Introduction
Presbycusis, also known as age-related hearing loss
(AHL), is a common neurodegenerative disorder and characterized by
gradual, progressive sensorineural hearing loss (1). Presbycusis affects 40% of patients
between 65 and 66 years old and 66.8% of patients between 73 and 74
years old (2). Presbycusis can
result in the speech processing deficits, the slowed central
processing of acoustic stimuli, the impaired sound localization,
social isolation, depression, and even cognitive impairment
(1,3–5).
Thus, have a better understanding of the pathogenesis of
presbycusis is of great importance to the therapy and prevention of
presbycusis.
It is well known that the intrastrial fluid-blood
barrier could separate the stria vascularis from peripheral
circulation. Especially, the integrity of the barrier is vital
important for maintaining inner ear homeostasis and sustaining the
endocochlear potential, which is an essential driving force for
hearing function (6–9). A previous study indicated that the
intrastrial fluid-blood barrier comprises a large number of
perivascular-resident macrophage-like melanocytes (PVM/Ms)
(10,11). Further study verified that absence
of PVM/Ms increases the permeability of the intrastrial fluid-blood
barrier to both low and high-molecular-weight tracers (12). Totally, these investigations may
indicate the important role of PVM/Ms in hearing function.
Macrophage migration inhibitory factor (MIF) is an
essential factor in axis and neural development. MIF is reported to
be strongly expressed in the inner ear and plays an important role
in inner ear of mice (13).
Additionally, MIF was proved to play an important role in cell
viability signal transmission, as well as in cell growth (14,15).
Thus, it raises the possibility that MIF may via regulating the
growth of PVM/Ms and exert important roles in hearing function.
To validate our speculation, PVM/Ms from stria
vascularis of lateral wall of cochlear in young and aged mice were
isolated. We found the expression of MIF was significantly
downregulated in aged mice. The viability of PVM/Ms in aged mice
was significantly decreased, and the apoptotic PVM/Ms number in
aged mice was markedly increased. Further studies showed that MIF
knockdown in young mice had more serious hearing loss as compared
to the control. MIF knockdown in PVM/Ms also revealed the
significant inhibited cell viability, the markedly induced cell
apoptosis as compared to the control. Totally, our results revealed
that MIF knockdown in vivo significantly accentuated hearing
loss in young mice, and MIF knockdown in vitro significantly
inhibited viability and induced apoptosis of PVM/Ms. Our study may
provide a potential therapy and prevention method for
presbycusis.
Materials and methods
Animals
All experiments were approved by the ethical
committee of the First Affiliated Hospital of Zhengzhou University.
C57BL/6J mice, which are known for their rapid development of
hearing loss, particularly in the high-frequency range, by one year
of age (16,17). Mice were purchased from the Chinese
Academy of Medical Sciences (Beijing, China). In our study, mice
aged 2 months and 10 months were termed as the young group and aged
group, respectively (n=20 in each group). The mice were housed 2 or
3 per cage and had free access to food and water, with the suitable
temperature and humidity conditions, along with a 12/12 h
light/dark cycle.
PVM/Ms isolation and culture
PVM/Ms were isolated from mice aged 2 months and 10
months, respectively, and were cultured as previously described
(12,18). The stria vascularis of lateral wall
of cochlear was firstly isolated. To produce PVM/Ms, the minced
stria vascularis was cultured on collagen-coated dishes in medium
254 (Invitrogen Life Technologies, Carlsbad, CA, USA), which
containing 10% FBS, 1% human melanocyte growth factor and 0.5%
gentamicin/amphotericin. The minced stria vascularis was incubated
at 37°C in 5% CO2 with the medium changed every 3 days.
Cell clones formed and melted at approximately 10 days. The cells
were detached from the cell colony with a solution of trypsin-EDTA
and then were purified. For purification of PVM/M cells, antibody
F4/80 or GST were added to the medium. Cells were incubated with
the antibodies for 10 min at 4°C, and then for isolation, and were
grown in medium 254 containing 1% human melanocyte growth
factor.
siRNA transfection
MIF silencing in vitro was performed on
passage-3 PVM/Ms seeded in 24-well plates as described previously
(12). The PVM/Ms
(1×105 cells/well) were transfected with si-MIF and
scrambled siRNA (Applied Biosystems) according to the
manufacturer's guidelines. After 4 days, the transfection
efficiency was determined by western blotting and cells were used
for cell viability and apoptosis assay.
