Introduction
Studies have demonstrated that mechanical unloading
may result in severe bone loss, as observed in astronauts
undergoing long-term space flight or in patients subjected to
long-duration immobility or bedrest (1,2).
Decreased bone formation caused by abnormal osteoblast function is
considered to be one of the primary causes of bone loss (3). Studies have demonstrated that real or
simulated microgravity may affect the processes of osteoblast
proliferation, differentiation and mineralization (4–6).
Genomic analysis revealed that the expression of hundreds of genes
in osteoblasts is altered upon exposure to microgravity (7,8).
Efforts have been made to identify and characterize the key
regulators or genes that control alterations in osteoblasts.
However, existing studies have not completely clarified the reason
for the delay in osteoblast development induced by microgravity; it
is therefore necessary to further examine more regulatory
mechanisms, in order to obtain a deep insight into this area and to
develop effective preventative measures.
Previously, high throughput genomic analysis and
transcriptome sequencing demonstrated that much of the genome is
transcribed to noncoding RNAs (ncRNAs) (9). Increasing numbers of ncRNAs, which
were previously considered to be transcriptional ‘noise’, have been
observed to be important regulators of gene expression in
development, physiology and disease (10,11).
A subset of these ncRNAs whose strand length exceeds 200
nucleotides are termed long noncoding RNAs (lncRNAs) (12). Studies have demonstrated that
lncRNAs are involved in a range of important cellular processes,
including chromatin modification, RNA processing and gene
transcription (11). ncRNAs are
able to regulate gene expression in close genomic proximity
(cis-acting) or target distant transcriptional activators or
inhibitors (trans-acting) via diverse mechanisms (13). Such regulatory effects have been
additionally observed in the osteogenic differentiation of
mesenchymal stem cells (MSCs). lncRNA-maternally expressed 3, which
is located near the bone morphogenetic protein 4 (BMP4) gene locus,
is able to dissociate the repressor transcription factor SOX-2 from
the BMP4 promoter and thereby activate the transcription of BMP4,
resulting in the osteogenic differentiation of bone marrow MSCs
(14). These previous studies
suggested that lncRNAs serve an important role upstream of gene
expression during normal osteoblast development. However, there are
no reports of lncRNA expression patterns in the
microgravity-induced inhibition of osteogenic development, and
whether mechanical signal-mediated osteoblast differentiation
relies on the modulation of lncRNA expression remains unclear.
The present study screened for
differentially-expressed lncRNAs and mRNAs in the MC3T3-E1
pre-osteoblast cells, prior to and following exposure to simulated
microgravity, using microarray profiling. A series of bioinformatic
analyses, including Gene Ontology (GO) analysis, pathway analysis,
genomic context analysis and co-expression analysis, were used to
predict potential functional lncRNAs. The results of the present
study suggested that lncRNAs may serve important roles during
osteoblast differentiation under simulated microgravity
conditions.
Materials and methods
Cell culture and osteogenic
differentiation
Mouse MC3T3-E1 pre-osteoblast cells were obtained
from the Chinese Academy of Sciences Cell Bank (Shanghai, China)
and cultured in Dulbecco's modified Eagle's medium (DMEM)
containing 10% fetal bovine serum (FBS) (both from Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA), 100 U/ml penicillin G
and 100 mg/ml streptomycin, at 37°C in a humidified atmosphere of
5% CO2 in air. To induce osteoblast differentiation, the
DMEM was supplemented with 10% FBS, 0.1 mM dexamethasone, 10 mM
β-glycerophosphate, and 50 µg/ml ascorbic acid, and was changed
every 2–3 days.
mRNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total mRNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
according to the manufacturer's instructions. The RNA concentration
was quantified using a NanoDrop 1000 Spectrophotometer (Thermo
Fisher Scientific, Inc.). RNA quality was tested using an Agilent
2100 bioanalyzer (Agilent Technologies, Inc., Santa Clara, CA, USA)
and measured using the RNA integrity number (RIN). RNA samples were
submitted for further analysis if the RIN score was >5.0.
For RT, first-strand cDNA was synthesized using the
PrimeScript® RT reagent kit (cat. no. DRR037; Takara
Bio, Inc., Otsu, Japan). The expression levels of target genes were
determined quantitatively using a CFX96 qPCR system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) using SYBR®
Premix Ex Taq™ II (cat. no. DRR820A; Takara Bio, Inc.),
according to the manufacturer's instructions. The primer pairs were
as follows: Runt-related transcription factor 2 (Runx2; GenBank
accession no. NM_053470) forward, 5′-CCATAACGGTCTTCACAAATCC-3′ and
reverse, 5′-GCGGGACACCTACTCTCATACT-3′; transcription factor Sp7
(Osx; accession no. NM_001037632) forward,
5′-CAGTAATCTTCGTGCCAGACC-3′ and reverse,
5′-CTTCTTTGTGCCTCCTTTTCC-3′; polypyrimidine tract-binding protein 2
(Ptbp2; accession no. NM_001310711) forward,
5′-AAAGTCGCTCTGAGTTGTTAT-3′ and reverse,
5′-GCGAAGAGTTTGTCCTCAACC-3′; transportin-1 (Tnpo1; accession no.
NM_001048267) forward, 5′-AATTCGCGGTGACTCAGTCTGG-3′ and reverse,
5′-TCCATCTTGGTTTGCGAGGC-3′; exostosin-1 (Ext1; accession no.
NM_010162) forward, 5′-GAAGAGCACAGTGGTCGGAA-3′ and reverse,
5′-CTCGATGGCCGCTAGAATGT-3′; NONMMUT044983 (NONCODE gene ID
NONMMUG027774.1) forward, 5′-CGGCAGGCCTAGTCTTGTAT-3′ and reverse,
5′-ACAGCAGAGAGAGCCAAGGA-3′; NONMMUT023474 (gene ID NONMMUG014520.1)
forward, 5′-TCTCGAACCCTAGGAGAGCA-3′ and reverse,
5′-GGGACAAGGTAAATGGCTCA-3′; NONMMUT018832 (gene ID NONMMUG011720.1)
forward, 5′-AACATCTGAGGCTTGGCACT-3′ and reverse,
5′-TCATGGTACTGGCATCTCCA-3′; and GAPDH (accession no. NM_008084)
forward, 5′-CAGTGCCAGCCTCGTCTCAT-3′ and reverse,
5′-AGGGGCCATCCACAGTCTTC-3′. GAPDH was used as an internal control.
