Introduction
Cervical cancer is the third most common type of
cancer and the fourth leading cause of cancer-associated mortality
in women worldwide (1). In
developing countries, it is the most common cause of
cancer-associated mortality, and the five-year survival rate for
patients with advanced cervical cancer is low (1). Thus, it is important to identify and
examine effective drugs in cervical cancer treatment. It has long
been reported that sclareol, a labdane-type diterpene isolated from
the Salvia sclarea plant can inhibit the proliferation and
induce the apoptosis of several cancer cell lines (2–6).
However, the exact mechanisms underlying the antitumorigenic effect
of sclareol in cervical carcinoma remain to be elucidated.
Caveolin-1 (Cav1) is an important component of
caveolae and is known to function as a scaffolding protein in the
regulation of several signaling pathways (7–9). The
loss of Cav1 has been demonstrated to be involved in the
tumorigenesis of several types of cancer, and the overexpression of
Cav1 has been shown to inhibit cell and tumor growth (10–12).
Thus Cav1 is regarded as a potential tumor suppressor. Although
numerous studies have been performed investigating the function of
Cav1 in several types of cancer, the exact role of Cav1 in cervical
carcinoma remains to be elucidated (10–12).
Copper-zinc superoxide dismutase (SOD1) is an
essential element of the superoxide radical eliminating system. It
is the most abundant anti-oxidant enzyme and is predominantly
localized in the cytoplasm, although its localization in the
mitochondrial intermembrane space and nucleus has also been
reported (13,14). Previous studies have revealed that
SOD1 is overexpressed in various types of cancer, and that the
downregulation of SOD1 leads to cancer cell death (15,16),
which indicates that SOD1 is important in tumorigenesis.
In the present study, the anticancer effect of
sclareol was investigated, and the expression levels of Cav1 and
SOD1 in a cervical cancer cell line were investigated, in order to
elucidate the potential mechanisms involved in the anticancer
effect of sclareol.
Materials and methods
Cell culture
HeLa cells were obtained from the American Type
Culture Collection (Rockville, MD, USA). The SW480, SW620, HepG2
and MCF-7 cancer cell lines were obtained from the Cell Resource
Center of the Chinese Academy of Sciences (Shanghai, China). All
cell lines were grown in Dulbecco's modified Eagle's medium (DMEM;
HyClone; GE Healthcare Life Sciences, Logan, UT, USA) supplemented
with 10% fetal bovine serum (HyClone; GE Healthcare Life Sciences).
All cell lines were incubated in a humidified atmosphere containing
5% CO2 at 37°C.
Reagents and antibodies
Sclareol, bortezomib, E64 and pepstatin A were
obtained from Sigma-Aldrich; Thermo Fisher Scientific, Inc.
(Waltham, MA, USA). The Cav1, SOD1, β-tubulin and p62 antibodies
were obtained from ProteinTech Group, Inc., Chicago, IL, USA), the
LC3 antibody was obtained from Sigma-Aldrich; Thermo Fisher
Scientific, Inc.), the Flag antibody was obtained from Prospec-Tany
TechnoGene, Ltd., (East Brunswick, NJ, USA), the HA tag antibody
was obtained from Cell Signaling Technology, Inc. (Beverly, MA,
USA).
MTT and cell apoptosis assays
The cells were plated into 96-well tissue culture
plates and seeded at a density of 10,000 cells per well. Viability
of HeLa cells was determined using an MTT assay following treatment
with vehicle or 10 µg/ml sclareol for 24, 48 or 72 h at 37°C. Cell
apoptosis assay of HeLa cells treated with 5 µg/ml sclareol was
performed for 24, 48 or 72 h at 37°C. The final volume of culture
medium in each well was 100 µl. A 10 µl volume of MTT solution (5
mg/ml) was added to the 100 µl medium in each well. The plates were
incubated at 37°C for 4 h, following which the supernatant was
removed and 100 µl DMSO was added to each well. The absorbance
signals were measured on a spectrophotometer at 490 nm. The cell
death-inducing effects of drug treatment were measured using a
CF488A-Annexin V and Propidium Iodide (PI) Apoptosis Assay kit
(Biotium, Inc., Hayward, CA, USA). The samples and assays were
prepared according to the manufacturer's protocol and then mounted
onto slides. Images were captured with a Nikon fluorescence
microscope (Nikon, Tokyo, Japan).
Western blot analysis
Samples were collected with 1X SDS sample buffer and
were separated by 10–12% SDS-PAGE (20 µg/lane). The proteins were
transferred onto an Immobilon-FL PVDF membrane (EMD Millipore,
Bedford, MA, USA). Protein quantification was performed using a BCA
assay kit (Thermo Fisher Scientific, Inc.). The membrane was first
blocked with 5% milk for 1 h, and then incubated overnight at 4°C
with the following primary antibodies: Cav 1 (cat. no. 16447-1-AP;
1:1,000; ProteinTech Group, Inc., Chicago, IL, USA), SOD1 (cat. no.
10269-1-AP; 1:1,000; ProteinTech Group, Inc.), β-tubilin (cat. no.
66240-1; 1:2,000; ProteinTech Group, Inc.), p62 (cat. no.
18420-1-AP; 1:1,000; ProteinTech Group, Inc.), LC3 (cat. no. L8918;
1:1,000; Sigma-Aldrich; Merck Millipore, Darmstadt, Germany), Flag
(cat. no. ANT-222; 1:1,000; Prospec-Tany TechnoGene, Ltd., East
Brunswick, NJ, USA) and HA tag (cat. no. C29F4; 1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA) diluted in the same
blocking buffer. Following three washes with Tris-buffered saline
with Tween 20 (TBST) containing 50 mM Tris-HCl (pH 7.5), 150 mM
NaCl and 0.2% Tween 20, the blot was incubated with Dylight
680/800-conjugated secondary antibodies (cat. nos. 042-06-15-06 and
042-06-18-0; 1:10,000; KPL, Inc. Gaithersburg, MD, USA) in the dark
for 2 h at room temperature. The blot was then washed again with
TBST and images were captured using a Li-Cor Odyssey Clx infrared
imaging system (LI-COR BioSciences, Inc., Lincoln, NE, USA).
Establishment of stable cell line
For the stable expression of Cav-1, the HeLa cells
were transduced with Cav1-Flag lentiviral particles using ViraPower
Lentiviral Expression system following the manufacturer's protocol
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol, and stable cell lines were selected using
blasticidin (Invitrogen; Thermo Fisher Scientific, Inc.).