The in vivo siRNA transfection was performed
as described previously (12,19).
Briefly, animals were anesthetized, and a 30-G needle was used to
make a single puncture in the anterior-inferior quadrant of the
tympanic membrane to allow exit of air from the middle ear during
drug injection. MIF was silenced with a 5-µl solution of siRNA (20
ng/µl) injected through the posterior-inferior quadrant. The middle
ear was filled completely with the solution for 5 days (n=3 mice
per group). Scrambled siRNA of the same concentration was given to
control group (n=3 mice per group). At 8 days after the in
vivo siRNA transfection, the transfection efficiency was
determined by western blotting and used for hearing tests.
Cell viability assay
Cells (4×103 cells/well) were seeded into
the 96-well plate overnight. MTT was added, and then incubated for
4 h. The formazan crystals formed from MTT by the living cells were
dissolved in 150 µl dimethyl sulfoxide (DMSO), and then detected
using a spectrophotometer at an absorption wavelength of 490
nm.
Flow cytometry analysis for
apoptosis
The Annexin V-FITC/PI Apoptosis Detection kit (Roche
Diagnostics, Rotkreuz, Switzerland) was used to detect cell
apoptosis. The cells were washed with cold PBS and resuspended.
Staining was performed according to the producer's manual. Flow
cytometry (BD Biosciences, Franklin Lakes, NJ, USA) was performed
immediately.
Hearing tests
The auditory brain stem response (ABR) test has been
described previously (20).
Briefly, hearing was measured with a click stimulus and at 4, 8, 16
and 32 kHz using tone pips (3 msec duration). Stimuli were
presented to both ears simultaneously at decreasing intensities (5
dB steps) in a recording system. For each animal for each stimulus,
the threshold response was recorded.
Reverse transcription-quantitative
polymerase chain reaction (qRT-PCR)
Total RNA from purified PVM/Ms was extracted
separately with a RNeasy kit (Qiagen, Valencia, CA, USA) according
to the manufacturer's suggestions. One microgram of total RNA was
reverse-transcribed using a SuperScript II (Invitrogen Life
Technologies) following the manufacturer's instructions. RT-qPCR
was performed using a SYBR-Green II RT-qPCR kit (Toyobo Co., Ltd.,
Osaka, Japan) and a Prism 7300 Sequence Detection System (Applied
Biosystems, Foster City, CA, USA). GAPDH was used as the internal
control.
Western blot analysis
Total protein was isolated from purified PVM/Ms and
the protein concentration was determined. SDS-PAGE was used to
separate the proteins and then the protein was transferred onto a
nitrocellulose membrane (Millipore Corp., Boston, MA, USA). The
membrane was then blocked and incubated with primary antibody.
Protein bands were detected by incubation with horseradish
peroxidase-conjugated secondary antibody and visualized with
chemiluminescence reagent (Pierce Chemical Co., Rockford, IL,
USA).
Statistical analysis
Data are presented as mean ± SEM. The unpaired
Student's t-test using Graph Pad Prism 5.0 (Hearne Scientific
Software, Chicago, IL, USA) was used to perform the statistical
analysis. P<0.05 was considered to indicate a statistically
significant difference.
