Introduction
Asthma is a common chronic inflammatory disorder
characterized by repeated attacks of wheezing, breathlessness,
cough and/or chest tightness (1).
It currently affects approximately 334 million people worldwide,
with China being one of the most afflicted countries, with
approximately 30 million asthmatics (2,3).
Following antigen exposure, CD4+T cells can
differentiate into at least four distinct subsets, including helper
T-cell (Th) 1, Th2, Th17 and regulatory T-cells (Treg) (4). The imbalance of Th1 and Th2 cells
contributes to the pathogenesis of asthma, constituting a target
for preventing and treating this disease (5,6).
However, in depth studies indicated that other immunological
mechanisms may be involved in regulating the formation of airway
inflammation in asthma, including Th17 and Treg cells (7). It has been suggested that reversal of
Th17/Treg cell imbalance maybe beneficial for suppressing chronic
Th2 cell-mediated inflammation in asthma (8–10).
Corticosteroids, β2-adrenoreceptor agonists and antileukotrienes or
leukotriene modifiers are currently used for asthma treatment;
whereas novel molecules including IL-5, IL-13, intercellular cell
adhesion molecule 1 and vascular cell adhesion molecule 1 have
yielded promising effects (11).
At present, the symptoms of a significant proportion of asthmatics
are poorly controlled, indicating the need for novel therapies.
Mesenchymal stem cells (MSCs) have gained attention
for their potential in treating and inhibiting further development
of pulmonary diseases, including bleomycin-induced lung fibrosis,
acute lung injury and lipopolysaccharide-induced acute lung injury
(12,13). For example, stem cells derived from
adipose tissue alleviate allergic airway inflammation and
ameliorate lung function by inducing Treg expansion (14). In an experimental model of severe
asthma, systemic transfer of bone marrow-derived MSCs (BM-MSCs)
resulted in decreased toluene diisocyanate-induced airway
inflammation and remodeling, as well as airway hyper-reactivity
(15). Compared with BM-MSCs,
human placenta MSCs (hPMSCs) are more easily obtained, propagated
and differentiated; there are therefore generally available and
suitable for large-scale culture in vitro (16,17).
Notably, hPMSCs retain immuno-tolerance properties, which are
closer to the clinical situation (18). hPMSCs suppress the activation and
proliferation of T lymphocytes (18,19).
Several studies have indicated the inhibitory
effects of MSCs on Th17 cell differentiation in vivo in
asthma models (20–23). However, the effects of hPMSCs on
Th17 and Treg cells in asthma remain unclear, with no data
regarding the immune responses of hPMSCs between the lymphatic
system and serum. Therefore, the aim of the present study was to
assess the therapeutic value of hPMSCs in asthma, evaluating the
impact of their transplantation on Th17/Treg balance in lymph and
serum samples from asthmatic animals.
Materials and methods
Animal model
A total of 60 male Sprague-Dawley rats (six-weeks
old) were purchased from Shandong Luye Pharmaceutical Co., Ltd.
(Yantai, China), and were bred in a specific pathogen-free animal
facility. The housing conditions were: Temperature was 18–26°C,
relative humidity was between 40–70%, 12-h during the day
(8:00-20:00) and 12-h by night (20:00-8:00) cycle mode, the noise
was below 85 decibels and the ammonia concentration was below 20
ppm and ventilated 8–12 times/h. The study protocol was approved by
the Institutional Animal Care and Use Committee of Binzhou Medical
University (Binzhou, China). The asthma rat model was established
as previously described (24).
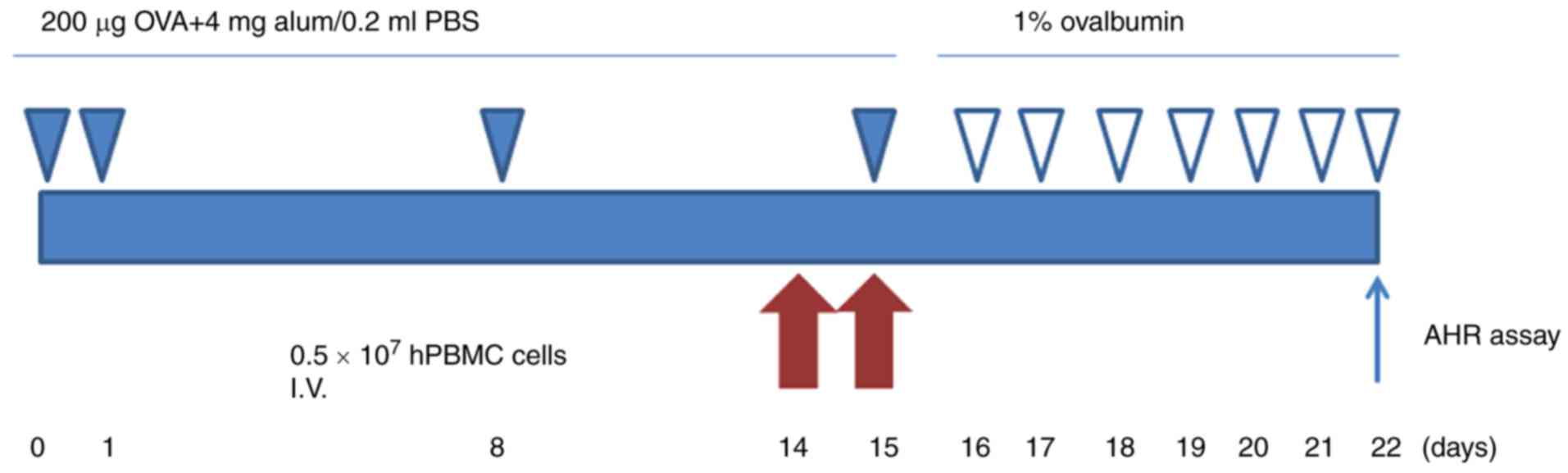
Rats were sensitized by hypodermic injection of 200 µg ovalbumin
(OVA; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) emulsified in
4 mg aluminum hydroxide in a total volume of 0.2 ml on day 0, 1, 8
and 15, respectively. Asthmatic rats were exposed to 1% OVA (grade
V; Sigma-Aldrich; Merck KGaA) in PBS for 30 min in a semi-closed
container on day 16, and every day afterwards for 1 week using an
ultrasonic nebulizer. On day 22, all rats were sacrificed (Fig. 1).
