Introduction
Chronic myeloid leukemia (CML) is a type of
hematopoietic disorder arising from chromosomal abnormality between
chromosomes 9 and 22 (1). This
chromosomal abnormality, known as the Philadelphia chromosome,
leads to myeloid proliferation (2). Although tyrosine kinase inhibitor
(TKI) imatinib mesylate (Gleevec) has been demonstrated to possess
clinical benefit for patients with chronic phase CML harboring
wild-type BCR-ABL, the CML patients harboring BCR-ABL mutants
(e.g., E255V, Y253H, M351T and T315I) do not respond to imatinib
(3). The majority of imatinib
resistance is caused by BCR-ABL mutants, however other mutants can
be overcome by the second-generation TKIs including dasatinib,
nilotinib and bosutinib (4). For
T315I mutation resistance to imatinib, effective therapeutics
remain to be developed. The third-generation TKIs, such as
ponatinib, show activity against CML with T315I BCR-ABL (5), however, the Food and Drug
Administration (FDA) suspended its clinical use in 2013 due to its
cardiovascular toxicity (6). Thus,
agents effective against T315I BCR-ABL mutation would be valuable
in treating refractory CML patients.
Pyrvinium
(6-(dimethylamino)-2-[2-(2,5-dimethyl-1-phenylpyrrol-3-yl)ethenyl]-1-methyl-quinolinium)
pamoate (Fig. 1A) is a
FDA-approved drug used for the treatment of pinworm infections
(7). Previous studies have
demonstrated that pyrvinium pamoate can effectively inhibit the
growth of tumor cells of diverse tissue origins, including the
pancreas, colon, breast, brain, myeloma and erythroleukemia
(8–14). It also has been demonstrated that
pyrvinium pamoate exhibits increased cytotoxicity in tumor cells
under hypoglycemic conditions and enhances the effects of
conventional chemotherapeutic drugs in tumor xenografts (10,12,15).
A previous study reported that pyrvinium pamoate was active against
CML patients harboring wild-type BCR-ABL (16). However, whether it is against T315I
BCR-ABL CML cells remains to be elucidated.
In the present study, the activity of pyrvinium
pamoate was determined in CML cells with T315I BCR-ABL. It was
identified that pyrvinium pamoate potently suppressed proliferation
and induced apoptosis, and inhibited the Wnt/β-catenin signaling
pathway in the CML cells with T315I BCR-ABL. These results suggest
that pyrvinium pamoate may be an effective drug for the treatment
of CML with T315I BCR-ABL mutation.
Materials and methods
Reagents and antibodies
Pyrvinium pamoate (CAS: 3546-41-6; chemical
structure, Fig. 1A) was purchased
from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany) and was
prepared as a 20 mM stock solution in dimethyl sulfoxide (DMSO) and
stored at −20°C. Antibodies and their sources were as follows:
Mouse monoclonal antibodies against poly ADP-ribose polymerase
(PARP; clone 4C10-5; cat no. 51-6639GR; dilution, 1:4,000),
X-linked inhibitor of apoptosis protein (XIAP; cat no. 610762;
dilution, 1:1,000), c-Myc (cat no. 51-1485GR; dilution, 1:1,000),
β-catenin (cat no. 610154; dilution, 1:1,000), cytochrome c
(clone 6H2.B4; cat no. 556432; dilution, 1:1,000), caspase-3 (cat
no. 610322; dilution, 1:1,000) and rabbit polyclonal antibodies
against active caspase-3 (cat no. 557038; dilution, 1:500) were
from BD Pharmingen (San Diego, CA, USA). Antibodies against B-cell
lymphoma (Bcl)-2-associated X (Bax; cat no. sc-493; dilution,
1:1,000), Bcl-XL (S-18) (cat no. sc-634; dilution,
1:1,000), glycogen synthase kinase (GSK)3β (cat no. sc-9166;
dilution, 1:1,000), cyclin D1 (cat no. sc-753; dilution, 1:1,000)
and mouse monoclonal antibodies against apoptosis-inducing factor
(AIF; cat no. sc-55519; dilution, 1:500), were from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA); antibodies against β-actin
(clone AC-15; cat no. A5441; dilution, 1:4,000) and α-tubulin
(clone B-5-1-2; cat no. T5168; dilution, 1:1,000) were from
Sigma-Aldrich; Merck KGaA; antibodies against caspase-8 (cat no.