Results
MIF was downregulated in PVM/Ms from
stria vascularis of lateral wall of cochlear in aged mice
PVM/Ms from stria vascularis of lateral wall of
cochlear in young and aged mice were isolated. Firstly, the
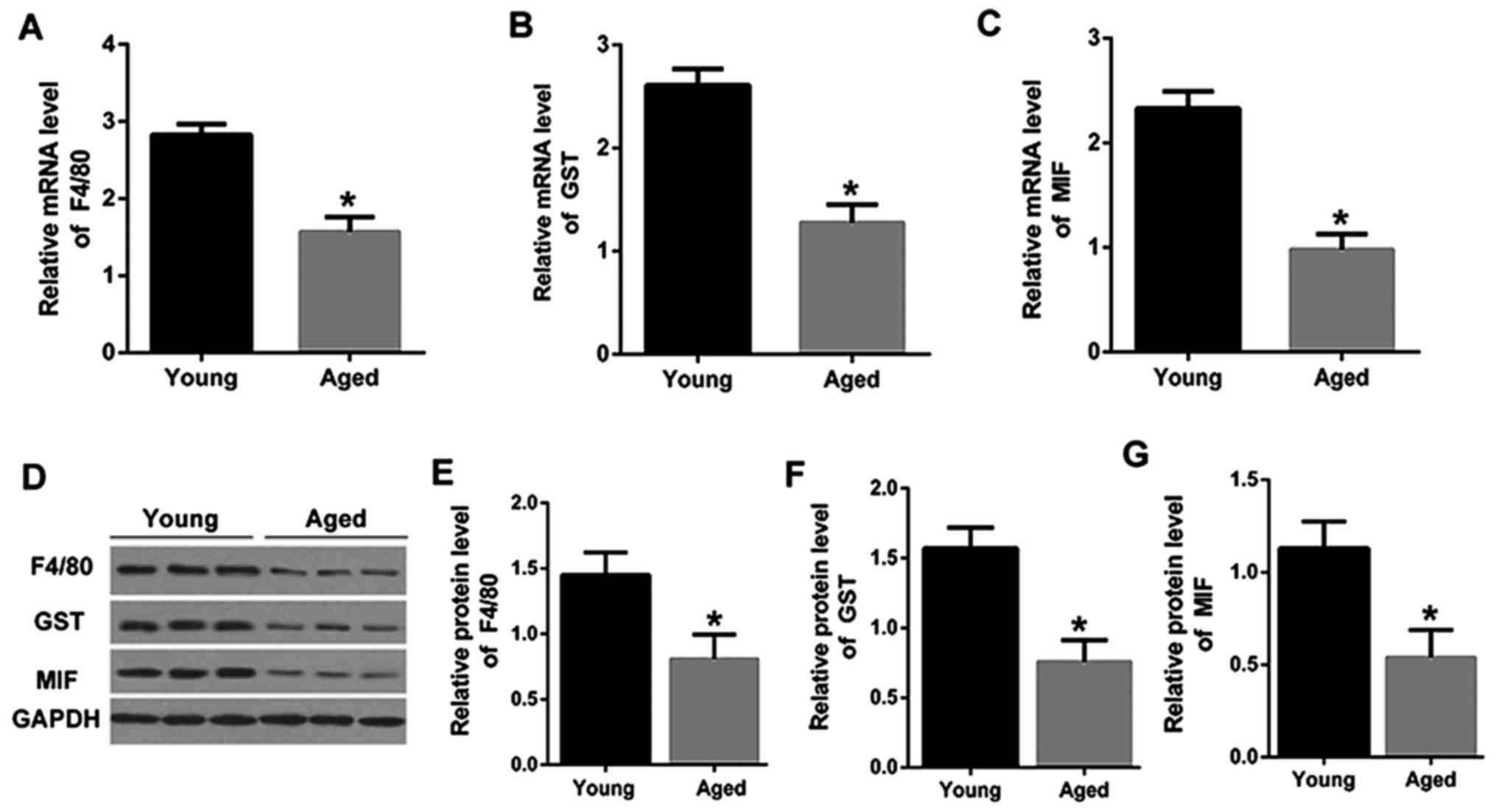
macrophage and melanocyte marker proteins, F4/80 and GST (21), were determined by qRT-PCR. As shown
in Fig. 1A and B, the mRNA
expression levels of F4/80and GST were significantly downregulated
in PVM/Ms from aged mice than that in the young mice. The mRNA
expression level of MIF was also significantly downregulated in
PVM/Ms from aged mice than that in the young mice (Fig. 1C). Consistently, the decreased
protein levels of F4/80, GST and MIF were also shown in aged mice
as compared to that in the young mice (Fig. 1D-G). The significant reduction in
F4/80 and GST indicated the decrease of PVM/Ms in aged mice,
revealing the vital role of PVM/Ms in aged mice. More importantly,
the reduction of MIF in aged mice was found, which may suggest the
potential role of MIF in PVM/Ms growth.
