Introduction
Gene expression and regulation are based among other
things on the posttranslational modification system of histone
proteins (1). Disruptor of telomeric
silencing 1 like histone lysine methyltransferase (DOT1L) is an
evolutionarily conserved histone methyltransferase that, according
to the current state of knowledge, exclusively catalyzes the
methylation of H3K79 (2,3). In cancer, this methylation is involved
in DNA damage repair and tumorigenesis. DOT1L was revealed to be
crucial in leukemic transformation and is associated with poor
prognosis in breast and colorectal cancer (4–7).
Sequencing studies (www.cbioportal.org) demonstrate DOT1L alterations in
≥13% of pancreatic cancer cases. The frequencies of DOT1L
copy-number variations determined using gold standard fluorescence
in situ hybridization (FISH), and their specific correlation
to protein expression, remain to be established.
At present, no relevant personalized treatable
target exists in pancreatic adenocarcinoma, thus conventional
chemotherapy and radiotherapy is still the standard systemic
therapy (8–10). Nevertheless the prognosis of
pancreatic adenocarcinoma remains poor, and surgery is the only
curative option for patients. However, only 20% of adenocarcinomas
are operably linked to a curative approach at the time of diagnosis
(11,12). Targeted therapy strategies are
urgently required for this severe form of carcinoma.
In the present study, DOT1L was evaluated as a
potential therapeutic target in pancreatic adenocarcinoma.
According to the results of genomic studies, immunohistochemical
analysis and FISH were performed, hypothesizing that the
identification of a subgroup of carcinoma cases may demonstrate a
significant DOT1L copy-number variation or amplification.
Patients and methods
Patients and tumor samples
In the present retrospective study 230 patients with
pancreatic adenocarcinoma who underwent surgical pancreatic
resections at the University Hospital of Cologne (Cologne, Germany)
between January 1999 and December 2014 were analyzed. For tissue
microarray analysis (TMA), two tissue cores from various areas of
each tumor were obtained and transferred into a TMA recipient
block. TMA construction was performed as described previously
(13,14). Patient characteristics are presented
in Table I. In brief, tissue
cylinders with a diameter of 1.2 mm each were punched from selected
tumor tissue blocks using a homemade semi-automated precision
instrument and inserted into empty recipient paraffin blocks. Four
μm-thick sections of the resulting TMA blocks were transferred to
an adhesive-coated slide system (Instrumedics Inc., Hackensack, NJ,
USA). Consecutive sections were used for FISH and
immunohistochemistry.
 | Table I.Clinicopathological characteristics of
230 pancreatic adenocarcinoma cases. |
Table I.
Clinicopathological characteristics of
230 pancreatic adenocarcinoma cases.
| Characteristic | No. of patients |
|---|
| Gender |
| Male | 124 |
|
Female | 106 |
| Age, years |
|
<50 | 51 |
|
50–60 | 70 |
|
>70 | 99 |
| Tumor stage |
| pT1 | 9 |
| pT2 | 25 |
| pT3 | 190 |
| pT4 | 6 |
| Lymph node
metastasis |
| N0 | 66 |
| N1 | 164 |
| Resection margin
status |
| R0 | 148 |
| R1 | 82 |
| Grading |
| G1 | 3 |
| G2 | 152 |
| G3 | 75 |
Immunohistochemistry
Immunohistochemistry (IHC) was performed on TMA
slides using the primary rabbit polyclonal antibody specific for
DOT1L (#NB100-40845; dilution, 1:50; Novus Biologicals, Ltd.,
Cambridge, UK). Immunohistochemical staining was performed using
the Ventana BenchMark stainer (Roche Diagnostics GmbH, Mannheim,
Germany) according to the manufacturer's protocol with on-slide
controls of the appendix. As a secondary antibody the ready-to-use
Ventana Detection kit (cat. no. 760–700; OptiView DAB IHC Detection
kit; Ventana, Roche Diagnostics GmbH, Mannheim, Germany) for
indirect biotin-free detection of primary rabbit antibody was used.
DOTL1 exhibits a nuclear staining pattern. DOT1L staining was
evaluated manually using light microscopy by two pathologists and
discrepant results were resolved by consensus review.