9746S; dilution, 1:1,000), caspase-9 (cat no. 9508S; dilution,
1:1,000), phosphorylated (p)-GSK3β (Ser9; cat no. 9336S; dilution,
1:1,000) and Mcl-1 (cat no. 4572S; dilution, 1:500) were from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Anti-survivin (cat
no. NB 500-201; dilution, 1:1,000) was from Novus Biologicals,
Bcl-2 (cat no. 05-729; dilution, 1:1,000) was from EMD Millipore
(Billerica, MA, USA) and cytochrome c oxidase subunit II
(COX II; dilution, 1:1,000) was purchased from Invitrogen (Thermo
Fisher Scientific, Inc., Waltham, MA, USA). The secondary Alexa
Fluor® 680CW goat anti-mouse (cat no. 926-68070;
dilution, 1:10,000) and 800CW goat anti-rabbit (cat no. 926-32211;
dilution, 1:10,000) antibodies were purchased from LI-COR
Biosciences (Lincoln, NE, USA).
Cell culture
K562 cells were purchased from American Type Culture
Collection (Manassas, VA, USA), and grown in RPMI-1640 (Invitrogen;
Thermo Fisher Scientific, Inc.) with 10% heat inactivated fetal
bovine serum (FBS; Biological Industries USA, Cromwell, CT, USA),
maintained in a humidified atmosphere with 5% CO2 at
37°C. KBM5 and KBM5-T315I cells were provided by Professor
Sai-Ching J. Yeung (Department of Emergency Medicine, University of
Texas MD Anderson Cancer Center, Houston, TX, USA) and grown in
Iscove's modified Dulbecco's medium (Invitrogen; Thermo Fisher
Scientific, Inc.) supplemented with 10% heat inactivated FBS as
previously described (17). The
32D myeloid cells stably expressing either 210 kDa wild-type
BCR-ABL (32D-BCR-ABL) or T315I BCR-ABL [32D-T315I; both provided by
Dr Lin Qiu (Harbin Institute of Hematology and Oncology, Harbin,
Heilongjiang, China)] were established and maintained in RPMI-1640
with 10% FBS as described previously (18,19).
Cell viability assay
Cell viability was determined by MTS assay
(CellTiter 96 Aqueous One Solution Cell Proliferation assay;
Promega Corporation, Madison, WI, USA) (20). Briefly, 2×104 cells in
100 µl were exposed to various concentrations of pyrvinium pamoate
for 72 h. Control cells were treated with medium containing
<0.1% DMSO. The absorbance/optical density were read on a
96-well plate reader at wave length 490 nm. The drug concentration
resulting in 50% inhibition of cell growth (IC50) was
determined (21).
Colony formation assays
The K562, KBM5, KBM5-T315I, 32D-BCR-ABL and
32D-T315I cells were treated with pyrvinium pamoate or DMSO
(control) for 24 h, then cells were harvested and washed with PBS,
and 2,000 cells were seeded in RPMI-1640 or Iscove's medium
containing 0.3% agar and 20% FBS in the absence of drug treatment.
Following incubation for another 10~14 days at 37°C and 5%
CO2, colonies with >50 cells were counted using an
inverted phase-contrast microscope (22).
Trypan blue exclusion assay
The K562, KBM5, KBM5-T315I, 32D-BCR-ABL and
32D-T315I cells were cultured at 2×105 cells/well in
6-well plates, then cells were treated with increasing
concentrations of pyrvinium pamoate, or with the fixed
concentration for different time periods. Viable and dead cells
were assessed by counting with a hemocytometer following trypan
blue exclusion assay (23).
Preparation of cell lysates and
cytosolic fractions
Whole lysates were prepared with
radioimmunoprecipitation assay buffer (1X PBS, 1% NP-40, 0.5%
sodium deoxycholate, 0.1% SDS) supplemented with freshly added 10
mM β-glycerophosphate, 1 mM sodium orthovanadate, 10 mM NaF, 1 mM
phenylmethylsulfonyl fluoride, and Roche Complete Mini Protease
Inhibitor Cocktail (Roche Diagnostics, Basel, Switzerland)
(24). The cytosolic fraction was
prepared with digitonin extraction buffer to detect the level of
cytochrome c and AIF in the cytosol, as described previously
(25).
