Introduction
Colon cancer is currently one of the most common
malignant cancers of the gastrointestinal tract. The incidence of
colon cancer has significantly increased in China within recent
years (1). The associated
resistance to cancer chemotherapy, in particular 5-fluorouracil
(5-FU), is an increasing problem within this line of treatment.
Initially presented in 1997, a small population of cells reported
to have the ability to self-renew were identified as cancer stem
cells (CSCs). Such cells pose a great challenge to cancer treatment
due to associated chemoresistance (2).
Metastasis-associated colon cancer 1 (MACC1) was
initially identified as a key regulator of the hepatocyte growth
factor-tyrosine-protein kinase signaling pathway by genome-wide
data analysis; MACC1 was also reported to be associated with colon
cancer metastasis (3–5). Significant upregulation of MACC1
expression has been observed within malignant tissues including
colon cancer of all stages, and liver and lung metastases, compared
with in normal tissues and adenomas (3).
MACC1 promotes carcinogenesis, growth and metastasis
of colorectal cancer (6,7). Furthermore, MACC1 is associated with
the promotion of tumor growth, invasion and metastasis in gastric
cancer (8,9), as well as poor prognosis in solid
cancers (10,11). As an easily detected biomarker in
cancer, MACC1 may serve as a prognostic factor of remission
following liver resection of colorectal cancer metastases (12). To the best of our knowledge,
MACC1-associated regulation of cell chemoresistance, the
development of CSC-like properties and the underlying mechanism
have yet to be investigated.
Numerous mechanisms have been associated with the
development of drug resistance. One of the major causes of
multi-drug resistance (MDR) is the overexpression of the
membrane-bound drug transporter protein P-glycoprotein (P-gp)
(13,14). P-gp is the protein product of the
MDR gene ATP binding cassette subfamily B member 1, and acts as an
energy-dependent drug efflux pump that requires two molecules of
adenosine 5′-triphosphate to remove a number of structurally
unrelated chemotherapeutic drugs.
The present study detected MACC1 expression and
examined its association with 5-FU resistance and CSC-like
properties in colon cancer cells. Increased 5-FU sensitivity,
reduced MDR protein 1 (MDR1) expression levels and reduced CSC-like
properties were associated with MACC1 knockdown, which may be
associated with inhibition of the phosphoinositide 3-kinase/protein
kinase B (PI3K/AKT) signaling pathway.
Materials and methods
Cell lines and reagents
SW620 and HCT116 cell lines were purchased from the
Culture Collection of the Chinese Academy of Sciences (Shanghai,
China). Cells were cultured in RPMI 1640 medium (Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA) supplemented with 10%
fetal bovine serum (PAN Biotech GmbH, Aidenbach, Germany) under a
humidified 5% CO2 atmosphere at 37°C. In addition,
5-FU-resistant (5-FUR) cell lines SW620/5-FUR and HCT116/5-FUR were
developed (15,16). Briefly, SW620/5-FUR and
HCT116/5-FUR cells were established by repeated subcultures in
stepwise-increased concentrations of 5-FU (5, 10 and 20 µg/ml;
Sigma-Aldrich; Merck KGaA, Darmstadt Germany, USA) over 6 months.
In the presence of 20 µg/ml 5-FU, exponential growth of SW620/5-FUR
and HCT116/5-FUR cells was observed. Resistant cell lines were
cultured under continuous 5-FU treatment (20 µg/ml) and were
incubated in drug-free medium for 1 week prior to use. Primary
antibodies against MACC1 (cat. no. ab106579) and MDR1 (cat. no.
ab129450) were purchased from Abcam (Cambridge, UK). Primary
antibodies against cluster of differentiation (CD) 44 (cat. no.
3570S), Nanog (cat. no. 4903) and AKT (cat. no. 4685) were
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Primary antibodies against CD133 (cat. no. 18470-1-AP) and GAPDH
(cat. no. 10494-1-AP) were obtained from Proteintech Group, Inc.
(Chicago, IL, USA). Primary antibodies against phosphorylated AKT
(p-AKT; Ser473; cat. no. 11054-1) were purchased from Signalway
Antibody LLC (College Park, MD, USA). The PI3K inhibitor LY294002
was purchased from Selleck Chemicals (Houston, TX, USA). Cells were
treated with LY294002 (20 µM) for 2 h at 37°C.
Establishment of stably transfected
cell lines
The short hairpin RNA (shRNA) sequence for MACC1
was: 5′-AATTATATGCCAGGACAGCTT-3′. As a negative control (NC), a
scrambled sequence was designed: 5′-AACAGTTATCTATGCGACAGT-3′.
Lenti-Easy Packaging Mix and Lentiviral Vector consist of shRNA
(Shanghai GeneChem Co., Ltd., Shanghai, China) were transfected
into 293T cells. The cell supernatant was collected, concentrated
by centrifugation (4,000 × g, 37°C, 10 min) and purified. The
lentivirus (LV)-MACC1-short hairpin RNA (shRNA)-green fluorescent
protein (GFP) was obtained. The two colonic cancer cell lines were
seeded on a 6-well plate (5×104 cells/well) and cultured
for 24 h. Lentivirus (LV)-MACC1-short hairpin RNA (shRNA)-green
fluorescent protein (GFP) was transduced into cells at a
multiplicity of infection (MOI) of 60 using polybrene (10 µg/ml;
Sigma-Aldrich; Merck KGaA) and Enhanced Infection Solution
(Shanghai GeneChem Co., Ltd., Shanghai, China), named positive
experimental group (shRNA group). Simultaneously, a non-target NC
virus LV-NC-shRNA-GFP was transduced into cells as a control, noted
as the vehicle group. After 12 h, the medium was replaced with
complete culture medium. Cells were cultured for 72 h in completed
medium and observed under a fluorescence microscope for
transduction efficiency. Then stably transduced cells were selected
with 10 µg/ml puromycin. Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) and western blotting were
performed to detect the inhibition rate of LV-mediated shRNA
targeting MACC1.
