Introduction
Breast cancer remains the most prevalent cancer in
females around the world today (1). In 2012, breast cancer was ranked
first in cancer incidence and was the leading cause of
cancer-associated mortalities among females worldwide, with an
estimated 1.7 million newly diagnosed cases and 521,900 mortalities
worldwide (2). Despite the
advances made in breast cancer management, including earlier
detection and more effective treatments strategies over the past
few decades, ~5 to 10% of females have metastatic disease when
first diagnosed with breast cancer, of which only one-fifth survive
5 years (3,4). Therefore, identifying novel molecular
correlates with tumorigenesis and development of breast cancer may
be helpful for its earlier diagnosis and treatment, so as to
improve the prognosis of breast cancer patients.
MicroRNAs (miRNAs/miRs) are small, endogenous,
noncoding RNAs that are ~22 nucleotides in length, and regulate
gene expression by destabilizing target mRNAs as well as inhibiting
their translation (5,6). Mechanistically, miRNAs function by
directly binding to the 3′ untranslated region (UTR) of its target
mRNA (7). An increasing number of
studies have demonstrated that miRNAs may serve as oncogenes or
tumor suppressors in human cancers, and aberrant expression of
miRNAs has been observed in various human cancers, including lung
(8), gastric (9) and prostate (10). Similarly, the role of miRNAs in the
initiation and development of breast cancer has been prevalently
investigated, including proliferation (11), apoptosis (12), migration and invasion (13). Therefore, various miRNAs may have
the potential to serve as diagnostic markers, prognosis predictors
and therapeutic targets in breast cancer patients.
It was reported that miR-372 belongs to the
Mir-371-372 gene cluster, which is located on chromosome 19q13.42
(14). Accumulating evidence has
suggested that miR-372 may regulate cell proliferation, apoptosis,
migration and invasion in many types of human cancers, and it may
serve oncogenic or suppressive roles in different human cancers.
For example, Cho et al (15) demonstrated the oncogenic role of
miR-372 in gastric cancer by controlling cell growth, the cell
cycle and apoptosis through downregulation of a tumor suppressor
gene, large tumor suppressor kinase 2. Chen et al (16) reported that miR-372 may function as
an oncogenic miRNA by directly targeting the 3′-UTR of PH domain
and leucine rich repeat protein phosphatase 2 in glioma. On the
contrary, Huang et al (17)
found that miRNA-372 was downregulated in renal cell carcinoma cell
lines and tissue specimens, and its over-expression inhibited cell
proliferation and invasion by suppressing IGF2BP1. Furthermore, Liu
et al (18) demonstrated
that miR-372 suppressed cell proliferation, migration, and
invasion, and promoted the apoptosis of endometrial carcinoma cells
through downregulating RhoC. However, the role of miR-372 in breast
cancer remains unknown.
To the best of our knowledge, the present study was
the first to investigate the expression level of miR-372 and its
role in breast cancer. Firstly, miR-372 expression levels in human
breast cancer tissues and cell lines were detected by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR).
Next, in vitro assays were conducted to investigate the
effect of miR-372 on breast cancer cell proliferation and
apoptosis. Finally, E2F1 was identified as a direct target of
miR-372 for its tumor suppressive role in breast cancer. These
results demonstrated that miR-372 inhibits proliferation and
induces apoptosis in breast cancer by directly targeting E2F1, and
may serve as a therapeutic target for the treatment of breast
cancer patients.
Materials and methods
Tissue specimens
A total of 20 paired clinical tissue specimens
(tumor and adjacent non-tumor tissues) were collected from patients
who were diagnosed with primary breast cancer and underwent surgery
in the Department of General Surgery, The Second Affiliated
Hospital and Yuying Children's Hospital of Wenzhou Medical
University (Wenzhou, China) between May and October 2015. All
tissue specimens were frozen in liquid nitrogen immediately and
stored at −80°C until use. Both tumor and non-tumor tissues were
confirmed histologically. No patients underwent radiation therapy
or chemotherapy prior to surgery. Written informed consent was
obtained from each participant and this study was approved by the
Ethics Committees of The Second Affiliated Hospital and Yuying
Children's Hospital of Wenzhou Medical University.