Isolation and identification of
hPMSCs
hPMSCs were harvested from the placental tissue of a
healthy pregnant mother following informed consent (obtained at the
Binzhou Medical University Hospital, Binzhou, China). The study
protocol was approved by the Ethics Committee of Binzhou Medical
University). In order to isolate hPMSCs, the placental tissue was
washed extensively with PBS, and digested with low glucose
(LG)-DMEM (GE Healthcare Life Sciences, Logan, UT, USA) containing
2.5 g/l trypsin (Sigma-Aldrich; Merck KGaA) and 1 g/l collagenase
IV (Sigma-Aldrich; Merck KGaA), at 37°C for 1 h. Digestion products
were centrifuged at 1,105 × g for 10 min. The pellet was filtered
through a nylon mesh to remove cellular debris, and incubated
overnight at 37°C in 5% CO2 in control medium. The
plates were then washed extensively with PBS to remove residual red
blood cells. A total of 1×107 cells/ml were seeded in
6-well plates, placed at 37°C in 5% CO2 cell culture
incubator. Cells were trypsinized to confluency, and used at the
third or fourth passage in experiments. Cell surface markers,
including CD34, CD45, CD73, CD90, CD105 and human leukocyte
antigen-antigen D related (HLA-DR) were assessed by flow cytometry
using specific kits from BD Biosciences (Franklin Lakes, NJ, USA),
according to the manufacturer's instructions.
All hPMSCs differentiation experiments were
conducted as previously reported (25). To induce neurogenic
differentiation, hPMSCs were incubated in LG-DMEM supplemented with
10% FBS, 1% penicillin-streptomycin, 20 ng/ml human basic
fibroblast growth factor, 10 ng/ml brain-derived neurotrophic
factor and 10 mM β-mercaptoethanol for 3 days, and stained for
Nestin, glial fibrillary acidic protein, β-tubulin,
microtubule-associated protein 2 and myelin basic protein.
Osteogenic differentiation was obtained after cell culture for 2
weeks or more in osteogenic medium (10% FBS, 0.1 mM dexamethasone,
10 µM E-glycerophosphate and 50 µg/ml ascorbic acid in DMEM) and
evaluation of extracellular matrix calcification by alizarin red
stain. Osteogenic differentiation was quantified by measuring
alizarin red stained areas in 6 wells by image analysis. For
adipogenic differentiation, the cells were cultured for 2 weeks in
adipogenic medium (10% FBS, 1 pM dexamethasone, 100 µg/ml
3-isobutyl-1-methylxanthine, 5 pg/ml insulin, and 60 pM
indomethacin in DMEM) and analyzed by Oil Red O staining. For
quantitation, 1 ml of isopropyl alcohol was added to the stained
culture dish.
Intravenous transplantation of
hPMSCs
hPMSCs were washed with PBS and resuspended in PBS
at a density of 1×107 cells/ml. Subsequently, 0.5 ml
cell suspension was injected into rats via the tail vein at 14 and
15 days. A total of 40 rats were divided into four groups, (n=10):
i) Control group (rats sensitized, pretreated and challenged with
PBS); ii) OVA group (rats sensitized and challenged with OVA and
pretreated with PBS); iii) control + hPMSCs group (rats sensitized
and challenged with PBS and pretreated with hPMSCs); iv) OVA +
hPMSCs group (rats sensitized with OVA, pretreated with hPMSCs,
then challenged with OVA).
Measurement of methacholine airway
hyperreactivity (AHR)
AHR was assessed as previously described (26). Briefly, unrestrained rats were
evaluated 24 h after the last challenge in the conscious state by
noninvasive whole-body plethysmography. The animals were placed in
the plethysmography chamber and exposed to increasing
concentrations of aerosolized methacholine (Chengdu
Chroma-Biotechnology Co., Ltd., Chengdu, China) at 0, 12.5, 5 and
50 mg/ml for 10 min. The enhanced pause (Penh) was calculated
automatically based on the mean pressure generated in the chamber
during inspiration and expiration combined with the time of each
phase. Penh values were obtained during each 3-min interval and
averaged.
Histopathological analysis
Lung tissues were fixed by infusing 4%
paraformaldehyde through the trachea for ~1 week. Then, the samples
were paraffin-embedded for light microscopic evaluation; 4 µm-thick
transverse sections obtained on a microtome were submitted to
hematoxylin and eosin. The degree of microscopic peri-bronchial and
peri-vascular inflammation was graded on a subjective scale of 0,
1, 2, 3 and 4 (9): Grade 0
indicated normal appearance, and grades 1, 2, 3 and 4 reflected
mild, moderate, distinct and severe inflammation, respectively.
Total lung inflammation and asthma scores were the sum of
peri-bronchial and peri-vascular inflammation scores; sections
obtained from the left lung were analyzed.
Bronchoalveolar lavage
Bronchoalveolar lavage fluid (BALF) samples were
collected by cannulation of the trachea. A syringe with normal
saline was inserted thrice along with the left lung bronchus for
tracheal lavage. The supernatant was discarded subseuquent to
centrifugation at 500 × g, 10 min, 25°C (Heraeus Biofuge; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), and cells were
submitted to Wright's staining. Total number of cells,
eosinophilia, lymphocyte, macrophage and neutrophils were obtained
by microscopic counting.
Lymph collection
Rats were anesthetized by intraperitoneal injection
of 4% chloral hydrate, and the abdominal cavity was exposed. The
superior mesenteric artery and lymph nodes around it were first
located, and a blunt dissection of the plasma membrane of lymphatic
vessels was conducted. Subsequently, a drainage tube was inserted
to extract 0.7–1 ml lymph. Following centrifugation at 500 × g, 10
min, 4°C for 10 min, the supernatant was collected and frozen at
−80°C until use.
Separation of mononuclear cells
(MNCs)
Blood was diluted with an equal volume of the PBS,
overlayed on Ficoll, and submitted to gradient centrifugation at
900 × g, 20 min, 4°C for 20 min. The MNCs were collected at the
interphase, and adjusted at 2–3×106/ml with fresh
RPMI-1640. The MNCs were seeded per well of 24-well plates, with
concanavalin A (ConA) added at 5 g/ml, incubated at 37°C in the
presence of 5% CO2 for 18 h.
