Introduction
Vascular endothelial cells are flat cells lining the
inner wall of blood vessels, which serve a key role as the
mechanical barrier between circulating blood and vascular smooth
muscle cells. In addition, the vascular endothelium is an important
endocrine organ, which is involved in maintaining vascular wall
tension, blood flow, vessel wall inflammation antagonism and
angiogenesis through the secretion of numerous vasoactive
substances. Due to their barrier function, endothelial cells are
more vulnerable to injury by various physical and chemical risk
factors. Injury to endothelial cells is a critical event in
angiogenesis, atherosclerosis, thrombosis, hypertension and heart
failure (1–5). Endothelial dysfunction, particularly
endocrine dysfunction, causes the secretion of a variety of active
substances, which may lead to dysfunction of the cardiovascular
system.
In endothelial cells, oxidative stress is regarded
as a critical pathogenic factor for endothelial cell injury, and
the accumulation of ROS may result in endothelial cell apoptosis
(6–8). Hydrogen peroxide
(H2O2) may penetrate the plasma membrane and
cause endothelial cell injury. In addition, it has been reported
that reactive oxygen species (ROS) are involved in the apoptosis of
endothelial cells (9,10) and the primary source of endogenous
ROS is H2O2 (11), which has been extensively used to
induce oxidative stress in in vitro models (12,13).
Radix astragali is the root of the perennial
herb Astragalus membranaceus (14). As a well-established traditional
Chinese medicine, it has been used for the treatment of
cardiovascular disease and is believed to possess immune
stimulatory, antiviral and antioxidative effects. Astragalus
polysaccharide (APS) is an important bioactive ingredient obtained
from Astragalus membranaceus that has a range of
pharmacological effects, including increasing levels of cyclic
guanosine monophosphate (cGMP) and cyclic adenosine monophosphate
(cAMP) in plasma and tissues, promotion of immune responses,
anti-inflammation (15),
protection of vessels, antioxidant effects, anti-insulin resistance
and antitumor effects (16–19).
Our previous studies reported that APS may inhibit
isoprenaline-induced cardiac hypertrophy (20,21).
It has been reported that Astragalus membranaceus and its
primary components, including APS, may ameliorate endothelial
dysfunction induced by homocysteine in which antioxidation is
involved. However, the effects of APS on endothelial injury induced
by ROS and the underlying mechanism remains to be fully
elucidated.
Materials and methods
Materials
APS was purchased from Nanjing Jingzhu
Pharmaceutical Technology Co., Ltd. (Nanjing, China).
H2O2 was purchased from Tianjin Guangfu Fine
Chemical Research Institute (Tianjin, China). Dihydroethidium (DHE)
was obtained from Shanghai Haoran Bio-Technology Co., Ltd.
(Shanghai, China). MTT and dimethyl sulfoxide (DMSO) were purchased
from Sigma-Aldrich; Merck Millipore (Darmstadt, Germany). DAPI,
Rhodamine 123 and the Cell Cycle and Apoptosis analysis kit were
purchased from Beyotime Institute of Biotechnology (Haimen, China).
The cGMP assay kit was supplied by Nanjing Jiancheng Bioengineering
Institute (Nanjing, China). Rabbit anti-Copper-zinc superoxide
dismutase (Cu/Zn-SOD; bs-10216R) and anti-endothelial nitric oxide
synthase (eNOS; bs-20609R) antibodies were purchased from Beijing
Boosen Biological Technology Co., Ltd (Beijing, China), and
anti-β-actin (HRP-600008) was from Wuhan Sanying Biotechnology
(Wuhan, China). All other chemicals and reagents used were of
analytical grade.
Cell culture
Human umbilical vein endothelial cells (HUVECs) were
obtained from Shanghai Zhong Qiao Xin Zhou Biotechnology Co., Ltd.
(Shanghai, China) and were maintained in high-glucose Dulbecco's
Modified Eagle's medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (HyClone; GE Healthcare
Life Sciences, Logan, UT, USA), 2 mmol/l L-glutamine, 5 µg/ml
endothelial growth factor (Shanghai Zhong Qiao Xin Zhou
Biotechnology Co., Ltd.), 100 U/ml penicillin and 100 µg/ml
streptomycin. Cells were incubated in a humidified incubator with
5% CO2 at 37°C with media replenishment every 2 days and
were passaged at 80–90% confluence. The cells were treated with
various concentrations of APS or H2O2 for 24
h incubated in serum-free medium, prior to an MTT assay. Additional
groups were pretreated with various concentrations of APS for 1 h
prior to H2O2 exposure in fresh medium,
following which they were harvested for further analysis. The cells
were assigned into the following five groups; A, untreated control
group; B, H2O2 (400 µM) model group; C,
H2O2+APS (0.1 µg/ml) group; D,
H2O2+APS (1 µg/ml) group; E,
H2O2+APS (10 µg/ml) group.
Cell viability assay
The MTT assay was used to evaluate cell viability
(22). In brief, HUVECs were
cultured in a 96-well plate (1×104 cells/well). A total of 24 h
following plating, cells were pretreated with 0.1–100 µg/ml APS for
1 h, following which 400 µM H2O2 was added
into the plate and cultured at 37°C for a further 24 h. The
viability of cells treated with APS (0.1–100 µg/ml) or
H2O2 alone (0–500 µM) was also assessed. MTT
was dissolved in PBS, added to each well and incubated at 37°C for
4 h, for a final concentration of 0.5 mg/ml. Subsequently, the
culture medium was carefully removed and replaced with 150 µl DMSO.
The plates were incubated for 10 min and measured at a wavelength
of 490 nm on a microplate reader (DNM-9602G; Beijing Perlong New
Technology Co., Ltd., Beijing, China). The viability of HUVECs was
expressed as a percentage of the control group.
Apoptosis analysis by flow
cytometry
Apoptosis was evaluated using the Cell Cycle and
Apoptosis analysis kit. Briefly, following treatment, floating and
attached cells were harvested by trypsin and washed with PBS
(concentration, 1×106 cells/ml). Cells were fixed with ice-cold 70%
alcohol at 4°C overnight, and staining was performed with propidium
(PI) according to the manufacturer's protocol. Samples were
analyzed on a flow cytometer (BD Biosciences, Franklin Lakes, NJ,
USA) using CellQuest software v6.1x (BD Biosciences). Cells in the
sub-G1 peak were considered apoptotic.
DAPI staining
The morphological characteristics of nuclei were
evaluated by DAPI staining as previously described (23). HUVECs were seeded onto a 6-well
plate at a density of 1×105 cells/well. Following treatment, cells
were fixed with 4% paraformaldehyde for 10 min. Cells were washed
once with PBS and stained with 2 µg/ml DAPI for 15 min at 37°C in
the dark. The stained cells were visualized under an inverted
fluorescence microscope.