FISH
FISH for evaluation of DOT1L gene copy numbers was
performed with the DOT1L/CEN19 Dual Color Probe (Empire Genomics,
Buffalo, NY, USA). Three-μm tissue sections on SuperFrost Plus
slides were mounted by heating at 56°C, followed by semi-automated
deparaffinization protease digestion washing steps (VP2000
processor system, Abbott Molecular, Wiesbaden, Germany). Protease
digestion was performed using the ready-to-use FISH pretreatment
kit (Vysis IntelliFISH Universal FFPE Tissue Pretreatment Protease;
Abbott Molecular, Wiesbaden, Germany). Slides were hybridized at
37°C overnight with the FISH probe. The slides were stained with
DAPI prior to analysis. Normal tissue including vessels,
fibroblasts, or non-tumor lung tissue served as an internal
positive control. Cases were only further evaluated if control
tissue nuclei exhibited one or two clearly distinct signals of each
color. Tumor tissue was scanned for amplification hot spots using
×63 objective (DM5500 fluorescent microscope; Leica Microsystems,
Inc., Buffalo Grove, IL, USA). If the signals were homogeneously
distributed, then random areas were used for counting the signals.
Sixty tumor cells in three areas were evaluated by counting green
DOT1L and orange centromere 19 (CEN19) signals. The reading
strategy followed that of the cMET-FISH probe to evaluate different
levels of amplification. Low-level amplification was defined as ≥4
DOT1L signals in ≥40% of cells. Intermediate-level amplification
was defined as ≥5 DOT1L in ≥50% of cells. High-level amplification
was defined as a DOT1L/CEN19 quotient of 2.0, ≥15 DOT1L signals in
≥10% of cells or an average DOT1L copy number of ≥6.
Procedures were followed as outlined in accordance
with ethical standards formulated in the Helsinki Declaration 1975
(and revised in 1983) with pre-approval by the Ethics Committee at
the University Hospital (Cologne, Germany; reference number:
09–232).
Results
Immunohistochemistry
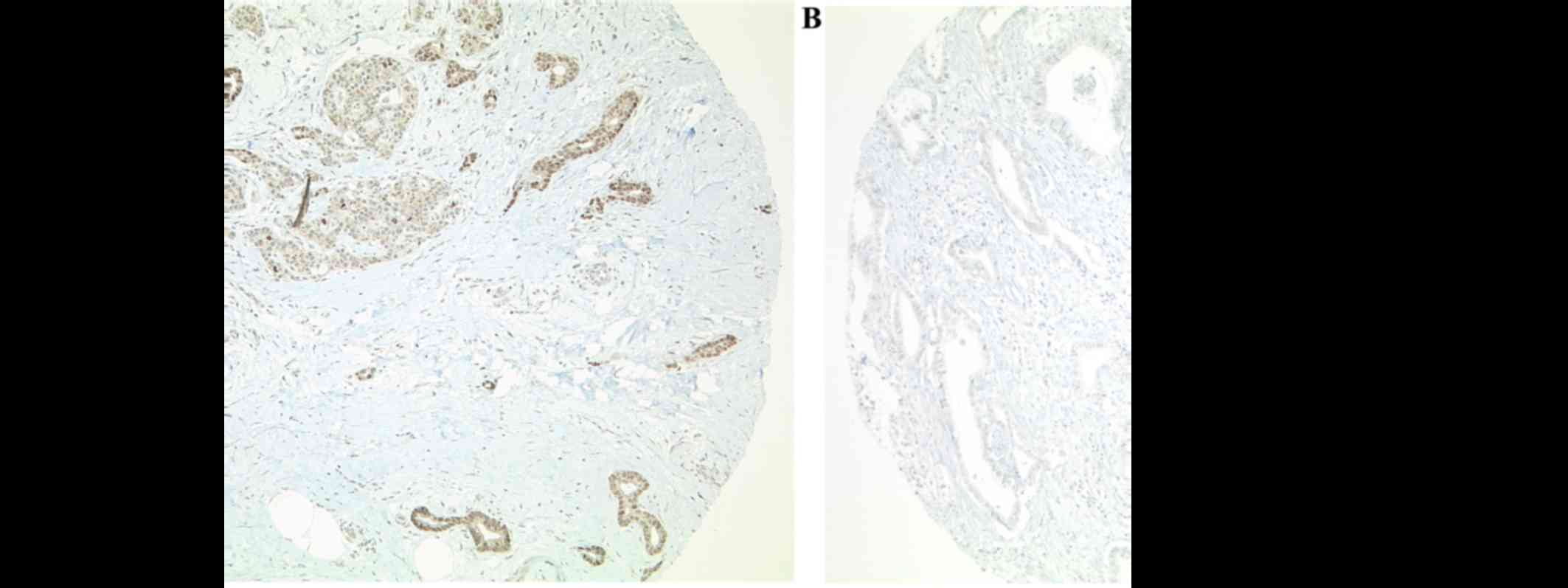
DOT1L immunohistochemistry was performed on TMA
slides, including 230 pancreatic carcinomas. Tissue from the
appendix vermiformis served as on-slide control. Of the 230
pancreatic carcinomas, 225 were evaluable. Ten carcinomas (4.4%)
revealed strong nuclear staining (Fig.