Co-immunoprecipitation assay
At 24 h post-transduction, the cells were washed
with phosphate-buffered saline (PBS) and lysed in 1% NP40 lysis
buffer for 30 min on ice. The cell lysates were collected in an EP
tube and then centrifuged at >18,000 × g for 10 min at
4°C, the supernatants were immunoprecipitated with the Flag
(1:1,000) and HA tag (1:1,000) antibodies overnight at 4°C.
Prewashed protein A/G-agarose beads (Thermo Fisher Scientific,
Inc.) were then added to the supernatant and incubated for another
2 h at 4°C. The immune complexes were washed with lysis buffer
three times, resuspended in sample buffer and then subjected to
western blot analysis.
Immunofluorescence assay
The cells were plated on coverslips at a density of
1.0×105 cells and transfected with the indicated plasmid
using Lipofectamine 2000. HeLa cells and HeLa cells stably
transfected with Cav1-Flag (Cav1) were transiently transfected with
a plasmid expressing lysosome-associated protein 2A (Lamp-2A)-EGFP.
At 24 h post-transfection, the cells were then fixed with 4%
paraformaldehyde for 15 min at room temperature, rinsed with PBS
and permeabilized with 0.25% Triton-x-100 for 10 min at room
temperature. Following rinsing with PBS three times, the cells were
blocked with 3% bovine serum albumin (Sigma-Aldrich; Merck
Millipore) overnight at 4°C. The EGFP (cat. no. 50430-2; 1:1,000;
Proteintech Group, Inc.), Flag (1:1,000) antibodies were added and
incubated overnight at 4°C and rinsed with PBS three times The
secondary antibody was added for 1 hour at room temperature,
following which the cells were rinsed with PBS three times, and
then incubated with DAPI for 10 min at room temperature. The cells
were then rinsed, mounted and sealed. Images were captured using an
Olympus confocal microscope (Olympus, Tokyo, Japan).
Plasmid construction and
transfection
The full-length Cav1 open reading frame (ORF) was
amplified by polymerase chain reaction (PCR) with a BamHI
site-containing 5′ primer and an XbaI site-containing 3′
primer, and cloned into the pcDNA3.1(+)-flag vector (Invitrogen;
Thermo Fisher Scientific, Inc.) between the BamHI and
XbaI sites to generate the Cav1-Flag expression construct.
The full-length heat-shock protein cognate 70 (HSC70) ORF was
amplified by PCR with a BamHI site-containing 5′ primer and
an XbaI site-containing 3′ primer, and cloned between the
BamHI and XbaI sites of the pcDNA3.1(+)-HA vector to
generate the HSC70-HA expression construct. Full-length LAMP-2A ORF
was amplified by PCR with a BamHI site-containing 5′ primer
and an XbaI site-containing 3′ primer, and cloned between
the BamHI and XbaI sites of the pcDNA3.1(+)-EGFP
vector to generate the LAMP-2A-EGFP expression construct. The human
cDNA preserved in the laboratory was utilized as the template. Flag
tag, HA tag and EGFP tag were cloned into the pcDNA3.1(+) vector
(Life Technologies; Thermo Fisher Scientific, Inc.) between
XbaI and ApaI to generate the pcDNA3.1(+)-tagged
vector. For the generation of Cav1Flag lentiviral particles, the