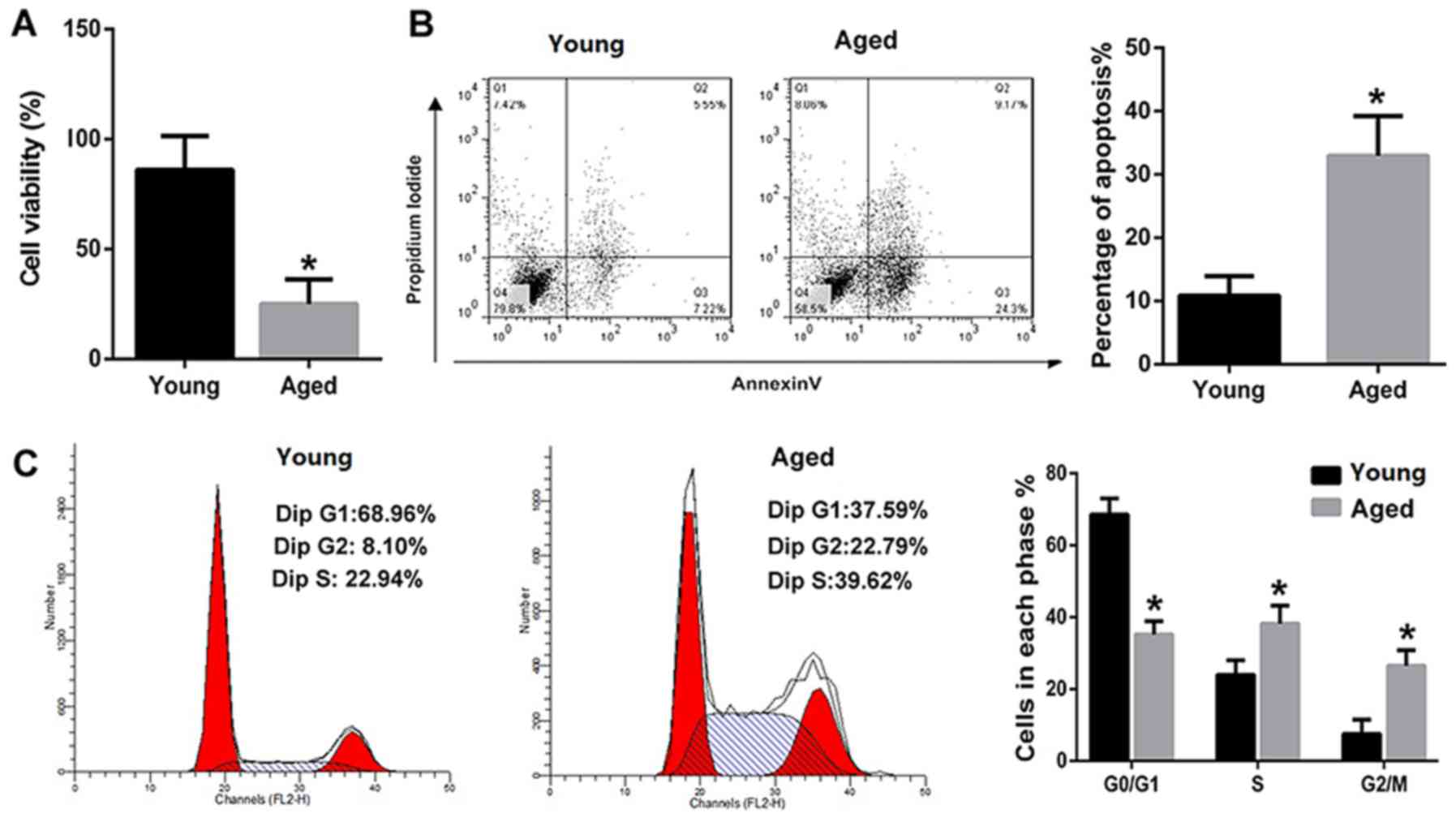
The viability of PVM/Ms was decreased
and the apoptotic PVM/Ms number was markedly increased in aged
mice
The viability and apoptosis of PVM/Ms from young and
aged mice were compared. The results revealed that the viability of
PVM/Ms from aged mice was significantly decreased as compared to
that from the aged mice (Fig. 2A).
Inversely, the apoptosis of PVM/Ms from aged mice was significantly
increased as compared to that from the aged mice (Fig. 2B). The cell population in G0/G1
phase of PVM/Ms from aged mice was significantly decreased as
compared to that from young mice; whereas the cell population in S
and G2/M phases of PVM/Ms from aged mice was significantly
increased as compared to that from young mice (Fig. 2C).