Preparation of cytoplasmic and nuclear
fractions
Control or drug-treated cells were pelleted by
centrifugation at 300 × g for 5 min at room temperature and rinsed
with PBS. Pellets were then resuspended in 200 µl ice-cold lysis
buffer (10 mM Hepes pH 7.9, 10 mM KCl, 0.1 mM EDTA, 0.4% NP-40 with
1 mM DTT, 0.5 mM PMSF, 1 mM sodium orthovanadate and the Roche
Complete Mini Protease Inhibitor Cocktail) by pipetting up and down
(without bubbling) ~10 times (22,26).
Following incubation on ice for 10 min, the lysates were
centrifuged at 10,000 × g for 10 min at 4°C. The supernatants were
transferred to fresh tubes and referred to as cytoplasm ice
extracts. Following washing with the lysis buffer, the pellets were
resuspended in nuclear protein extraction buffer with inhibitors
(20 mM HEPES, pH 7.9, 0.4 M NaCl, 1 mM EDTA with 1 mM DTT, 0.5 mM
PMSF, 0.2 mM sodium orthovanadate and the Roche Complete Mini
Protease Inhibitor Cocktail) and centrifuged at 10,000 × g for 10
min at 4°C. The resultant supernatants were kept as nuclear
fractions (22,26).
Western blotting analysis
The whole cells were lysed in
radioimmunoprecipitation assay buffer on ice for 30 min. Equal
volumes (20 µl) of lysate were electrophoresed with 10%
SDS-polyacrylamide gels and electrotransferred to nitrocellulose
membranes. The membranes were blocked in PBS with 0.05% Tween-20
containing 5% skimmed milk and incubated with the primary antibody
at 4°C overnight, and then incubated with the secondary 680/800
antibodies. Protein expression was assessed by the Odyssey infrared
imaging system (LI-COR Biosciences). β-actin was used as a protein
loading control.
Statistical analysis
All experiments were performed a minimum of three