Cell lines
The BT-474, MCF-7, MDA-MB-436 and MDA-MB-231 human
breast cancer cell lines were purchased from the Shanghai Institute
of Cell Biology, Chinese Academy of Sciences (Shanghai, China). The
MCF10A healthy breast cell line was purchased from American Type
Culture Collection (Manassas, VA, USA). All breast cancer cell
lines were cultured in Dulbecco's modified Eagle's medium (DMEM;
Gibco; Thermo Fisher Scientific, Waltham, MA, USA) supplemented
with 10% fetal bovine serum (FBS, Gibco; Thermo Fisher Scientific,
Inc.), 100 µg/ml penicillin and 100 µg/ml streptomycin (Invitrogen;
Thermo Fisher Scientific, Inc.). MCF-10A cells were cultured in
DMEM/F12 (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
5% horse serum (Gibco; Thermo Fisher Scientific, Inc.), 20 ng/ml
EGF (Peprotech, Inc. Rocky Hill, NJ, USA), 0.5 mg/ml hydrocortisone
(Stemcell Technologies, Inc., Vancouver, BC, Canada), 10 µg/ml
insulin (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and 100
ng/ml cholera toxin (Sigma-Aldrich; Merck MGaA). Cells were
maintained at 37°C in a humidified atmosphere containing 5%
CO2.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from tissue samples or cell
lines using TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Waltham, MA, USA) according to the manufacturer's protocol and then
reverse transcribed into cDNA using a reverse transcription kit
(Promega Corporation, Madison, WI, USA). For E2F1 mRNA, qPCR was
performed using a SYBR Green Master Mix (Takara Bio, Inc., Otsu,
Japan) and GAPDH served as an internal control. The RT-qPCR
conditions were 95°C for 3 min; 40 cycles of 95°C for 12 sec, and
62°C for 1 min. For miR-372, TaqMan assays (Applied Biosystems;
Thermo Fisher Scientific, Inc.) were performed following the
manufacturer's protocol and small nuclear U6 RNA served as an
internal control. The relative expression levels were normalized to
internal controls using the comparative 2−ΔΔCq method.
(19) Primers for miR-372 were as
follows: Forward, 5′-ACACTCCAGCTGGGAAAGTGCTGCGACATTT-3′ and
reverse, 5′-GTGCAGGGTCCGAGGT-3′. Primers for E2F1 were as follows:
Forward, 5′-CCCATCCCAGGAGGTCACTT-3′ and reverse,
5′-CTGCAGGCTCACTGCTCTC-3′. All experiments were conducted in
triplicate.
Cell transfection
All transfections were performed using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacture's protocol. miR-372
mimics (miR-372), miR-372 inhibitors (anti-miR-383) and their
negative controls (miR-NC and anti-miR-NC) were purchased from
Guangzhou RiboBio Co., Ltd. (Guangzhou, China). Small interfering
(si)RNA targeting human E2F1 mRNA (si-E2F1) and the scramble
(si-NC) were designed by Shanghai GenePharma Co., Ltd. (Shanghai,
China). At 48 h after transfection, cells were collected and
RT-qPCR was conducted to confirm the transfection efficiency. All
assays were conducted in triplicate.
MTT assay
Cell proliferation was analyzed by MTT assay
according to the manufacturer's protocol. Briefly, cells were
seeded into 96-well plates at a density of 1×104 cells
per well and incubated for 0, 24, 48, 72 or 96 h. Following this,
100 µl MTT (5 mg/ml) was added to each well and incubated for 4 h.
Next, 150 µl dimethyl sulfoxide was added to halt the reaction. A
microplate was used to determine the absorbance at a wavelength of
490 nm. Each assay was performed in triplicate and repeated three
times.
Flow cytometry
Cell apoptosis was analyzed by an Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis
detection kit (Nanjing KeyGen Biotech, Co., Ltd., Nanjing, China)
following the manufacturer's protocol. After transfection for 48 h,
cells were harvested by 0.25% trypsin and centrifuged at 251 × g
for 5 min at room temperature. Subsequently, cells were washed
twice with cold PBS and resuspended in 250 µl binding buffer
(Nanjing KeyGen Biotech, Co., Ltd.). FITC Annexin V FITC (5 µl) and
1 µl PI solution were added and cells were incubated for 15 min at
room temperature. The apoptosis rate was detected using a Calibur
Flow Cytometer (BD Biosciences, San Jose, CA, USA). Three
independent experiments were repeated.
Luciferase report assay
TargetScan version 6.2 (http://www.targetscan.org) was used to investigate the
potential targets of miR-372 and then it was confirmed by western
blotting and a luciferase reporter assay. For the luciferase
reporter assay, cells were seeded into 24-well plates 24 h prior to
transfection. Cells were co-transfected with the E2F1 wild-type
(Wt) or mutant (Mut) 3′UTR reporter vector and miR-372 mimics or NC
using Lipofectamine 2000 according to the manufacturer's protocol.
After transfection for 48 h, cells were lysed and luciferase
activity against Renilla luciferase activity was measured
using a Dual Luciferase Reporter Assay System (Promega Corporation)
according to the manufacturer's protocol. Experiments were
independently repeated three times.
Western blot analysis
Proteins were lysed from treated cells using
radioimmunoprecipitation assay buffer containing protease inhibitor
(Sigma-Aldrich; Merck KGaA) and collected by centrifugation (16,000
× g at 4°C) for 15 min. The protein concentration was determined
using a bicinchoninic Acid Protein Assay kit (Sigma-Aldrich; Merck
KGaA). Total protein (40 µg) was separated via 10% SDS-PAGE,
transferred to a polyvinylidene difluoride membrane (GE Healthcare
Life Sciences, Chalfont, UK) and blocked with 5% non-fat milk for 1
h at room temperature. The membranes were probed with primary
antibodies against E2F1 (Abcam, Cambridge UK; cat. no. ab112580;
1:1,000) and β-actin (Santa Cruz Biotechnology, Inc., Dallas, TX,
USA; cat. no. sc-130300; 1:2,000) overnight at 4°C, and then
incubated with horseradish peroxidase conjugated secondary
antibodies (Santa Cruz Biotechnology, Inc.; cat. no. goat
anti-mouse sc-2005 and goat anti-rabbit sc-2004; 1:5,000) for 1 h
at room temperature. Finally, signals were detected using an
enhanced chemiluminescence system with the enhanced
chemiluminescense kit (EMD Millipore, Billerica, MA, USA). Each
reaction was performed in triplicate.