Determination of mitochondrial
membrane potential
The mitochondrial membrane potential of HUVECs was
assessed using the fluorescent dye Rhodamine 123 (24–26).
Rhodamine 123, a selective fluorescent dye, permeates cell
membranes and is sequestered by active mitochondria, therefore
staining the mitochondria of living cells, and is widely used for
the detection of mitochondrial membrane potential. Flavovirens
fluorescence represents normal cells. HUVECs were cultured in a
6-well plate at a density of 1×105 cells/well. Following treatment,
the culture medium was carefully removed and cells were washed
twice with PBS. The cells were subsequently stained with 2 µM
Rhodamine 123 for 20 min at 37°C in the dark. Cells were analyzed
under an inverted fluorescence microscope. The fluorescence
intensity was calculated with LAS Software (V4.3) (Leica
Microsystems GmbH, Wetzlar, Germany).
Measurement of intracellular cGMP
levels
HUVECs were pretreated with 0.1, 1 or 10 µg/ml APS
for 1 h, followed by exposure to 400 µM H2O2
for 24 h. The cells were subsequently harvested with a Falcon
scraper, centrifuged at 4°C for 5 min at 800 × g and sonicated. The
homogenate was prepared in PBS and centrifuged at 1,000 × g for 5
min at 4°C. The cGMP levels in the supernatants were determined
with a kit according to the manufacturer's protocol. cGMP levels
were expressed as nmol/g protein. Protein concentration was
calculated with a Bicinchoninic Acid (BCA) Protein assay kit
(Beijing Dingguo Changsheng Biotechnology Co., Ltd., Beijing,
China). cGMP is a second messenger of NO, and its level reflects
the level of nitric oxide (NO) (27).
Measurement of intracellular ROS
production
The production of ROS was measured using DHE. DHE,
an oxidative fluorescent dye, may be oxidized to ethidium bromide
and intercalated into DNA in the presence of superoxide anions.
HUVECs were seeded on a 6-well plate (1×105 cells/well). Following
treatment, cells were washed with PBS and incubated with 2 mM DHE
for 30 min at 37°C. Cells were washed with PBS and analyzed under
an inverted fluorescence microscope. The fluorescence intensity was
calculated using LAS Software V4.3.
Western blot analysis
Following treatment, cells were harvested and lysed
with ice-cold radio immunoprecipitation assay buffer (Wuhan Boster
Biotechnology Co., Ltd., Wuhan, China). Protein concentration was
determined using a BCA Protein assay kit. Protein extracts (30 µg)
were electrophoretically separated by 12 or 10% SDS-PAGE and
transferred onto polyvinylidene difluoride membranes. Membranes
were blocked with 5% nonfat milk powder and incubated overnight at
4°C with the following primary antibodies: Anti-eNOS (1:1,500),
anti-Cu/Zn-SOD (1:500) and β-actin (1:500). Following washing with
TBS containing Tween-20 (10 mM Tris, 100 mM NaCl, and 0.1%
Tween-20), membranes were incubated with horseradish
peroxidase-conjugated secondary antibodies (anti-rabbit IgG and
anti-mouse IgG; 1706515 and 1706516; 1:1,500; Bio-Rad Laboratories,
Inc., Hercules, CA, USA) for 2 h at room temperature. Membranes
were washed with TBS, and immunocomplexes were visualized using the
Amersham ECL start Western Blotting Detection Reagent enhanced
chemiluminescence system (GE Healthcare Life Sciences). β-actin
served as an internal control for the experiments. The results were
quantified by the Quantity One software v4.62 (Bio-Rad
Laboratories, Inc.).
Statistical analysis
All data are expressed as the mean ± standard
deviation. Each experiment was performed at least three times.
Statistical analysis was performed using Student's t-test or
one-way analysis of variance with SPSS software version 17.0 (SPSS
Inc., Chicago, IL, USA). The post hoc least significant difference
test was used to evaluate differences between two groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
Effects of APS on the viability of
HUVECs
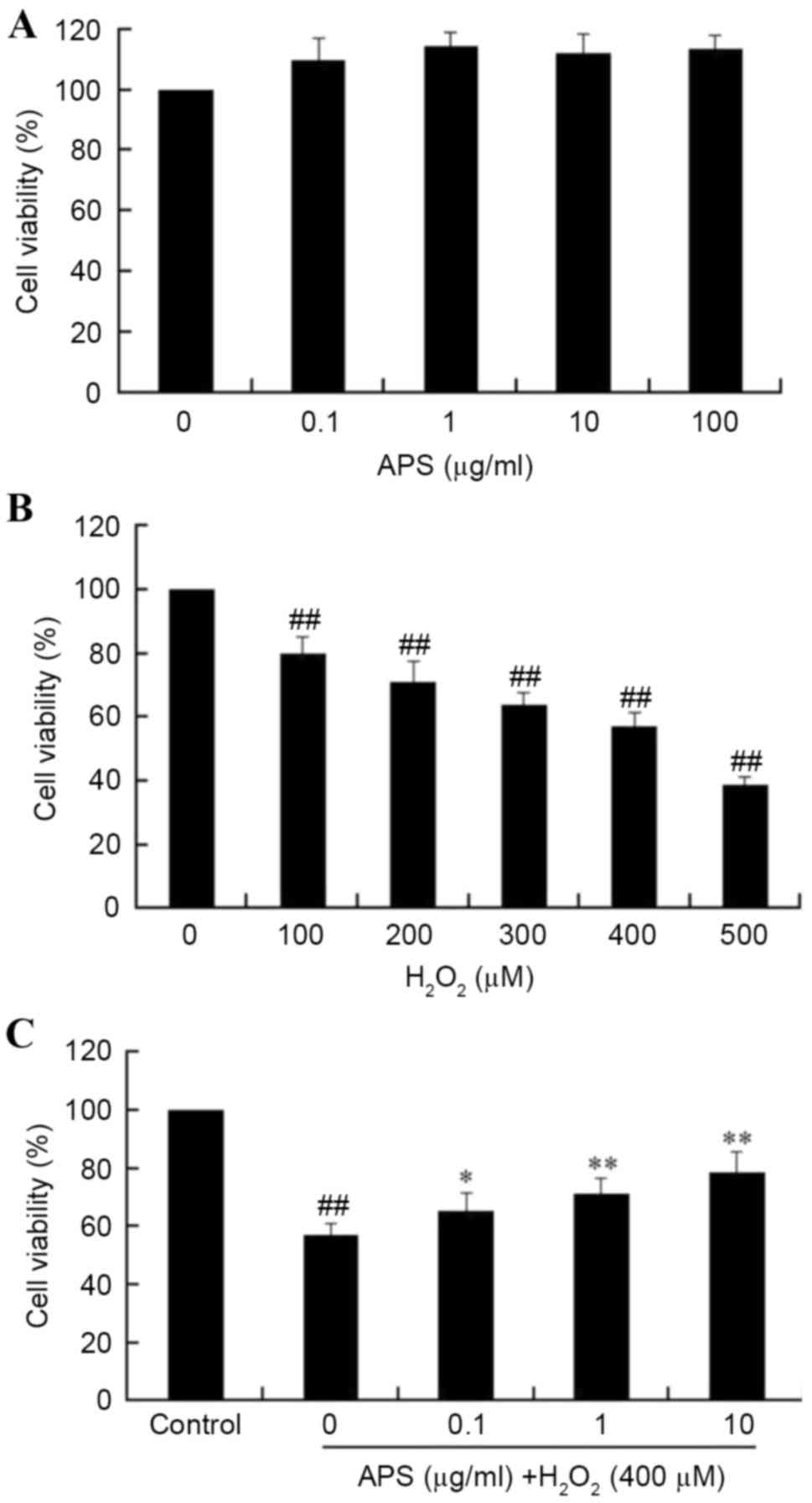
Initially, the cytotoxicity of APS on HUVECs was
measured using the MTT assay. As presented in Fig. 1A, incubation of HUVECs with 0.1–100
µg/ml APS for 24 h did not significantly affect the cell viability
(P>0.05). However, treatment of HUVECs with 100 to 500 µM
H2O2 for 24 h resulted in a
concentration-dependent decrease in cell viability, when compared
with the control group (P<0.05; Fig. 1B). Pretreatment of HUVECs with 0.1,
1 or 10 µg/ml APS for 1 h prior to exposure to
H2O2 significantly increased viability in a
dose-dependent manner (Fig. 1C).