times, and data are presented as the mean ± standard deviation.
Differences between multiple groups were analyzed by one-way
analysis of variance, with post-hoc intergroup comparisons made
using Tukey's test, unless otherwise stated. GraphPad Prism 5.0
software (GraphPad Software, Inc., San Diego, CA, USA) was used for
statistical analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
Pyrvinium pamoate effectively
inhibited the proliferation of CML cells harboring T315I
BCR-ABL
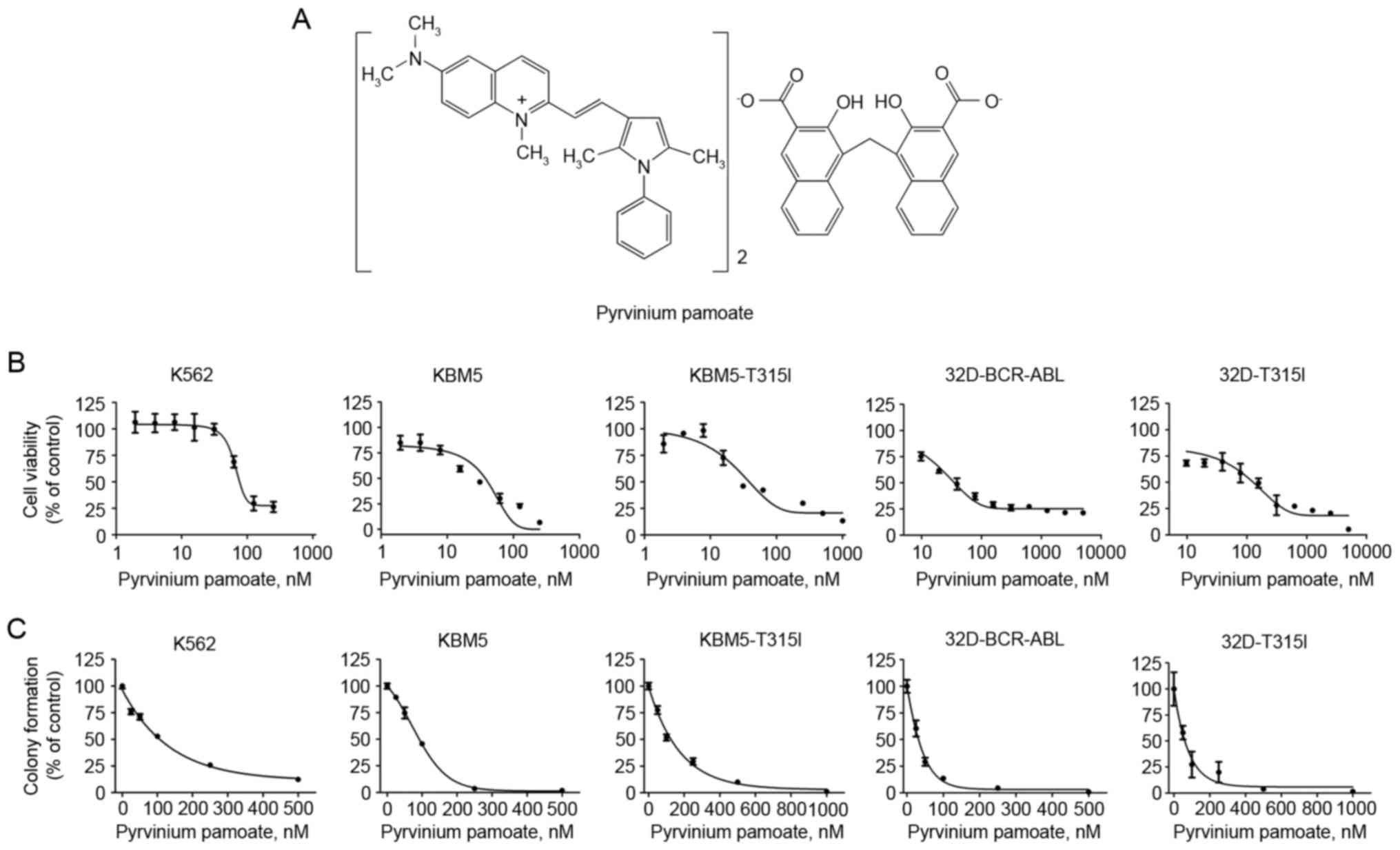
Firstly, the effect of pyrvinium pamoate on cell
viability was examined. A total of three lines of human CML cells
were incubated with increasing concentrations of pyrvinium pamoate
(Fig. 1A) for 72 h. The MTS assay
indicated that K562 and KBM5 cells, both of which harbor wild-type
BCR-ABL, were sensitive to pyrvinium pamoate with IC50
values of 14.2 and 26.2 nM, respectively (Fig. 1B), KBM5-T315I cells harboring T315I
BCR-ABL exhibited a similar sensitivity to pyrvinium pamoate with
an IC50 value 29.2 nM. The ability of pyrvinium pamoate
to inhibit the viability was further confirmed in a pair of murine
32D myeloid cells stably transfected with either the wild-type or
T315I BCR-ABL, the IC50 values were 49.7 and 133 nM,
respectively (Fig. 1B). In an
additional independent set of experiments, similar results were
obtained in colony formation experiments, which indicated that
pyrvinium pamoate inhibited the clonogenicity of CML cells bearing
either wild-type BCR-ABL or T315I BCR-ABL (Fig. 1C).
These results demonstrated that pyrvinium pamoate
could be equally effective in inhibiting the proliferation of CML
cells with T315I BCR-ABL mutation as to that of wild-type CML
cells.