For MACC1 overexpression, ectopic MACC1 coding
sequence was amplified by PCR. The primer sequences were: Forward
5′-CCGCTCGAGATGCTAATCACTGAAAGAAAAC-3′ and reverse
5′-CCGCTCGAGCTATACTTCCTCAGAAGTGGAGAAT-3′. Lenti-Easy Packaging Mix
and Lentiviral Vector consist of MACC1 plasmid (Shanghai GeneChem
Co., Ltd.,) were transfected into 293T cells. Cell supernatant were
collected, concentrated and purified, Then the (LV)-MACC1-GFP) was
obtained. Colonic cancer cells were seeded on a 6-well plate
(5×104 cells/well) and cultured for 24 h. LV-MACC1-GFP
was transduced into cells in the MAAC1 group at an MOI of 40 using
polybrene (10 µg/ml; Sigma-Aldrich; Merck KGaA) and Enhanced
Infection Solution (Shanghai GeneChem Co., Ltd.). Simultaneously, a
non-target NC virus LV-NC-GFP was transduced into cells in the
vehicle group. After 12 h the medium was replaced with complete
culture medium, and stably transduced cells were selected with 10
µg/ml puromycin.
RT-qPCR
Cells were separated into three groups, including
control group (no tranduction), vehicle group (transduction with
LV-scrambled sequence-GFP) and shRNA group (transduction with
LV-MACC1-shRNA-GFP). Total RNA was extracted from the cells
following treatment for the indicated duration with RNAiso Plus
(cat. no. 9108; Takara Biotechnology Co., Ltd., Dalian, China).
Following genomic DNA elimination reaction and
reverse-transcription reaction using PrimeScript™ RT reagent Kit
with gDNA Eraser (Perfect Real Time; cat. no. RR047A; Takara
Biotechnology Co., Ltd.) in a total volume of 20 µl according to
the manufacturer's protocol. First strand cDNA synthesis was
conducted using 1,000 ng total RNA. Quantification of target and
reference (GAPDH) genes was performed in triplicate using SYBR
Premix Ex Taq II (Tli RNaseH Plus; cat. no. RR820A; Takara
Biotechnology Co., Ltd.) on CFX96 Touch™ Real-Time PCR Detection
system (Bio-Rad Laboratories, Inc., Hercules, CA, USA). PCR
thermocycling conditions is following: initial denaturation at 95°C
30 sec; 2nd step PCR 40 cycles of 95°C for 5 sec, 60°C for 30 sec)
and dissociation. Following normalization to the GAPDH gene,
expression levels for each target gene were calculated using the
2−ΔΔCq method (17).
Primers used in RT-qPCR were as follow: MACC1,
5′-CCTTCGTGGTAATAATGCTTCC-3′ forward and
5′-AGGGCTTCCATTGTATTGAGGT-3′ reverse; CD44 forward,
5′-ATCATCTTGGCATCCCTCTTG-3′ and reverse,
5′-CACCATTTCCTGAGACTTGCTG-3′; CD133 forward,
5′-ACAATCCTGTTATGACAAGCCCA-3′ and reverse,
5′-GGAAAGTCCTTGTAGACCCAGAAA-3′; Nanog forward,
5′-CCTATGCCTGTGATTTGTGGG-3′ and reverse, 5′-TTGCCTTTGGGACTGGTGG-3′;
Bmi-1 forward, 5′-AAGGAGGAGGTGAATGATAA-3′ and reverse,
5′-ATCAATCTGGAAAGTATTAGG-3′; Oct-4 forward,
5′-GTGAGAGGCAACCTGGAGAA-3′ and reverse, 5′-GAACCACACTCGGACCACAT-3′;
GAPDH forward, 5′-ACCACAGTCCATGCCATCCAC-3′ and reverse,
5′-TCCACCACCCTGTTGCTGTA-3′. Each experiment was performed in
triplicate.
Western blotting
Cells were lysed in lysis buffer (50 mM Tris, 150 mM
NaCl, 1% NP-40, 0.1% SDS, 10 mM EDTA, 1 mM PMSF, and 0.5% sodium
deoxycholate) containing phosphatase inhibitor. Following
centrifuging at 13,201 × g at 4°C for 15 min, the supernatants of
lysates were collected for use. The proteins concentration was
quantified by using the bicinchoninic assay protein assay kit
(Beyotime Institute of Biotechnology, Shanghai, China). Lysates
were denatured by heating. Equal amounts of protein samples (20 µg)
were loaded per well and separated by 10% SDS-PAGE, and were
subsequently transferred electrophoretically onto polyvinylidene
fluoride membranes. Following blocking with 5% non-fat milk at room
temperature for 1 h, membranes were incubated with primary
antibodies (all 1:1,000) at 4°C overnight and were then incubated
with peroxidase-conjugated goat anti-rabbit immunoglobulin (Ig;
cat. no. ZB-2301; OriGene Technologies, Inc., Beijing, China) or
peroxidase-conjugated goat anti-mouse IgG (cat. no. ZB-2305;
OriGene Technologies, Inc.; 1:5,000) for 1 h at room temperature.
Specific immune complexes were detected using a chemiluminescence
reagent (EMD Millipore, Billerica, MA, USA) with FUSION-FX7
advanced system (Vilber Lourmat, Marne La Vallée, France). Specific
bands were quantified using FusionCapt Advance software (version
16.08a; Vilber Lourmat). Each experiment was performed in
triplicate.
MTT assay
Cells (5.0×103 cells/well) were seeded in
96-well plates. Following treatment with a series of 5-FU dosages
(5, 10, 20, 40, 80 µg/ml) at 37°C for 48 h, medium was removed and
50 µl MTT (Sigma-Aldrich; Merck KGaA) was added to each well.
Subsequently, cells were incubated at 37°C for 4 h in the dark. The
reaction was terminated by the addition of 150 µl dimethyl
sulfoxide (Sigma-Aldrich; Merck KGaA), and the absorbance of
samples was measured at 570 nm.