These results indicated that APS may protect HUVECs from oxidative
stress-induced injury. Based on these results, further studies
employed a H2O2 concentration of 400 µM, and
APS concentrations of 0.1, 1.0 and 10 µg/ml.
Effect of APS on
H2O2-induced apoptosis in HUVECs
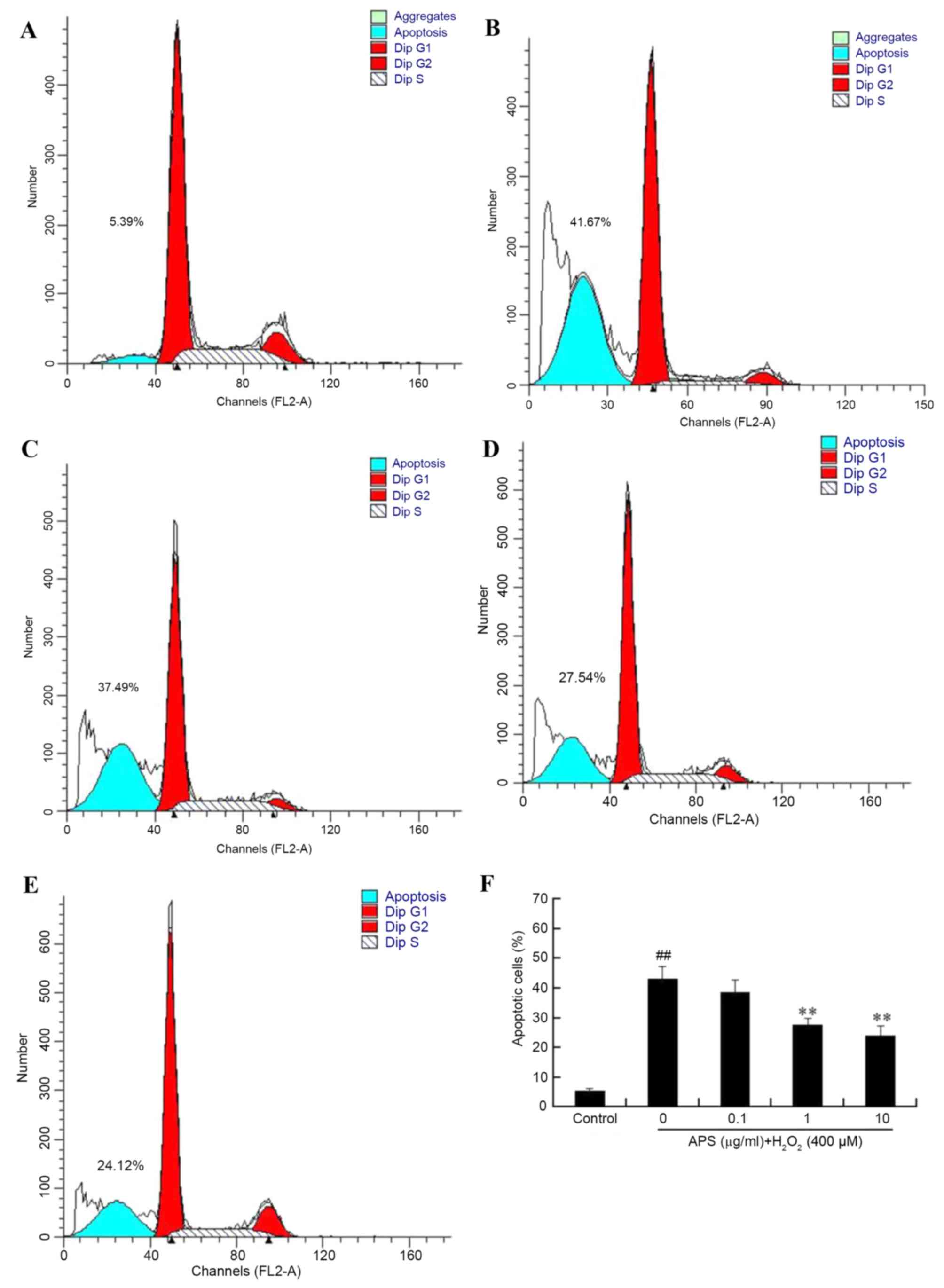
To further evaluate the protective effect of APS
against H2O2-induced damage in HUVECs,
apoptosis rates were measured by flow cytometry. As presented in
Fig. 2, the percentage of
apoptotic cells in the control group was 5.44±0.57%. Following
exposure to 400 µM H2O2 for 24 h, the
percentage of apoptosis increased to 43.14±4.25%. However,
pretreatment of the cells with 0.1, 1 and 10 µg/ml APS for 1 h
prior to H2O2 exposure reduced the percentage
of apoptotic cells to 38.58±4.13, 27.60±2.25 and 24.10±3.26%,
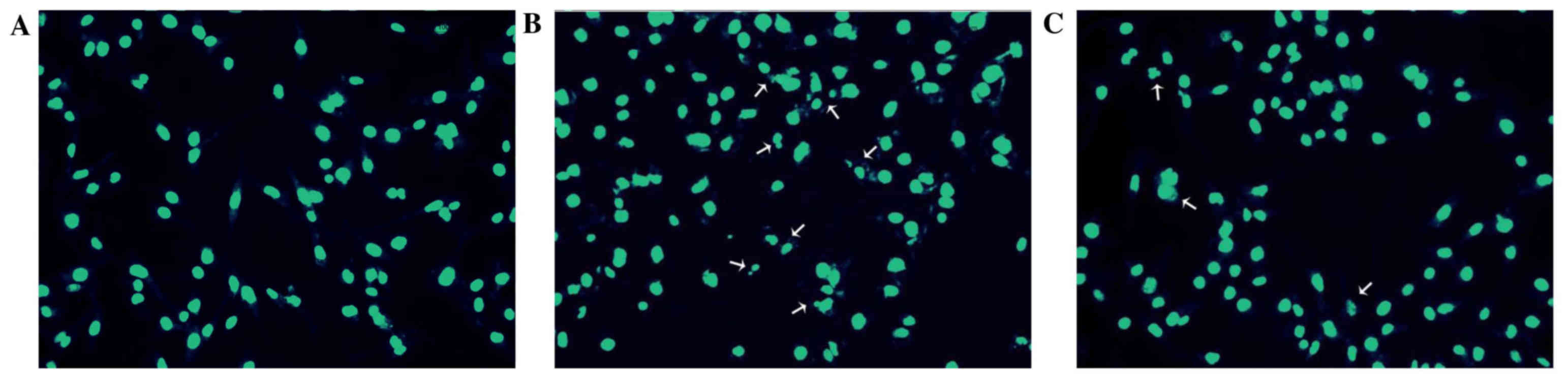
respectively. In addition, the morphology of HUVEC nuclei was
evaluated by DAPI staining. Compared with the control group
(Fig. 3A), nuclei of the
H2O2 group (Fig.