1).
FISH
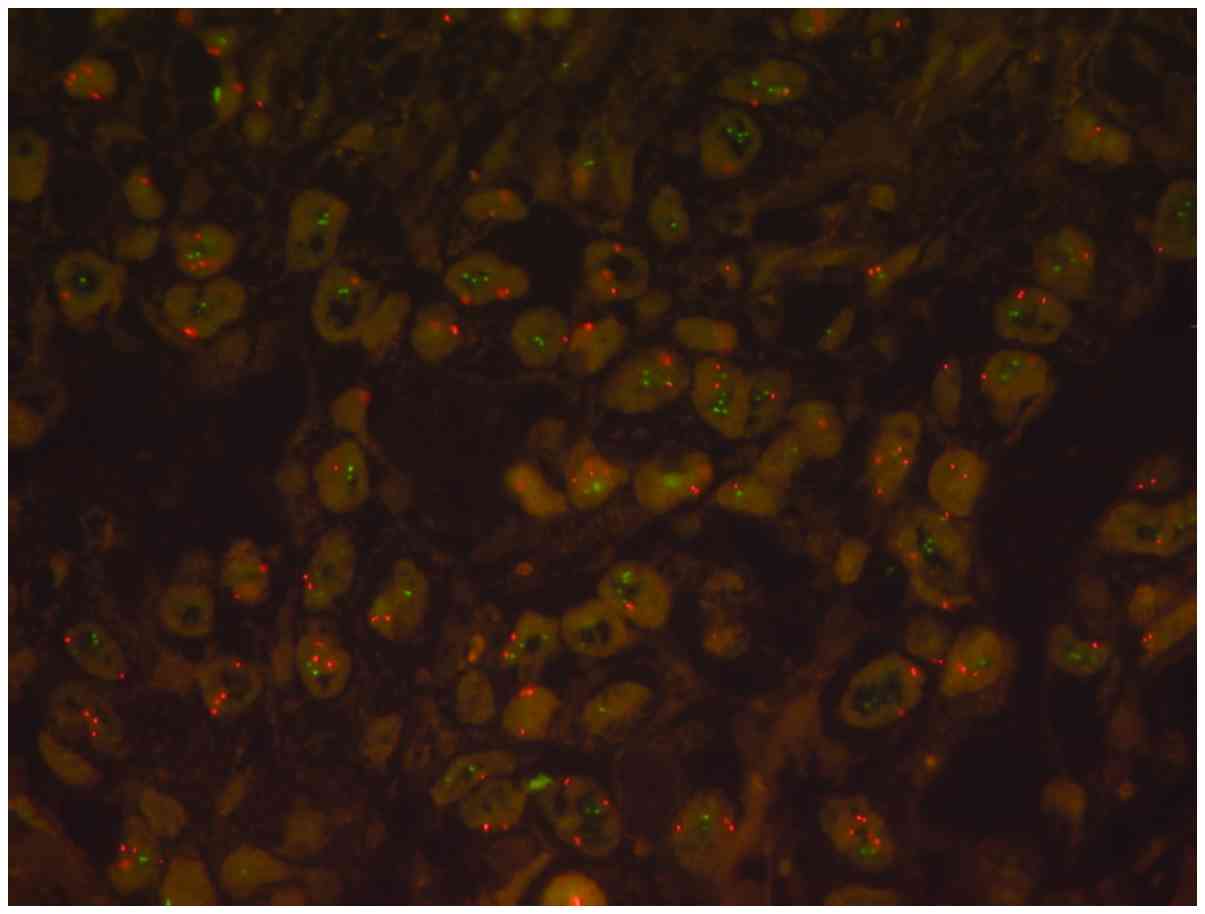
The DOT1L-FISH-signal pattern revealed an even
distribution of the signals in the majority of cases. A total of
224 pancreatic adenocarcinomas were able to be analyzed using FISH;
there were 9 cases with single tumor cells exhibiting microclusters
of >5 DOT1L-signals accompanied by elevated CEN-signals, but no
amplification according to the evaluation criteria (using common
low-, intermediate- and high level amplification criteria of MET
analyses) was detected (Table II).