Statistical analysis
All data are presented as the mean ± standard
deviation and were analyzed using SPSS version 17.0 (SPSS Inc.,
Chicago, IL, USA) and GraphPad Prism software (version 5.0;
GraphPad Software, Inc., La Jolla, CA, USA). The difference between
groups was analyzed by Student's t test when comparing only two
groups and one-way analysis or post hoc test when comparing more
than two groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-372 is downregulated in breast
cancer specimens and cell lines
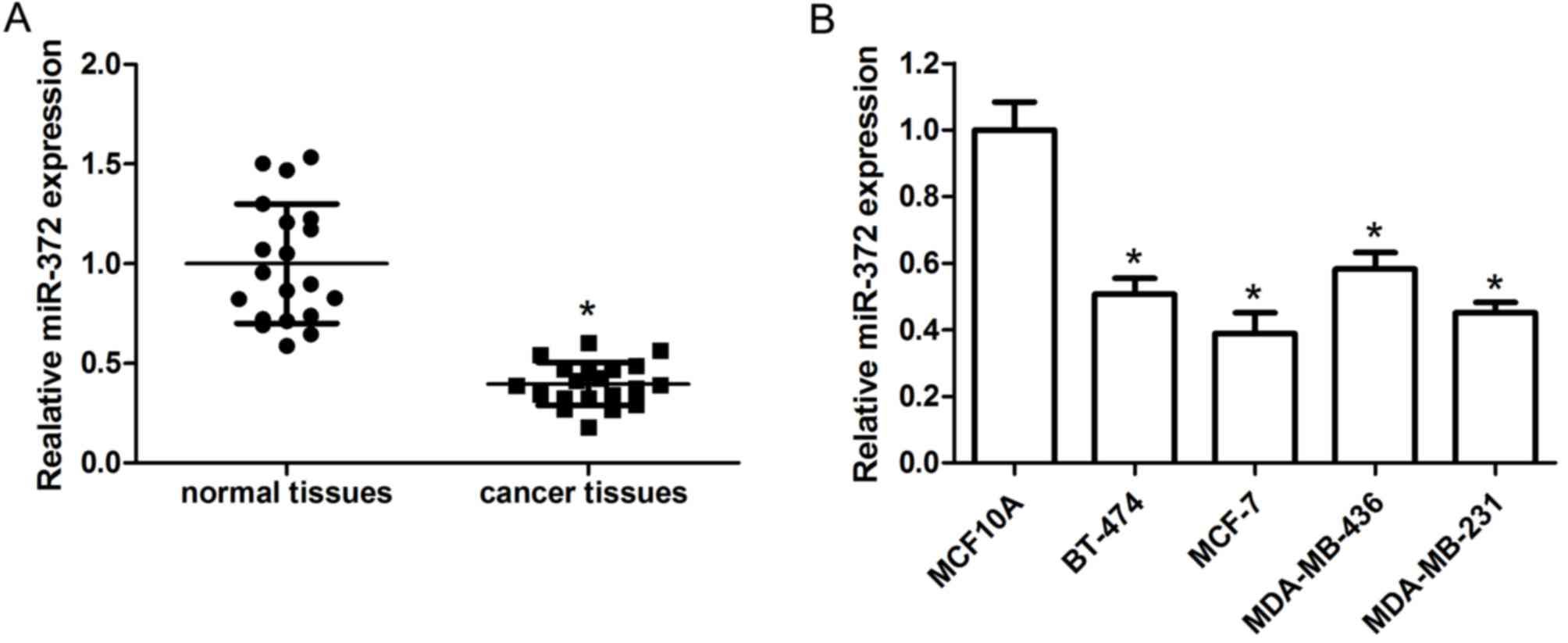
The expression level of miR-372 in human breast
cancer tissues and adjacent non-tumor tissues was detected by
RT-qPCR. The results demonstrated that miR-372 expression levels
were obviously reduced in breast cancer tissue specimens compared
with adjacent non-tumor tissues (Fig.