3B) were shrunken, irregular and fragmented, typical features
of apoptosis. However, APS (1 µg/ml) pretreatment (Fig. 3C) reduced these alterations.
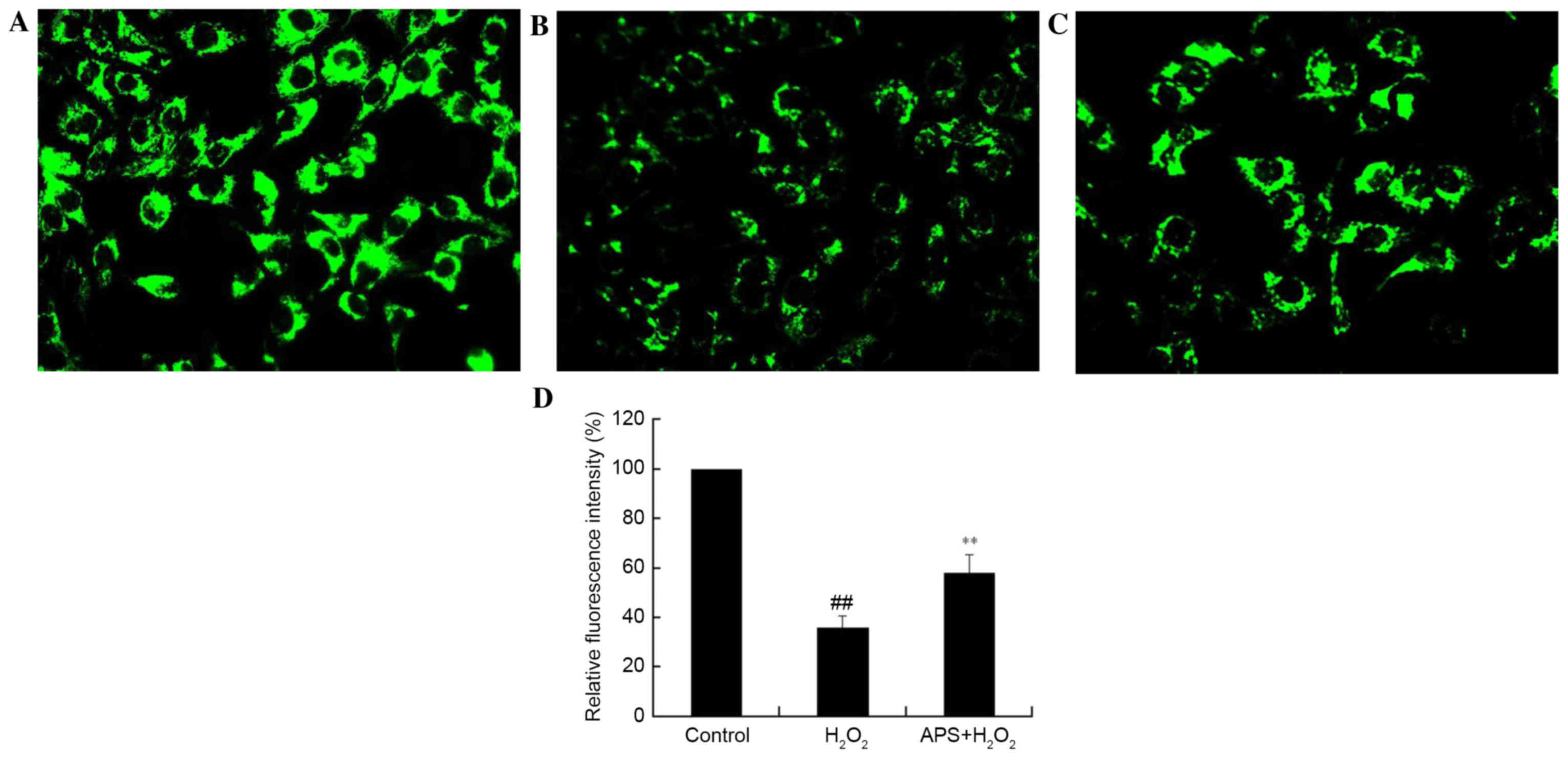
Effect of APS on the mitochondrial
membrane potential of HUVECs
The disruption of mitochondria has been reported to
be involved in programmed cell death. The maintenance of the
mitochondrial membrane potential is essential for mitochondrial
integrity and bioenergetic function. To determine whether the
anti-apoptotic effect of APS is associated with the inhibition of
mitochondrial disruption, mitochondrial membrane potential
variation in HUVECs was assessed by Rhodamine 123 staining.
Rhodamine 123 is sequestered by active mitochondria of normal
HUVECs and revealed yellow-green flavovirens fluorescence. Compared
with control cells (Fig. 4A),
H2O2-treated cells presented a weak green
fluorescence intensity, which reflected the loss of mitochondrial
membrane potential (Fig. 4B); this
decrease in fluorescence was partially reversed by APS (1 µg/ml)
pretreatment (Fig. 4C).
Quantification of fluorescence intensity revealed that these
alterations were significant (Fig.
4D). This may be a possible underlying mechanism by which APS
protects against injury.