These elevated DOT1L signals were detected in cases where polysomy
was already detected. Polysomy was slightly increased, ranging from
1.55–2.5 average CEN-signals/cell (Fig.
2).
| Table II.Fluorescence in situ
hybridization analysis of copy number variations of DOT1L in 9
pancreatic adenocarcinoma cases. |
Table II.
Fluorescence in situ
hybridization analysis of copy number variations of DOT1L in 9
pancreatic adenocarcinoma cases.
| Patient | Green signals/60
cells | Orange signals/60
cells | Ratio
green:orange | Average green
signals | Average orange
signals |
|---|
| 1 | 162 | 93 | 1.74 | 2.7 | 1.55 |
| 2 | 174 | 150 | 1.16 | 2.9 | 2.5 |
| 3 | 168 | 94 | 1.78 | 2.8 | 1.6 |
| 4 | 186 | 126 | 1.48 | 3.1 | 2.1 |
| 5 | 176 | 132 | 1.32 | 2.9 | 2.2 |
| 6 | 193 | 145 | 1.32 | 3.2 | 2.4 |
| 7 | 167 | 127 | 1.3 | 2.8 | 2.1 |
| 8 | 158 | 118 | 1.34 | 2.6 | 2.0 |
| 9 | 195 | 130 | 1.5 | 3.2 | 2.2 |
Discussion
According to the analysis of sequencing data,
certain cancer types exhibit DOT1L gene alterations. Concordant
with the biology of MET, the present study hypothesized that a
significant elevation of gene copy number of DOT1L may correlate
with protein overexpression. The augmentation of DOT1L protein
expression may further be associated with an increased response to
a DOT1L-inhibitor therapy. DOT1L is a druggable target causing cell
cycle arrest and leading to chemosensitivity in leukemia (15,16). The
DOT1L inhibitor EPZ-5676 is under clinical investigation in a phase
I study for mixed lineage leukemia (www.ClinicalTrials.gov; cat. no. NCT01684150). The
pharmaco-physiological mechanism of DOTL1 inhibition using the drug
EPZ-5676 in mixed lineage leukemia is not fully understood. The
maintenance of the MLL-AF6 fusion protein requires H3K79
methylation (17). MLL fusion
proteins recruit DOT1L, leading to aberrant DOT1L overexpression
and H3K79 methylation (18,19). In breast carcinoma, DOT1L is
associated with an aggressive phenotype and has also been revealed
to be a potential therapeutic target (4,20,21).
Therefore, the present study immunohistochemically
evaluated the frequency of pancreatic carcinoma cells that
overexpress DOT1L, and how the protein expression correlates with
the DOT1L gene copy number. No association between DOT1L gene copy
number elevation and protein overexpression in tumor cells of the
same cases were identified. Post-transcriptional modifications of
DOT1L that inhibit appropriate protein expression may be one
explanation. On the other hand, DOT1L protein overexpression (not
dependent on an elevation of gene copy number) may be
pharmaco-physiologically important if further studies are able to
establish the expected association, in which more DOT1L protein be
related to more intense methylation of H3K79. At present, the
methylation of H3K79 is considered to accelerate the activity of
various genes involved in cell cycle activity (22). Due to this perception, an inhibition
of DOT1L in cancer may be important. Initial evidence suggests that
the DOT1L inhibitor EPZ-5676 is effective in mixed lineage leukemia
(MLL) (23,24).