Flow cytometric analysis of Treg and
Th17 cellsin lymph and MNC samples
Following 18-h culture, MNCs were collected. For
Treg detection, the cells were stained with fluorescent-labeled
anti-CD25-phycoerythrin (PE), anti-CD4-fluorescein isothiocyanate
(FITC) and anti-forkhead box P3 (Foxp3)-PerCP/Cy5 (0.2 mg/ml)
antibodies (1:1; cat. nos. 12-0390-82, 11-0040-82, 45-5773-82,
respectively; eBioscience; Thermo Fisher Scientific, Inc.). PE-Rat
IgG1 κ, FITC- Rat IgG2a κ and PE-Cy5 Rat IgG2a Isotype control
antibodies (1:1; cat. nos. 12-4301-82, 11-4321-80 and 35-4321-82,
respectively; eBioscience; Thermo Fisher Scientific, Inc.) were
used as controls. For Th17 detection, the cells were treated with
vancomycin (1 µg/ml), and Brefeldin (10 µg/ml; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) in the last 5 h of incubation, and
stained with fluorescent-labeled anti-CD4-FITC and anti-IL17-PE
(1:1; cat. nos. 11-0040-82 and 12-7177-81, respectively;
eBioscience; Thermo Fisher Scientific, Inc.) antibodies. FITC-rat
IgG2a κ isotype and PE-rat IgG2a κ isotype antibodies (1:1; cat.
no. s 11-4321-80 and 12-4321-41, respectively; eBioscience; Thermo
Fisher Scientific, Inc.) were used as controls.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from lung tissues and MNCs
using RNAiso Plus (Takara Bio, Inc., Otsu, Japan), and 2 µg was
reverse transcribed using the Revert Aid First Strand cDNA
Synthesis kit (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
RT-qPCR was conducted using 0.4 µl of the forwards and reverse
primers, each (synthesized by Sangon Biotech Co., Ltd., Shanghai,
China), 2.0 µl cDNA, and 10 µl SYBR green-based PCR master mix
(Takara Bio, Inc.) on a Rotor Gene 3000 Real-Time PCR System from
Corbett Research (Sydney, Australia). Reactions were performed at
95°C for 30 sec (initial denaturation), followed by 40 cycles of
95°C for 5 sec and 60°C for 20 sec. Fluorescence readings were
monitored during the 60°C step. Data were analyzed by the
2−ΔΔCq method using the LightCycler Data Analysis
software (version 4.0.5.415; Roche Diagnostics, Basel, Switzerland)
(27). The following primers were
used: Foxp3 forward, 5′-ttcacctatgccaccctcat-3′ and reverse,
5′-actgctcccttctcactctcc-3′; GAPDH forward,
5′-acagcaacagggtggtggac-3′ and reverse, 5′-tttgagggtacagcgaactt-3′;
IL-10 forward, 5′-gctatgttgcctgctcttactg-3′ and reverse,
5′-tctggctgactgggaagtg-3′; RORγt forward,
5′-gacttttcccacttcctacagc-3′ and reverse,
5′-cagatgctccactctcctcttt-3′; IL-17 forward,
5′-gaaagtcctcaactcccttagctc-3′ and reverse,
5′-cctcccagatcacagaaggata-3′.
Western blotting assay
Western blot analysis was performed to detect the
protein expression levels of RORγt and Foxp3 in asthma models and
hPMSC-treated groups. Lung tissues were lysed using RIPA lysis
buffer (Beyotime Institute of Biotechnology, Haimen, China). A
total of ~50 mg amounts of protein were separated by 10% SDS-PAGE
and transferred onto PVDF membranes. After blocking with 5% skimmed
milk, the membranes were probed with a rabbit antibodies against
rat RORγt (1:500; ab78007; Abcam, Cambridge, UK) and mouse
antibodies for Foxp3 (1:1,000; ab22510; Abcam) followed by
horseradish peroxidase-conjugated goat anti-rabbit IgG (1:1,000;
cat. no. A0208; Beyotime Institute of Biotechnology, China) IgG
antibody incubation. Protein bands were detected with an enhanced
chemiluminescence kit (Pierce, Biotechnology, Inc., Rockford, IL,
USA) according to the manufacturer's instructions. GAPDH was used
as a loading control.
Measurement of cytokines
Lung tissues, lymph supernatants and serum specimens
from the four groups were collected and analyzed using commercially
available Quantikine kits, including rat IL-10 Immunoassay ELISA
kit, R1000 IL-17 and IL-10 mouse IL-17 Immunoassay ELISA kit, M1700
(both R&D Systems, Inc., Minneapolis, MN, USA).
Statistical analysis
Data are mean ± standard deviation. Multigroup
comparsions of the means were carried out by one-way analysis of
variance test with post hoc contrasts by Student-Newman-Keuls test.
Statistical analyses were performed using the SPSS 18.0 software
(SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
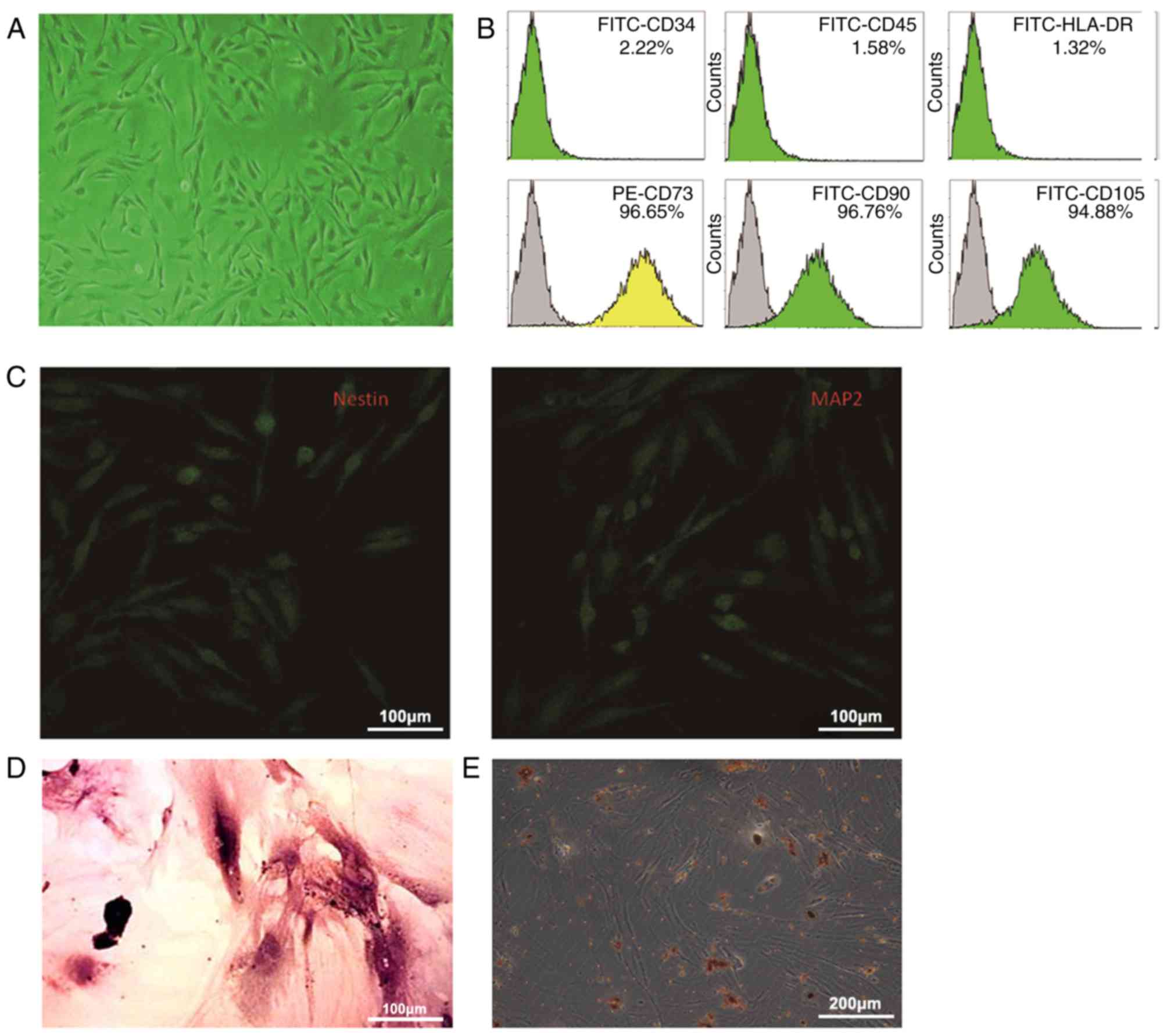
hPMSCs exhibit multipotency
The hPMSCs obtained had a spindle-shaped
fibroblast-like appearance (Fig.