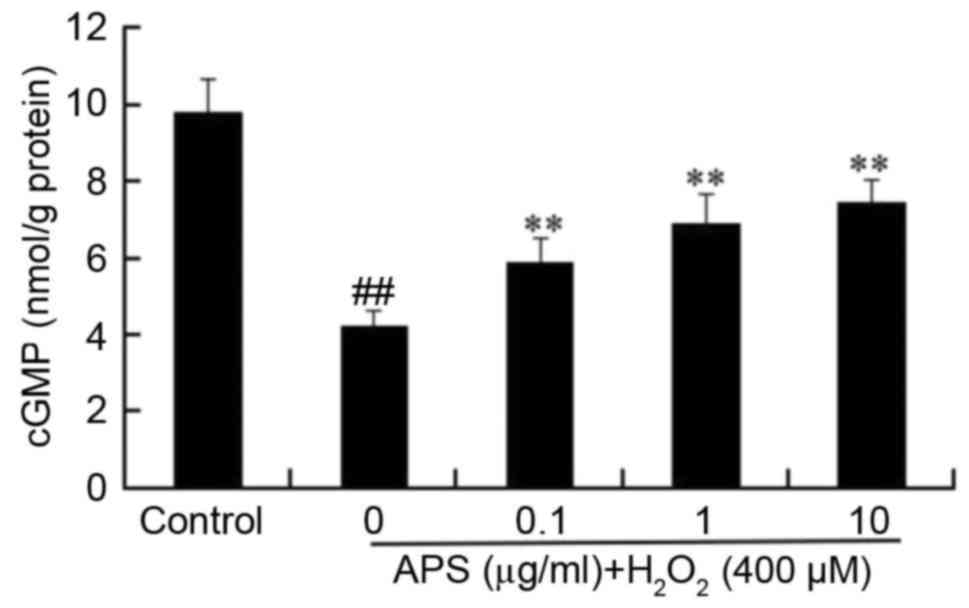
Effect of APS on cGMP levels in
HUVECs
To further investigate the protective effects of
APS, the influence of H2O2 on cGMP
accumulation was measured. As presented in Fig. 5, basal production of cGMP decreased
from 9.8±0.8 nmol/g protein in control cells to 4.2±0.4 nmol/g
protein in H2O2-treated cells. However,
pretreatment of cells with 0.1–10 µg/ml APS for 1 h prior to
H2O2 exposure increased the levels of cGMP
compared with H2O2 treatment alone, in a
dose-dependent manner.
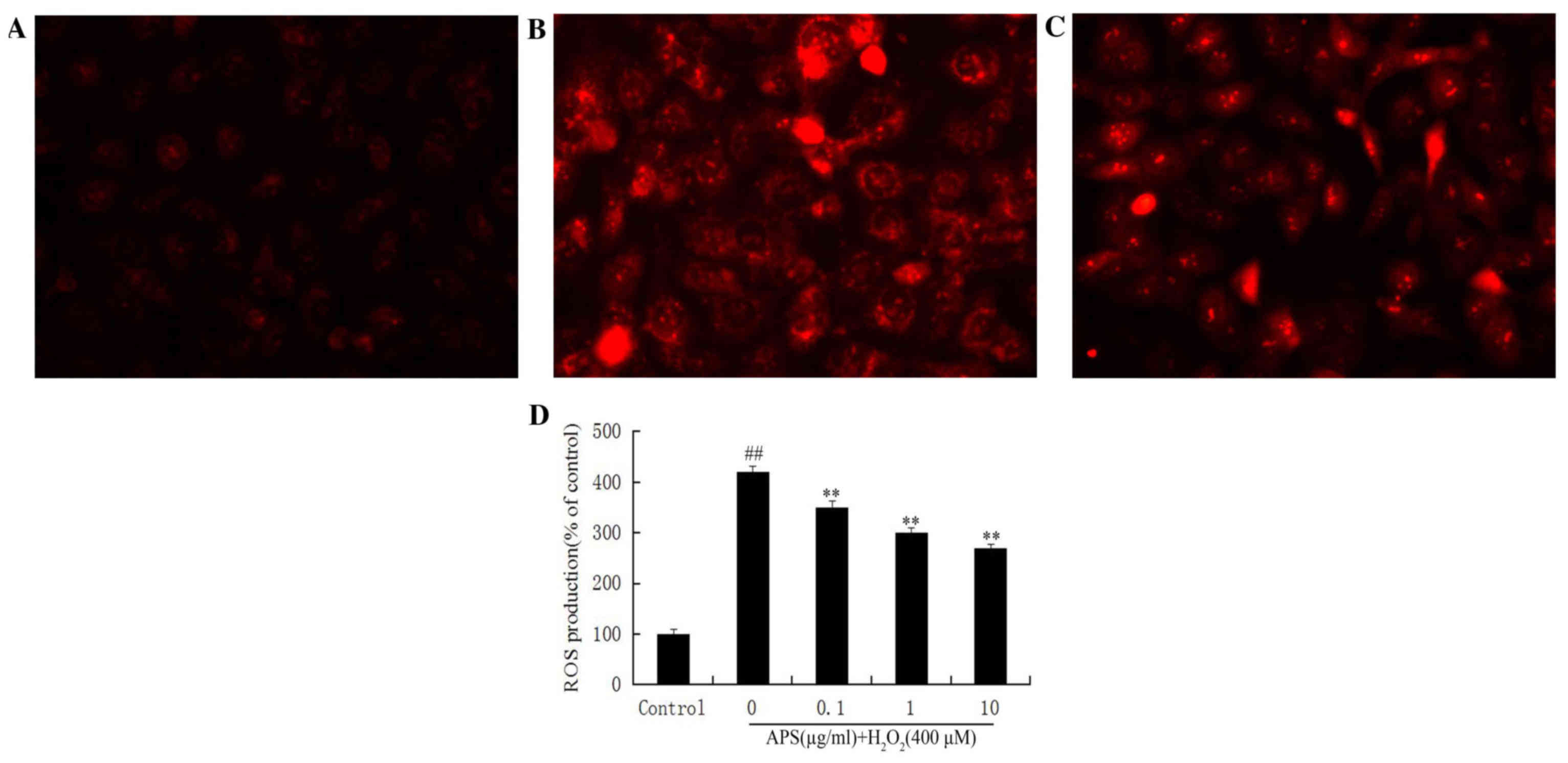
Effect of APS on ROS formation in
HUVECs
The secondary generation of ROS is another potential
mechanism underlying cell damage induced by
H2O2. Therefore, intracellular ROS levels
were measured by DHE staining. Compared with the control group
(Fig. 6A), DHE fluorescence in
HUVECs exposed to H2O2 was increased
(Fig. 6B). This increase was
attenuated by pretreatment with APS (Fig. 6C). Quantification of fluorescence
revealed significantly increased ROS production in
H2O2-treated cells (P<0.01), and a
significant and dose-dependent effect of APS (P<0.01; Fig. 6D).