The qPCR thermal cycling was performed as follows: Initial
incubation for 15 sec at 95°C, followed by denaturing for 40 cycles
at 95°C for 5 sec and annealing for 31 sec at 60°C. The relative
expression of target genes was calculated using the
2−ΔΔCq method (15).
Alkaline phosphatase (ALP) activity
assay
Cells were seeded at 1×106 cells/well in
6-well plates (Corning Inc., Corning, NY, USA) and cultured for 24
h. When ALP activity was determined, confluent cell layers were
washed with PBS, lysed with 0.1 mol/l M-PER mammalian protein
extraction reagent (Pierce; Thermo Fisher Scientific, Inc.) for
15–30 min, and centrifuged at 12,000 × g for 15 min at room
temperature, according to the manufacturer's instructions. The
supernatants were collected for determining ALP activity using an
ALP assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing,
China). Protein concentrations were measured using a Bicinchoninic
Acid Protein Assay kit (Pierce; Thermo Fisher Scientific, Inc.).
ALP activity (IU/l) was defined as the release of 1 nmol
p-nitrophenol/min/µg total cellular protein.
Simulated microgravity exposure
The Rotating Wall Vessel Bioreactor (RWVB) clinostat
is an effective, ground-based device which is used to simulate
microgravity (7). Microgravity is
achieved by keeping cells rotating uniformly around a horizontal
axis. Therefore, there is a vector-averaged reduction in the
apparent gravity acting on the cells when rotated by 360°. A
2D-RWVB (China Astronaut Research and Training Center, Beijing,
China) was used in the present study, as previously described
(16). MC3T3-E1 cells were seeded
on coverslips and incubated until cell confluence reached 70%. The
coverslips were subsequently fixed in the bioreactor and placed
12.5 mm away from the rotational axis. The bioreactor was
completely filled with culture medium. Gentle aspiration was
performed to clear away air bubbles in order to avoid shear stress
during rotation. The bioreactor was fixed onto the clinostat and
rotated around a horizontal axis at 24 rpm. The group rotating
around a vertical axis was regarded as the control. The entire
system was placed in a humidified incubator at 37°C under 5%
CO2.
Microarray profiling
An Affymetrix GeneChip® Mouse
Transcriptome assay (version 1.0; Affymetrix Inc., Santa Clara, CA,
USA) provided global profiling of transcripts in the mouse genome,
which were selected from the most authoritative databases,
including NONCODE (www.noncode.org/index.php) and ENSEMBL (www.ensembl.org/index.html). In the microarray,
every transcript was detected with 10 specific probes in order to
improve the confidence of the statistical results. Protein-coding
and non-coding genes were represented on a separate array to supply
coincident hybridization. RNA labeling and array hybridization were
performed according to the manufacturer's protocol. The microarray
work was performed by Shanghai Ming Information Technology Co.,
Ltd. (Shanghai, China).
GO and pathway analysis
GO analysis was applied to analyze the primary
function of the differentially-expressed genes according to the
principles of Gene Ontology, which is able to organize genes into
hierarchical categories and reveal the gene regulatory network on
the basis of biological process and molecular function (17). Pathway analysis was additionally
performed to examine the significant pathways of the
differentially-expressed genes according to the Kyoto Encyclopedia
of Genes and Genomes, Biocarta and Reatome databases (18). An online Bioinformatics enrichment
tool (DAVID; david.ncifcrf.gov) was used to
perform the GO and pathway analysis in the present study (19). Biological processes of GO terms
were illustrated in the GO analysis. The P-value indicated the
significance of GO term and pathway term enrichment in the
differentially-expressed mRNA list (P<0.05 was considered to be
statistically significant).
Construction of the lncRNA-mRNA
co-expression network
A co-expression regulatory network is an undirected
graph, where each node corresponds to coding or ncRNAs, and a pair
of nodes is connected by an edge if there is a significant
co-expression association between them (20). lncRNA-mRNA networks were built to
identify the interactions between the differentially-expressed
genes and lncRNAs, according to the normalized signal intensity of
specific expression in the gene and lncRNA. For lncRNA-gene pair,
the Pearson correlation was calculated and the significant
correlation pairs with which to construct the network were selected
(21). In network analysis, degree
centrality, which is defined as the number of connections between
one node and another, is the simplest and most important measure of
the centrality of a gene or lncRNA within a network which
determines its relative importance (22).
Statistical analysis
Experiments were repeated a minimum of three times.