Cav1-Flag cDNA fragment was inserted into a pLenti6 lentiviral
vector (Invitrogen Life Technologies; Thermo Fisher Scientific,
Inc.), and the resultant plasmid was co-transfected with ViraPower
Lentiviral Packaging mix (Invitrogen Life Technologies; Thermo
Fisher Scientific, Inc.) into HEK293FT cells with Lipofectamine
2000. The virus-containing cell culture medium was harvested 40 h
later and used to transduce the HeLa cells. The sequences of the
primers are listed in Table I.
 | Table I.Sequences of primers for plasmid
construction. |
Table I.
Sequences of primers for plasmid
construction.
| Primer name | Sequence
(5′-3′) |
|---|
| Cav1-F |
gatcggatccgccaccatgtctgggggcaaatacgtag |
| Cav1-R |
gtagttgaacgtctttctttatagatctctag |
| HSC70-F |
gatcggatccgccaccatgtccaagggacctgcagttg |
| HSC70-R |
gggtggtaacttctccaactaagatctctag |
| LAMP-2A-F |
gatcggatccgccaccatggtgtgcttccgcctcttc |
| LAMP-2A-R |
gtacgacctatactcgttaaaagatctctag |
Statistical analysis
Experimental data are expressed as the mean ±
standard error of the mean. Data analyses were performed using
ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Differences between groups were compared using one-way analysis of
variance followed by Tukey's post-hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
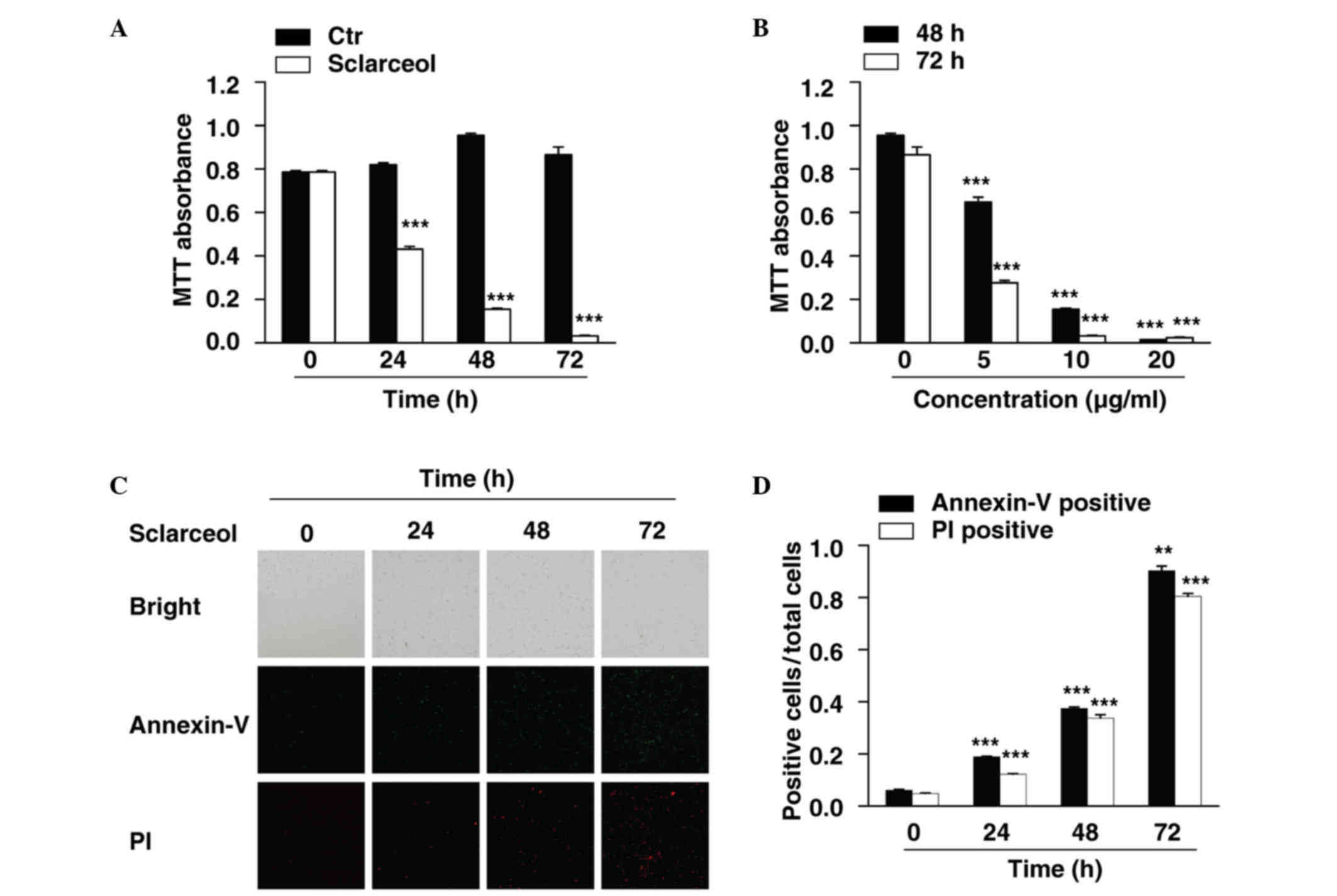
Sclareol inhibits cell proliferation
and induces apoptosis in HeLa cells
In the present study, HeLa cells were used to
investigate the anticancer effects of sclareol in cervical
carcinoma. An MTT assay was first performed to measure the
viability of the HeLa cells. The cells were treated with sclareol
for 24, 48 and 72 h, and cell viability was measured at each time
point. The results showed that sclareol significantly inhibited the
proliferation of the HeLa cells, and the inhibitory effect was
time-dependent (Fig. 1A). In
addition, when the cells were treated with different concentrations
of sclareol, the proliferation of the HeLa cells was inhibited in a
dose-dependent manner (Fig.
1B).
Treatment with sclareol also induced the apoptosis
of the HeLa cells. An Annexin V-fluorescein isothiocyanate
(FITC)/PI staining assay was used to assess the proportion of
apoptotic cells when HeLa cells were treated with sclareol for
indicated durations. The results indicated that the Annexin
V-FITC-positive and PI-positive cells were significantly increased
in a time-dependent manner when treated with sclareol (Fig. 1C and D).
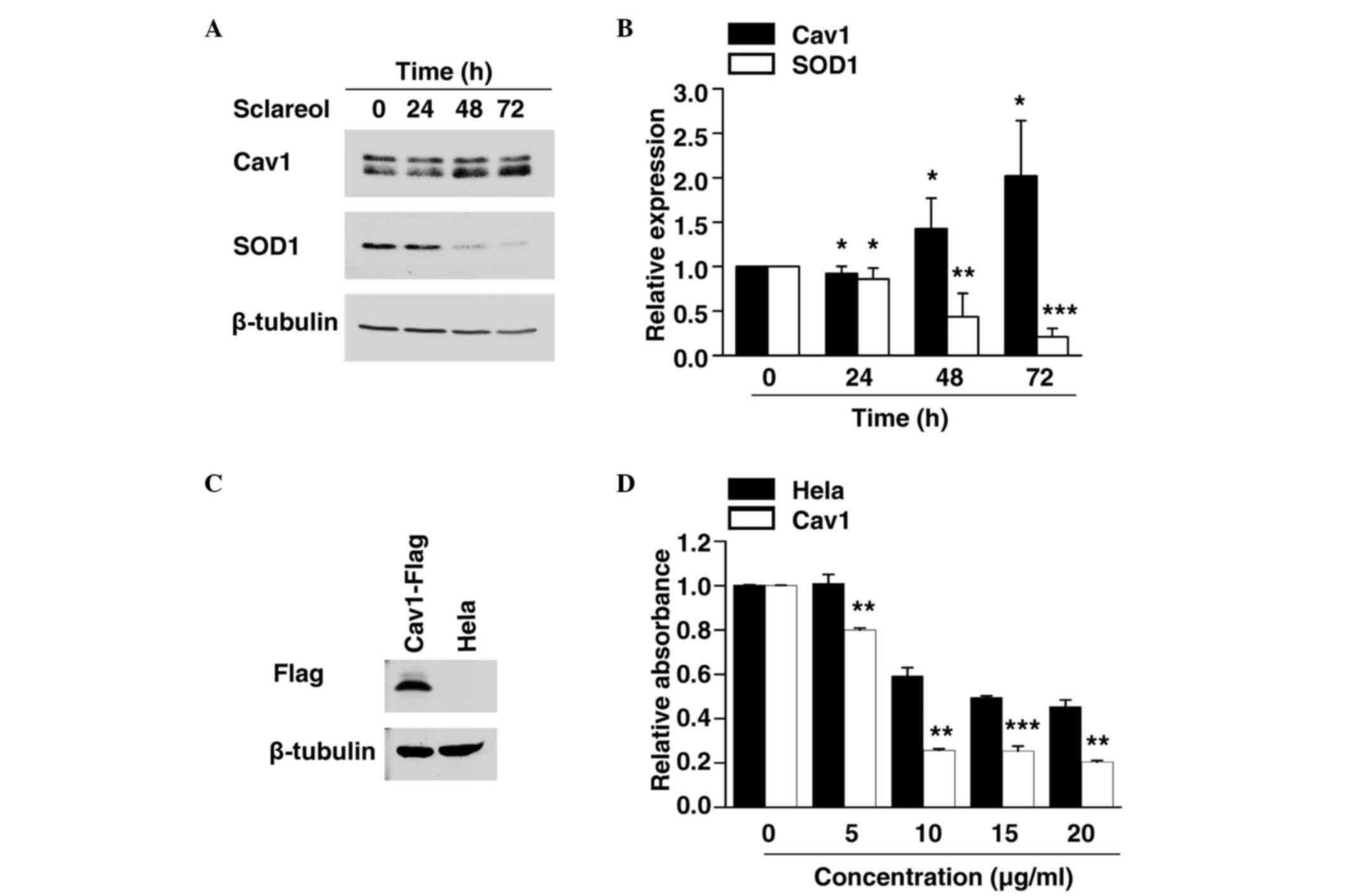
Sclareol induces the expression of
Cav1
As the exact mechanism involved in the anticancer
effect of sclareol remains to be fully elucidated, the present
study examined the expression of various cancer-associated proteins
to identify the potential target of sclareol. The cells were
exposed to sclareol for 24, 48 and 72 h, and were then subjected to
western blot analysis. The results showed that sclareol induced the
protein levels of Cav1 in a time-dependent manner, compared with
the control group (Fig. 2A and
B).