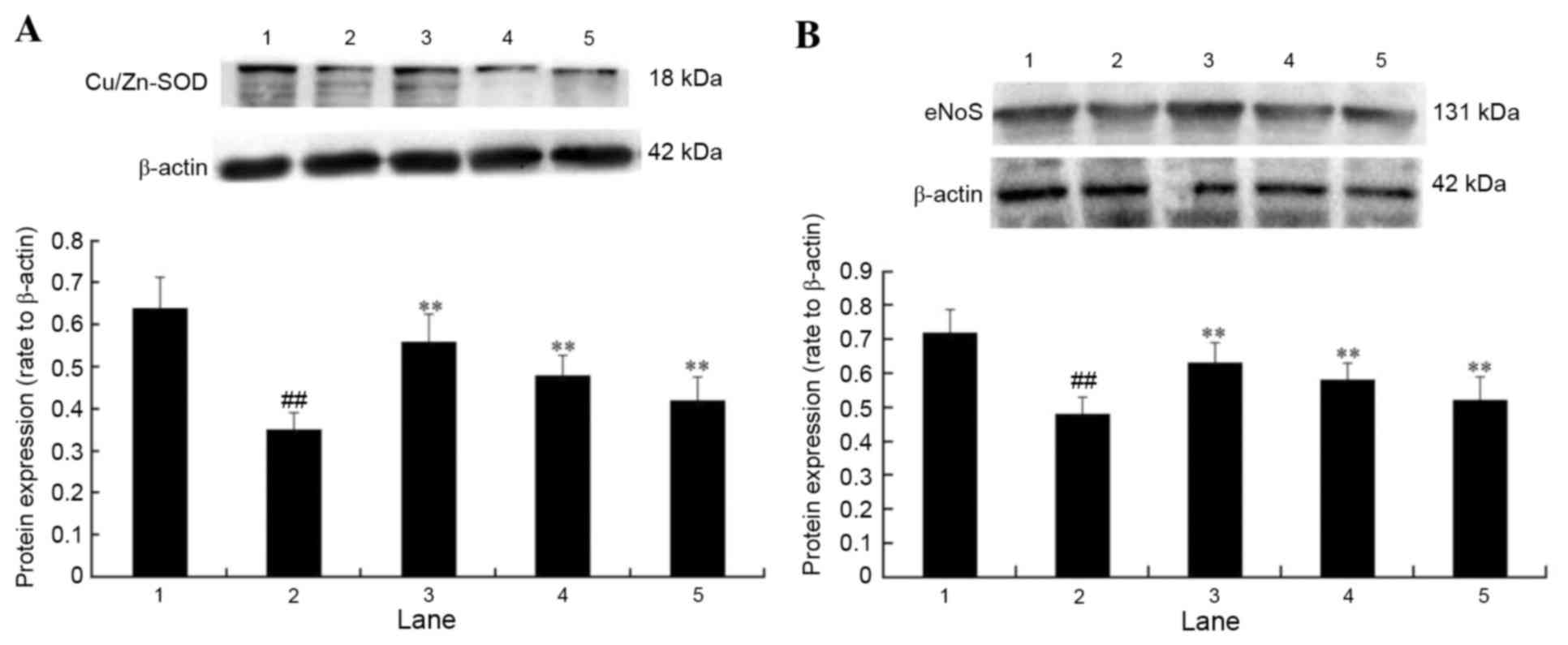
Effect of APS on the protein
expression levels of eNOS and Cu/Zn-SOD in
H2O2-treated cells
The protein expression levels of eNOS and Cu/Zn-SOD
were examined to investigate the potential underlying molecular
mechanisms contributing to APS cytoprotection. Protein expression
levels of Cu/Zn-SOD (Fig. 7A) and
eNOS (Fig. 7B) in HUVECs exposed
to H2O2 were significantly decreased
(P<0.01). However, pretreatment with 10, 1 or 0.1 µg/ml APS
significantly attenuated the H2O2-mediated
decrease of eNOS and Cu/Zn-SOD expression (P<0.01), which may
therefore contribute to the APS-mediated protective effects of
HUVECs.
Discussion
Vascular endothelial cells form a crucial functional
barrier between tissues and the circulation, which acts as a
receptor that senses physical or chemical stimuli occurring inside
the vessel. In addition, this barrier may secrete a variety of
substances that contribute to the response of the blood vessel to
different physiologic and pathologic stimuli, including regulation
of coagulation and participation in the immune response.
Abnormalities in endothelial cell structure and function
potentially lead to numerous cardiovascular diseases. Therefore,
endothelial function has been identified as a biomarker/mediator of
cardiovascular risk factors (28–30)
that may act as an independent predictor of cardiovascular disease
(3,31). APS, isolated from Astragalus
membranaceus, has a variety of pharmacological effects,
including anti-inflammatory, antiviral and antioxidant effects, and
it increases levels of cGMP and cAMP in plasma and tissues
(32). It has been reported that
APS has cytoprotective effects on the erythroid lineage K562 cells
(33). In addition, APS may
improve the response of C2C12 skeletal muscle myotubes and
myoblasts to peroxide-induced injury in vitro by inhibiting
apoptosis (34). However, the
effect and underlying mechanisms of APS on
H2O2-induced injury in HUVECs remain to be
elucidated.
In the present study, the effects of APS on
H2O2-induced injury in HUVECs in vitro
and the possible underlying mechanisms were investigated. The
results indicated that APS protected HUVECs from
H2O2-induced apoptosis. Furthermore, APS
markedly decreased intracellular ROS levels, increased protein
expression levels of eNOS and Cu/Zn-SOD, partially restored the
mitochondrial membrane potential and increased levels of cGMP,
compared with cells treated with H2O2 alone.
The underlying mechanisms of APS against HUVEC apoptosis may
involve an increase in antioxidant defense systems and an increase
in the production and bioavailability of NO.
NO, an endothelium-dependent vasodilator, is an
important mediator in the regulation of endothelial cell function.
In addition, it has numerous biochemical activities, including
directly scavenging superoxide, maintaining endothelial integrity,
attenuating leukocyte adhesion and inhibiting platelet aggregation
(5). Endothelial NO produced by
eNOS may inhibit apoptosis, and is regarded as a survival factor
for endothelial cells (35). It
has been reported that the reduced formation of NO or impairment of
NO effects may be associated with aortic sclerosis (36). Additional studies have indicated
that reduced NO bioavailability enhances atherogenesis in animal
models (8), and contributes to
endothelial dysfunction (37). In
the current study, the intracellular cGMP levels were detected to
evaluate the bioavailability of NO, as it is the second messenger
of NO. The results suggested that APS pretreatment may increase the
levels of cGMP and the protein expression levels of eNOS, compared
with cells treated with H2O2 alone. This
indicated that APS may protect HUVECs from injury induced by
H2O2 via increasing the production and
bioavailability of NO.