Data were statistically analyzed using SPSS 19.0 software and
expressed as the mean ± standard deviation. A one-way repeated
measures analysis of variance followed by Dunnett's post hoc test
was used to compare the time course-dependent variables. Two group
comparisons were performed using a two-tailed t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Differentially-expressed lncRNAs and
mRNAs in MC3T3-E1 cells exposed to simulated microgravity
MC3T3-E1 cells were cultured in an RWVB clinostat
and tested following rotation for 72 h. The effect of simulated
microgravity on osteoblasts was assessed by observing the gene
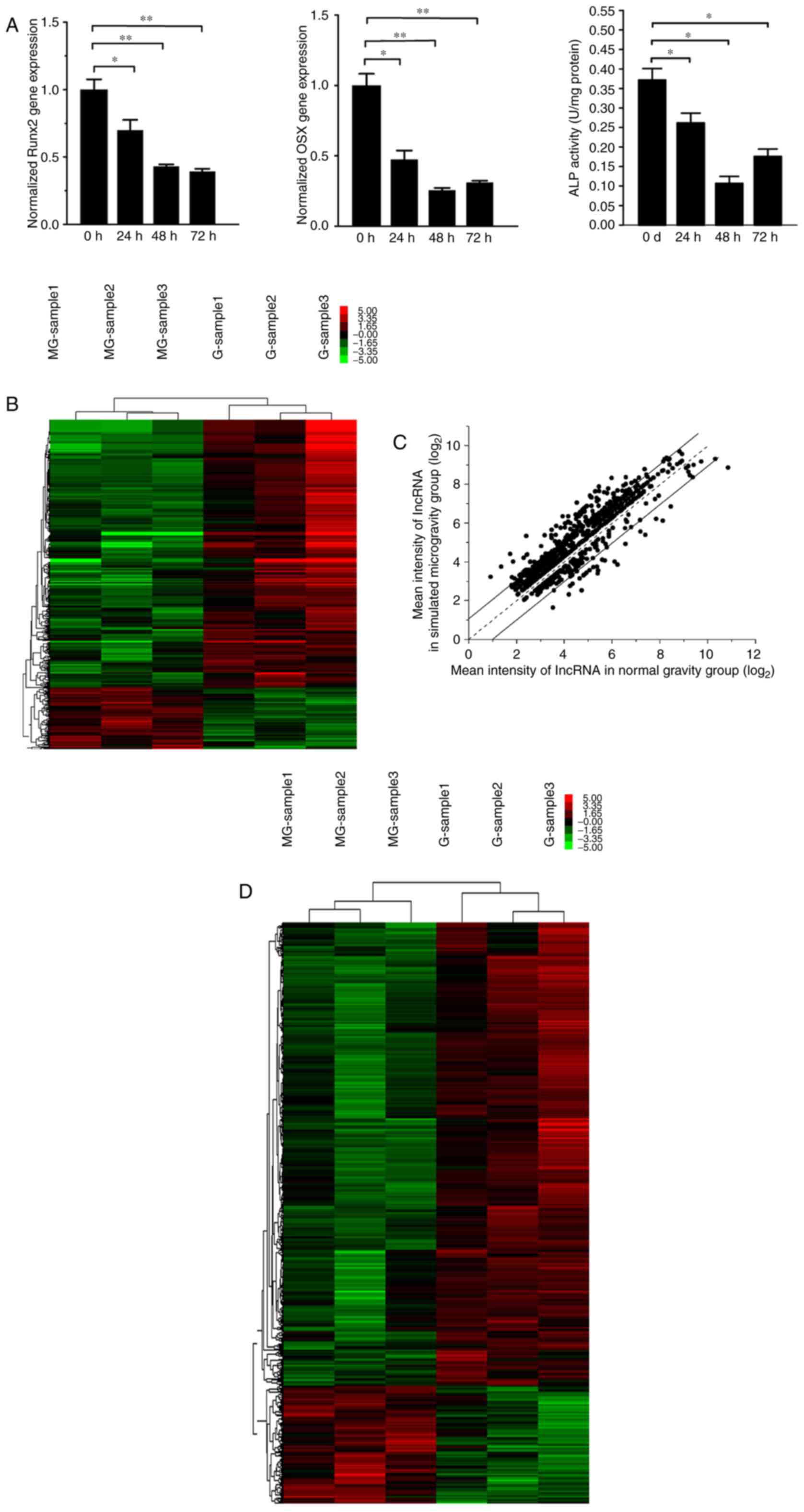
expression of Runx2 and Osx, in addition to the activity of ALP,
which were considered to be important transcription factors for
osteoblast differentiation. The data demonstrated that the gene
expression of Runx2 and Osx and the protein activity of ALP were
significantly decreased, of which, Osx and ALP reached lowest
levels at 48 h and Runx2 decreased slightly further at 72 h
(Fig. 1A). The results suggested
that osteo-differentiation process of MC3T3-E1 cells was inhibited
by simulated microgravity.
In order to detect the differentially-expressed
lncRNAs and mRNAs in MC3T3-E1 when exposed to simulated
microgravity, a transcriptome assay was performed to compare
expression profiles between the normal gravity and simulated
microgravity groups. The expression levels of 1,481 ncRNAs were
observed to be significantly altered, which is presented in the
hierarchical clustering analysis heat map (Fig. 1B). Among them, 857 lncRNAs whose
length exceeded 200 nucleotides, including 168 upregulated and 689
downregulated, were screened out and are exhibited on a scatter
plot (Fig. 1C). In addition, 2,264
mRNAs were demonstrated to be differentially-expressed, including
459 upregulated mRNAs and 1,805 downregulated mRNAs, presented on a
clustering heat map (Fig. 1D).
Construction of the co-expression
network between differentially-expressed lncRNAs and mRNAs
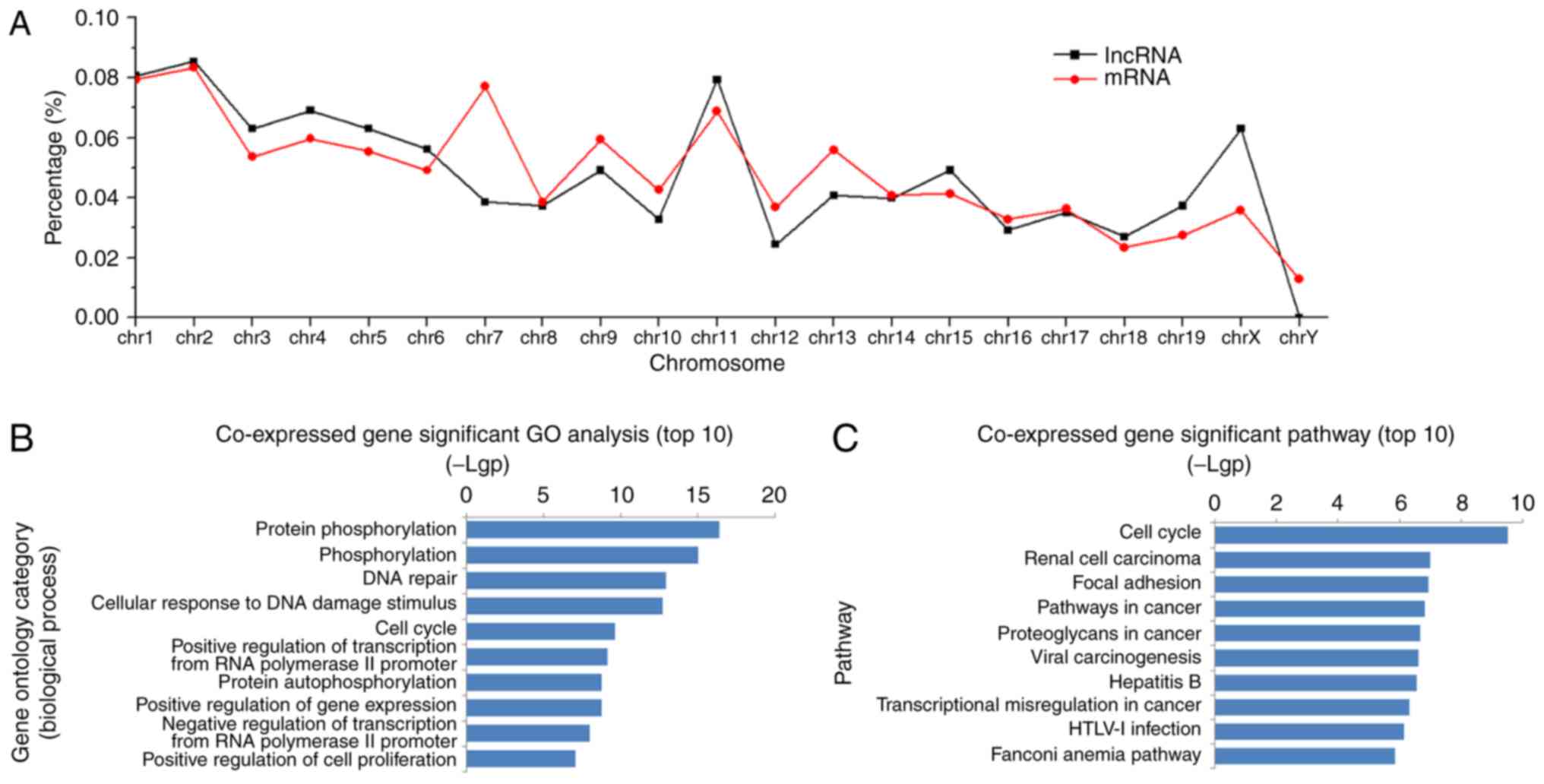
Locational distributions of the aberrantly-expressed
lncRNAs and mRNAs were analyzed synchronously. Significantly
altered lncRNAs and mRNAs were spread across the chromosomes, with