Sphere-forming assay
Cells (1×104/well) were cultured in
6-well ultralow attachment surface dishes (Corning Inc., Corning,
NY, USA). The cells were cultured in serum-free Dulbecco's modified
Eagle's medium/F12 (Hyclone; GE Healthcare Life Sciences)
supplemented with 20 ng/ml recombinant human epithelial growth
factor (Peprotech EC Ltd., London, UK), 10 ng/ml human basic
fibroblast growth factor (Peprotech EC Ltd.) and B27 (Gibco; Thermo
Fisher Scientific Inc., Waltham, MA, USA). The number of spheres
>50 µm3 was counted after 14 days of cell culture
with an inverted microscope (CKX41; Olympus Corporation, Tokyo,
Japan).
Statistical analysis
Data are expressed as the mean ± standard deviation
of three independent experiments. Data were analyzed by two-tailed
unpaired Student's t-test between two groups and by one-way
analysis of variance with Tukey's multiple comparison test among
three groups. These analyses were performed using GraphPad Prism
Software Version 6.0 (GraphPad Software, Inc., La Jolla, CA,
USA).
Results
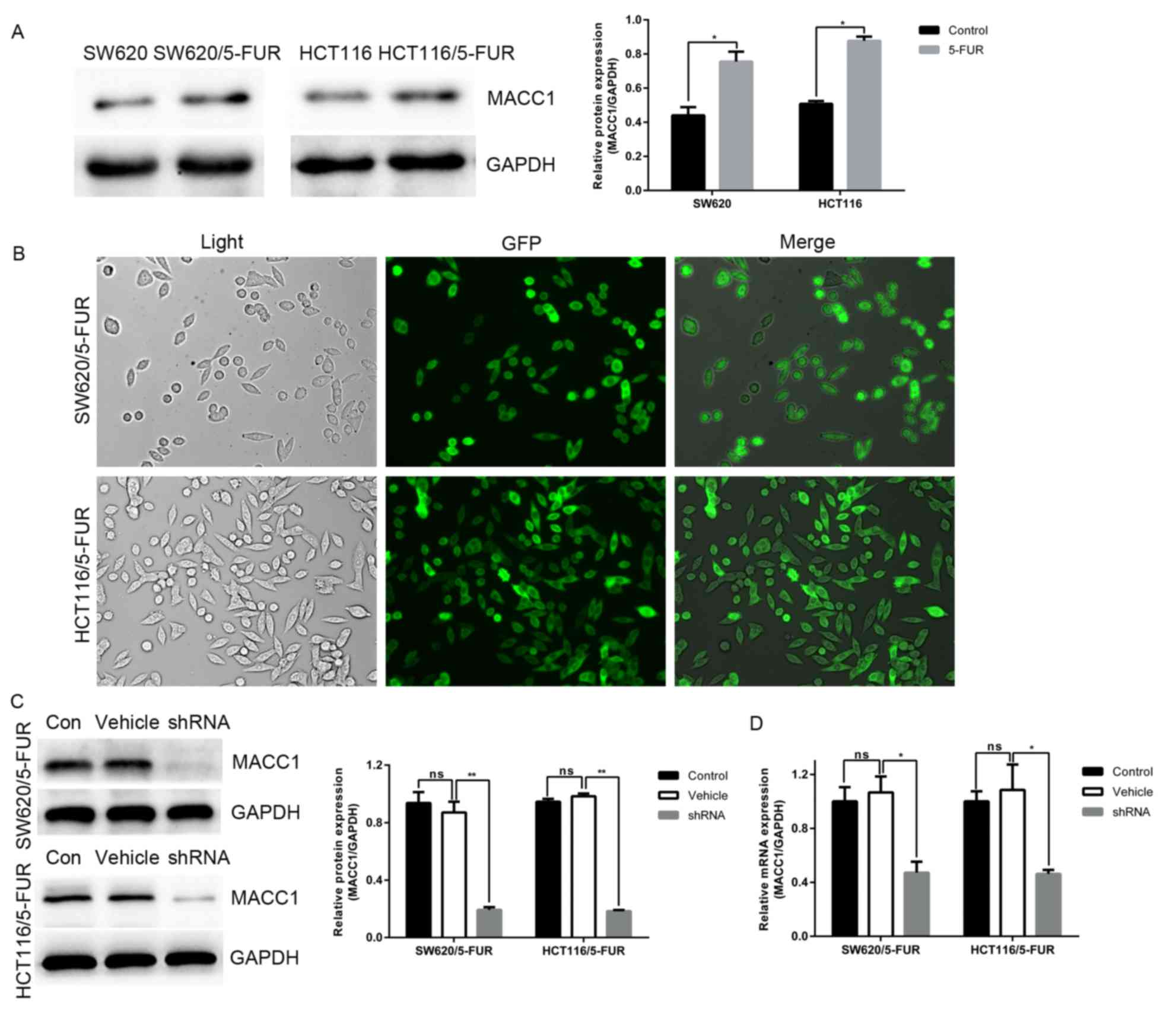
MACC1 expression is increased in 5-FUR
colon cancer cells and is decreased by RNA interference
Human colon cancer cell lines SW620 and HCT116 were
employed to generate SW620/5-FUR and HCT116/5-FUR cell lines via
stepwise exposure to increasing doses of 5-FU. Unexpectedly,
compared with parental cells, the expression of MACC1 was
significantly increased in 5-FUR cells (Fig. 1A). To determine the role of MACC1
in colon cancer cells, LV-MACC1-shRNA-GFP and LV-NC-shRNA-GFP were
transduced into SW620/5-FUR and HCT116/5-FUR colon cancer cells. A
total of 72 h post-transduction, cells with green fluorescence were
observed under fluorescence microscopy (Fig. 1B). Cells were divided into three
groups: Blank control cells (control group), NC cells (vehicle
group) and MACC1-shRNA cells (shRNA group). Expression levels of
MACC1 in transduced cells selected via puromycin were measured by
RT-qPCR and western blot analysis. Compared with the control and
vehicle groups, mRNA and protein expression levels of MACC1 were
significantly downregulated in the shRNA group (Fig. 1C and D). Cells of the vehicle and
shRNA groups were used for further analysis.