Expression of SOD1 is negatively
correlated with Cav1
SOD1 has previously been identified as a novel
cancer gene (13,14). The results of the present study
showed that the protein level of SOD1 was significantly decreased
in a time-dependent manner when treated with sclareol (Fig. 2A and B), and the decreased
expression of SOD1 was negatively correlated with the increased
expression of Cav1.
Cells overexpressing Cav1 are more
sensitive to sclareol treatment
The present study established a HeLa cell line
stably expressing Cav1. The stable Cav1-expressing cell line and
normal HeLa cells were treated with vehicle or sclareol at the
indicated concentration for 24 h, following which the cell
viability was measured using an MTT assay. It was found that stable
expression of Cav1 enhanced the sensitivity of the cells to
sclareol treatment (Fig. 2C and
D). These results indicated that the upregulation of Cav1 was
responsible for the anticancer effect of sclareol.
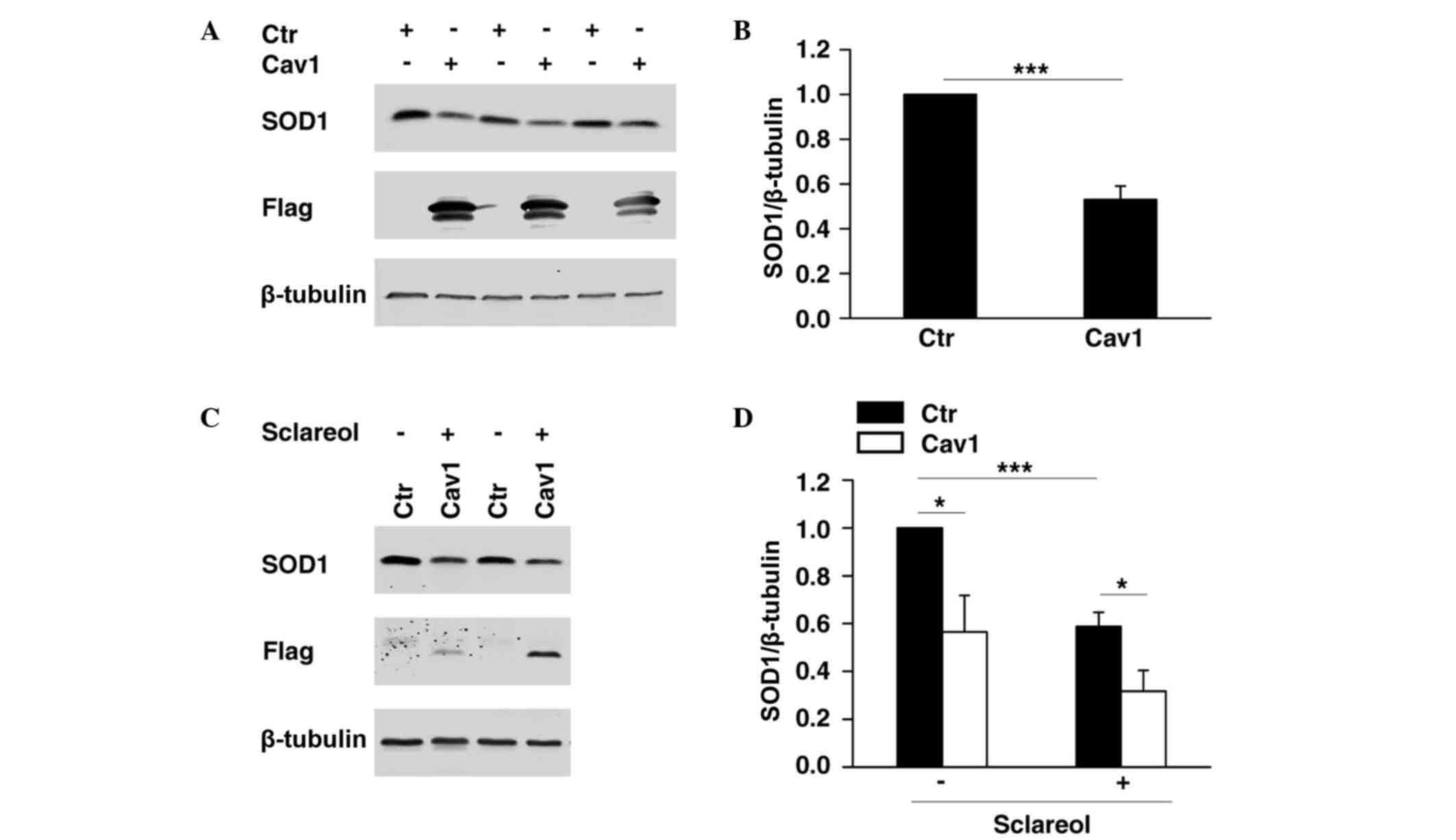
Cav1 promotes the downregulation of
SOD1
It was previously demonstrated that SOD1 is
important in cancer development (10,11),
whereas Cav1 has been reported to be a cancer suppressor. As shown
in Fig. 2, the present study found
that SOD1 was negatively correlated with Cav1. To determine whether
the expression of SOD1 was directly regulated by Cav1, HeLa cells
overexpressing Flag-tagged Cav1 were established, and western blot
analysis was performed to determine the protein level of SOD1. The
results showed that the overexpression of Cav1 significantly
promoted the downregulation of SOD1 (Fig. 3A and B). In the HeLa cell line
stably expressing Cav1, it was found that stable expression of Cav1
enhanced the downregulation of SOD1 when treated with sclareol
(Fig. 3C and D).
Sclareol sensitizes cells to the
antiproliferative effect of bortezomib by targeting Cav1 and
SOD1
Previous investigations have revealed that Cav1 is
degraded through the ubiquitin-proteasome pathway (17,18).
As the results of the present study showed that sclareol inhibited
cancer cell proliferation through the upregulation of Cav1, whether
bortezomib, an anticancer drug and a proteasome inhibitor, enhances
the anticancer effect of sclareol was investigated. The HeLa cells
were treated with sclareol only, bortezomib only or sclareol and
bortezomib together, and subjected to western blot analysis and an
MTT assay. By measuring the cell viability of each group, it was
found that sclareol significantly enhanced the antiproliferative
effect of bortezomib, compared with the cells treated with either
drug separately (Fig. 4A). The
results of the western blot analysis showed that the protein level
of Cav1 increased following treatment with bortezomib (Fig. 4B and C). In addition, the HeLa
cells treated with sclareol and bortezomib together exhibited a
lower level of SOD1 and higher level of Cav1, compared with the
vehicle group (Fig. 4B and C).