2A). Flow cytometry indicated that they expressed the
cell-surface markers CD73, CD90 and CD105, however not HLA-DR, CD34
and CD45 (Fig. 2B). In addition,
hPMSCs could be differentiated into neuronal cells, osteoblasts and
fat cells (Fig. 2C-E).
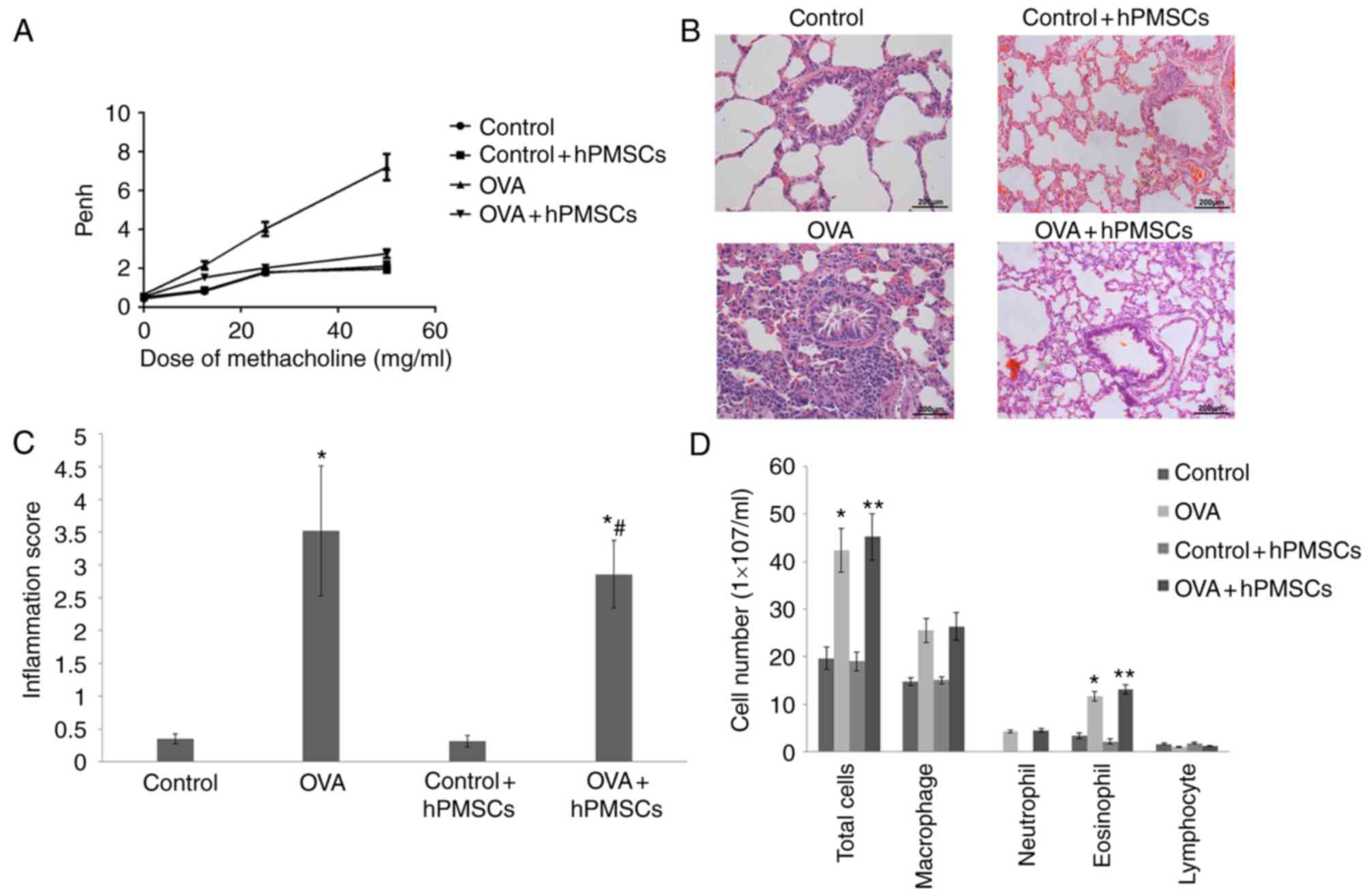
Methacholine AHR
In order to assess the effect of hPMSCs on lung
function, AHR was measured. As presented in Fig. 3A, Penh values in the four groups
increased with the methacholine concentrations. However, Penh
values in the OVA group at 25–50 mg/ml methacholine were
significantly higher than those in the control, control + hPMSCs
and OVA + hPMSCs groups. These results indicated that hPMSCs
treatment significantly alleviated AHR in response to methacholine
in asthmatic rats (P<0.01).
hPMSCs engraftment results in
significantly fewer inflammatory cells in BALF
The numbers of total cells, macrophages, neutrophils
and eosinophils were markedly increased in the OVA group compared
with those of control + hPMSCs and control groups (Fig. 3). Meanwhile, the hPMSCs + OVA group
had more total cells and eosinophils compared with the OVA group.