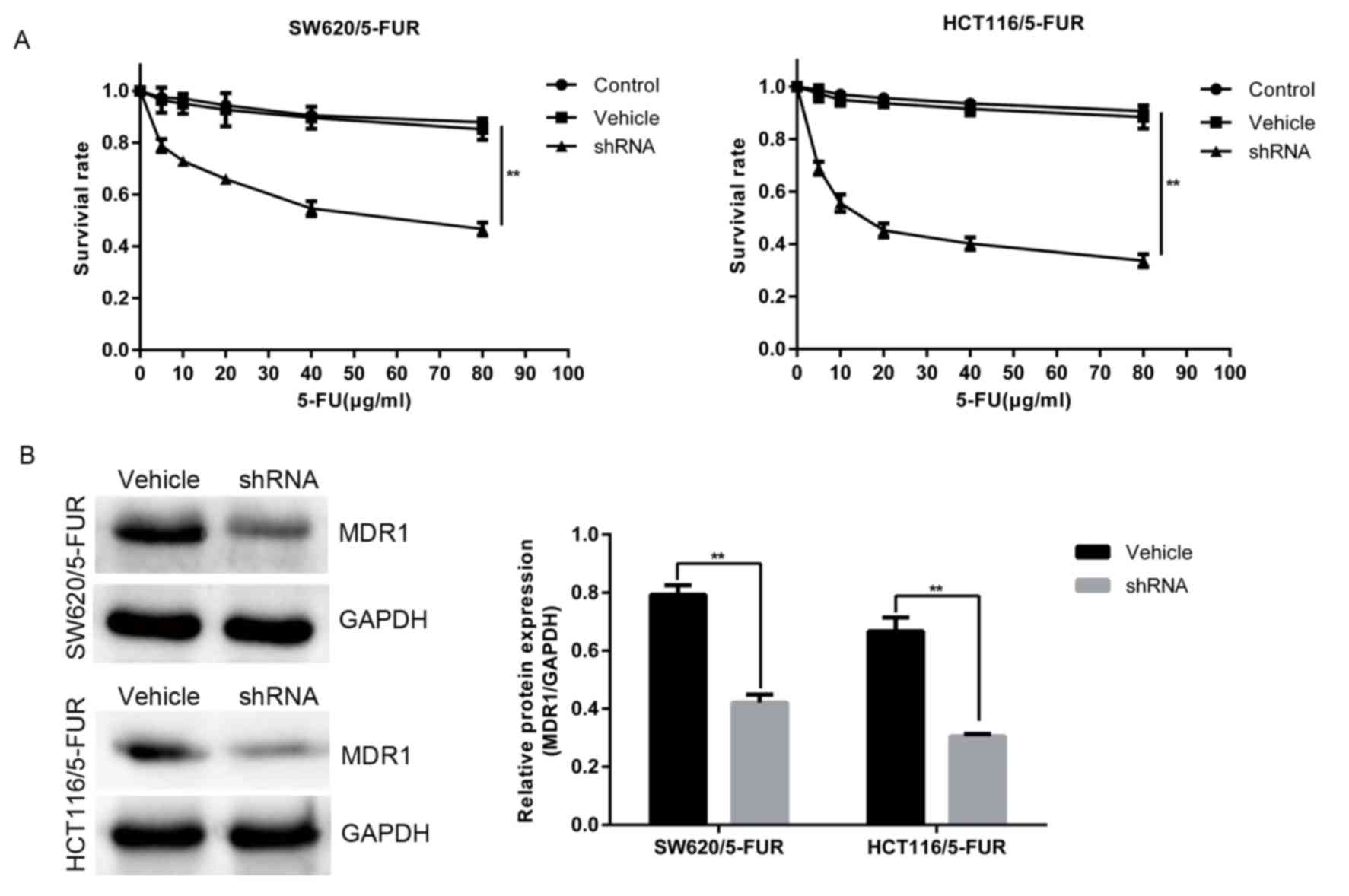
MACC1 knockdown sensitizes colon
cancer cells to 5-FU and reduces MDR1 expression
To determine whether MACC1 promotes resistance to
5-FU in colon cancer cells, survival rate was detected in the
vehicle and shRNA groups treated with a series of 5-FU dosages for
48 h via an MTT assay. Lower rates of survival were noted in the
shRNA group, compared with in the vehicle group (Fig. 2A). Furthermore, MACC1 knockdown
reduced the protein levels of MDR1 (Fig. 2B). Collectively, the results of the
present study revealed that MACC1 promoted chemoresistance and MDR1
upregulation in SW620/5-FUR and HCT116/5-FUR cells.