the largest amounts on chromosomes 1, 2 and 11. A relatively
consistent variation pattern was observed between aberrant lncRNA
and mRNA species, except for on chromosome 7 (Fig. 2A). In order to elucidate the
detailed association, a large and complex lncRNA-mRNA co-expression
network was generated according to the normalized signal intensity
of differentially-expressed lncRNAs and mRNAs. This co-expression
network consisted of 916 nodes and 4,813 connections between 354
lncRNAs and 562 mRNAs. Within this network, there were 3,798 pairs
presenting as positive regulatory associations and 1,015 pairs as
negative associations. Subsequently, mRNAs in the network were
submitted for GO enrichment analysis. The results demonstrated that
the aberrant mRNAs were frequently enriched in such biological
process as phosphorylation, response to DNA damage, the cell cycle,
regulation of gene expression and cell proliferation (Fig. 2B), and were involved in the cell
signaling pathways of the cell cycle, focal adhesion, cancer and
viral infection (Fig. 2C).
Genomic context analysis of the
aberrantly-expressed lncRNAs
In order to further elicit information from the
co-expression network and examine the possible functional lncRNAs,
the nearby mRNAs located within 100 kb up or downstream of
aberrantly-expressed lncRNAs were identified and matched with the
aberrantly-expressed mRNAs in the microarray data. Among them, 132
differentially-expressed lncRNAs and nearby mRNAs pairs were
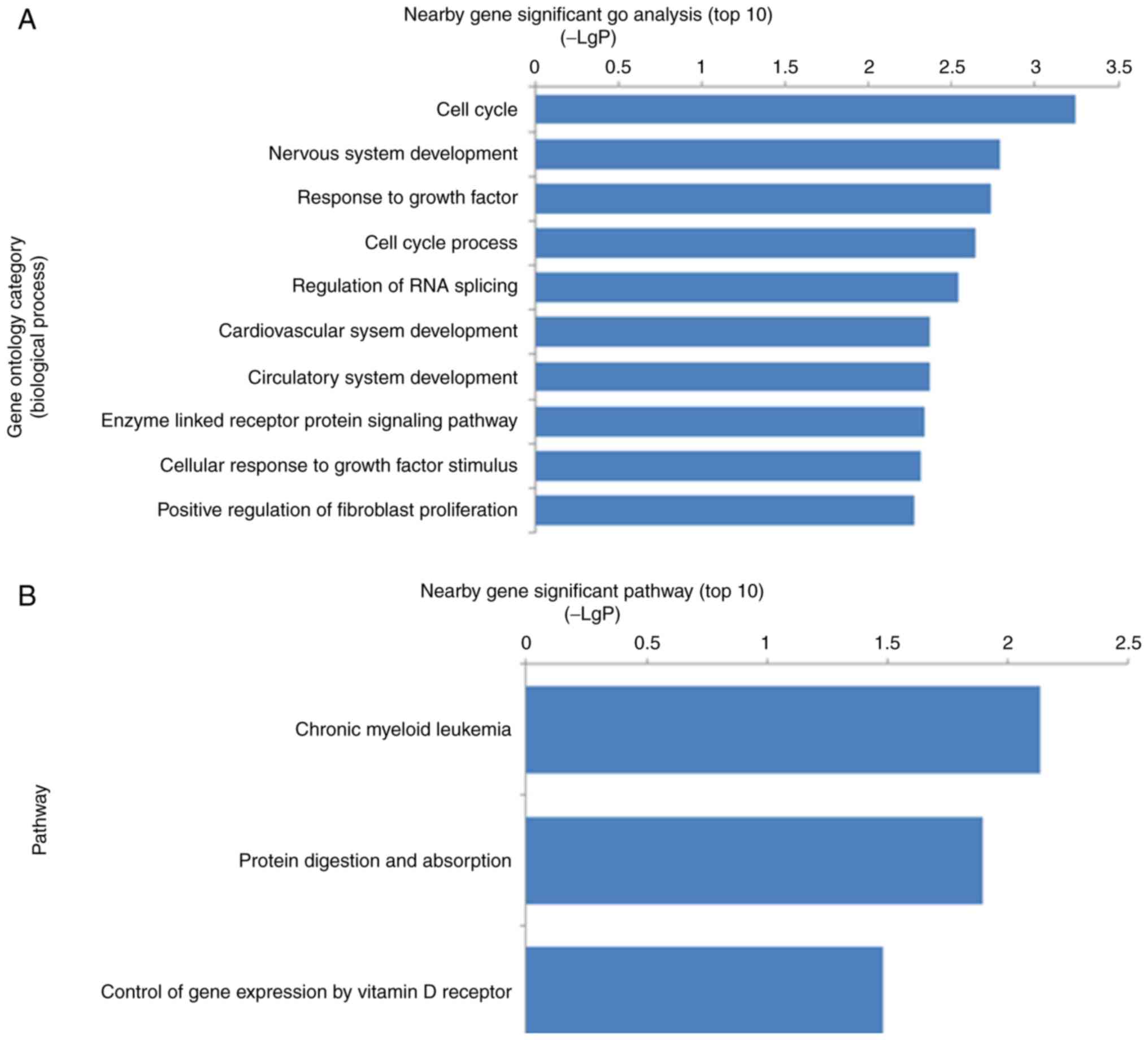
identified (data not shown). GO analysis indicated that the nearby
genes were involved in the significant biological processes,
including the cell cycle, nervous system development, response to
growth factors, regulation of RNA splicing, circulatory system
development, enzyme-linked receptor protein signaling pathway and
positive regulation of fibroblast proliferation (Fig. 3A). Pathway analysis demonstrated
that the most significant pathways were chronic myeloid leukemia,
protein digestion and absorption, and control of gene expression by
vitamin D receptor (Fig. 3B). A
literature review further demonstrated that ≥15 mRNAs included in
17 lncRNA-mRNA pairs have been reported to be involved in the
regulation of osteoblast proliferation, differentiation and
mineralization (Table I).
