase in p-EGFR following RL121 treatment in PC3 cells at 48 h
compared to DU145-treated cells.
The serine-threonine kinase AKT is an important
regulator of protein synthesis and cell cycle progression (49,50).
Accumulating evidence has indicated that overexpressed AKT and
p-AKT activity has been associated with prostate cancer progression
(51–56). The combination of curcumin (25
µM) and PEITC (10 µM) has been shown to inhibit the
expression of AKT and levels of p-AKT in PC3 cells (46). Moreover, the curcumin analogue,
FLLL12, has been shown to inhibit p-AKT expression in PC3 cells
(23). RL121 is a more potent and
direct inhibitor of AKT, as 1 µM significantly decreased the
ratio of p-AKT/AKT in PC3 cells. This is a critical action of RL112
as, when PTEN is absent, there is an increase in the signaling
through both the RAS/RAF/MEK/ERK and AKT/mTOR pathways (57) and thus these pathways would
dominate. Therefore, the direct inhibition of AKT will be an
important driver of cell death in PC3 cells and will also drive the
decrease observed in NF-κB (p65/RelA) following RL112
treatment.
mTOR, a member of the PI3K kinase superfamily, plays
a key role in the regulation of protein synthesis, cell growth,
proliferation, differentiation and survival (58). In a previous study,
immunohistochemical analysis revealed that the expression of mTOR
and cytoplasmic p-mTOR was significantly increased in prostate
cancer tissue compared to the normal prostatic epithelium with mTOR
levels in cancer cells being 2-fold higher than those in benign
tissue (59). In this study, RL118
had no effect on mTOR levels; however, by contrast, RL121 decreased
the ratio of p4E-BP1/4E-BP1 protein levels by 55% in DU145 cells
and 81% in PC3 cells. It is not likely that RL121 inhibited 4E-BP1
directly as in the DU145 cells, the 55% decrease in the ratio of
p4E-BP1/4E-BP1 levels correlated with a 48% decrease in the ratio
of p-mTOR/mTOR, most likely directed through the inhibition of AKT.
As stated previously in PC3 cells, when PTEN is absent, there is an
increase in signaling through the RAS/RAF/MEK/ERK pathways as well
as the AKT/mTOR pathway (57). It
is likely that ERK can act directly on 4E-BP1 and indirectly via
TSC2/mTOR, as following ionizing radiation, it has been shown to
stimulate protein synthesis via ATM-dependent ERK phosphorylation
(60,61). Thus, in PC3 cells, the ratio of
p-4E-BP1/4E-BP1 changes independently of changes in mTOR.
Therefore, RL121 has several advantages in PC3 cells as it can
modulate changes in AKT directly and also decrease the in ratio of
p4E-BP1/4-EBP1 in an environment of highly active mTOR. Of the 9
novel curcumin analogs screened, RL118 and RL121 exhibited the most
potent anticancer activity in the PC3 and DU145 cell lines. While
both analogs warrant further examination in vivo, RL121 has
a wider range of activity in prostate cancers that lack PTEN, and
may thus be considered to be the lead compound to emerge from this
study.
References
|
1
|
Greenlee RT, Murray T, Bolden S and Wingo
PA: Cancer statistics, 2000. CA Cancer J Clin. 50:7–33. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Ward E, Brawley O and Jemal A:
Cancer statistics, 2011: The impact of eliminating socioeconomic
and racial disparities on premature cancer deaths. CA Cancer J
Clin. 61:212–236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Karantanos T, Corn PG and Thompson TC:
Prostate cancer progression after androgen deprivation therapy:
Mechanisms of castrate resistance and novel therapeutic approaches.
Oncogene. 32:5501–5511. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marques RB, Dits NF, Erkens-Schulze S, van
Weerden WM and Jenster G: Bypass mechanisms of the androgen
receptor pathway in therapy-resistant prostate cancer cell models.