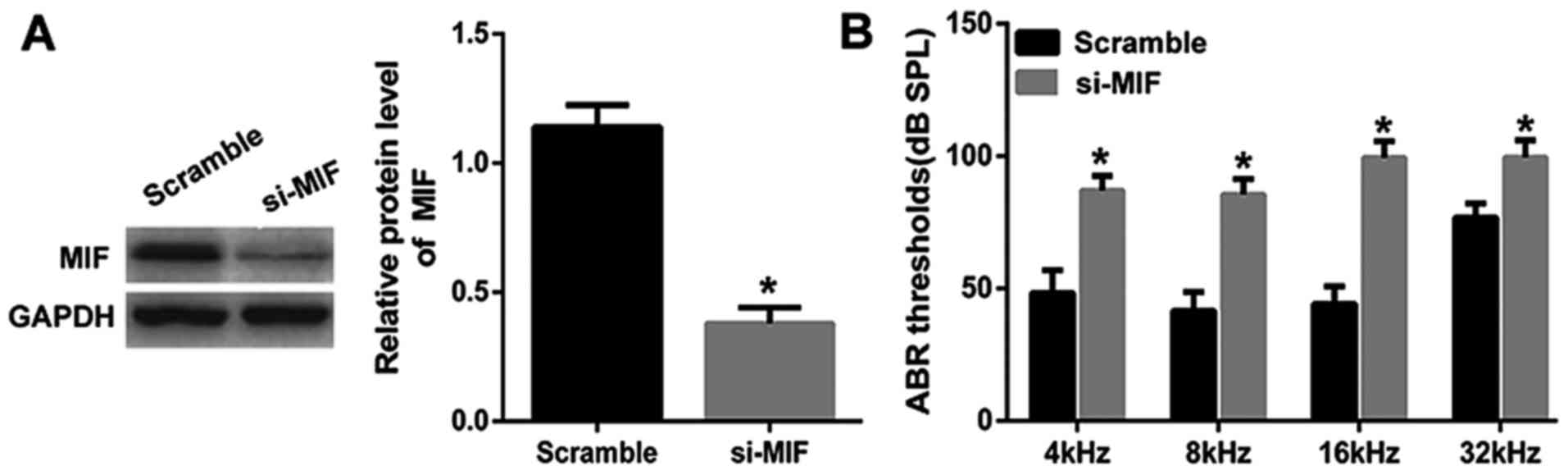
MIF knockdown in young mice leads to
hearing loss
Considering the decreased cell viability of PVM/Ms
and decreased protein level of MIF in aged mice, we knocked down
MIF in young mice to explore the important role of MIF. MIF was
knocked down in young mice and the in vivo transfection
efficiency was determined by the western blotting (Fig. 3A). Through the ABR test, the
results showed that the average threshold of the MIF knockdown
group was maintained at a significant lower level than the control
group, indicating the significant hearing loss in young mice with
MIF knockdown (Fig. 3B).
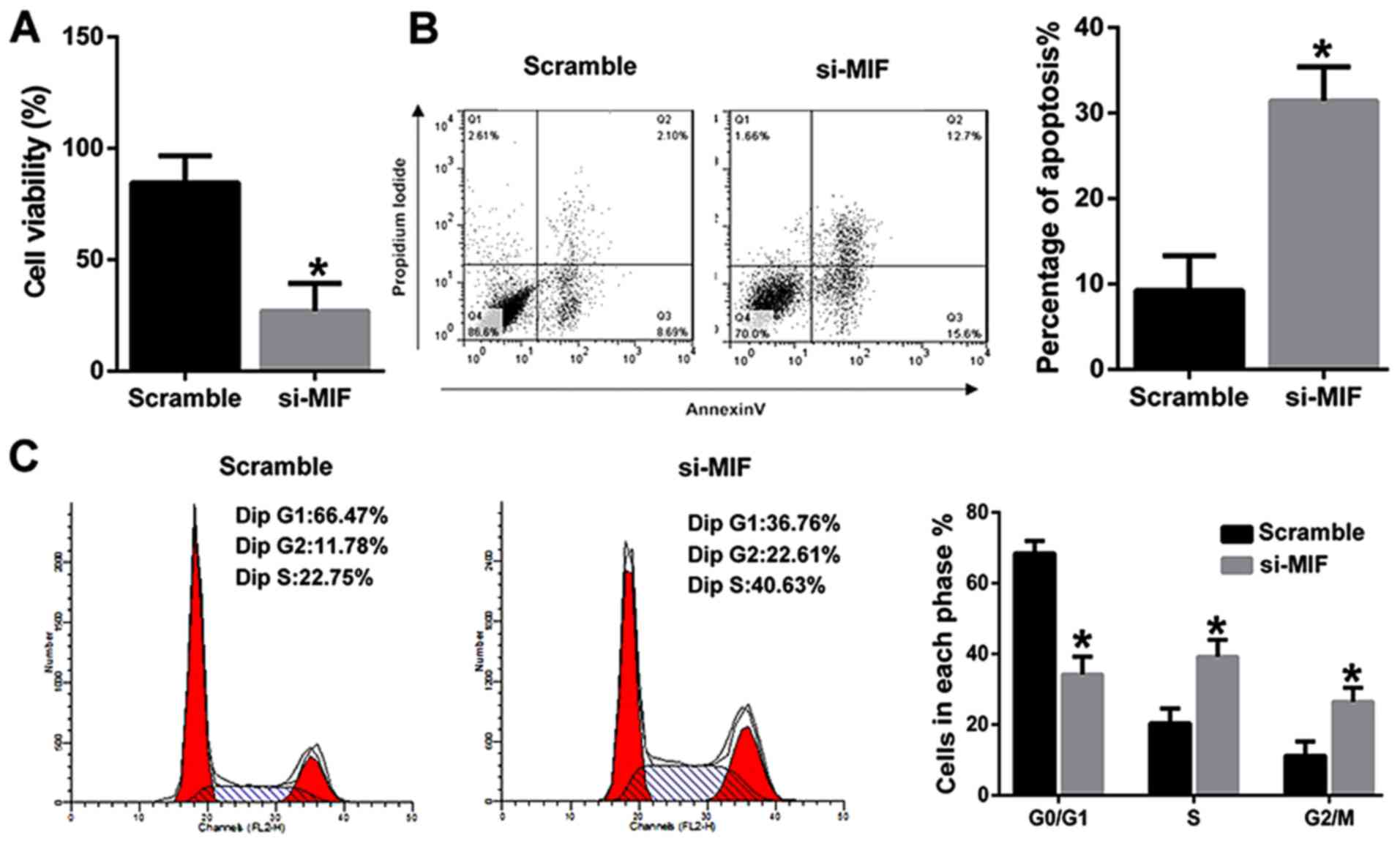
MIF knockdown inhibited the viability
and induced the apoptosis of PVM/Ms from young mice
To further explore the underlying mechanism, PVM/Ms
were isolated from young mice. MIF was then knocked down and
transfected into PVM/Ms, the viability and apoptosis were
determined respectively. The results revealed that MIF knockdown
significantly inhibited the viability of PVM/Ms as compared to the
control (Fig. 4A). Inversely, MIF
knockdown significantly induced the apoptosis of PVM/Ms as compared
to the control (Fig. 4B). The cell
population in G0/G1 phase of PVM/Ms with MIF knockdown was
significantly decreased as compared to the control; whereas the
cell population in S and G2/M phases of PVM/Ms with MIF knockdown
was significantly increased as compared to the control (Fig. 4C). Totally, we found that the