 | Table I.Osteoblast function-associated genes
located near to deregulated lncRNAs. |
Table I.
Osteoblast function-associated genes
located near to deregulated lncRNAs.
| Author, year | Deregulated
lncRNAa | lncRNA fold
change | Nearby gene | Expression
styleb | Location of nearby
gene |
Osteoblast-associated function of nearby
genec | (Refs.) |
|---|
| Hong et al,
2010 | NONMMUT000809 | 0.47 | Col5a2 | Downregulated | Same strand with
overlap | Positive on
differentiation | (23) |
| Verlinden et
al, 2013 | NONMMUT001206 | 0.83 | Nrp2 | Downregulated | Same strand with
overlap | Positive on
proliferation | (24) |
| Tang et al,
2011 | NONMMUT044983 | 0.54 | Ptbp2 | Downregulated | Same strand with
overlap | Positive on
development | (25) |
| Lekva et al,
2012 | NONMMUT044301 | 0.81 | Txnip | Downregulated | Complementary with
overlap | Negative on
differentiation | (26) |
| Ogasawara et
al, 2004 |
KnowTID_00005517 | 0.83 | Cdk6 | Downregulated | Upstream, 10,000
bp | Negative on
differentiation | (27) |
| Fan et al,
2016 | NONMMUT054273 | 0.80 | Ptpn11 | Downregulated | Same strand with
overlap | Positive on
differentiation | (28) |
| Sun et al,
2015 | NONMMUT058601 | 0.51 | Cacna1c | Downregulated | Same strand with
overlap | Positive on
proliferation | (29) |
| Chang et al,
2009 |
KnowTID_00006206 | 0.82 | Cdkn1b | Downregulated | Upstream, 10,000
bp | Positive on
differentiation | (30) |
| Yano et al,
2014 | NONMMUT055574 | 0.54 | Col1a2 | Downregulated | Same strand with
overlap | Positive on
differentiation | (31) |
| Haasper et
al, 2008 |
KnowTID_00006709 | 3.21 | Fosb | Upregulated | Upstream, 10,000
bp | Positive on
differentiation | (32) |
| Choi et al,
2015 |
KnowTID_00007668 | 0.66 | Cbl | Downregulated | Upstream, 10,000
bp | Negative on
differentiation | (33) |
| Yi et al,
2017 | NONMMUT069918 | 0.42 | Tcf12 | Downregulated | Same strand with
overlap | Negative on
differentiation | (34) |
| Di et al,
2014 | NONMMUT018832 | 0.68 | Tnpo1 | Downregulated | Complementary
strand with overlap | Positive on
differentiation | (35) |
| Matsumoto et
al, 2010 | NONMMUT023472 | 0.55 | Ext1 | Downregulated | Same strand
overlap | Positive on
development | (36) |
| Matsumoto et
al, 2010 | NONMMUT023474 | 0.50 | Ext1 | Downregulated | Same strand with
overlap | Positive on
development | (36) |
| Matsumoto et
al, 2010 | NONMMUT023475 | 0.55 | Ext1 | Downregulated | Same strand with
overlap | Positive on
development | (36) |
| Lu et al,
2016 | NONMMUT025233 | 0.65 | Crebbp | Downregulated | Upstream, 10,000
bp | Positive on
differentiation | (37) |
Bioinformatics screening of potential
functional lncRNAs
In the co-expression network, degree centrality was
considered to be the simplest and most important measure by which
to estimate the core regulatory factors. In order to obtain more
accurate candidate lncRNAs, the degree centrality of the 17 lncRNAs
whose nearby genes had been reported to be associated with
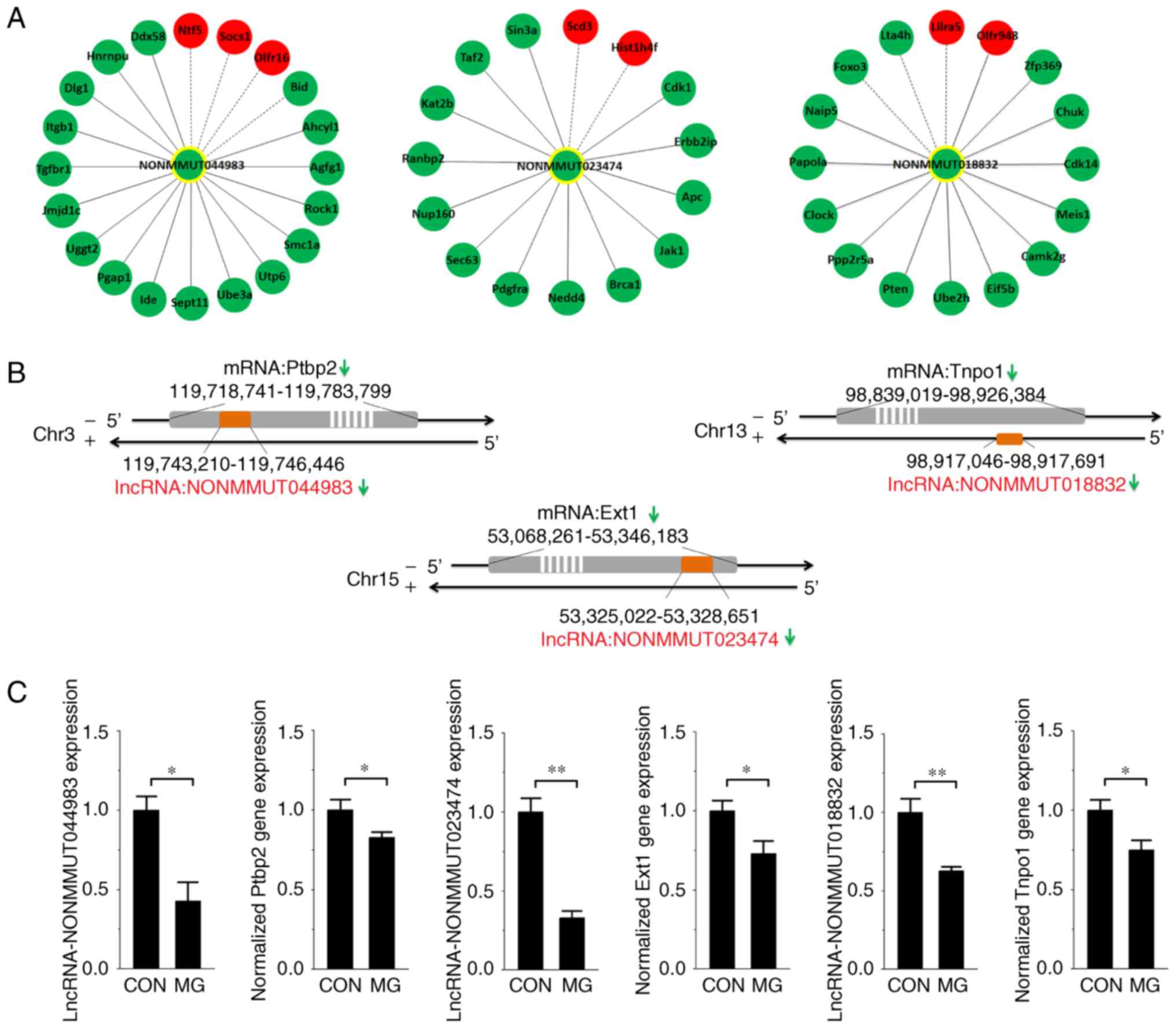
osteoblast function was calculated. The lncRNAs NONMMUT044983,
NONMMUT023474 and NONMMUT018832 were screened out via the degree
centrality analysis (degree >15). Their associations with
deregulated mRNAs were obtained from the overall co-expression
network and rebuilt (Fig. 4A). The
locations of the lncRNAs compared with mRNAs on the chromosome are
exhibited in Fig. 4B. In addition,
RT-qPCR analysis was performed to determine the expression of the
three lncRNAs and their nearby genes in MC3T3-E1 cells exposed to
the RWVB clinostat for 48 h. The results of the present study
demonstrated that the expression of each lncRNA and mRNA was
significantly decreased under simulated microgravity (Fig. 4C), consistent with the microarray