PLoS One. 5:e135002010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shakibaei M, Mobasheri A, Lueders C, Busch
F, Shayan P and Goel A: Curcumin enhances the effect of
chemotherapy against colorectal cancer cells by inhibition of NF-κB
and Src protein kinase signaling pathways. PLoS One. 8:e572182013.
View Article : Google Scholar
|
|
6
|
Sadzuka Y, Nagamine M, Toyooka T, Ibuki Y
and Sonobe T: Beneficial effects of curcumin on antitumor activity
and adverse reactions of doxorubicin. Int J Pharm. 432:42–49. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chuang SE, Kuo ML, Hsu CH, Chen CR, Lin
JK, Lai GM, Hsieh CY and Cheng AL: Curcumin-containing diet
inhibits diethylnitrosamine-induced murine hepatocarcinogenesis.
Carcinogenesis. 21:331–335. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Strimpakos AS and Sharma RA: Curcumin:
Preventive and therapeutic properties in laboratory studies and
clinical trials. Antioxid Redox Signal. 10:511–545. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Khor TO, Keum YS, Lin W, Kim JH, Hu R,
Shen G, Xu C, Gopalakrishnan A, Reddy B, Zheng X, et al: Combined
inhibitory effects of curcumin and phenethyl isothiocyanate on the
growth of human PC-3 prostate xenografts in immunodeficient mice.
Cancer Res. 66:613–621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dorai T, Gehani N and Katz A: Therapeutic
potential of curcumin in human prostate cancer-I. curcumin induces
apoptosis in both androgen-dependent and androgen-independent
prostate cancer cells. Prostate Cancer Prostatic Dis. 3:84–93.
2000. View Article : Google Scholar
|
|
11
|
Anand P, Kunnumakkara AB, Newman RA and
Aggarwal BB: Bioavailability of curcumin: Problems and promises.
Mol Pharm. 4:807–818. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Adams BK, Cai J, Armstrong J, Herold M, Lu
YJ, Sun A, Snyder JP, Liotta DC, Jones DP and Shoji M: EF24, a
novel synthetic curcumin analog, induces apoptosis in cancer cells
via a redox-dependent mechanism. Anticancer Drugs. 16:263–275.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wei X, Zhou D, Wang H, Ding N, Cui XX,
Wang H, Verano M, Zhang K, Conney AH, Zheng X, et al: Effects of
pyridine analogs of curcumin on growth, apoptosis and NF-κB
activity in prostate cancer PC-3 cells. Anticancer Res.
33:1343–1350. 2013.PubMed/NCBI
|
|
14
|
Yadav B, Taurin S, Rosengren RJ,
Schumacher M, Diederich M, Somers-Edgar TJ and Larsen L: Synthesis
and cytotoxic potential of heterocyclic cyclohexanone analogues of
curcumin. Bioorg Med Chem. 18:6701–6707. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yadav B, Taurin S, Larsen L and Rosengren
RJ: RL66 a second-generation curcumin analog has potent in vivo and
in vitro anticancer activity in ER-negative breast cancer models.
Int J Oncol. 41:1723–1732. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yadav B, Taurin S, Larsen L and Rosengren
RJ: RL71, a second-generation curcumin analog, induces apoptosis
and downregulates Akt in ER-negative breast cancer cells. Int J
Oncol. 41:1119–1127. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Somers-Edgar TJ, Taurin S, Larsen L,
Chandramouli A, Nelson MA and Rosengren RJ: Mechanisms for the
activity of heterocyclic cyclohexanone curcumin derivatives in
estrogen receptor negative human breast cancer cell lines. Invest
New Drugs. 29:87–97. 2011. View Article : Google Scholar
|
|
18
|
Stuart EC, Jarvis RM and Rosengren RJ: In
vitro mechanism of action for the cytotoxicity elicited by the
combination of epigallocatechin gallate and raloxifene in
MDA-MB-231 cells. Oncol Rep. 24:779–785. 2010.PubMed/NCBI
|
|
19
|
Beresford SA, Davies MA, Gallick GE and
Donato NJ: Differential effects of
phosphatidylinositol-3/Akt-kinase inhibition on apoptotic
sensitization to cytokines in LNCaP and PCc-3 prostate cancer
cells. J Interferon Cytokine Res. 21:313–322. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Samaan N, Zhong Q, Fernandez J, Chen G,
Hussain AM, Zheng S, Wang G and Chen QH: Design, synthesis, and
evaluation of novel heteroaromatic analogs of curcumin as
anti-cancer agents. Eur J Med Chem. 75:123–131. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mazumder A, Gould M, Taurin S, Nicholson H
and Rosengren R: Raloxifene potentiates the cytotoxicity induced by
RL91, a second generation curcumin analog, in PC3 prostate cancer
cells. Toxicologist. 132:1462013.