viability and apoptosis of PVM/Ms that transfected with si-MIF was
comparable to that in the aged mice (Fig. 2).
Discussion
In present study, we firstly found the significant
reduction in F4/80 and GST in aged mice. Since F4/80 and GST are
the marker proteins of macrophage and melanocyte (21), to some extent, the results
indicated the decrease of PVM/Ms in aged mice. We also found that
the viability of PVM/Ms was decreased and the apoptotic PVM/Ms
number was markedly increased in aged mice that that in young mice.
Besides, the cell population in G0/G1 phase of PVM/Ms from aged
mice was significantly decreased and the cell population in S and
G2/M phases of PVM/Ms from aged mice was significantly increased as
compared to that from young mice. The progression of cell cycle was
arrested at S and G2/M phases as compared to that from young mice.
To some extent, the results were consistent with the previous
results demonstrating that PVM/Ms are essential for maintaining
cochlear vascular architecture and stability (22). Altogether, these investigations
indicated the important role of PVM/Ms on hearing function. A
previous study has suggested than the PVM/Ms are in close contact
with vessels through cytoplasmic processes and have an important
role in maintaining the integrity of the intrastrial fluid-blood
barrier and hearing function. Besides, they also showed that
absence of PVM/Ms increases the permeability of the intrastrial
fluid-blood barrier to both low and high-molecular-weight tracers.