Lymphocyte numbers were similar among all four groups. Then,
inflammation scores were evaluated in each treatment group, taking
into consideration the proportion of alveolar collapse, broncho
constriction index, inflammatory cell infiltration and mucosal
membrane thickening. The resulting values were higher in the OVA
group compared with those of the control and control + hPMSCs
groups. Notably, the OVA + hPMSCs group exhibited a significantly
reduced score compared with the OVA group value (Fig. 3C). These results indicated that
hPMSCs administration alleviated inflammation in OVA-treated
rats.
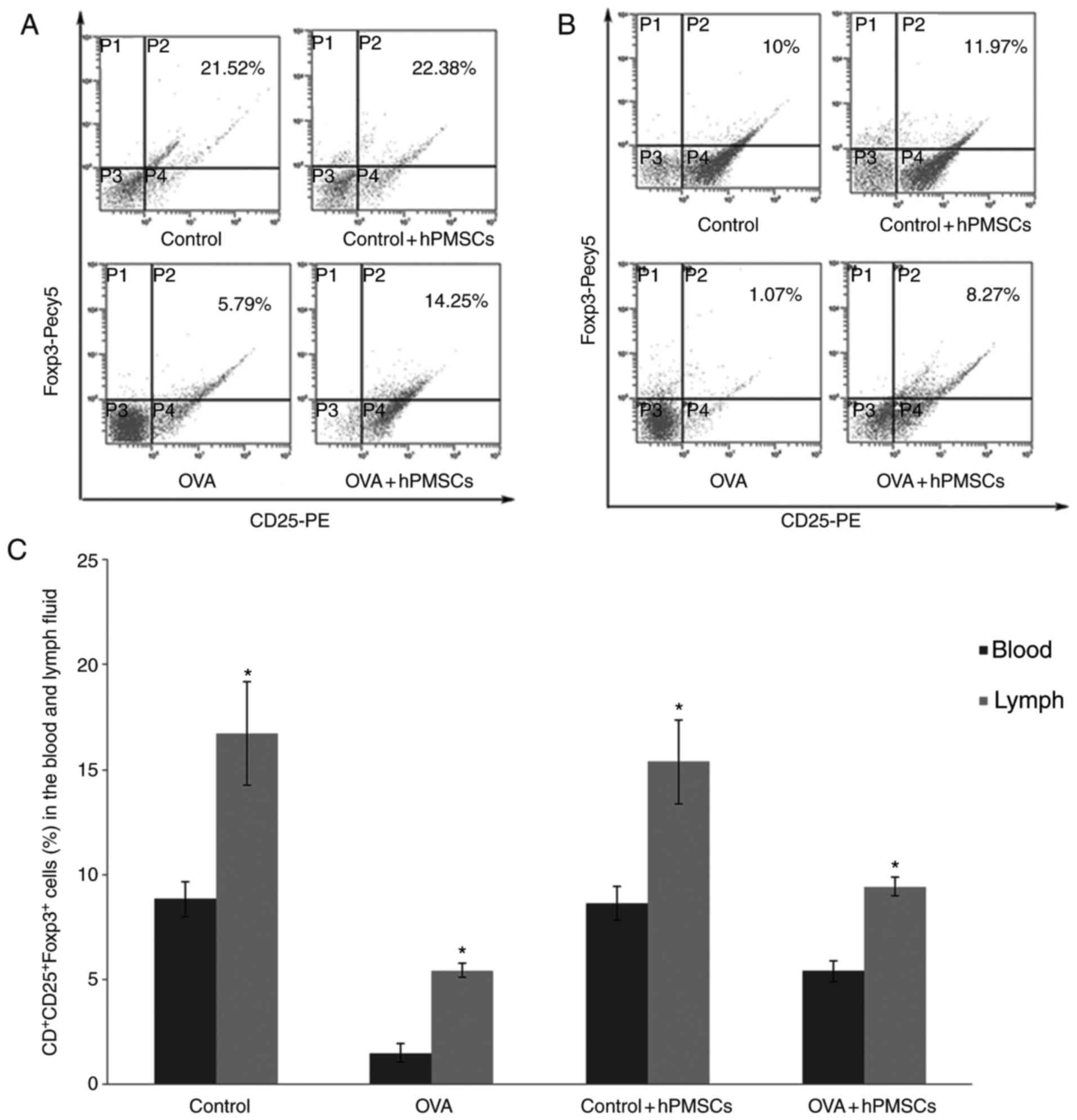
hPMSCs engraftment alters the
frequencies of Treg andTh17 cells in serum and lymph
To investigate the mechanism of hPMSCs effects,
CD4+CD25+regulatory T cells and Th17 cells
were quantified. The amount of
CD4+CD25+Foxp3+ (Treg cells) in
the lymph were significantly lower in the OVA group (P<0.05)
compared with control values. However, hPMSC transplantation
significantly increased the lymph ratio of
CD4+CD25+ Treg cells in OVA-treated animals
(OVA+ hPMSCs group; P<0.05) (Fig.
4A and C). A similar trend was obtained for serum samples, with
the OVA group exhibiting significantly lower amounts of Treg cells
than the control and hPMSCs treatment groups (P<0.05);
administration of hPMSCs markedly increased
CD4+CD25+Foxp3+ (Treg cells)
proportions in serum from OVA treated animals (Fig. 4B and C).
The percentages of Th17 positive cells in serum and
lymph from OVA treated animals were higher compared with control
values (P<0.05; Fig. 4D-F), and
were decreased following hPMSCs administration (P<0.05). A
greater number of Th17 cells were observed in the lymph compared
with the serum in all groups (P<0.05; Fig. 4D-F).