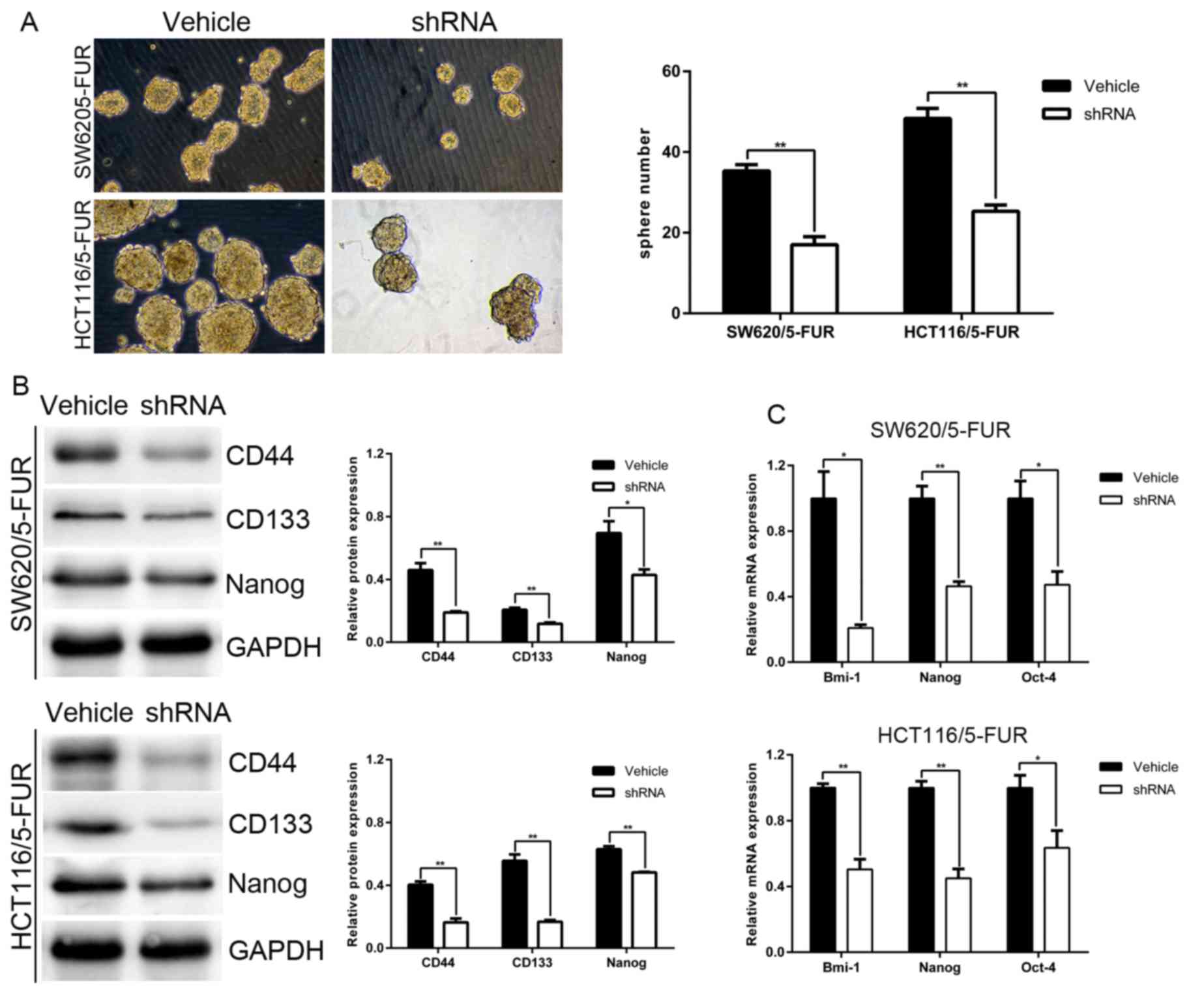
MACC1 induces CSC-like properties of
colon cancer cells
The effects of MACC1 on the CSC-like phenotype were
investigated in SW620/5-FUR and HCT116/5-FUR cells. Cells in the
shRNA group exhibited reduced sphere-forming capacity compared with
cells in the vehicle group. Western blotting and RT-qPCR were
employed to identify the genes responsible for the maintenance and
regulation of CSC-like properties observed in SW620/5-FUR and
HCT116/5-FUR cells. The results of the present study revealed a
significant decrease in the expression levels of the pluripotent
markers CD44, CD133, Bmi-1, Oct-4 and Nanog in the shRNA group
compared with in the vehicle group (Fig. 3B and C). Collectively, these
results suggested that MACC1 knockdown may reduce CSC-like
properties in SW620/5-FUR and HCT116/5-FUR cells.
MACC1 overexpression inhibits 5-FU
sensitivity and promotes CSC-like properties in colon cancer
cells
Western blot analysis was used to detect the
expression levels of MACC1 in stably transduced cells. Compared
with the control and vehicle groups, MACC1 protein levels were
significantly upregulated in MACC1 group cells (Fig. 4A). To validate whether MACC1
overexpression promoted 5-FU resistance and stem cell-like
properties in colon cancer cells, the survival rate was analyzed in
the control, vehicle and MACC1 groups treated with a series of 5-FU
doses over 24 h. The MACC1 group exhibited a higher rate of
survival compared with the control and vehicle groups (Fig. 4B). Furthermore, MACC1
overexpression significantly increased MDR1 expression, and the
expression of pluripotent markers CD44, CD133 and Nanog, compared
with in the vehicle group (Fig.
4C). Collectively, these results demonstrated that MACC1
overexpression reduced 5-FU sensitivity, and induced CD44, CD133
and Nanog expression in SW620 and HCT116 cells.
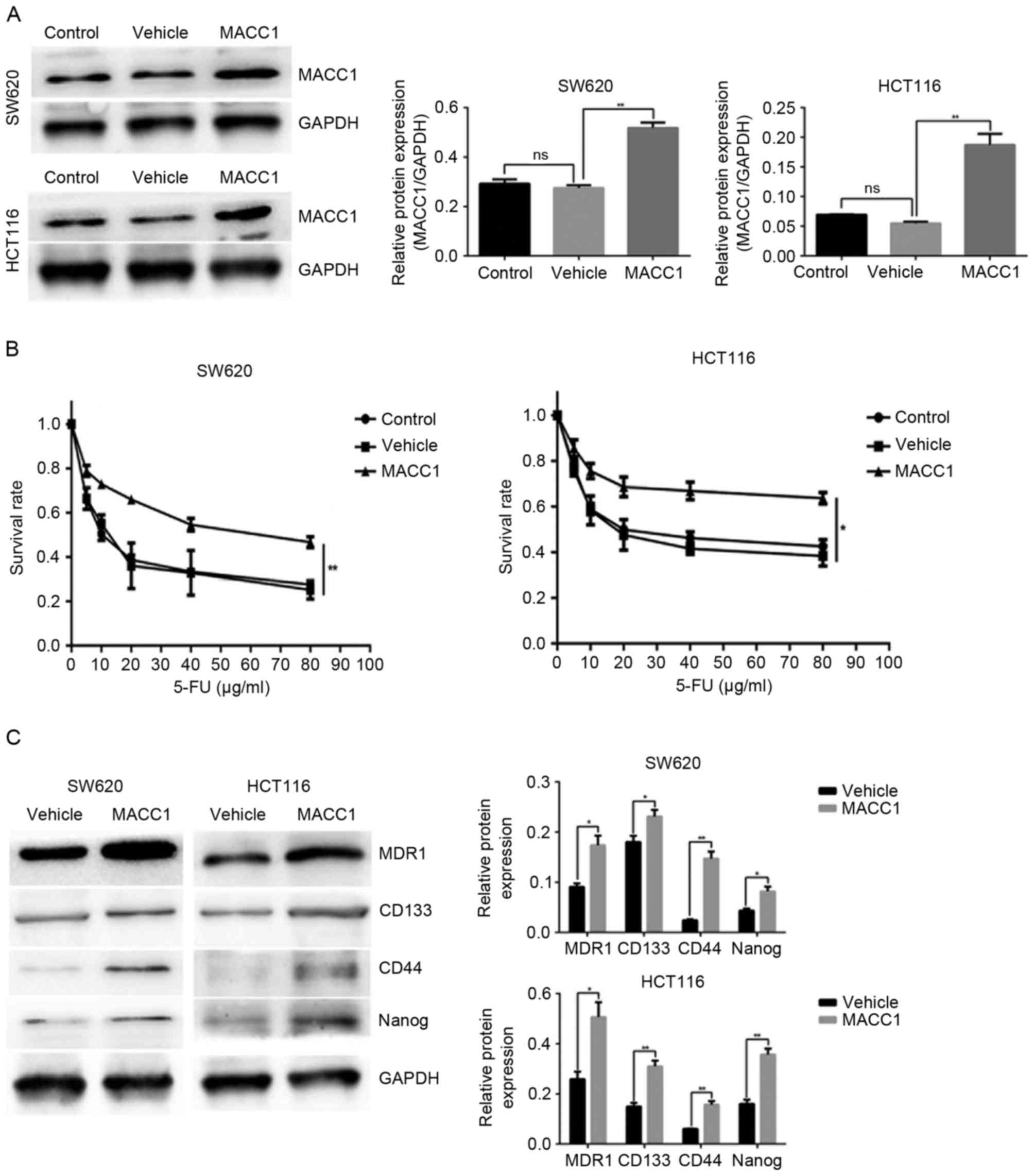 | Figure 4.MACC1 overexpression inhibits 5-FU
sensitivity and promotes cancer stem cell-like properties in colon
cancer cells. Cells transduced with nothing comprised the control
group, cells transduced with LV-negative control-GFP comprised the