|
|
22
|
Zhang X, Chen M, Zou P, Kanchana K, Weng
Q, Chen W, Zhong P, Ji J, Zhou H, He L, et al: Curcumin analog WZ35
induced cell death via ROS-dependent ER stress and G2/M cell cycle
arrest in human prostate cancer cells. BMC Cancer. 15:8662015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lin L, Hutzen B, Ball S, Foust E, Sobo M,
Deangelis S, Pandit B, Friedman L, Li C, Li PK, et al: New curcumin
analogues exhibit enhanced growth-suppressive activity and inhibit
AKT and signal transducer and activator of transcription 3
phosphorylation in breast and prostate cancer cells. Cancer Sci.
100:1719–1727. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chiu TL and Su CC: Curcumin inhibits
proliferation and migration by increasing the Bax to Bcl-2 ratio
and decreasing NF-kappaBp65 expression in breast cancer MDA-MB-231
cells. Int J Mol Med. 23:469–475. 2009.PubMed/NCBI
|
|
25
|
Wong RS: Apoptosis in cancer: From
pathogenesis to treatment. J Exp Clin Cancer Res. 30:872011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Khan N, Adhami VM and Mukhtar H: Apoptosis
by dietary agents for prevention and treatment of prostate cancer.
Endocr Relat Cancer. 17:R39–R52. 2010. View Article : Google Scholar
|
|
27
|
Mandalapu D, Saini KS, Gupta S, Sharma V,
Yaseen Malik M, Chaturvedi S, Bala V, Hamidullah, Thakur S,
Maikhuri JP, et al: Synthesis and biological evaluation of some
novel triazole hybrids of curcumin mimics and their selective
anticancer activity against breast and prostate cancer cell lines.
Bioorg Med Chem Lett. 26:4223–4232. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei X, Du ZY, Cui XX, Verano M, Mo RQ,
Tang ZK, Conney AH, Zheng X and Zhang K: Effects of cyclohexanone
analogues of curcumin on growth, apoptosis and NF-κB activity in
PC-3 human prostate cancer cells. Oncol Lett. 4:279–284.
2012.PubMed/NCBI
|
|
29
|
Oliver KM, Taylor CT and Cummins EP:
Hypoxia. Regulation of NFkappaB signalling during inflammation: The
role of hydroxylases. Arthritis Res Ther. 11:2152009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Walmsley SR, Print C, Farahi N,
Peyssonnaux C, Johnson RS, Cramer T, Sobolewski A, Condliffe AM,
Cowburn AS, Johnson N, et al: Hypoxia-induced neutrophil survival
is mediated by HIF-1α-dependent NF-kappaB activity. J Exp Med.
201:105–115. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Maxwell PJ, Gallagher R, Seaton A, Wilson
C, Scullin P, Pettigrew J, Stratford IJ, Williams KJ, Johnston PG
and Waugh DJ: HIF-1 and NF-kappaB-mediated upregulation of CXCR1
and CXCR2 expression promotes cell survival in hypoxic prostate
cancer cells. Oncogene. 26:7333–7345. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Suh J, Payvandi F, Edelstein LC, Amenta
PS, Zong WX, Gélinas C and Rabson AB: Mechanisms of constitutive
NF-kappaB activation in human prostate cancer cells. Prostate.