1A). The level of miR-372 in the BT-474, MCF-7, MDA-MB-436 and
MDA-MB-231 human breast cancer cell lines and the MCF10A normal
breast cell line was further examined. The data indicated that
miR-372 was downregulated in breast cancer cell lines compared with
MCF10A cells (Fig. 1B).
miR-372 inhibits cell proliferation
and induces apoptosis in MCF-7 cells
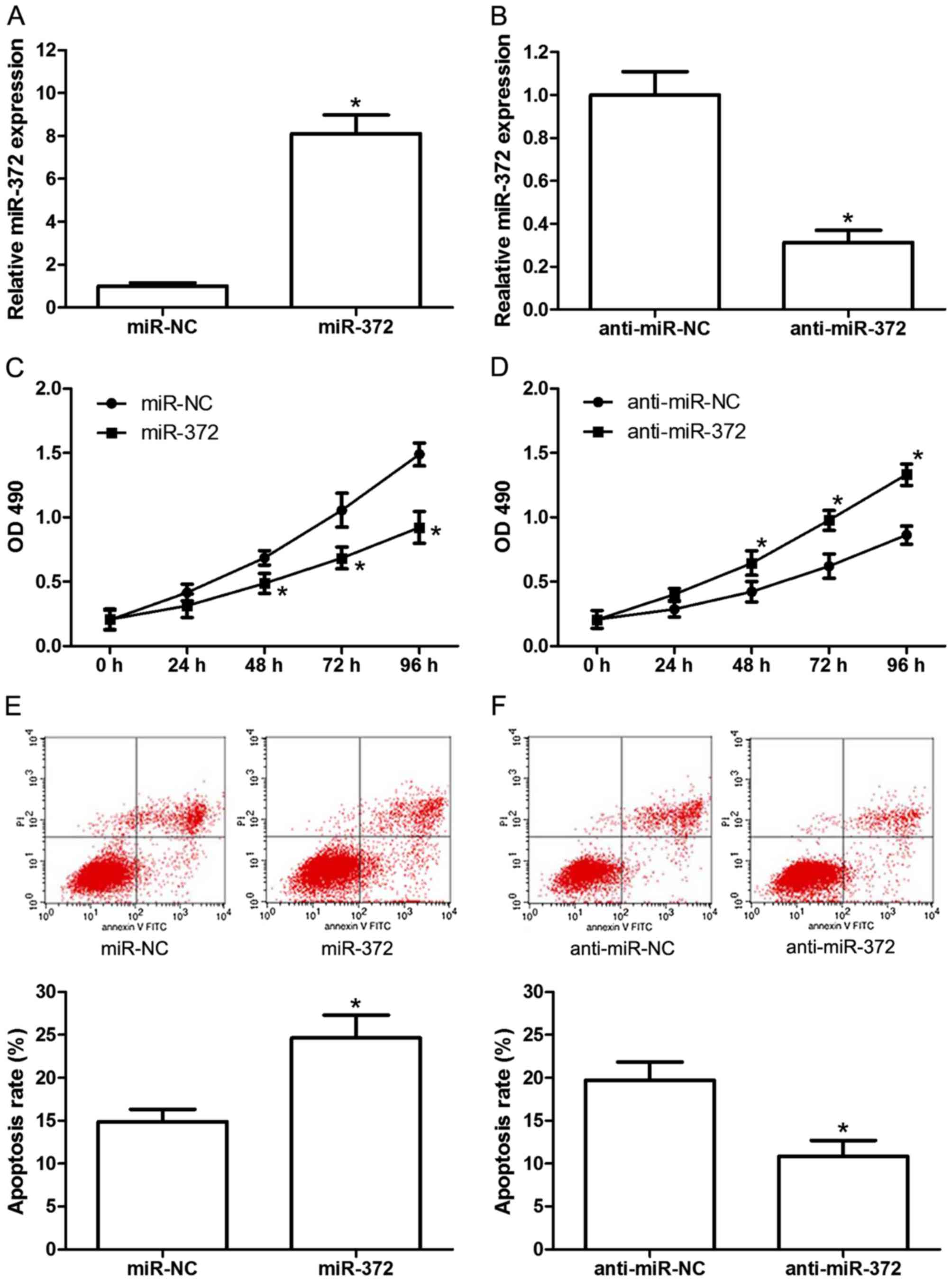
To investigate the effects of miR-372 on the
proliferation and apoptosis of breast cancer cells, miR-372 mimics
(miR-372), miR-372 inhibitors (anti-miR-372) or their negative
controls (miR-NC and anti-miR-NC) were transfected into the MCF-7
human breast cancer cell line. The transfection efficiency was
confirmed by RT-qPCR (Fig. 2A and
B). Subsequently, cell proliferation and apoptosis were
analyzed by MTT assay and flow cytometry, respectively. The results
demonstrated that miR-372 mimics significantly inhibited MCF-7 cell
proliferation (Fig. 2C), and
miR-372 inhibitors significantly increased proliferation (Fig. 2D), compared with the negative
control. Additionally, miR-372 mimics significantly induced
apoptosis (Fig. 2E) and miR-372
inhibitors markedly reduced apoptosis (Fig. 2F) compared with the control group.
These results revealed the tumor suppressive role of miR-372 in
human breast cancer development.
E2F1 is a direct target of miR-372 in
breast cancer
TargetScan 6.2 was used to investigate the
functional target of miR-372 in breast cancer, and E2F1 was
identified as a candidate target (Fig.