Effect of hPMSC administration on
levels of cytokines andtranscription factors in asthmatic rats
To further assess the effects of hPMSC
administration in asthmatic rats, cytokine levels were measured in
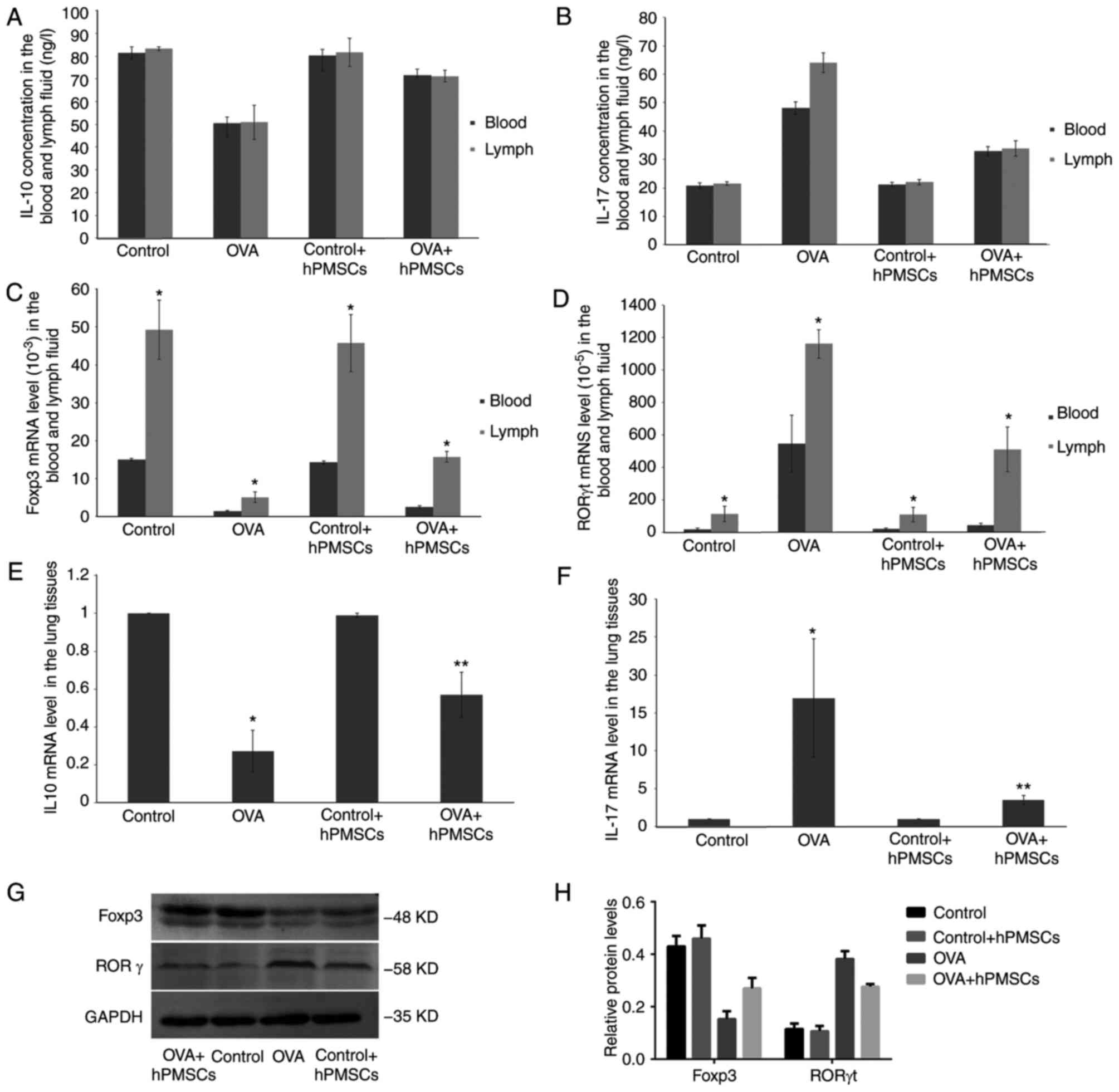
serum and lymph samples. As presented in Fig. 5A, lower IL-10 amounts detected by
ELISA were obtained in the OVA group compared with control values,
both in serum and lymph specimens. In addition, IL-17 exhibited the
opposite trend (Fig. 5B). Notably,
administration of hPMSCs in OVA-treated rats (OVA + hPMSCs group)
resulted in increased IL-10 and reduced IL-17 levels, both in serum
and lymph samples, compared with the OVA group (P<0.05).
However, no significant differences in IL-10 and IL-17 levels
between lymph and peripheral blood samples were observed in either
group (Fig. 5A and B).
To further characterize the effects of hPMSCs on
immune response, gene expression levels of select molecules were
assessed by RT-qPCR. Notably, mRNA levels of the Treg master
transcription factor Foxp3 were significantly increased in
hPMSCs-treated asthmatic rats, both in blood and lymph samples
(Fig. 5C). An opposite trend was
obtained for the Th17 transcription factor RORγt, in lymph and
peripheral blood samples (Fig.
5D). Foxp3 and RORγt mRNA levels in lymph samples were
significantly increased compared with the levels in the serum in
each group (P<0.05).
Subsequently, IL-10 and IL-17 gene expression levels
were assessed in lung tissue samples by RT-qPCR. In agreement with
the above, IL-10 mRNA levels were significantly decreased in the
OVA group compared with control rat values (P<0.05; Fig. 5E), while IL-17 levels were
increased. However, IL-10 upregulation and IL-17 downregulation
were reversed by hPMSCs treatment (OVA + hPMSCs groups) (P<0.05;
Fig. 5F). Finally, tissue protein
levels of Foxp3 and RORγt were assessed by western blotting. As
presented in Fig. 5G and H, Foxp3
protein levels were decreased, and RORγt amounts increased in the
OVA group compared with control values; meanwhile, transplantation
of hPMSCs resulted in increased Foxp3 and RORγt protein amounts
compared with the asthma (OVA) group (Fig. 5G and H).
Discussion
The present study indicated that treatment with
hPMSCs improved lung inflammation and corrected the Th17/Treg
balance, which may be mediated IL-17/IL-10 and Foxp3/RORγt ratios
between peripheral blood and mesenteric lymph nodes.
A previous study demonstrated that MSCs have an
important therapeutic potential in several clinical disorders such
as asthma (28). It is known that
hPMSCs generated from the placenta are morphologically and
functionally similar to BM-MSCs; however, the immune inhibitory
effects of hPMSCs on T cell proliferation are more prominent than
those of BM-MSCs (29,30). In the present study, it was
demonstrated that the placenta had the advantages of producing more
MSCs compared with bone marrow, with reduced ethical issues (as the
placenta is medical waste). These results suggest that hPMSCs maybe
an alternative source of stem cells for medicinal use.
Th17 cells that produce IL-17 participate in the
development of neutrophilic inflammation in asthma via expression
of ROR; IL-17 expression is upregulated in the airways of asthmatic
patients, correlating with eosinophilic airway inflammation
(31). Previous studies
demonstrated that allergen sensitization induces Th17-dependent
airway neutrophilia and AHR (32),
and the degree of AHR in patients with asthma is positively
correlated with sputum IL-17 levels (33,34).