According to sequence data, all DOT1L-altered
pancreatic carcinoma harbor a KRAS mutation as well (The Cancer
Genome Atlas data, www.cbioportal.org). This is not an unexpected
finding, due to the fact that >90% of all pancreatic carcinomas
have KRAS mutations (25). The
present study speculates that CNVs of DOT1L in KRAS mutated tumors
are likely to be an epiphenomenon. In accordance with this
assumption, the KRAS mutation may knock out any therapeutically
intended DOT1L inhibition. Furthermore, the distribution of CNVs or
protein expression of DOT1L identified in the present study is
highly heterogeneous. Tumor cells presenting with CNVs of DOT1L
were intermixed with carcinoma cells with regular gene copy
numbers. A similar result was observed in immunohistochemically
positive tumors: Certain tumor cell groups were DOT1L positive and
others were negative.
In conclusion, a subset of ~4% of pancreatic
adenocarcinomas exhibit CNVs of DOT1L with
heterogeneously-distributed single tumor cells showing
microclusters of >5 DOT1L-signals (≤14 signals) or nuclear
protein overexpression. None of the tumor samples meets the
criteria of amplification. There is no correlation between CNVs and
protein overexpression. The significance of possible druggability
in pancreatic adenocarcinoma, particularly in the protein positive
tumor subgroup, requires further study. However, the present study
does not predict a successful therapeutic effect using DOT1L
inhibitors in adenocarcinomas of the pancreas.
References
|
1
|
Wang X, Chen CW and Armstrong SA: The role
of DOT1L in the maintenance of leukemia gene expression. Curr Opin
Genet Dev. 36:68–72. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lacoste N, Utley RT, Hunter JM, Poirier GG
and Côte J: Disruptor of telomeric silencing-1 is a
chromatin-specific histone H3 methyltransferase. J Biol Chem.
277:30421–30424. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Farooq Z, Banday S, Pandita TK and Altaf
M: The many faces of histone H3K79 methylation. Mutat Res Rev Mutat
Res. 768:46–52. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee JY and Kong G: DOT1L: A new
therapeutic target for aggressive breast cancer. Oncotarget.
6:30451–30452. 2015.PubMed/NCBI
|
|
5
|
Kryczek I, Lin Y, Nagarsheth N, Peng D,
Zhao L, Zhao E, Vatan L, Szeliga W, Dou Y, Owens S, et al:
IL-22(+)CD4(+) T cells promote colorectal cancer stemness via STAT3
transcription factor activation and induction of the
methyltransferase DOT1L. Immunity. 40:772–784. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nguyen AT, Taranova O, He J and Zhang Y:
DOT1L, the H3K79 methyltransferase, is required for
MLL-AF9-mediated leukemogenesis. Blood. 117:6912–6922. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Oktyabri D, Ishimura A, Tange S, Terashima
M and Suzuki T: DOT1L histone methyltransferase regulates the
expression of BCAT1 and is involved in sphere formation and cell
migration of breast cancer cell lines. Biochimie. 123:20–31. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Caparello C, Meijer LL, Garajova I,
Falcone A, Le Large TY, Funel N, Kazemier G, Peters GJ, Vasile E
and Giovannetti E: FOLFIRINOX and translational studies: Towards
personalized therapy in pancreatic cancer. World J Gastroenterol.
22:6987–7005. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takaori K, Bassi C, Biankin A, Brunner TB,
Cataldo I, Campbell F, Cunningham D, Falconi M, Frampton AE, Furuse
J, et al: IAP/EPC study group on the clinical managements of
pancreatic cancer: International Association of Pancreatology
(IAP)/European Pancreatic Club (EPC) consensus review of guidelines
for the treatment of pancreatic cancer. Pancreatology. 16:14–27.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gall TM, Tsakok M, Wasan H and Jiao LR:
Pancreatic cancer: Current management and treatment strategies.