3A). A luciferase reporter assay and western blotting were
performed for confirmation. The luciferase reporter assay
demonstrated a significant downregulation of luciferase activity in
MCF-7 cells when co-transfected with miR-372 mimics (miR-372) and
Wt, but not the Mut 3′-UTR of E2F1 (Fig. 3B). The results of western blotting
indicated that protein expression levels of E2F1 in MCF-7 cells
decreased following transfection with miR-372 mimics (miR-372), but
increased when transfected with anti-miR-372 (Fig. 3C). Furthermore, the mRNA expression
levels of E2F1 in human breast cancer tissues and adjacent
non-tumor tissues were assessed. E2F1 was demonstrated to be
upregulated in human breast cancer tissues compared with adjacent
normal tissues (Fig. 3D), and its
level was negatively correlated with miR-372 levels in breast
cancer tissues (Fig. 3E). All the
data indicated that E2F1 is a direct target of miR-372 in breast
cancer.
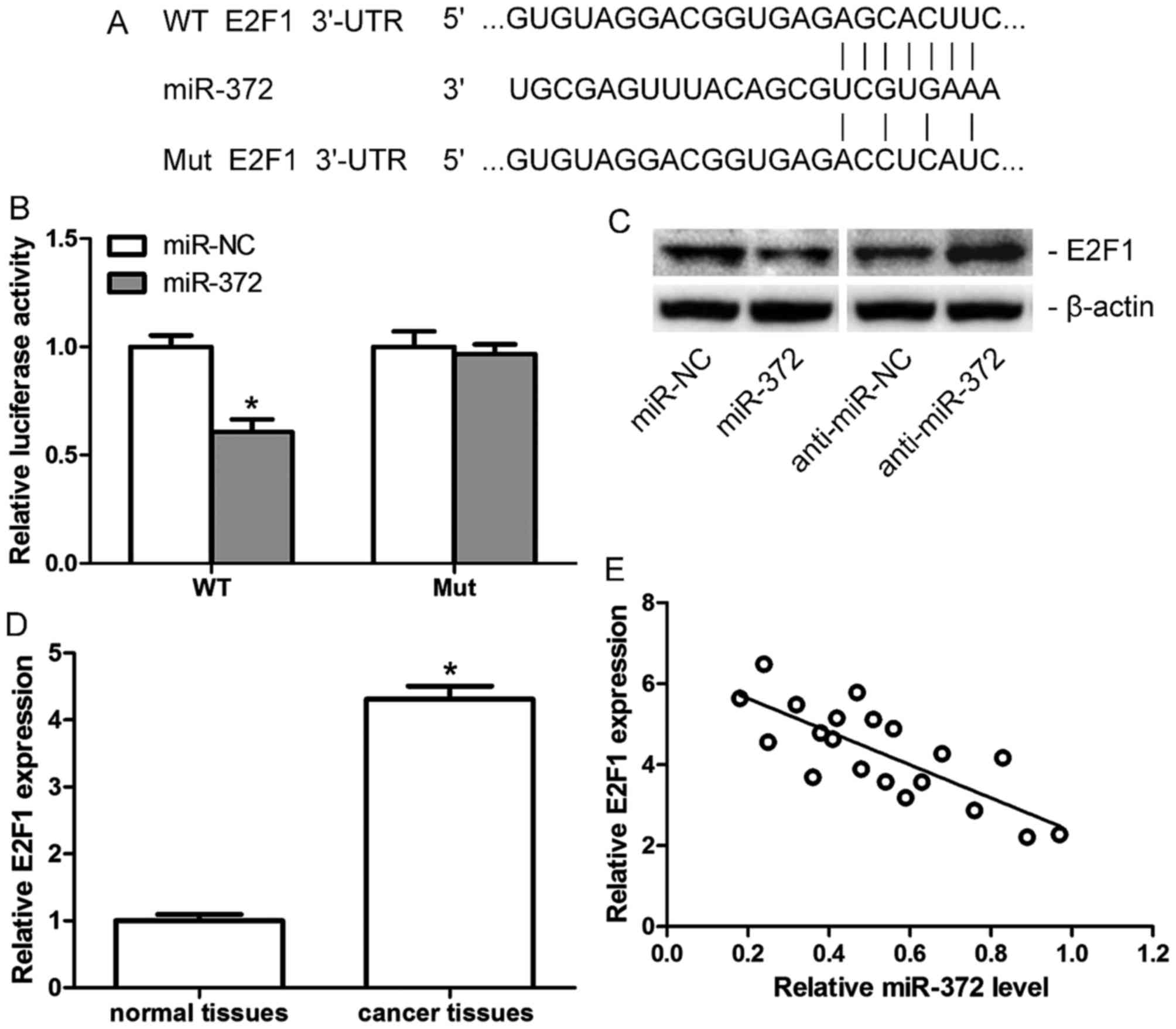 | Figure 3.E2F1 is a direct target of miR-372 in
breast cancer. (A) The sequences of miR-372 against WT or Mut
3′-UTR of E2F1, as predicted by Targetscan. (B) Relative luciferase
activity of MCF-7 cells co-transfected with miR-372 mimics or
miR-NC and WT or Mut 3′-UTR of E2F1. (C) E2F1 protein expression
levels in MCF-7 cells transfected with miR-372, anti-miR-372 or