vehicle group and cells transduced with LV-MACC1-GFP comprised the
MACC1 group. (A) Effects of MACC1 overexpression were identified by
western blot analysis. Bar graphs demonstrate relative expression
levels of MACC1 protein. **P<0.01 vs. vehicle (B) Cells of the
control, vehicle and MACC1 groups were cultured in the presence of
5-FU at the indicated concentrations for 24 h, cell viability was
determined using an MTT assay. *P<0.05, **P<0.01 vs. the
vehicle (C) Protein levels of MDR1, CD133, Nanog and CD44 in the
vehicle and MACC1 groups were analyzed by western blotting. Bar
graphs indicate the relative protein expression levels. *P<0.05,
**P<0.01 vs. vehicle. 5-FU, 5-fluorouracil; CD, cluster of
differentiation; GFP, green fluorescent protein; LV, lentivirus;
MACC1, metastasis-associated colon cancer 1; MDR1, multi-drug
resistance protein 1; ns, not significant. |
MACC1 promotes chemoresistance and
CSC-like properties via activation of the PI3K/AKT signaling
pathway in colon cancer cells
Recent studies have revealed that AKT serves an
important role in chemoresistance (18). Therefore, the phosphorylation
levels of AKT (Ser473) in the control, vehicle and shRNA groups of
HCT116/5-FUR cells were measured by western blotting. The results
of the present study indicated that MACC1 knockdown decreased the
protein expression levels of p-AKT (Fig. 5A). To determine the possible roles
of PI3K/AKT in MACC1-induced chemoresistance and stem cell
properties, LY294002 was applied; chemoresistance and sphere
forming ability of HCT116/5-FUR cells were investigated. The
results of the present study suggested that LY294002 significantly
decreased 5-FU-associated resistance in HCT116/5-FUR cells
(Fig. 5B). Protein expression
levels of MDR1, CD133 and CD44 were decreased in response of
HCT116/5-FUR cells to LY294002, compared with in the control group
(Fig. 5C). Additionally, treatment
with the PI3K inhibitor significantly reduced the sphere formation
in HCT116/5-FUR cells (Fig. 5D).
These results indicated that MACC1 may promote 5-FU-associated
resistance and CSC-like properties of colon cancer cells via
stimulation of the PI3K/AKT signaling pathway.