data.
Discussion
Osteoblasts, the main functional cells of bone
formation, have been reported to be associated with the bone loss
induced by microgravity (38,39).
A number of important regulators or genes have been studied to
examine how microgravity may affect osteoblast function. In the
present study, global genome analysis was performed and it was
observed that 857 lncRNAs and 2,264 mRNAs were significantly
altered in MC3T3-E1 cells exposed to simulated microgravity.
Bioinformatic analyses were performed to identify potential
functional lncRNAs. To the best of our knowledge, the present study
is the first to suggest that lncRNAs may serve important roles in
osteoblast differentiation under simulated microgravity conditions
and provide a novel perspective on the effects of microgravity on
osteoblast function.
Previous studies have revealed that a number of
lncRNAs are not transcriptional noise and serve important
regulatory roles (11,13). However, unlike the relatively
definite regulatory mechanisms of miRNAs, lncRNA function may not
be accurately predicted from the sequence or structure. A previous
study demonstrated that the potential function of lncRNA may be
inferred from their co-expressed genes (40). In the present study, a relatively
consistent variation pattern, except for on chromosome 7, was
observed between the differentially-expressed lncRNAs and mRNAs,
suggesting that they were more co-expressed or inversely-expressed
than may be expected by chance. The co-expression regulatory
network was built according to the normalized signal intensity of
the differentially-expressed lncRNAs and mRNAs. The co-expressed
lncRNAs/genes in the network were hypothesized to be controlled by
the same transcriptional regulatory pathway, to be functionally
similar, or to be members of the same pathway or protein complex
(41). Therefore, the functions of
the deregulated lncRNAs may cover the biological process of the
cell cycle, regulation of gene expression or cell proliferation,
which were inferred from functional enrichment analysis of their
co-expressed genes.
Apart from expression-based approaches, the genomic
context of lncRNAs is frequently studied in order to elucidate
their functions. An important localized regulatory mechanism
through which lncRNAs may affect the expression of nearby
protein-coding genes has been confirmed (42,43).
For example, lncRNA-Evf2, generated from the homeobox protein
Dlx-5/6 ultra conserved region, is able to act concurrently with
the transcription factor Dlx2 to increase the transcriptional
activity of the Dlx-5/6 enhancer in a target- and
homeodomain-specific manner (43).
Studying the nearby coding mRNAs may enhance the understanding of
the function and potential regulatory mechanisms of lncRNAs. In the
present study, 132 differentially-expressed lncRNAs and their
nearby coding mRNA pairs were identified. The 132
differentially-expressed mRNAs were involved in the significant
biological processes of the cell cycle, nervous system development
and response to growth factors, or in the signaling pathways of
chronic myeloid leukemia, protein digestion and absorption, and
control of gene expression by vitamin D receptor. In particular,
the vitamin D receptor pathway is associated with osteogenic
development (44). Certain
deregulated lncRNAs were more likely to affect osteoblast function
by acting on their nearby osteoblast-associated mRNAs. Notably, 15
of the nearby mRNAs have been previously reported to be involved in
osteoblast development. The corresponding 17 lncRNAs are therefore
important candidates for examining the function of lncRNAs in
osteoblast development.