Similar results were observed when other cancer cells were treated
with the drug combination of bortezomib and sclareol (Fig. 4D).
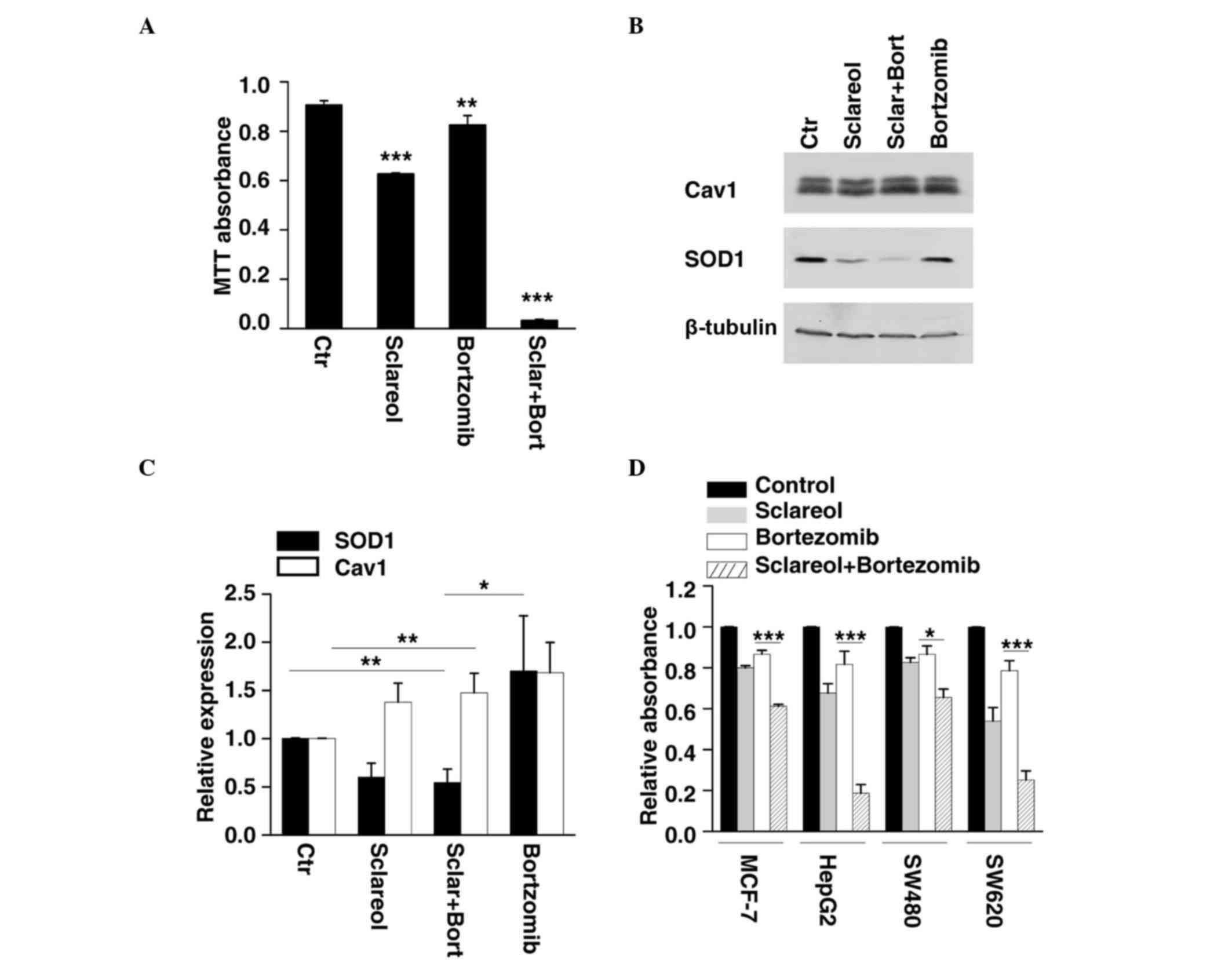 | Figure 4.Sclareol synergizes with bortezomib to
inhibit cancer cell growth. (A) HeLa cells were treated with
sclareol (5 µg/ml) only, bortezomib (5 nM) only, or sclareol (5
µg/ml) with bortezomib (5 nM) for 48 h, and subjected to MTT
analysis. Data are presented as the mean + standard error of the
mean of three independent experiments. *P<0.05, **P<0.01 and
***P<0.005, compared with the vehicle group. (B) Treated HeLa
cells were also subjected to western blot analysis. (C)
Quantification of the relative protein levels of SOD1 and Cav1 in
HeLa cells treated with vehicle (Ctr) or indicated drugs. Data are
presented as the mean + standard error of the mean of three
independent experiments. *P<0.05, **P<0.01 and ***P<0.005,
compared with the control group. (D) Sclareol sensitized cells to
the anti-proliferative effect of bortezomib in several cancer cell
lines. (A) MCF-7, HepG2, SW480 and SW620 cells were also subjected
to MTT analysis. Data are presented as the mean + standard error of
the mean of three independent experiments. *P<0.05, **P<0.01
and ***P<0.005 compared with control group. Ctr, control; Cav1,
caveolin-1; SOD1, superoxide dismutase 1; Sclar, sclareol; Bort,
bortzomib. |
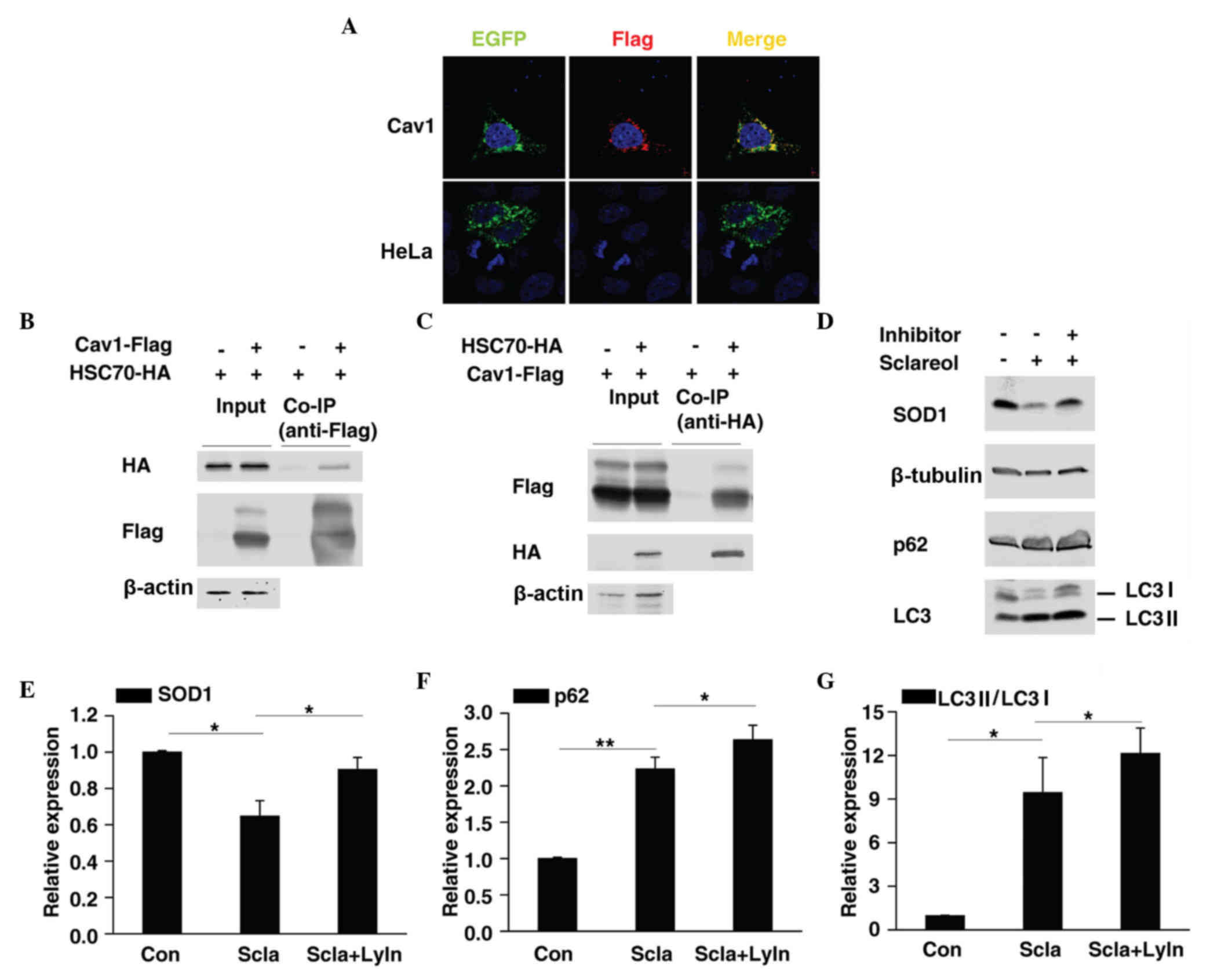
Cav1 may be involved in the
lysosome-mediated degradation of SOD1
Previous reports have shown that Cav1 mediates the
degradation of several proteins (19–21),
thus it is possible that Cav1 promotes the degradation of SOD1.
HSC70 and LAMP-2A are two important components of
chaperone-mediated autophagy (CMA) (22). The present study found that Cav1
not only colocalized with LAMP-2A (Fig. 5A) but also interacted with HSC70
(Fig. 5B and C), thus it is
possible that Cav1 may be involved in CMA-mediated protein
degradation. CMA substrates are usually delivered to the lysosome
for degradation (23). In the
present study, HeLa cells were treated with sclareol only or
sclareol together with lysosome inhibitors and then subjected to
western blot analysis. Of note, as with p62/SQSTM1 and LC3 II,
which are two autophagy-associated proteins previously reported to
be degraded in lysosomes (24,25),
inhibition of lysosome activity suppressed the downregulation of
SOD1 caused by sclareol treatment (Fig. 5D-G), which suggested that sclareol
treatment may promote the downregulation of SOD1 through lysosome
mediated degradation via Cav1.
Discussion
Sclareol exhibits antiproliferative and cytotoxic
activity against various human cancer cell lines (2–6), and
inhibits tumor growth in vivo (3,6). The
anticancer effect of sclareol has long been reported, however, the
exact role of sclareol and its molecular mechanisms in cervical