ROS, the free radicals present in all vascular
cells, have been demonstrated to serve an important role in
endothelial injury (38). Numerous
studies have suggested that ROS are involved in vascular remodeling
and endothelial dysfunction (39).
Endothelial cells express a variety of enzymes from which ROS may
be generated. Therefore, endothelial cells are regarded as an
important source of vascular ROS production (40). In addition, mitochondria are
important physiological sources of ROS. Furthermore, it has been
reported that H2O2 may cause endothelial cell
injury by inducing mitochondrial dysfunction, including loss of
mitochondrial membrane potential (41).
Reduced bioavailability of NO contributes to
endothelial dysfunction. The overproduction of ROS may inactivate
NO by reacting with it to decrease NO synthesis and reduce NO
bioavailability. Previous studies have suggested that the imbalance
between ROS and NO levels, rather than the levels of each, may be a
primary cause of endothelial dysfunction in numerous cardiovascular
diseases. The results of the present study suggested that APS may
protect endothelial cells against injury induced by
H2O2 via increasing the protein expression
levels of Cu/Zn-SOD. This may lead to superoxide scavenging,
restoring mitochondrial membrane potential and decreasing the
intracellular ROS levels detected by DHE staining.
In conclusion, the present study demonstrated that
APS had a protective effect against
H2O2-induced injury and apoptosis in HUVECs.
The underlying mechanisms of the protective effects of APS may
involve restoration of the balance between ROS and NO levels via
increasing the cell antioxidant capacity and NO
bioavailability.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81374008).
Glossary
Abbreviations
Abbreviations:
|
APS
|
Astragalus polysaccharide
|
|
ROS
|
reactive oxygen species
|
|
HUVECs
|
human umbilical vein endothelial
cells
|
References
|
1
|
Karsan A and Harlan JM: Modulation of
endothelial cell apoptosis: Mechanisms and pathophysiological
roles. J Atheroscler Thromb. 3:75–80. 1996. View Article : Google Scholar
|
|
2
|
Harrison D, Griendling KK, Landmesser U,
Hornig B and Drexler H: Role of oxidative stress in
atherosclerosis. Am J Cardiol. 91:7A–11A. 2003. View Article : Google Scholar
|
|
3
|
Szymanski MK, Buikema JH, Van Veldhuisen
DJ, Koster J, van der Velden J, Hamdani N, Hillege JL and
Schoemaker RG: Increased cardiovascular risk in rats with primary
renal dysfunction; mediating role for vascular endothelial
function. Basic Res Cardiol. 107:2422012. View Article : Google Scholar :
|
|
4
|
Li JM and Shah AM: Endothelial cell
superoxide generation: Regulation and relevance for cardiovascular
pathophysiology. Am J Physiol Regul Integr Comp Physiol.
287:R1014–R1030. 2004. View Article : Google Scholar
|
|
5
|
Loscalzo J: Nitric oxide insufficiency,
platelet activation, and arterial thrombosis. Circ Res. 88:756–762.
2001. View Article : Google Scholar
|
|
6
|
Birukov KG: Cyclic stretch, reactive
oxygen species, and vascular remodeling. Antioxid Redox Signal.
11:1651–1667. 2009. View Article : Google Scholar :
|
|
7
|
Urso C and Caimi G: Oxidative stress and
endothelial dysfunction. Minerva Med. 102:59–77. 2011.
|
|
8
|
Lu X, Dang CQ, Guo X, Molloi S, Wassall
CD, Kemple MD and Kassab GS: Elevated oxidative stress and
endothelial dysfunction in right coronary artery of right
ventricular hypertrophy. J Appl Physiol (1985). 110:1674–1681.
2011. View Article : Google Scholar :
|
|
9
|
Lee YJ, Kang IJ, Bünger R and Kang YH:
Mechanisms of pyruvate inhibition of oxidant-induced apoptosis in
human endothelial cells. Microvasc Res. 66:91–101. 2003. View Article : Google Scholar
|
|
10
|
Maio R, Perticone M, Sciacqua A, Tassone
EJ, Naccarato P, Bagnato C, Iannopollo G, Sesti G and Perticone F:
Oxidative stress impairs endothelial function in nondipper
hypertensive patients. Cardiovasc Ther. 30:85–92. 2012. View Article : Google Scholar
|
|
11
|
Nohl H, Kozlov AV, Gille L and Staniek K:
Cell respiration and formation of reactive oxygen species: Facts
and artefacts. Biochem Soc Trans. 31:1308–1311. 2003. View Article : Google Scholar
|
|
12
|
Yang B, Oo TN and Rizzo V: Lipid rafts
mediate H2O2 prosurvival effects in cultured endothelial cells.
FASEB J. 20:1501–1503. 2006. View Article : Google Scholar
|
|
13
|
Wang YK, Hong YJ, Wei M, Wu Y, Huang ZQ,
Chen RZ and Chen HZ: Curculigoside attenuates human umbilical vein
endothelial cell injury induced by H2O2. J Ethnopharmacol.
132:233–239. 2010. View Article : Google Scholar
|
|
14
|
Liu J, Chen HB, Guo BL, Zhao ZZ, Liang ZT
and Yi T: Study of the relationship between genetics and geography
in determining the quality of Astragali Radix. Biol Pharm Bull.
34:1404–1412. 2011. View Article : Google Scholar
|
|
15
|
Yang J, Wang HX, Zhang YJ, Lu ML, Zhang J,
Li ST, Zhang SP and Li G: Astragaloside IV attenuates inflammatory
cytokines by inhibiting TLR4/NF-κB signaling pathway in
isoproterenol-induced myocardial hypertrophy. J Ethnopharmacol.
150:1062–1070. 2013. View Article : Google Scholar
|
|
16
|
Wang D, Hu Y, Sun J, Kong X, Zhang B and
Liu J: Comparative study on adjuvanticity of compound Chinese
herbal medicinal ingredients. Vaccine. 23:3704–3708. 2005.
View Article : Google Scholar
|
|
17
|
Huang WM, Liang YQ, Tang LJ, Ding Y and
Wang XH: Antioxidant and anti-inflammatory effects of Astragalus
polysaccharide on EA.hy926 cells. Exp Ther Med. 6:199–203.
2013.
|
|
18
|
Lu J, Chen X, Zhang Y, Xu J, Zhang L, Li
Z, Liu W, Ouyang J, Han S and He X: Astragalus polysaccharide
induces anti-inflammatory effects dependent on AMPK activity in
palmitate-treated RAW264.7 cells. Int J Mol Med. 31:1463–1470.