To identify the most likely functional lncRNAs,
degree centrality within the co-expression network was considered.
Larger degree centrality values indicate an increased possibility
of a regulatory role. Therefore, the lncRNAs NONMMUT044983,
NONMMUT018832 and NONMMUT023474 were screened out via degree
centrality at the forefront. According to the degree of functional
association and similarity between co-expression genes, it may be
possible to infer the potential function of these lncRNAs from
their co-expressed genes, although GO analysis was not feasible due
to the insufficient sample size. Alternative analyses of the
functions of the three lncRNAs focused on the nearby genes. Their
nearby coding mRNAs exhibited an association with osteoblast
development, according to previous literature. Ptbp2 regulates the
mutually exclusive exons 8a and 8 in the CaV1.2 calcium channel
transcript (25), and the latter
serves fundamental roles in cellular responses to external stimuli,
including mechanical forces and hormonal signals, in osteoblastic
lineage bone cells (45,46). Tnpo1 is able to facilitate the
nuclear translocation of oxytocin receptors and result in
osteoblast maturation. The knockdown of Tnpo1 may abrogate the
oxytocin-induced expression of osteoblast differentiation genes
Osx, cyclic AMP-dependent transcription factor Atf4 and osteocalcin
(35). In addition, it has been
reported that conditional ablation of Ext1 may lead to
dysregulation of BMP signaling and severe skeletal defects
(36). Microarray and RT-qPCR
analyses suggested that the expression of the three lncRNAs and
their nearby genes were significantly decreased under simulated
microgravity. Therefore, it was hypothesized that downregulation of
these lncRNAs may abrogate the expression of their nearby
osteoblast-associated genes and result in inhibited osteogenesis
under simulated microgravity. Future studies are required to
investigate the potential regulatory mechanisms.
In conclusion, 857 differentially-expressed lncRNAs
were identified in the present study when the osteo-differentiating
MC3T3-E1 cells were exposed to simulated microgravity. A number of
potential functional lncRNAs, and the possible regulatory
mechanisms through which lncRNAs may control osteoblast
differentiation in a microgravity environment were predicted using
bioinformatics analysis. Although further experiment studies are
required to test this hypothesis, the results of the present study
demonstrated that lncRNAs are novel and potent candidates for
studies into the effects and mechanism of microgravity in
osteoblast function.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 31570939,
81701856, 81471815 and 31170889).
References
|
1
|
Bloomfield SA: Disuse osteopenia. Curr
Osteoporos Rep. 8:91–97. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ohshima H: Secondary osteoporosis UPDATE.
Bone loss due to bed rest and human space flight study. Clin
Calcium. 20:709–716. 2010.(In Japanese).
|
|
3
|
Morey ER and Baylink DJ: Inhibition of
bone formation during space flight. Science. 201:1138–1141. 1978.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zayzafoon M, Gathings WE and McDonald JM:
Modeled microgravity inhibits osteogenic differentiation of human
mesenchymal stem cells and increases adipogenesis. Endocrinology.
145:2421–2432. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rodionova NV: The dynamics of
proliferation and differentiation of osteogenic cells under
supportive unloading. Tsitol Genet. 45:22–27. 2011.PubMed/NCBI
|
|
6
|
Hu LF, Li JB, Qian AR, Wang F and Shang P:
Mineralization initiation of MC3T3-E1 preosteoblast is suppressed
under simulated microgravity condition. Cell Biol Int. 39:364–372.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Capulli M, Rufo A, Teti A and Rucci N:
Global transcriptome analysis in mouse calvarial osteoblasts
highlights sets of genes regulated by modeled microgravity and
identifies a ‘mechanoresponsive osteoblast gene signature’. J Cell
Biochem. 107:240–252. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Patel MJ, Liu W, Sykes MC, Ward NE, Risin
SA, Risin D and Jo H: Identification of mechanosensitive genes in
osteoblasts by comparative microarray studies using the rotating
wall vessel and the random positioning machine. J Cell Biochem.
101:587–599. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Claverie JM: Fewer genes, more noncoding
RNA. Science. 309:1529–1530. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kapranov P, Cheng J, Dike S, Nix DA,
Duttagupta R, Willingham AT, Stadler PF, Hertel J, Hackermüller J,
Hofacker IL, et al: RNA maps reveal new RNA classes and a possible
function for pervasive transcription. Science. 316:1484–1488. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mercer TR, Dinger ME and Mattick JS: Long
noncoding RNAs: Insights into functions. Nat Rev Genet. 10:155–159.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Brosnan CA and Voinnet O: The long and the
short of noncoding RNAs. Curr Opin Cell Biol. 21:416–425. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhuang W, Ge X, Yang S, Huang M, Zhuang W,
Chen P, Zhang X, Fu J, Qu J and Li B: Upregulation of lncRNA MEG3
promotes osteogenic differentiation of mesenchymal stem cells from
multiple myeloma patients by targeting BMP4 transcription. Stem
Cells. 33:1985–1997. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hu Z, Wang Y, Sun Z, Wang H, Zhou H, Zhang
L, Zhang S and Cao X: miRNA-132-3p inhibits osteoblast
differentiation by targeting Ep300 in simulated microgravity. Sci
Rep. 5:186552015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Draghici S, Khatri P, Tarca AL, Amin K,
Done A, Voichita C, Georgescu C and Romero R: A systems biology
approach for pathway level analysis. Genome Res. 17:1537–1545.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
da W Huang, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009.PubMed/NCBI
|
|
20
|
Stuart JM, Segal E, Koller D and Kim SK: A
gene-coexpression network for global discovery of conserved genetic
modules. Science. 302:249–255. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Prieto C, Risueño A, Fontanillo C and De
las Rivas J: Human gene coexpression landscape: Confident network
derived from tissue transcriptomic profiles. PLoS One. 3:e39112008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Barabási AL and Oltvai ZN: Network