carcinoma remain to be elucidated. The present study demonstrated
that sclareol inhibited proliferation and induced apoptosis in HeLa
cells, and the combination of sclareol with another anticancer
drug, bortezomib, significantly inhibited tumor cell proliferation
in several types of cancer cell. It was also shown that sclareol
treatment induced the expression of Cav1 in HeLa cells.
Although the role of Cav1 in tumorigenesis is
controversial, increasing evidence indicates that Cav1 functions as
a tumor suppressor in various types of cancer. The downregulation
of Cav1 has been implicated in the development and progression of
cutaneous squamous cell carcinoma (26), lung carcinoma (12), alveolar rhabdomyosarcomas (10), colon cancer (11), sporadic vestibular schwannomas
(27) and breast cancer (28). The overexpression of Cav1 decreases
tumor cell growth (10–12) and enhances the sensitivity of
cancer cells to anticancer drugs (21). However, the exact function of Cav1
in cervical carcinoma remains to be elucidated. In the present
study, the upregulation of Cav1 correlated with decreased cell
viability under sclareol treatment, which indicated that Cav1 may
function as a tumor suppressor in HeLa cells.
Cav1 is ubiquitinated and targeted to lysosomes for
degradation (17,18), and previous studies have
demonstrated that Cav-1 is also a molecular target of bortezomib,
which is involved in antitumorigenesis and proteasome inhibition
(29), and is degraded through the
proteasome pathway (30). The data
obtained in the present study showed that sclareol increased the
protein level of Cav1, and the combined treatment with bortezomib
further enhanced the expression of Cav1. Based on these findings,
it is possible that the combination of sclareol with bortezomib may
have novel synergistic antitumor effects via upregulation of the
cancer suppressor, Cav1. In line with this hypothesis, the present
study found that bortezomib synergized with sclareol to reduce the
viability of various types of cancer cell, including HeLa, HepG2,
MCF-7, SW480 and SW620 cells, whereas treatment with bortezomib or
sclareol alone at the same concentration had markedly lower
inhibitory effects. Thus, the combination of bortezomib with
sclareol may be a novel and promising approach in cancer
therapy.
Previous reports have revealed the potential
mechanism involved in the anticancer effect of Cav1. Cav-1 has been
shown to prevent cancer growth via inhibition of the
mitogen-activated protein kinase pathway (10) and to enhance the degradation of
epidermal growth factor receptor (EGFR) (21). In the present study, the increased
protein level of Cav1 was negatively correlated with the expression
of SOD1, and the overexpression of Cav1 significantly suppressed
the protein level of SOD1. These findings indicated a potential
association between Cav1 and SOD1.