Treg development and function are dependent on CD4
and CD25 receptors and the expression of the master transcription
factor Foxp3; Treg not only inhibits Th1 and Th2 cell immune
responses (35), however
additionally the proliferation and function of Th17 cells by
contact-dependent suppression or release of the anti-inflammatory
cytokines IL-10 and TGF-β. Previous studies have demonstrated that
BM-MSCs inhibit Th2-mediated allergic airway inflammation by
altering antigen-specific CD4 T lymphocyte differentiation
(36,37). As indicated above, hPMSC
administration increased IL-10 production and decreased IL-17
levels to inhibit Th17 cells in lymph and blood, correcting the
Th17/Treg imbalance in asthmatic rats. Th17/Treg associated with
inflammation in asthma maybe mediated by Foxp3/RORγt alteration.
The IL-17/IL-10 balance in the lung, lymph and circulation
indicates that the complex regulatory role of hPMSCs requires
further investigation.
Treg expressing Foxp3 serves an anti-inflammatory
role and maintain immune tolerance to self-components through
cell-to-cell direct contact or the release of cytokines such as
IL-10. Foxp3, as a master regulator, is involved in modulating Treg
mediated immunosuppression by inducing the production of TGF-β and
IL-10 (38,39). RORγt is a Th17 cell-specific
transcription factor. RORγt knockout mice exhibit reduced amounts
of Th17 cells and decreased incidence of autoimmune diseases
(40). Notably, Th17 cells and
IL-17 were demonstrated to induce airway remodeling (41). Studies have indicated that Th17
cell cytokine and protein expression levels are significantly
higher in the lung tissue, sputum and BALF from patients with
asthma (31). The present study
demonstrated that Foxp3 levels in lymph and serum samples increased
in asthmatic rats treated with hPMSCs treatment, while RORγt levels
decreased. These results indicated that hPMSCs may have ameliorated
inflammation in asthma by boosting Foxp3 and IL-10 production to
increase Treg cell amounts, while repressing RORγt and IL-17 to
reduce Th17 cell number. This ultimately resulted in corrected
Th17/Treg balance in asthma.
In summary, it was demonstrated for, to the best of
our knowledge, the first time, that hPMSCs alleviated allergic
airway inflammation in a rat model of asthma. It appeared that
Th17/Treg rebalance was induced by hPMSCs administration. This
protection may be mediated by Treg regulation, partly involving
increased IL-10 levels, in addition to the ratio of associated
transcription factors including Foxp3/RORγt. These results indicate
that hPMSCs affects immunosuppression, and may represent an
improved cell source to replace BM-MSCs for immune disease
treatment. The mechanisms of hPMSCs on the immune association
between lymph and serum require further investigation.
Acknowledgements
The present study was supported by grants from the
Shandong Province Natural Science Foundation of China (grant nos.
ZR2010HM085 and ZR2011HM081), the National Natural Science
Foundation of China (grant no. 81273200) and the Taishan Scholar
Foundation.
Glossary
Abbreviations
Abbreviations:
|
MSCs
|
mesenchymal stem cells
|
|
Treg
|
regulatory T cell
|
|
hPMSCs
|
human placenta MSCs
|
|
Th
|
helper T-cell
|
|
BM-MSCs
|
bone marrow-derived mesenchymal stem
cells
|
|
AHR
|
airway hyperreactivity
|
|
BALF
|
bronchoalveolar lavage fluid
|
|
MNCs
|
mononuclear cells
|
|
ConA
|
concanavalin A
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
References
|
1
|
Zhang J, Dai J, Yan L, Fu W, Yi J, Chen Y,
Liu C, Xu D and Wang Q: Air pollutants, climate, and the prevalence
of pediatric asthma in urban areas of China. Biomed Res Int.
2016:29351632016.PubMed/NCBI
|
|
2
|
Asher I and Pearce N: Global burden of
asthma among children. Int J Tuberc Lung Dis. 18:1269–1278. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen ZH, Wang PL and Shen HH: Asthma
research in China: A five-year review. Respirology. 18 Suppl
3:S10–S19. 2013. View Article : Google Scholar
|
|
4
|
Maddur MS, Miossec P, Kaveri SV and Bayry
J: Th17 cells: Biology, pathogenesis of autoimmune and inflammatory
diseases, and therapeutic strategies. Am J Pathol. 181:8–18. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lloyd CM and Hawrylowicz CM: Regulatory T
cells in asthma. Immunity. 31:438–449. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jiang H, Wu X, Zhu H, Xie Y, Tang S and
Jiang Y: FOXP3(+)Treg/Th17 cell imbalance in lung tissues of mice
with asthma. Int J Clin Exp Med. 8:4158–4163. 2015.PubMed/NCBI
|
|
7
|
Weaver CT, Hatton RD, Mangan PR and
Harrington LE: IL-17 family cytokines and the expanding diversity
of effector T cell lineages. Annu Rev Immunol. 25:821–852. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Souwer Y, Szegedi K, Kapsenberg ML and de
Jong EC: IL-17 and IL-22 in atopic allergic disease. Curr Opin
Immunol. 22:821–826. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hou X, Wan H, Ai X, Shi Y, Ni Y, Tang W
and Shi G: Histone deacetylase inhibitor regulates the balance of
Th17/Treg in allergic asthma. Clin Respir J. 10:371–379. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wei Y, Liu B, Sun J, Lv Y, Luo Q, Liu F
and Dong J: Regulation of Th17/Treg function contributes to the
attenuation of chronic airway inflammation by icariin in
ovalbumin-induced murine asthma model. Immunobiology. 220:789–797.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kanagaratham C and Radzioch D: Allergic
Asthma: A summary from genetic basis, mouse studies, to diagnosis
and treatment. Curr Pharm Des. 22:6261–6272. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Moodley Y, Manuelpillai U and Weiss DJ:
Cellular therapies for lung disease: A distant horizon.
Respirology. 16:223–237. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sinclair K, Yerkovich ST and Chambers DC:
Mesenchymal stem cells and the lung. Respirology. 18:397–411. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cho KS, Park MK, Kang SA, Park HY, Hong
SL, Park HK, Yu HS and Roh HJ: Adipose-derived stem cells
ameliorate allergic airway inflammation by inducing regulatory T
cells in a mouse model of asthma. Mediators Inflamm.
2014:4364762014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee SH, Jang AS, Kwon JH, Park SK, Won JH
and Park CS: Mesenchymal stem cell transfer suppresses airway
remodeling in a toluene diisocyanate-induced murine asthma model.