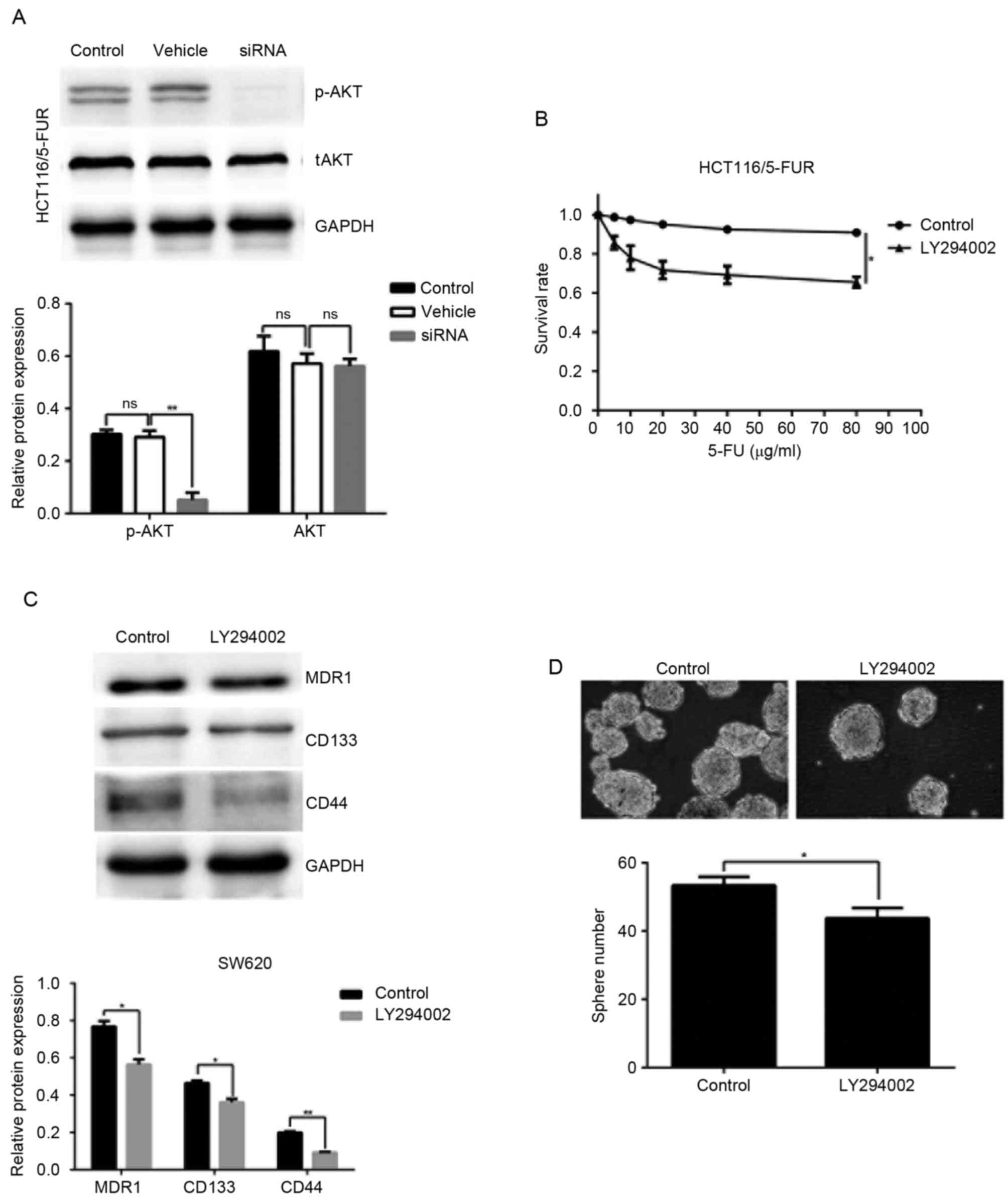 | Figure 5.MACC1 promotes chemoresistance and
cancer stem cell-like properties through the phosphoinositide
3-kinase/AKT pathway in 5FUR colon cancer cells. (A) Representative
western blot image demonstrates the expression of tAKT and p-AKT in
the control, vehicle and shRNA groups. Bar graphs indicate the
relative protein expression levels. (B) Control group and LY294002
(10 µM) group cells were cultured in the presence of 5-FU at the
indicated concentrations for 24 h, and the cell viability was
determined using an MTT assay. (C) Protein levels of MDR1, CD133
and CD44 in the control and LY294002 (10 µM) groups were analyzed
by western blotting. (D) Sphere forming ability was evaluated in
the control and LY294002 (10 µM) group cells (magnification, ×200).
Bar graph demonstrate the relative sphere numbers. *P<0.05,
**P<0.01. 5-FU, 5-fluorouracil; CD, cluster of differentiation;
AKT, protein kinase B; MACC1, MACC1, metastasis-associated colon
cancer 1; MDR1, multi-drug resistance protein 1; ns, not
significant; p-AKT, phosphorylated protein kinase B; shRNA, short
hairpin RNA; tAKT, total protein kinase B. |
Discussion
The present study demonstrated that MACC1 may induce
the development of 5-FU resistance and CSC-like properties in colon
cancer cells. The results indicated that MACC1 was upregulated in
5-FUR colon cancer cells, whereas MACC1 knockdown promoted 5-FU
sensitivity and reduced MDR1 protein expression. In addition to
decreased sphere formation, MACC1 knockdown also reduced the
expression levels of pluripotent markers CD44, CD133 and Nanog.
Furthermore, the results of the present study suggested that the
PI3K/AKT signaling pathway was involved in the regulatory effects
of MACC1 in 5-FUR cells. To the best of our knowledge, the present
study is the first to reveal that MACC1 promotes chemoresistance
and CSC-like properties in colon cancer.
Patients with colon cancer that metastasizes to the
liver are treated with conventional first-line treatments.
Combinations of 5-FU, leucovorin and oxaliplatin or irinotecan are
employed and have demonstrated efficiency in phase III trials
(19). However, chemoresistance
develops in almost all patients (2). Therefore, an improved understanding
of drug-associated mechanisms of resistance is essential in
exploring approaches for the prevention or reversal of drug
resistance. MDR is a phenomenon that often accompanies drug
treatment and is characterized by relapse or attenuation of drug
efficacy; MDR is almost unavoidable in patients with colorectal
cancer (CRC) receiving 5-FU-based chemotherapy. The MDR1 phenotype
involves the development of resistance to numerous
chemotherapeutics observed within cancer cells (20); the mechanisms responsible for MDR1
upregulation in CRC cells remain to be elucidated. In the present
study, MACC1 knockdown promoted sensitivity to 5-FU and inhibited
MDR1 protein expression in SW620/5-FUR and HCT116/5-FUR cells.
MACC1 may therefore facilitate the development of chemoresistance
in colon cancer.
CSCs are characterized by three unique properties:
The capacity of self-renewal, the ability to exhibit all
characteristics of the cancer cells of the parental tumor, and the
expression of a distinctive set of surface biomarkers (21). Antitumor therapies may lead to a
reduction of tumor mass yet fail to result in tumor regression due
to the lack of CSC targeting. In the present study, MACC1 knockdown
reduced sphere formation and the expression of pluripotent markers,
including CD44, CD133 and Nanog; MACC1 may serve as a potential
target for the elimination of CSCs in colon cancer. Further
research is required to determine the role of MACC1 in CSC-like
properties and to understand the underlying mechanisms.