miR-NC and anti-miR-NC, as assessed by western blotting. (D)
Relative mRNA expression levels of E2F1 in breast cancer tissues
and adjacent non-tumor tissues, as detected by reverse
transcription-quantitative polymerase chain reaction. (E)
Correlation between E2F1 expression and miR-372 level in breast
cancer tissues. Data are presented as the mean ± standard deviation
of three independent experiments. *P<0.05. miR-372,
microRNA-372; NC, negative control; UTR, untranslated region; WT,
wild-type; Mut, mutant; E2F1, transcription factor E2F1. |
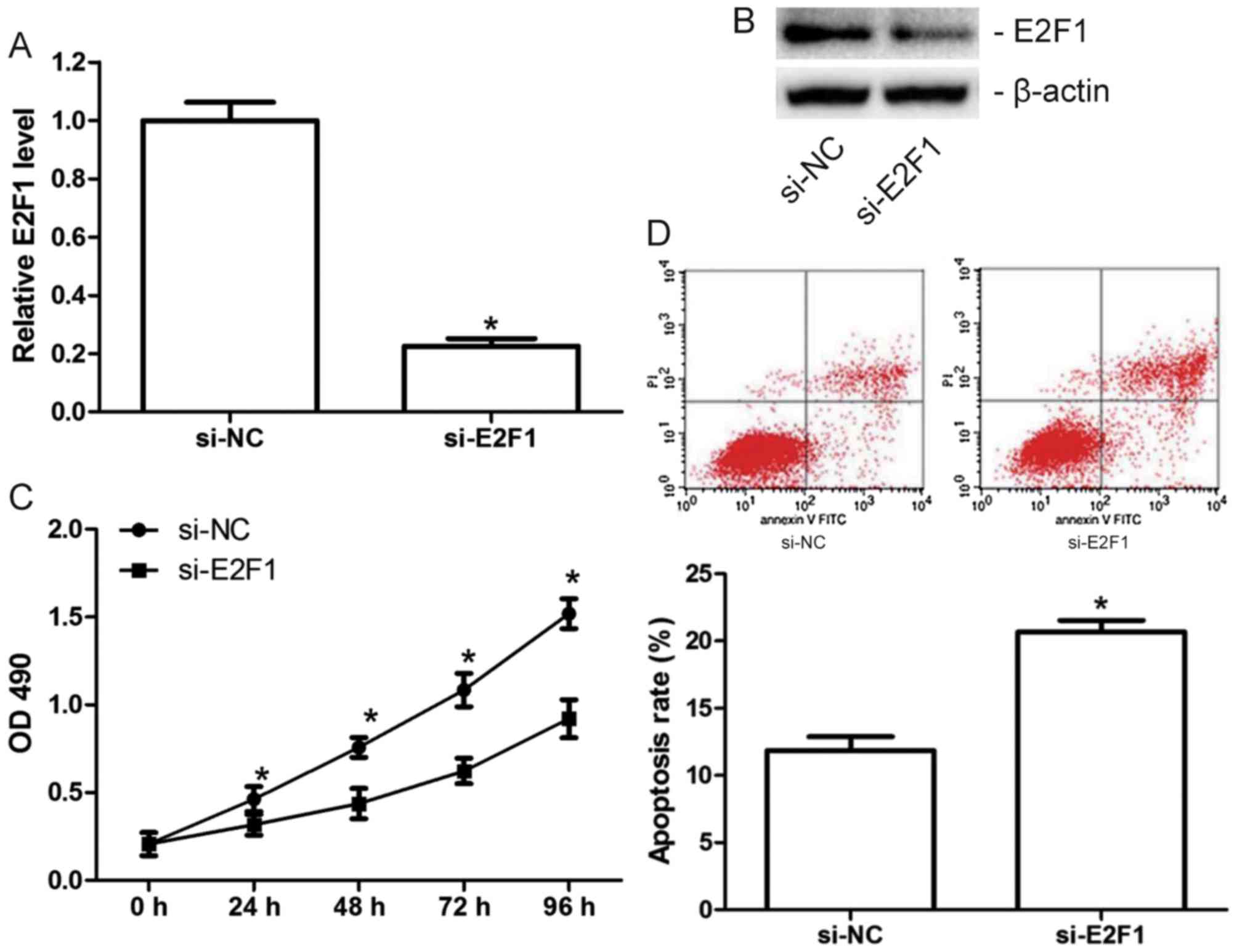
Knockdown of E2F1 inhibits
proliferation and induces apoptosis in MCF-7 cells
As miR-372 inhibits proliferation and induces
apoptosis in MCF-7 cells, and E2F1 is a direct target of miR-372,
MCF-7 cells were transfected with si-E2F1 or the scramble (si-NC)
to examine the effect of E2F1 on MCF-7 cell proliferation and
apoptosis. The transfection efficiency was confirmed by RT-qPCR
(Fig. 4A) and western blotting
(Fig. 4B). Results of functional
experiments demonstrated that knockdown of E2F1 significantly
inhibited proliferation (Fig. 4C)
and induced apoptosis in MCF-7 cells (Fig. 4D), which suggested that the tumor
suppressive role of miR-372 in breast cancer is involved in its
suppression of E2F1.