2013.
|
|
19
|
Shao BM, Xu W, Dai H, Tu P, Li Z and Gao
XM: A study on the immune receptors for polysaccharides from the
roots of Astragalus membranaceus, a Chinese medicinal herb. Biochem
Biophys Res Commun. 320:1103–1111. 2004. View Article : Google Scholar
|
|
20
|
Luan A, Tang F, Yang Y, Lu M, Wang H and
Zhang Y: Astragalus polysaccharide attenuates isoproterenol-induced
cardiac hypertrophy by regulating TNF-α/PGC-1α signaling mediated
energy biosynthesis. Environ Toxicol Pharmacol. 39:1081–1090. 2015.
View Article : Google Scholar
|
|
21
|
Dai H, Jia G, Liu X, Liu Z and Wang H:
Astragalus polysaccharide inhibits isoprenaline-induced cardiac
hypertrophy via suppressing Ca2+-mediated calcineurin/NFATc3 and
CaMKII signaling cascades. Environ Toxicol Pharmacol. 38:263–271.
2014. View Article : Google Scholar
|
|
22
|
Li ZL, Liu JC, Hu J, Li XQ, Wang SW, Yi DH
and Zhao MG: Protective effects of hyperoside against human
umbilical vein endothelial cell damage induced by hydrogen
peroxide. J Ethnopharmacol. 139:388–394. 2012. View Article : Google Scholar
|
|
23
|
Gong G, Qin Y, Huang W, Zhou S, Yang X and
Li D: Rutin inhibits hydrogen peroxide-induced apoptosis through
regulating reactive oxygen species mediated mitochondrial
dysfunction pathway in human umbilical vein endothelial cells. Eur
J Pharmacol. 628:27–35. 2010. View Article : Google Scholar
|
|
24
|
Ishikawa K, Takenaga K, Akimoto M,
Koshikawa N, Yamaguchi A, Imanishi H, Nakada K, Honma Y and Hayashi
J: ROS-generating mitochondrial DNA mutations can regulate tumor
cell metastasis. Science. 320:661–664. 2008. View Article : Google Scholar
|
|
25
|
Gonda K, Tsuchiya H, Sakabe T, Akechi Y,
Ikeda R, Nishio R, Terabayashi K, Ishii K, Matsumi Y, Ashla AA, et
al: Synthetic retinoid CD437 induces mitochondria-mediated
apoptosis in hepatocellular carcinoma cells. Biochem Biophys Res
Commun. 370:629–633. 2008. View Article : Google Scholar
|
|
26
|
Han YS and Lee CS: Antidepressants reveal
differential effect against 1-methyl-4-phenylpyridinium toxicity in
differentiated PC12 cells. Eur J Pharmacol. 604:36–44. 2009.
View Article : Google Scholar
|
|
27
|
Calderone A, Thaik CM, Takahashi N, Chang
DL and Colucci WS: Nitric oxide, atrial natriuretic peptide, and
cyclic GMP inhibit the growth-promoting effects of norepinephrine
in cardiac myocytes and fibroblasts. J Clin Invest. 101:812–818.
1998. View Article : Google Scholar :
|
|
28
|
Bleske BE, Hwang HS, Zineh I, Ghannam MG
and Boluyt MO: Evaluation of immunomodulatory biomarkers in a
pressure overload model of heart failure. Pharmacotherapy.
27:504–509. 2007. View Article : Google Scholar
|
|
29
|
Cai H and Harrison DG: Endothelial
dysfunction in cardiovascular diseases: The role of oxidant stress.
Circ Res. 87:840–844. 2000. View Article : Google Scholar
|
|
30
|
Vita JA and Keaney JF Jr: Endothelial
function: A barometer for cardiovascular risk? Circulation.
106:640–642. 2002. View Article : Google Scholar
|
|
31
|
Poulikakos D, Ross L, Recio-Mayoral A,
Cole D, Andoh J, Chitalia N, Sharma R, Kaski J Carlos and Banerjee
D: Left ventricular hypertrophy and endothelial dysfunction in
chronic kidney disease. Eur Heart J Cardiovasc Imaging. 15:56–61.
2014. View Article : Google Scholar
|
|
32
|
Zhang BQ, Hu SJ, Qiu LH, Zhu JH, Xie XJ,
Sun J, Zhu ZH, Xia Q and Bian K: Effects of Astragalus membranaceus
and its main components on the acute phase endothelial dysfunction
induced by homocysteine. Vascul Pharmacol. 46:278–285. 2007.
View Article : Google Scholar
|
|
33
|
Yang M, Qian XH, Zhao DH and Fu SZ:
Effects of Astragalus polysaccharide on the erythroid lineage and
microarray analysis in K562 cells. J Ethnopharmacol. 127:242–250.
2010. View Article : Google Scholar
|
|
34
|
Lu L, Wang DT, Shi Y, Yin Y, Wei LB, Zou
YC, Huang B, Zhao Y, Wang M, Wan H, et al: Astragalus
polysaccharide improves muscle atrophy from dexamethasone- and
peroxide-induced injury in vitro. Int J Biol Macromol. 61:7–16.
2013. View Article : Google Scholar
|
|
35
|
Dimmeler S and Zeiher AM: Nitric oxide-an
endothelial cell survival factor. Cell Death Differ. 6:964–968.
1999. View Article : Google Scholar
|
|
36
|
Nightingale AK, Sverdlov AL, Rajendran S,
Mishra K, Heresztyn T, Ngo DT and Horowitz JD: Lack of association
between aortic sclerosis and left ventricular hypertrophy in
elderly subjects. Int J Cardiol. 150:33–38. 2011. View Article : Google Scholar
|
|
37
|
Clapp BR, Hingorani AD, Kharbanda RK,
Mohamed-Ali V, Stephens JW, Vallance P and MacAllister RJ:
Inflammation-induced endothelial dysfunction involves reduced
nitric oxide bioavailability and increased oxidant stress.
Cardiovasc Res. 64:172–178. 2004. View Article : Google Scholar
|
|
38
|
Zorov DB, Filburn CR, Klotz LO, Zweier JL
and Sollott SJ: Reactive oxygen species (ROS)-induced ROS release:
A new phenomenon accompanying induction of the mitochondrial
permeability transition in cardiac myocytes. J Exp Med.
192:1001–1014. 2000. View Article : Google Scholar :
|
|
39
|
Touyz RM and Schiffrin EL: Reactive oxygen
species in vascular biology: Implications in hypertension.
Histochem Cell Biol. 122:339–352. 2004. View Article : Google Scholar
|
|
40
|
Cai H: Hydrogen peroxide regulation of
endothelial function: Origins, mechanisms and consequences.
Cardiovasc Res. 68:26–36. 2005. View Article : Google Scholar
|
|
41
|
Liu YM, Jiang B, Bao YM and An LJ:
Protocatechuic acid inhibits apoptosis by mitochondrial dysfunction
in rotenone-induced PC12 cells. Toxicol In Vitro. 22:430–437. 2008.
View Article : Google Scholar
|