The expression of SOD1 has been linked to the
resistance of cancer cells to pro-oxidant drugs and the anticancer
drug. bortezomib (31), which
raises the possibility that an SOD1 inhibitor can be used, not only
as an antitumorigenic drug, but also as a molecule to challenge the
drug resistance of cancer cells. The data obtained in the present
study showed that the treatment of HeLa cells with sclareol led to
downregulation of the protein level of SOD1, whereas the
overexpression of Cav1 enhanced the suppressed expression of SOD1.
These findings suggested that sclareol may function as an SOD1
inhibitor via the upregulation of Cav1. As the first anticancer
drug to function as a proteasome inhibitor, bortezomib has been
approved for the clinical treatment of patients with multiple
myeloma (32), and has also been
demonstrated to be an effective cell growth inhibitor in various
cancer cell lines (33–36). However, the emergence of drug
resistance has seriously affected the therapeutic effect of
bortezomib. The overexpression of SOD1 has been shown to increase
the resistance of cancer cells to bortezomib (31), and the results of the present study
showed that combined sclareol and bortezomib treatment resulted in
markedly lower protein levels of SOD1, compared with bortezomib
treatment alone, which suggested that combined sclareol and
bortezomib treatment may be a promising therapy in overcoming the
challenge of bortezomib resistance.
Cav1 is important in Derlin-1- and p97-mediated
cyclooxygenase-2 ubiquitination and degradation (19,20),
and previous reports have shown that Cav1 enhances the
ubiquitination and degradation of EGFR via endocytosis (21). However, how Cav1 promotes the
suppressed expression of SOD1 remains to be fully elucidated. The
present study found that inhibition of the lysosome rescued the
decreased protein level of SOD1, which was induced by sclareol,
thus Cav1 may interact with SOD1 to facilitate its degradation
inside the lysosome. However, the protein level of SOD1 cannot be
fully rescued by lysosome inhibition, which indicates Cav1 is not
the only molecule to promote the degradation of SOD1 and that SOD1
is not only degraded by the lysosome-mediated pathway. Further
detailed investigation is required to elucidate the exact mechanism
involved.
In conclusion, the present study demonstrated the
inhibitory effect of sclareol on the proliferation of HeLa cells.
In addition, it was found that the anticancer effect of sclareol
was enhanced when combined with bortezomib, which was demonstrated
in various cancer cells. Further investigations revealed that the
tumor suppressor, Cav1, enhanced the decreased expression of SOD1,
which may be responsible for the anticancer effect of sclareol.
These findings not only elucidated the potential mechanisms
underlying the anticancer effect of sclareol, but also provided a
novel and promising approach for cancer therapy and resolving drug
resistance in cancer cells.
Acknowledgements
This study was supported by research grants from the
Natural Science Foundation of Chengdu University (grant no.
2011XJZ14) and the Natural Science Foundation of China (grant no.
51402027).
References
|
1
|
Jemal A, Center MM, DeSantis C and Ward
EM: Global patterns of cancer incidence and mortality rates and
trends. Cancer Epidemiol Biomarkers Prev. 19:1893–1907. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Noori S, Hassan ZM and Salehian O:
Sclareol reduces CD4+CD25+FoxP3+Treg cells in a breast cancer model
in vivo. Iran J Immunol. 10:10–21. 2013.PubMed/NCBI
|
|
3
|
Noori S, Hassan ZM, Mohammadi M, Habibi Z,
Sohrabi N and Bayanolhagh S: Sclareol modulates the Treg
intra-tumoral infiltrated cell and inhibits tumor growth in vivo.
Cellular Immunol. 263:148–153. 2010. View Article : Google Scholar
|
|
4
|
Wang L, He HS, Yu HL, Zeng Y, Han H, He N,
Liu ZG, Wang ZY, Xu SJ and Xiong M: Sclareol, a plant diterpene,
exhibits potent antiproliferative effects via the induction of
apoptosis and mitochondrial membrane potential loss in osteosarcoma
cancer cells. Mol Med Rep. 11:4273–4278. 2015.PubMed/NCBI
|
|
5
|
Sashidhara KV, Rosaiah JN, Kumar A, Bid
HK, Konwar R and Chattopadhyay N: Cell growth inhibitory action of
an unusual labdane diterpene, 13-epi-sclareol in breast and uterine
cancers in vitro. Phytotherapy Res. 21:1105–1108. 2007. View Article : Google Scholar
|
|
6
|
Dimas K, Hatziantoniou S, Tseleni S, Khan
H, Georgopoulos A, Alevizopoulos K, Wyche JH, Pantazis P and
Demetzos C: Sclareol induces apoptosis in human HCT116 colon cancer
cells in vitro and suppression of HCT116 tumor growth in
immunodeficient mice. Apoptosis. 12:685–694. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Simmons GE, Taylor HE and Hildreth JEK:
Caveolin-1 suppresses human immunodeficiency virus-1 replication by
inhibiting acetylation of NF-κB. Virology. 432:110–119. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Garrean S, Gao XP, Brovkovych V, Shimizu
J, Zhao YY, Vogel SM and Malik AB: Caveolin-1 regulates NF-kappaB
activation and lung inflammatory response to sepsis induced by
lipopolysaccharide. J Immunol. 177:4853–4860. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang XM, Kim HP, Song R and Choi AM:
Caveolin-1 confers antiinflammatory effects in murine macrophages
via the MKK3/p38 MAPK pathway. Am J Respir Cell Mol Biol.
34:434–442. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huertas-Martinez J, Rello-Varona S,
Herrero-Martin D, Barrau I, García-Monclús S, Sáinz-Jaspeado M,
Lagares-Tena L, Núñez-Álvarez Y, Mateo-Lozano S, Mora J, et al:
Caveolin-1 is down-regulated in alveolar rhabdomyosarcomas and
negatively regulates tumor growth. Oncotarget. 5:9744–9755. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bender FC, Reymond MA, Bron C and Quest
AF: Caveolin-1 levels are down-regulated in human colon tumors and
ectopic expression of caveolin-1 in colon carcinoma cell lines
reduces cell tumorigenicity. Cancer Res. 60:5870–5878.
2000.PubMed/NCBI
|
|
12
|
Belanger MM, Roussel E and Couet J:
Caveolin-1 is down-regulated in human lung carcinoma and acts as a
candidate tumor suppressor gene. Chest. 125 Supply 5:S1062004.