Recent studies revealed that PI3K/AKT serves an
important role in chemoresistance (18, 22). Activation of the
PI3K/AKT pathway contributes to numerous cancer therapy-associated
resistance and is deemed a poor prognostic factor of cancer
(23). The results of the present
study suggested that the protein expression levels of p-AKT were
lower, indicating reduced activation of PI3K/AKT, in the
MACC1-depleted colon cancer cells. Subsequent treatment of
HCT116/5-FUR cells with LY294002 led to a decrease in cell survival
rate and sphere formation compared with in untreated HCT116/5-FUR
cells. These results indicated that the PI3K/AKT signaling pathway
may serve a key role in 5-FU resistance and CSC-like properties via
MACC1; however, this process requires further investigation.
References
|
1
|
Chen W, Zheng R, Zeng H, Zhang S and He J:
Annual report on status of cancer in China, 2011. Chin J Cancer
Res. 27:2–12. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dallas NA, Xia L, Fan F, Gray MJ, Gaur P,
van Buren G II, Samuel S, Kim MP, Lim SJ and Ellis LM:
Chemoresistant colorectal cancer cells, the cancer stem cell
phenotype, and increased sensitivity to insulin-like growth
factor-I receptor inhibition. Cancer Res. 69:1951–1957. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stein U, Walther W, Arlt F, Schwabe H,
Smith J, Fichtner I, Birchmeier W and Schlag PM: MACC1, a newly
identified key regulator of HGF-MET signaling, predicts colon
cancer metastasis. Nat Med. 15:59–67. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Stein U, Smith J, Walther W and Arlt F:
MACC1 controls Met: What a difference an Sp1 site makes. Cell
Cycle. 8:2467–2469. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stein U, Burock S, Herrmann P, Wendler I,
Niederstrasser M, Wernecke KD and Schlag PM: Circulating MACC1
transcripts in colorectal cancer patient plasma predict metastasis
and prognosis. PLoS One. 7:e492492012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhen T, Dai S, Li H, Yang Y, Kang L, Shi
H, Zhang F, Yang D, Cai S, He Y, et al: MACC1 promotes
carcinogenesis of colorectal cancer via β-catenin signaling
pathway. Oncotarget. 5:3756–3769. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tang J, Chen J, Chen L, Tang J, Cui Z, Liu
C and Wang Z: Metastasis associated in colon cancer 1 (MACC1)
promotes growth and metastasis processes of colon cancer cells. Eur
Rev Med Pharmacol Sci. 20:2825–2834. 2016.PubMed/NCBI
|
|
8
|
Xie QP, Xiang C, Wang G, Lei KF and Wang
Y: MACC1 upregulation promotes gastric cancer tumor cell metastasis
and predicts a poor prognosis. J Zhejiang Univ Sci B. 17:361–366.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang T, He W, Cui F, Xia J, Zhou R, Wu Z,
Zhao Y and Shi M: MACC1 mediates acetylcholine-induced invasion and
migration by human gastric cancer cells. Oncotarget. 7:18085–18094.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang G, Fu Z and Li D: MACC1
overexpression and survival in solid tumors: A meta-analysis. Tumor
Biol. 36:1055–1065. 2015. View Article : Google Scholar
|
|
11
|
Stein U: MACC1-a novel target for solid
cancers. Expert Opin Ther Targets. 17:1039–1052. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Isella C, Mellano A, Galimi F, Petti C,
Capussotti L, De Simone M, Bertotti A, Medico E and Muratore A:
MACC1 mRNA levels predict cancer recurrence after resection of
colorectal cancer liver metastases. Ann Surg. 257:1089–1095. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang X, Yang P, Shen J, Osaka E, Choy E,
Cote G, Harmon D, Zhang Z, Mankin H, Hornicek FJ and Duan Z:
Prevention of multidrug resistance (MDR) in osteosarcoma by
NSC23925. Br J Cancer. 110:2896–2904. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gottesman MM, Fojo T and Bates SE:
Multidrug resistance in cancer: Role of ATP-dependent transporters.
Nat Rev Cancer. 2:48–58. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Toden S, Okugawa Y, Jascur T, Wodarz D,
Komarova NL, Buhrmann C, Shakibaei M, Boland CR and Goel A:
Curcumin mediates chemosensitization to 5-fluorouracil through
miRNA-induced suppression of epithelialto-mesenchymal transition in
chemoresistant colorectal cancer. Carcinogenesis. 36:355–367. 2014.
View Article : Google Scholar
|
|
16
|
Toden S, Tran HM, Tovar-Camargo OA,
Okugawa Y and Goel A: Epigallocatechin-3-gallate targets cancer
stem-like cells and enhances 5-fluorouracil chemosensitivity in
colorectal cancer. Oncotarget. 7:16158–16171. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Das D, Satapathy SR, Siddharth S, Nayak A
and Kundu CN: NECTIN-4 increased the 5-FU resistance in colon
cancer cells by inducing the PI3K-AKT cascade. Cancer Chemother
Pharmacol. 76:471–479. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lenz HJ: First-line combination treatment
of colorectal cancer with hepatic metastases: Choosing a targeted
agent. Cancer Treat Rev. 34 Suppl 2:S3–S7. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Amiri-Kordestani L, Basseville A, Kurdziel
K, Fojo AT and Bates SE: Targeting MDR in breast and lung cancer:
Discriminating its potential importance from the failure of drug
resistance reversal studies. Drug Resist Updat. 15:50–61. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Maenhaut C, Dumont JE, Roger PP and van
Staveren WC: Cancer stem cells: A reality, a myth, a fuzzy concept
or a misnomer? An analysis. Carcinogenesis. 31:149–58. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao M, Luo R, Liu Y, Gao L, Fu Z, Fu Q,
Luo X, Chen Y, Deng X, Liang Z, et al: miR-3188 regulates
nasopharyngeal carcinoma proliferation and chemosensitivity through
a FOXO1-modulated positive feedback loop with
mTOR-p-PI3K/AKT-c-JUN. Nat Commun. 7:113092016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
LoPiccolo J, Blumenthal GM, Bernstein WB
and Dennis PA: Targeting the PI3K/Akt/mTOR pathway: Effective
combinations and clinical considerations. Drug Resist Updat.
11:32–50. 2008. View Article : Google Scholar : PubMed/NCBI
|