52:183–200. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Abdulghani J, Gu L, Dagvadorj A, Lutz J,
Leiby B, Bonuccelli G, Lisanti MP, Zellweger T, Alanen K, Mirtti T,
et al: Stat3 promotes metastatic progression of prostate cancer. Am
J Pathol. 172:1717–1728. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lindholm PF, Bub J, Kaul S, Shidham VB and
Kajdacsy-Balla A: The role of constitutive NF-kappaB activity in
PC-3 human prostate cancer cell invasive behavior. Clin Exp
Metastasis. 18:471–479. 2000. View Article : Google Scholar
|
|
35
|
Lessard L, Bégin LR, Gleave ME, Mes-Masson
AM and Saad F: Nuclear localisation of nuclear factor-kappaB
transcription factors in prostate cancer: An immunohistochemical
study. Br J Cancer. 93:1019–1023. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Domingo-Domenech J, Oliva C, Rovira A,
Codony-Servat J, Bosch M, Filella X, Montagut C, Tapia M, Campás C,
Dang L, et al: Interleukin 6, a nuclear factor-kappaB target,
predicts resistance to docetaxel in hormone-independent prostate
cancer and nuclear factor-kappaB inhibition by PS-1145 enhances
docetaxel antitumor activity. Clin Cancer Res. 12:5578–5586. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ross JS, Kallakury BV, Sheehan CE, Fisher
HA, Kaufman RP Jr, Kaur P, Gray K and Stringer B: Expression of
nuclear factor-κ B and IκBα proteins in prostatic adenocarcinomas:
Correlation of nuclear factor-κB immunoreactivity with disease
recurrence. Clin Cancer Res. 10:2466–2472. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chakraborty M, Qiu SG, Vasudevan KM and
Rangnekar VM: Par-4 drives trafficking and activation of Fas and
Fasl to induce prostate cancer cell apoptosis and tumor regression.
Cancer Res. 61:7255–7263. 2001.PubMed/NCBI
|
|
39
|
Jin R, Sterling JA, Edwards JR, DeGraff
DJ, Lee C, Park SI and Matusik RJ: Activation of NF-kappa B
signaling promotes growth of prostate cancer cells in bone. PLoS
One. 8:e609832013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Huang S, Pettaway CA, Uehara H, Bucana CD
and Fidler IJ: Blockade of NF-kappaB activity in human prostate
cancer cells is associated with suppression of angiogenesis,
invasion, and metastasis. Oncogene. 20:4188–4197. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang CH, Yue J, Sims M and Pfeffer LM: The
curcumin analog EF24 targets NF-κB and miRNA-21, and has potent
anticancer activity in vitro and in vivo. PLoS One. 8:e711302013.
View Article : Google Scholar
|
|
42
|
Olivera A, Moore TW, Hu F, Brown AP, Sun
A, Liotta DC, Snyder JP, Yoon Y, Shim H, Marcus AI, et al:
Inhibition of the NF-κB signaling pathway by the curcumin analog,
3,5-Bis(2-pyridinylmethylidene)-4-piperidone (EF31):
Anti-inflammatory and anti-cancer properties. Int Immunopharmacol.
12:368–377. 2012. View Article : Google Scholar
|
|
43
|
Jorissen RN, Walker F, Pouliot N, Garrett
TP, Ward CW and Burgess AW: Epidermal growth factor receptor:
Mechanisms of activation and signalling. Exp Cell Res. 284:31–53.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nicholson RI, Gee JM and Harper ME: EGFR
and cancer prognosis. Eur J Cancer. 37(Suppl 4): S9–S15. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Di Lorenzo G, Tortora G, D'Armiento FP, De
Rosa G, Staibano S, Autorino R, D'Armiento M, De Laurentiis M, De
Placido S, Catalano G, et al: Expression of epidermal growth factor
receptor correlates with disease relapse and progression to
androgen-independence in human prostate cancer. Clin Cancer Res.