biology: Understanding the cell's functional organization. Nat Rev
Genet. 5:101–113. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hong D, Chen HX, Yu HQ, Liang Y, Wang C,
Lian QQ, Deng HT and Ge RS: Morphological and proteomic analysis of
early stage of osteoblast differentiation in osteoblastic
progenitor cells. Exp Cell Res. 316:2291–2300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Verlinden L, Kriebitzsch C, Beullens I,
Tan BK, Carmeliet G and Verstuyf A: Nrp2 deficiency leads to
trabecular bone loss and is accompanied by enhanced osteoclast and
reduced osteoblast numbers. Bone. 55:465–475. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang ZZ, Sharma S, Zheng S, Chawla G,
Nikolic J and Black DL: Regulation of the mutually exclusive exons
8a and 8 in the CaV1.2 calcium channel transcript by polypyrimidine
tract-binding protein. J Biol Chem. 286:10007–10016. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lekva T, Ueland T, Bøyum H, Evang JA,
Godang K and Bollerslev J: TXNIP is highly regulated in bone
biopsies from patients with endogenous Cushing's syndrome and
related to bone turnover. Eur J Endocrinol. 166:1039–1048. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ogasawara T, Kawaguchi H, Jinno S, Hoshi
K, Itaka K, Takato T, Nakamura K and Okayama H: Bone morphogenetic
protein 2-induced osteoblast differentiation requires Smad-mediated
down-regulation of Cdk6. Mol Cell Biol. 24:6560–6568. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fan D, Liu S, Jiang S, Li Z, Mo X, Ruan H,
Zou GM and Fan C: The use of SHP-2 gene transduced bone marrow
mesenchymal stem cells to promote osteogenic differentiation and
bone defect repair in rat. J Biomed Mater Res A. 104:1871–1881.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun Z, Cao X, Hu Z, Zhang L, Wang H, Zhou
H, Li D, Zhang S and Xie M: miR-103 inhibits osteoblast
proliferation mainly through suppressing Cav1.2 expression in
simulated microgravity. Bone. 76:121–128. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chang SF, Chang TK, Peng HH, Yeh YT, Lee
DY, Yeh CR, Zhou J, Cheng CK, Chang CA and Chiu JJ: BMP-4 induction
of arrest and differentiation of osteoblast-like cells via p21 CIP1
and p27 KIP1 regulation. Mol Endocrinol. 23:1827–1838. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yano H, Hamanaka R, Nakamura-Ota M, Adachi
S, Zhang JJ, Matsuo N and Yoshioka H: Sp7/Osterix induces the mouse
pro-α2 (I) collagen gene (Col1a2) expression via the proximal
promoter in osteoblastic cells. Biochem Biophys Res Commun.
452:531–536. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Haasper C, Jagodzinski M, Drescher M,
Meller R, Wehmeier M, Krettek C and Hesse E: Cyclic strain induces
FosB and initiates osteogenic differentiation of mesenchymal cells.
Exp Toxicol Pathol. 59:355–363. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Choi YH, Han Y, Lee SH, Jin YH, Bahn M,
Hur KC, Yeo CY and Lee KY: Cbl-b and c-Cbl negatively regulate
osteoblast differentiation by enhancing ubiquitination and
degradation of Osterix. Bone. 75:201–209. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yi S, Yu M, Yang S, Miron RJ and Zhang Y:
Tcf12, A member of basic helix-loop-helix transcription factors,
mediates bone marrow mesenchymal stem cell osteogenic
differentiation in vitro and in vivo. Stem Cells. 35:386–397. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Di Benedetto A, Sun L, Zambonin CG, Tamma
R, Nico B, Calvano CD, Colaianni G, Ji Y, Mori G, Grano M, et al:
Osteoblast regulation via ligand-activated nuclear trafficking of
the oxytocin receptor. Proc Natl Acad Sci USA. 111:pp. 16502–16507.
2014; View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Matsumoto Y, Matsumoto K, Irie F, Fukushi
J, Stallcup WB and Yamaguchi Y: Conditional ablation of the heparan
sulfate-synthesizing enzyme Ext1 leads to dysregulation of bone
morphogenic protein signaling and severe skeletal defects. J Biol
Chem. 285:19227–19234. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lu J, Qu S, Yao B, Xu Y, Jin Y, Shi K,
Shui Y, Pan S, Chen L and Ma C: Osterix acetylation at K307 and
K312 enhances its transcriptional activity and is required for
osteoblast differentiation. Oncotarget. 7:37471–37486. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kacena MA, Todd P and Landis WJ:
Osteoblasts subjected to spaceflight and simulated space shuttle
launch conditions. In Vitro Cell Dev Biol Anim. 39:454–459. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bucaro MA, Fertala J, Adams CS, Steinbeck
M, Ayyaswamy P, Mukundakrishnan K, Shapiro IM and Risbud MV: Bone
cell survival in microgravity: Evidence that modeled microgravity
increases osteoblast sensitivity to apoptogens. Ann N Y Acad Sci.
1027:64–73. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Necsulea A, Soumillon M, Warnefors M,
Liechti A, Daish T, Zeller U, Baker JC, Grützner F and Kaessmann H:
The evolution of lncRNA repertoires and expression patterns in
tetrapods. Nature. 505:635–640. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Matthew T Weirauch: Gene coexpression
networks for the analysis of DNA microarray dataApplied Statistics
for Network Biology: Methods in Systems Biology. Dehmer M,
Emmert-Streib F, Graber A and Salvador A: Wiley-VCH Verlag GmbH
& Co. KGaA; Weinheim, Germany: 2011
|
|
42
|
Martianov I, Ramadass A, Barros A Serra,
Chow N and Akoulitchev A: Repression of the human dihydrofolate
reductase gene by a non-coding interfering transcript. Nature.
445:666–670. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Feng J, Bi C, Clark BS, Mady R, Shah P and
Kohtz JD: The Evf-2 noncoding RNA is transcribed from the Dlx-5/6
ultraconserved region and functions as a Dlx-2 transcriptional
coactivator. Genes Dev. 20:1470–1484. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Olivares-Navarrete R, Sutha K, Hyzy SL,
Hutton DL, Schwartz Z, McDevitt T and Boyan BD: Osteogenic
differentiation of stem cells alters vitamin D receptor expression.
Stem Cells Dev. 21:1726–1735. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Duncan RL, Akanbi KA and Farach-Carson MC:
Calcium signals and calcium channels in osteoblastic cells. Semin
Nephrol. 18:178–190. 1998.PubMed/NCBI
|
|
46
|
Iqbal J and Zaidi M: Molecular regulation
of mechanotransduction. Biochem Biophys Res Commun. 328:751–755.
2005. View Article : Google Scholar : PubMed/NCBI
|