Besides, in the PVM/M-depleted animals, substantial drop in
endocochlear potential with accompanying hearing loss was found
(12).
Based on these results, it seems that factors that
affect PVM/Ms growth were also associated with the hearing
function. In our study, we found the reduction of MIF in aged mice.
Additionally, MIF knockdown in young mice not only leads to hearing
loss, but also inhibited the viability and induced the apoptosis of
PVM/Ms. Besides, the cell population in G0/G1 phase of PVM/Ms with
MIF knockdown was significantly decreased, and the cell population
in S and G2/M phases of PVM/Ms with MIF knockdown was significantly
increased as compared to the control. The progression of cell cycle
was arrested at S and G2/M phases as compared to the control. Dong
et al indicated that genes involved in apoptosis were
associated with human presbycusis, and provided important evidence
to support the suggestion that apoptosis was involved in the
pathogenesis of human presbycusis (23). MIF was reported to regulate cell
proliferation, survival and apoptosis in many studies (24–26).
However, the effects of MIF on the growth of PVM/Ms were not
reported previously. In our study, MIF may be identified as a
contribute factor to the pathogenesis of presbycusis.
MIF was observed in the spiral ligament, Reissner's
membrane, stria vascularis, saccular macula, spiral ganglion cells,
and membranous labyrinth, which suggest the important role of MIF
in the inner ear of mice (13).
Researchers also indicated that MIF-deficient mice had significant
loss of cochlear hair cells and prolonged hearing loss after
intense noise exposure, indicating that MIF may play an important
role in recovery from acoustic trauma and management of MIF may be
a novel therapeutic option for noise-induced hearing loss (27). However, the underlying mechanism
has not been clear yet, our study may provide a possibility, that
MIF may exert its role in hearing function through regulating
PVM/Ms growth.
Totally, our results revealed that MIF knockdown
in vivo significantly accentuated hearing loss in young
mice, and MIF knockdown in vitro significantly inhibited
viability and induced apoptosis of PVM/Ms. Our study may provide a
potential therapy and prevention method for presbycusis.
Acknowledgements
This study was supported by grants from the National
Natural Science Youth Foundation of China (no. 81500799) and Youth
Funds from the First Affiliated Hospital of Zhengzhou
University.
References
|
1
|
Heman-Ackah SE, Huang TC and Juhn SK:
R442-Antioxidant therapy prevents presbycusis In C57BL6 mice.
Otolaryngology Head & Neck Surgery. 139:2S12008. View Article : Google Scholar
|
|
2
|
Gopinath B, Rochtchina E, Wang JJ,
Schneider J, Leeder SR and Mitchell P: Prevalence of age-related
hearing loss in older adults: Blue mountains study. Arch Intern
Med. 169:415–416. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Frisina DR and Frisina RD: Speech
recognition in noise and presbycusis: Relations to possible neural
mechanisms. Hear Res. 106:95–104. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gates GA and Mills JH: Presbycusis.
Lancet. 366:1111–1120. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
del Campo HN Martin, Measor KR and Razak
KA: Parvalbumin immunoreactivity in the auditory cortex of a mouse
model of presbycusis. Hear Res. 294:31–39. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Juhn SK, Hunter BA and Odland RM:
Blood-labyrinth barrier and fluid dynamics of the inner ear. Int
Tinnitus J. 7:72–83. 2001.PubMed/NCBI
|
|
7
|
Juhn SK, Rybak LP and Prado S: Nature of
blood-labyrinth barrier in experimental conditions. Ann Otol Rhinol
Laryngol. 90:135–141. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Salt AN, Mleichar I and Thalmann MD:
Mechanisms of endocochlear potential generation by stria
vascularis. Laryngoscope. 97:984–991. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takeuchi S, Ando M, Sato T and Kakigi A:
Three-dimensional and ultrastructural relationships between
intermediate cells and capillaries in the gerbil stria vascularis.