Discussion
Over the past few decades, the detection
non-malignant lesions of breast cancer has increased, including
atypical ductal hyperplasia, atypical lobular hyperplasia, ductal
carcinoma in situ and lobular carcinoma in situ
(20). Its early detection is due
to an increase in the identification of molecular biomarkers. Apart
from established biomarkers, including estrogen receptor,
progesterone receptor, receptor tyrosine-protein kinase erbB-2 and
Ki67, certain other novel molecular such as bcyclin E, B-Myb,
twist, and cyclin D-binding Myb-like Protein 1β have also been
identified as novel biomarkers for breast cancer early diagnosis
and prognosis prediction (21). A
recently published review indicated that miRNAs may serve important
roles in initiation, development, metastasis and drug resistance of
breast cancer (22). Furthermore,
miRNAs are hypothesized to have prognostic, diagnostic and
therapeutic potential in breast cancer (23). Thus, identifying novel miRNAs
involved in the initiation and development of breast cancer may be
helpful for breast cancer patients.
E2F is a family of transcription factors that
regulate the expression of genes essential for a wide range of
cellular functions, including cell cycle progression, DNA repair,
DNA replication, differentiation, proliferation and apoptosis
(24). The E2F family is composed
of at least eight members that can be divided into distinct
subgroups based on their structural and functional similarities
(25). E2F1, the most classic
member of the E2F family, exhibits dual properties in tumor
development by acting as a tumor suppressor or oncogene (26). Accumulating evidence has suggested
that E2F1 is regulated by various microRNA in tumor development.
For example, Yang et al (27) demonstrated that miR-106a suppresses
proliferation and induces apoptosis in human glioma cells, and
identified E2F1 as a direct functional target of miR-106a. Dar
et al (28) revealed that
miR-205 inhibits melanoma cell proliferation and induces apoptosis
via regulation of E2F1 protein level. Xiao et al (29) also reported that miR-329 could
markedly suppress cell proliferation and colony formation by
targeting E2F1 (29).
The present study demonstrated that miR-372 is
significantly down-regulated in human breast cancer tissues and
cell lines compared with healthy controls, which suggested that
miR-372 may serve as a tumor suppressor in breast cancer
development. Subsequetly, MCF-7 cells were transfected with miR-372
mimics or inhibitors to investigate its role in breast cancer. The
results indicated that miR-372 could inhibit proliferation and
induce apoptosis in breast cancer cells. Furthermore, E2F1 was
identified as a direct target of miR-372 in the MCF-7 breast cancer
cell line, and the mRNA level of E2F1 was negatively correlated
with miR-372 levels in breast cancer tissues. Additionally, it was
further demonstrated that knockdown of E2F1 inhibited proliferation
and induced apoptosis in MCF-7 cells.
In conclusion, to the best of our knowledge, the
present study investigated the expression level and roles of
miR-372 in breast cancer for the first time. miR-372 was
demonstrated to be significantly down-regulated in breast cancer
tissues and cell lines compared with normal controls. Furthermore,
the results indicated that miR-372 may inhibit proliferation and
induce apoptosis in breast cancer cell lines by directly targeting
E2F1. These findings suggested that miR-372 may have the potential
to serve as a novel molecule for the diagnosis and therapy of
patients with breast cancer.
Acknowledgements
The present study was supported by the Foundation of
Science and Technology Bureau of Wenzhou (grant nos. Y20100257 and
Y20160403), the Foundation of Traditional Chinese Medicine of
Zhejiang Province (grant no. 2009C520014) and the Foundation of
Public Welfare Technology Application of Zhejiang Province (grant
no. 2015C33100).
References
|
1
|
Cai Y, Yan X, Zhang G, Zhao W and Jiao S:
MicroRNA-205 increases the sensitivity of docetaxel in breast
cancer. Oncol Lett. 11:1105–1109. 2016.PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xiao X, Huang X, Ye F, Chen B, Song C, Wen
J, Zhang Z, Zheng G, Tang H and Xie X: The miR-34a-LDHA axis
regulates glucose metabolism and tumor growth in breast cancer. Sci
Rep. 6:217352016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cardoso F, Harbeck N, Fallowfield L,
Kyriakides S and Senkus E; ESMO Guidelines Working Group, : Locally
recurrent or metastatic breast cancer: ESMO clinical practice
guidelines for diagnosis, treatment and follow-up. Ann Oncol. 23
Suppl 7:vii11–19. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in Cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang Y and Blelloch R: Cell cycle
regulation by MicroRNAs in embryonic stem cells. Cancer Res.