8:3438–3444. 2002.PubMed/NCBI
|
|
46
|
Kim JH, Xu C, Keum YS, Reddy B, Conney A
and Kong AN: Inhibition of EGFR signaling in human prostate cancer
PC-3 cells by combination treatment with β-phenylethyl
isothiocyanate and curcumin. Carcinogenesis. 27:475–482. 2006.
View Article : Google Scholar
|
|
47
|
Shinde SR and Maddika S: PTEN modulates
EGFR late endocytic trafficking and degradation by
dephosphorylating Rab7. Nat Commun. 7:106892016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Rush JS, Quinalty LM, Engelman L, Sherry
DM and Ceresa BP: Endosomal accumulation of the activated epidermal
growth factor receptor (EGFR) induces apoptosis. J Biol Chem.
287:712–722. 2012. View Article : Google Scholar :
|
|
49
|
Dillon RL, White DE and Muller WJ: The
phosphatidyl inositol 3-kinase signaling network: Implications for
human breast cancer. Oncogene. 26:1338–1345. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
51
|
Graff JR, Konicek BW, McNulty AM, Wang Z,
Houck K, Allen S, Paul JD, Hbaiu A, Goode RG, Sandusky GE, et al:
Increased AKT activity contributes to prostate cancer progression
by dramatically accelerating prostate tumor growth and diminishing
p27Kip1 expression. J Biol Chem. 275:24500–24505. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Shimizu Y, Segawa T, Inoue T, Shiraishi T,
Yoshida T, Toda Y, Yamada T, Kinukawa N, Terada N, Kobayashi T, et
al: Increased Akt and phosphorylated Akt expression are associated
with malignant biological features of prostate cancer in Japanese
men. BJU Int. 100:685–690. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Morgan TM, Koreckij TD and Corey E:
Targeted therapy for advanced prostate cancer: Inhibition of the
PI3K/Akt/mTOR pathway. Curr Cancer Drug Targets. 9:237–249. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ayala G, Thompson T, Yang G, Frolov A, Li
R, Scardino P, Ohori M, Wheeler T and Harper W: High levels of
phosphorylated form of Akt-1 in prostate cancer and non-neoplastic
prostate tissues are strong predictors of biochemical recurrence.
Clin Cancer Res. 10:6572–6578. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kreisberg JI, Malik SN, Prihoda TJ,
Bedolla RG, Troyer DA, Kreisberg S and Ghosh PM: Phosphorylation of
Akt (Ser473) is an excellent predictor of poor clinical outcome in
prostate cancer. Cancer Res. 64:5232–5236. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mimeault M, Johansson SL and Batra SK:
Pathobiological implications of the expression of EGFR, pAkt, NF-κB
and MIC-1 in prostate cancer stem cells and their progenies. PLoS
One. 7:e319192012. View Article : Google Scholar
|
|
57
|
Song MS, Salmena L and Pandolfi PP: The
functions and regulation of the PTEN tumour suppressor. Nat Rev Mol
Cell Biol. 13:283–296. 2012.PubMed/NCBI
|
|
58
|
Pópulo H, Lopes JM and Soares P: The mTOR
signalling pathway in human cancer. Int J Mol Sci. 13:1886–1918.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kremer CL, Klein RR, Mendelson J, Browne
W, Samadzedeh LK, Vanpatten K, Highstrom L, Pestano GA and Nagle
RB: Expression of mTOR signaling pathway markers in prostate cancer
progression. Prostate. 66:1203–1212. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liu G, Zhang Y, Bode AM, Ma WY and Dong Z:
Phosphorylation of 4E-BP1 is mediated by the p38/MSK1 pathway in
response to UVB irradiation. J Biol Chem. 277:8810–8816. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Haystead TA, Haystead CM, Hu C, Lin TA and
Lawrence JC Jr: Phosphorylation of PHAS-I by mitogen-activated
protein (MAP) kinase. Identification of a site phosphorylated by
MAP kinase in vitro and in response to insulin in rat adipocytes. J
Biol Chem. 269:23185–23191. 1994.PubMed/NCBI
|