Hear Res. 155:103–112. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dai M, Yang Y, Omelchenko I, Nuttall AL,
Kachelmeier A, Xiu R and Shi X: Bone marrow cell recruitment
mediated by inducible nitric oxide synthase/stromal cell-derived
factor-1alpha signaling repairs the acoustically damaged cochlear
blood-babyrinth barrier. Am J Pathol. 177:3089–3099. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shi X: Resident macrophages in the
cochlear blood-labyrinth barrier and their renewal via migration of
bone-marrow-derived cells. Cell Tissue Res. 342:21–30. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang W, Dai M, Fridberger A, Hassan A,
Degagne J, Neng L, Zhang F, He W, Ren T, Trune D, et al:
Perivascular-resident macrophage-like melanocytes in the inner ear
are essential for the integrity of the intrastrial fluid-blood
barrier. Proc Natl Acad Sci USA. 109:pp. 10388–10393. 2012;
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kariya S, Okano M, Maeda Y, Hirai H,
Higaki T, Noyama Y, Haruna T, Nishihira J and Nishizaki K: Role of
macrophage migration inhibitory factor in age-related hearing loss.
Neuroscience. 279:132–138. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Obikane Y: The role of macrophage
migration inhibitory factor (MIF) in cell proliferation signal
transmission. Hokkaido Igaku Zasshi. 79:659–666. 2004.(In
Japanese). PubMed/NCBI
|
|
15
|
Nishihira J: Macrophage migration
inhibitory factor (MIF): Its essential role in the immune system
and cell growth. J Interferon Cytokine Res. 20:751–762. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Keithley EM, Canto C, Zheng QY,
Fischel-Ghodsian N and Johnson KR: Age-related hearing loss and the
ahl locus in mice. Hear Res. 188:21–28. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hunter KP and Willott JF: Aging and the
auditory brainstem response in mice with severe or minimal
presbycusis. Hear Res. 30:207–218. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Neng L, Zhang W, Hassan A, Zemla M,
Kachelmeier A, Fridberger A, Auer M and Shi X: Isolation and
culture of endothelial cells, pericytes and perivascular resident
macrophage-like melanocytes from the young mouse ear. Nat Protoc.
8:709–720. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kaur T, Mukherjea D, Sheehan K, Jajoo S,
Rybak LP and Ramkumar V: Short interfering RNA against STAT1
attenuates cisplatin-induced ototoxicity in the rat by suppressing
inflammation. Cell Death Dis. 2:e1802011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Makary CA, Shin J, Kujawa SG, Liberman MC
and Merchant SN: Age-related primary cochlear neuronal degeneration
in human temporal bones. J Assoc Res Otolaryngol. 12:711–717. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang F, Zhang J, Neng L and Shi X:
Characterization and inflammatory response of perivascular-resident
macrophage-like melanocytes in the vestibular system. J Assoc Res
Otolaryngol. 14:635–643. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Neng L, Zhang J, Yang J, Zhang F, Lopez
IA, Dong M and Shi X: Structural changes in thestrial
blood-labyrinth barrier of aged C57BL/6 mice. Cell Tissue Res.
361:685–696. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dong Y, Li M, Liu P, Song H, Zhao Y and
Shi J: Genes involved in immunity and apoptosis are associated with
human presbycusis based on microarray analysis. Acta Oto-laryngol.
134:601–608. 2014. View Article : Google Scholar
|
|
24
|
Nguyen MT, Lue H, Kleemann R, Thiele M,
Tolle G, Finkelmeier D, Wagner E, Braun A and Bernhagen J: The
cytokine macrophage migration inhibitory factor reduces
pro-oxidative stress-induced apoptosis. J Immunol. 170:3337–3347.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Baumann R, Casaulta C, Simon D, Conus S,
Yousefi S and Simon HU: Macrophage migration inhibitory factor
delays apoptosis in neutrophils by inhibiting the
mitochondria-dependent death pathway. FASEB J. 17:2221–2230. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Saksida T, Stosic-Grujicic S, Timotijevic
G, Sandler S and Stojanovic I: Macrophage migration inhibitory
factor deficiency protects pancreatic islets from palmitic
acid-induced apoptosis. Immunol Cell Biol. 90:688–698. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kariya S, Okano M, Maeda Y, Hirai H,
Higaki T, Noyama Y, Haruna T, Nishihira J and Nishizaki K:
Macrophage migration inhibitory factor deficiency causes prolonged
hearing loss after acoustic overstimulation. Otol & Neurot.
36:1103–1108. 2015. View Article : Google Scholar
|