View Article : Google Scholar
|
|
13
|
Fischer LR, Igoudjil A, Magrané J, Li Y,
Hansen JM, Manfredi G and Glass JD: SOD1 targeted to the
mitochondrial intermembrane space prevents motor neuropathy in the
Sod1 knockout mouse. Brain. 134(Pt 1): 196–209. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Igoudjil A, Magrané J, Fischer LR, Kim HJ,
Hervias I, Dumont M, Cortez C, Glass JD, Starkov AA and Manfredi G:
In vivo pathogenic role of mutant SOD1 localized in the
mitochondrial intermembrane space. J Neurosci. 31:15826–15837.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Somwar R, Erdjument-Bromage H, Larsson E,
Shum D, Lockwood WW, Yang G, Sander C, Ouerfelli O, Tempst PJ,
Djaballah H and Varmus HE: Superoxide dismutase 1 (SOD1) is a
target for a small molecule identified in a screen for inhibitors
of the growth of lung adenocarcinoma cell lines. Proc Natl Acad Sci
USA. 108:pp. 16375–16380. 2011; View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Glasauer A, Sena LA, Diebold LP, Mazar AP
and Chandel NS: Targeting SOD1 reduces experimental non-small-cell
lung cancer. J Clin Invest. 124:117–128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kirchner P, Bug M and Meyer H:
Ubiquitination of the N-terminal region of caveolin-1 regulates
endosomal sorting by the VCP/p97 AAA-ATPase. J Biol Chem.
288:7363–7372. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hayer A, Stoeber M, Ritz D, Engel S, Meyer
HH and Helenius A: Caveolin-1 is ubiquitinated and targeted to
intralumenal vesicles in endolysosomes for degradation. J Cell
Biol. 191:615–629. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen SF, Wu CH, Lee YM, Tam K, Tsai YC,
Liou JY and Shyue SK: Caveolin-1 interacts with Derlin-1 and
promotes ubiquitination and degradation of cyclooxygenase-2 via
collaboration with p97 complex. J Biol Chem. 288:33462–33469. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen SF, Liou JY, Huang TY, Lin YS, Yeh
AL, Tam K, Tsai TH, Wu KK and Shyue SK: Caveolin-1 facilitates
cyclooxygenase-2 protein degradation. J Cell Biochem. 109:356–362.
2010.PubMed/NCBI
|
|
21
|
Feldman R and Martinez JD: Growth
suppression by ursodeoxycholic acid involves caveolin-1 enhanced
degradation of EGFR. Biochim Biophys Acta. 1793:1387–1394. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qi L, Zhang XD, Wu JC, Lin F, Wang J,
DiFiglia M and Qin ZH: The role of chaperone-mediated autophagy in
huntingtin degradation. PloS One. 7:e468342012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cuervo AM, Dice JF and Knecht E: A
population of rat liver lysosomes responsible for the selective
uptake and degradation of cytosolic proteins. J Biol Chem.
272:5606–5615. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liang X, Wei SQ, Lee S, Fung JK, Zhang M,
Tanaka A, Choi AM and Jin Y: p62 sequestosome 1/light chain 3b
complex confers cytoprotection on lung epithelial cells after
hyperoxia. Am J Respir Cell Mol Biol. 48:489–496. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gómez-Sánchez R, Yakhine-Diop SM,
Rodríguez-Arribas M, Bravo-San Pedro JM, Martínez-Chacón G,
Uribe-Carretero E, de Castro DC Pinheiro, Pizarro-Estrella E,
Fuentes JM and González-Polo RA: mRNA and protein dataset of
autophagy markers (LC3 and p62) in several cell lines. Data Brief.
7:641–647. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Trimmer C, Bonuccelli G, Katiyar S, Sotgia
F, Pestell RG, Lisanti MP and Capozza F: Cav1 suppresses tumor
growth and metastasis in a murine model of cutaneous SCC through
modulation of MAPK/AP-1 activation. Am J Pathol. 182:992–1004.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Aarhus M, Bruland O, Saetran HA, Mork SJ,
Lund-Johansen M and Knappskog PM: Global gene expression profiling
and tissue microarray reveal novel candidate genes and
down-regulation of the tumor suppressor gene CAV1 in sporadic
vestibular schwannomas. Neurosurgery. 67:998–1019. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shi Y, Tan SH, Ng S, Zhou J, Yang ND, Koo
GB, McMahon KA, Parton RG, Hill MM, Del Pozo MA, et al: Critical
role of CAV1/caveolin-1 in cell stress responses in human breast
cancer cells via modulation of lysosomal function and autophagy.
Autophagy. 11:769–784. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Podar K, Shringarpure R, Tai YT, Simoncini
M, Sattler M, Ishitsuka K, Richardson PG, Hideshima T, Chauhan D
and Anderson KC: Caveolin-1 is required for vascular endothelial
growth factor-triggered multiple myeloma cell migration and is
targeted by bortezomib. Cancer Res. 64:7500–7506. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mao M, Sudhahar V and Fukai T: P54:
Nitroglycerin-induced loss of caveolin-1 results in eNOS
dysfunction, peroxynitrite production and nitrate tolerance. Nitric
Oxide 31, Supplement. 1:S362013. View Article : Google Scholar
|
|
31
|
Salem K, McCormick ML, Wendlandt E, Zhan F
and Goel A: Copper-zinc superoxide dismutase-mediated redox
regulation of bortezomib resistance in multiple myeloma. Redox
Biol. 4:23–33. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen D, Frezza M, Schmitt S, Kanwar J and
Dou QP: Bortezomib as the first proteasome inhibitor anticancer
drug: Current status and future perspectives. Curr Cancer Drug
Targets. 11:239–253. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kao C, Chao A, Tsai CL, Chuang WC, Huang
WP, Chen GC, Lin CY, Wang TH, Wang HS and Lai CH: Bortezomib
enhances cancer cell death by blocking the autophagic flux through
stimulating ERK phosphorylation. Cell Death Dis. 5:e15102014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kretowski R, Stypulkowska A and
Cechowska-Pasko M: Efficient apoptosis and necrosis induction by
proteasome inhibitor: Bortezomib in the DLD-1 human colon cancer
cell line. Mol Cell Biochem. 398:165–173. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Miyamoto Y, Nakagawa S, Wada-Hiraike O,
Seiki T, Tanikawa M, Hiraike H, Sone K, Nagasaka K, Oda K, Kawana
K, et al: Sequential effects of the proteasome inhibitor bortezomib
and chemotherapeutic agents in uterine cervical cancer cell lines.
Oncol Rep. 29:51–57. 2013.PubMed/NCBI
|
|
36
|
Ando M, Hoyos V, Yagyu S, Tao W, Ramos CA,
Dotti G, Brenner MK and Bouchier-Hayes L: Bortezomib sensitizes
non-small cell lung cancer to mesenchymal stromal cell-delivered
inducible caspase-9-mediated cytotoxicity. Cancer Gene Ther.
21:472–482. 2014. View Article : Google Scholar : PubMed/NCBI
|