Postgrad Med J. 91:601–607. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wagner M, Redaelli C, Lietz M, Seiler CA,
Friess H and Büchler MW: Curative resection is the single most
important factor determining outcome in patients with pancreatic
adenocarcinoma. Br J Surg. 91:586–594. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Badger SA, Brant JL, Jones C, McClements
J, Loughrey MB, Taylor MA, Diamond T and McKie LD: The role of
surgery for pancreatic cancer: A 12-year review of patient outcome.
Ulster Med J. 79:70–75. 2010.PubMed/NCBI
|
|
13
|
Simon R, Mirlacher M and Sauter G: Tissue
microarrays. Methods Mol Med. 114:257–268. 2005.PubMed/NCBI
|
|
14
|
Helbig D, Ihle MA, Pütz K, Tantcheva-Poor
I, Mauch C, Büttner R and Quaas A: Oncogene and therapeutic target
analyses in atypical fibroxanthomas and pleomorphic dermal
sarcomas. Oncotarget. 7:21763–21774. 2016.PubMed/NCBI
|
|
15
|
Liu W, Deng L, Song Y and Redell M: DOT1L
inhibition sensitizes MLL-rearranged AML to chemotherapy. PLoS One.
9:e982702014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Stein EM and Tallman MS: Mixed lineage
rearranged leukaemia: Pathogenesis and targeting DOT1L. Curr Opin
Hematol. 22:92–96. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Deshpande AJ, Chen L, Fazio M, Sinha AU,
Bernt KM, Banka D, Dias S, Chang J, Olhava EJ, Daigle SR, et al:
Leukemic transformation by the MLL-AF6 fusion oncogene requires the
H3K79 methyltransferase Dot1l. Blood. 121:2533–2541. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Okada Y, Feng Q, Lin Y, Jiang Q, Li Y,
Coffield VM, Su L, Xu G and Zhang Y: hDOT1L links histone
methylation to leukemogenesis. Cell. 121:167–178. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wong M, Polly P and Liu T: The histone
methyltransferase DOT1L: Regulatory functions and a cancer therapy
target. Am J Cancer Res. 5:2823–2837. 2015.PubMed/NCBI
|
|
20
|
Zhang L, Deng L, Chen F, Yao Y, Wu B, Wei
L, Mo Q and Song Y: Inhibition of histone H3K79 methylation
selectively inhibits proliferation, self-renewal and metastatic
potential of breast cancer. Oncotarget. 5:10665–10677. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cho MH, Park JH, Choi HJ, Park MK, Won HY,
Park YJ, Lee CH, Oh SH, Song YS, Kim HS, et al: DOT1L cooperates
with the c-Myc-p300 complex to epigenetically derepress CDH1
transcription factors in breast cancer progression. Nat Commun.
6:78212015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim W, Choi M and Kim JE: The histone
methyltransferase Dot1/DOT1L as a critical regulator of the cell
cycle. Cell Cycle. 13:726–738. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Daigle SR, Olhava EJ, Therkelsen CA,
Basavapathruni A, Jin L, Boriack-Sjodin PA, Allain CJ, Klaus CR,
Raimondi A, Scott MP, et al: Potent inhibition of DOT1L as
treatment of MLL-fusion leukemia. Blood. 122:1017–1025. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Klaus CR, Iwanowicz D, Johnston D,
Campbell CA, Smith JJ, Moyer MP, Copeland RA, Olhava EJ, Scott MP,
Pollock RM, et al: DOT1L inhibitor EPZ-5676 displays synergistic
antiproliferative activity in combination with standard of care
drugs and hypomethylating agents in MLL-rearranged leukemia cells.
J Pharmacol Exp Ther. 350:646–656. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Morris JP IV, Wang SC and Hebrok M: KRAS,
Hedgehog, Wnt and the twisted developmental biology of pancreatic
ductal adenocarcinoma. Nat Rev Cancer. 10:683–695. 2010. View Article : Google Scholar : PubMed/NCBI
|