69:4093–4096. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van Kouwenhove M, Kedde M and Agami R:
MicroRNA regulation by RNA-binding proteins and its implications
for cancer. Nat Rev Cancer. 11:644–656. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang T, Hu Y, Ju J, Hou L, Li Z, Xiao D,
Li Y, Yao J, Wang C, Zhang Y, et al: Downregulation of miR-522
suppresses proliferation and metastasis of non-small cell lung
cancer cells by directly targeting DENN/MADD domain containing 2D.
Sci Rep. 6:193462016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao Y, Lu G, Ke X, Lu X, Wang X, Li H,
Ren M and He S: miR-488 acts as a tumor suppressor gene in gastric
cancer. Tumour Biol. 37:8691–8698. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang L, Song G, Tan W, Qi M, Zhang L, Chan
J, Yu J, Han J and Han B: MiR-573 inhibits prostate cancer
metastasis by regulating epithelial-mesenchymal transition.
Oncotarget. 6:35978–35990. 2015.PubMed/NCBI
|
|
11
|
Song C, Liu LZ, Pei XQ, Liu X, Yang L, Ye
F and Xie X, Chen J, Tang H and Xie X: miR-200c inhibits breast
cancer proliferation by targeting KRAS. Oncotarget. 6:34968–34978.
2015.PubMed/NCBI
|
|
12
|
Wang L, Li L, Guo R, Li X, Lu Y, Guan X,
Gitau SC, Wang L, Xu C, Yang B and Shan H: miR-101 promotes breast
cancer cell apoptosis by targeting Janus kinase 2. Cell Physiol
Biochem. 34:413–422. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li G, Yao L, Zhang J, Li X, Dang S, Zeng
K, Zhou Y and Gao F: Tumor-suppressive microRNA-34a inhibits breast
cancer cell migration and invasion via targeting oncogenic TPD52.
Tumour Biol. 37:7481–7491. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Voorhoeve PM, le Sage C, Schrier M, Gillis
AJ, Stoop H, Nagel R, Liu YP, van Duijse J, Drost J, Griekspoor A,
et al: A genetic screen implicates miRNA-372 and miRNA-373 as
oncogenes in testicular germ cell tumors. Cell. 124:1169–1181.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cho WJ, Shin JM, Kim JS, Lee MR, Hong KS,
Lee JH, Koo KH, Park JW and Kim KS: miR-372 regulates cell cycle
and apoptosis of ags human gastric cancer cell line through direct
regulation of LATS2. Mol Cells. 28:521–527. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen X, Hao B, Han G, Liu Y, Dai D, Li Y,
Wu X, Zhou X, Yue Z, Wang L, et al: miR-372 regulates glioma cell
proliferation and invasion by directly targeting PHLPP2. J Cell
Biochem. 116:225–232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang X, Huang M, Kong L and Li Y: miR-372
suppresses tumour proliferation and invasion by targeting IGF2BP1
in renal cell carcinoma. Cell Prolif. 48:593–599. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu BL, Sun KX, Zong ZH, Chen S and Zhao
Y: MicroRNA-372 inhibits endometrial carcinoma development by
targeting the expression of the Ras homolog gene family member C
(RhoC). Oncotarget. 7:6649–6664. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fu SW, Lee W, Coffey C, Lean A, Wu X, Tan
X, Man YG and Brem RF: miRNAs as potential biomarkers in early
breast cancer detection following mammography. Cell Biosci.
6:62016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Inoue K and Fry EA: Novel molecular
markers for breast cancer. Biomark Cancer. 8:25–42. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Takahashi RU, Miyazaki H and Ochiya T: The
Roles of microRNAs in breast cancer. Cancers (Basel). 7:598–616.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mulrane L, Klinger R, McGee SF, Gallagher
WM and O'Connor DP: microRNAs: A new class of breast cancer
biomarkers. Expert Rev Mol Diagn. 14:347–363. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhan L, Zhang Y, Wang W, Song E, Fan Y and
Wei B: E2F1: A promising regulator in ovarian carcinoma. Tumour
Biol. 37:2823–2831. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
DeGregori J and Johnson DG: Distinct and
overlapping roles for E2F family members in transcription,
proliferation and apoptosis. Curr Mol Med. 6:739–748. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Knoll S, Emmrich S and Pützer BM: The
E2F1-miRNA cancer progression network. Adv Exp Med Biol.
774:135–147. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang G, Zhang R, Chen X, Mu Y, Ai J, Shi
C, Liu Y, Shi C, Sun L, Rainov NG, et al: MiR-106a inhibits glioma
cell growth by targeting E2F1 independent of p53 status. J Mol Med
(Berl). 89:1037–1050. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dar AA, Majid S, de Semir D, Nosrati M,
Bezrookove V and Kashani-Sabet M: miRNA-205 suppresses melanoma
cell proliferation and induces senescence via regulation of E2F1
protein. J Biol Chem. 286:16606–16614. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xiao B, Tan L, He B, Liu Z and Xu R:
MiRNA-329 targeting E2F1 inhibits cell proliferation in glioma
cells. J Transl Med. 11:1722013. View Article : Google Scholar : PubMed/NCBI
|