Allergy Asthma Immunol Res. 3:205–211. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Vellasamy S, Sandrasaigaran P, Vidyadaran
S, George E and Ramasamy R: Isolation and characterisation of
mesenchymal stem cells derived from human placenta tissue. World J
Stem Cells. 4:53–61. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Abumaree MH, Al Jumah MA, Kalionis B,
Jawdat D, Al Khaldi A, AlTalabani AA and Knawy BA: Phenotypic and
functional characterization of mesenchymal stem cells from
chorionic villi of human term placenta. Stem Cell Rev. 9:16–31.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Luan X, Li G, Wang G, Wang F and Lin Y:
Human placenta-derived mesenchymal stem cells suppress T cell
proliferation and support the culture expansion of cord blood
CD34+ cells: A comparison with human bone marrow-derived
mesenchymal stem cells. Tissue Cell. 45:32–38. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu KJ, Wang CJ, Chang CJ, Hu HI, Hsu PJ,
Wu YC, Bai CH, Sytwu HK and Yen BL: Surface expression of HLA-G is
involved in mediating immunomodulatory effects of placenta-derived
multipotent cells (PDMCs) towards natural killer lymphocytes. Cell
Transplant. 20:1721–1730. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang M, Zhao X, Liu Y, Tian Y, Ran X and
Jiang Y: A role for WNT1-inducible signaling protein-1 in airway
remodeling in a rat asthma model. Int Immunopharmacol. 17:350–357.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Trzil JE, Masseau I, Webb TL, Chang CH,
Dodam JR, Cohn LA, Liu H, Quimby JM, Dow SW and Reinero CR:
Long-term evaluation of mesenchymal stem cell therapy in a feline
model of chronic allergic asthma. Clin Exp Allergy. 44:1546–1557.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ogulur I, Gurhan G, Aksoy A, Duruksu G,
Inci C, Filinte D, Kombak FE, Karaoz E and Akkoc T: Suppressive
effect of compact bone-derived mesenchymal stem cells on chronic
airway remodeling in murine model of asthma. Int Immunopharmacol.
20:101–109. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zeng SL, Wang LH, Li P, Wang W and Yang J:
Messenchymal stem cells abrogate experimental asthma by altering
dendritic cell fuction. Mol Med Rep. 12:2511–2520. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yen BL, Huang HI, Chien CC, Jui HY, Ko BS,
Yao M, Shun CT, Yen ML, Lee MC and Chen YC: Isolation of
multipotent cells from human term placenta. Stem Cells. 23:3–9.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee RH, Kim B, Choi I, Kim H, Choi HS, Suh
K, Bae YC and Jung JS: Characterization and expression analysis of
mesenchymal stem cells from human bone marrow and adipose tissue.
Cell Physiol Biochem. 14:311–324. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee CG, Link H, Baluk P, Homer RJ,
Chapoval S, Bhandari V, Kang MJ, Cohn L, Kim YK, McDonald DM and
Elias JA: Vascular endothelial growth factor (VEGF) induces
remodeling and enhances TH2-mediated sensitization and inflammation
in the lung. Nat Med. 10:1095–1103. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mariñas-Pardo L, Mirones I, Amor-Carro O,
Fraga-Iriso R, Lema-Costa B, Cubillo I, Rodríguez Milla MÁ,
García-Castro J and Ramos-Barbón D: Mesenchymal stem cells regulate
airway contractile tissue remodeling in murine experimental asthma.
Allergy. 69:730–740. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Srour N and Thébaud B: Stem cells in
animal asthma models: A systematic review. Cytotherapy.
16:1629–1642. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li X, Bai J, Ji X, Li R, Xuan Y and Wang
Y: Comprehensive characterization of four different populations of
human mesenchymal stem cells as regards their immune properties,
proliferation and differentiation. Int J Mol Med. 34:695–704. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Doe C, Bafadhel M, Siddiqui S, Desai D,
Mistry V, Rugman P, McCormick M, Woods J, May R, Sleeman MA, et al:
Expression of the T helper 17-associated cytokines IL-17A and
IL-17F in asthma and COPD. Chest. 138:1140–1147. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wilson RH, Whitehead GS, Nakano H, Free
ME, Kolls JK and Cook DN: Allergic sensitization through the airway
primes Th17-dependent neutrophilia and airway hyperresponsiveness.
Am J Respir Crit Care Med. 180:720–730. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Barczyk A, Pierzchala W and Sozanska E:
Interleukin-17 in sputum correlates with airway hyperresponsiveness
to methacholine. Respir Med. 97:726–733. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lathrop MJ, Brooks EM, Bonenfant NR,
Sokocevic D, Borg ZD, Goodwin M, Loi R, Cruz F, Dunaway CW, Steele
C and Weiss DJ: Mesenchymal stromal cells mediate Aspergillus
hyphal extract-induced allergic airway inflammation by inhibition
of the Th17 signaling pathway. Stem Cells Transl Med. 3:194–205.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim HP and Leonard WJ: CREB/ATF-dependent
T cell receptor-induced FoxP3 gene expression: A role for DNA
methylation. J Exp Med. 204:1543–1551. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Goodwin M, Sueblinvong V, Eisenhauer P,
Ziats NP, LeClair L, Poynter ME, Steele C, Rincon M and Weiss DJ:
Bone marrow-derived mesenchymal stromal cells inhibit Th2-mediated
allergic airways inflammation in mice. Stem Cells. 29:1137–1148.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Giuliani M, Fleury M, Vernochet A,
Ketroussi F, Clay D, Azzarone B, Lataillade JJ and Durrbach A:
Long-lasting inhibitory effects of fetal liver mesenchymal stem
cells on T-lymphocyte proliferation. PLoS One. 6:e199882011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fontenot JD, Gavin MA and Rudensky AY:
Foxp3 programs the development and function of CD4+CD25+ regulatory
T cells. Nat Immunol. 4:330–336. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ge X, Bai C, Yang J, Lou G, Li Q and Chen
R: Intratracheal transplantation of bone marrow-derived mesenchymal
stem cells reduced airway inflammation and up-regulated
CD4+CD25+ regulatory T cells in asthmatic
mouse. Cell Biol Int. 37:675–686. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ivanov II, McKenzie BS, Zhou L, Tadokoro
CE, Lepelley A, Lafaille JJ, Cua DJ and Littman DR: The orphan
nuclear receptor RORgammat directs the differentiation program of
proinflammatory IL-17+ T helper cells. Cell. 126:1121–1133. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao J, Lloyd CM and Noble A: Th17
responses in chronic allergic airway inflammation abrogate
regulatory T-cell-mediated tolerance and contribute to airway
remodeling. Mucosal Immunol. 6:335–346. 2013. View Article : Google Scholar : PubMed/NCBI
|