Pyrvinium pamoate induced the
apoptosis in CML cells harboring T315I BCR-ABL
Whether pyrvinium pamoate was able to induce cell
death was investigated. The cells were cultured with increasing
concentrations of pyrvinium pamoate for 36 h, and cell death was
measured by trypan blue exclusion assay. Pyrvinium pamoate led to
marked increases in cell death in a dose- and time-dependent manner
in the CML cells harboring either wild-type or T315I BCR-ABL
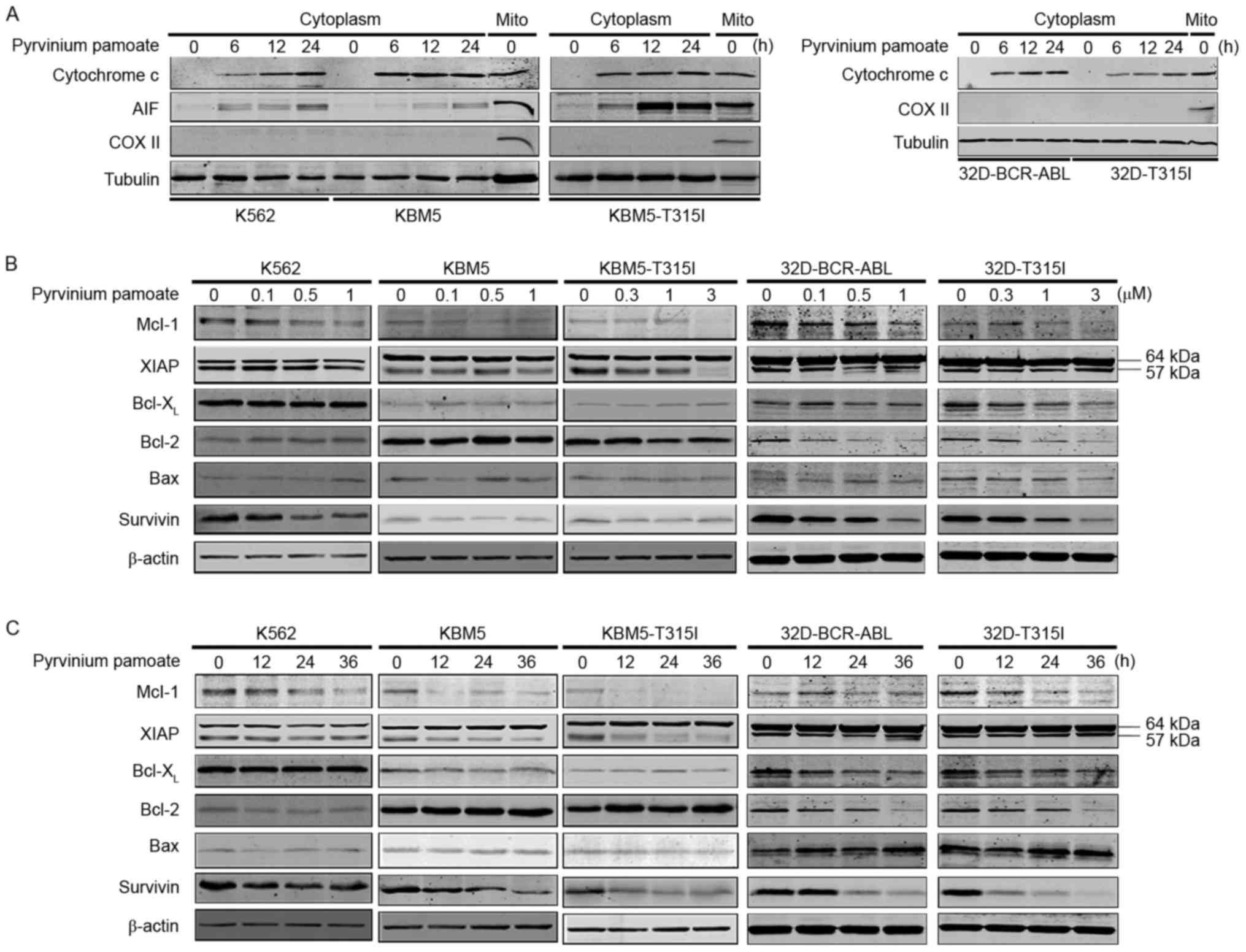
(Fig. 2A and B). In addition,
pyrvinium pamoate induced a dose- and time-dependent cleavage of
PARP and caspase-3 in all five lines of BCR-ABL-expressing cells
(Fig. 2C and D). The levels of
caspase-9 and −8 were also decreased in a dose- and time-dependent
manner, suggesting their activation by pyrvinium pamoate (Fig. 2C and D). Pyrvinium pamoate
additionally induced a time-dependent release of AIF and cytochrome
c into cytosol (Fig. 3A).
Additionally, in all the drug-treated BCR-ABL-expressing cells, a
decrease in the protein levels of anti-apoptotic proteins Mcl-1,
survivin and XIAP was observed (Fig.
3B and C) but not of other Bcl-2 family members. Taken
together, pyrvinium pamoate effectively induced apoptosis in five
lines of BCR-ABL-expressing cells regardless of their BCR-ABL
status.
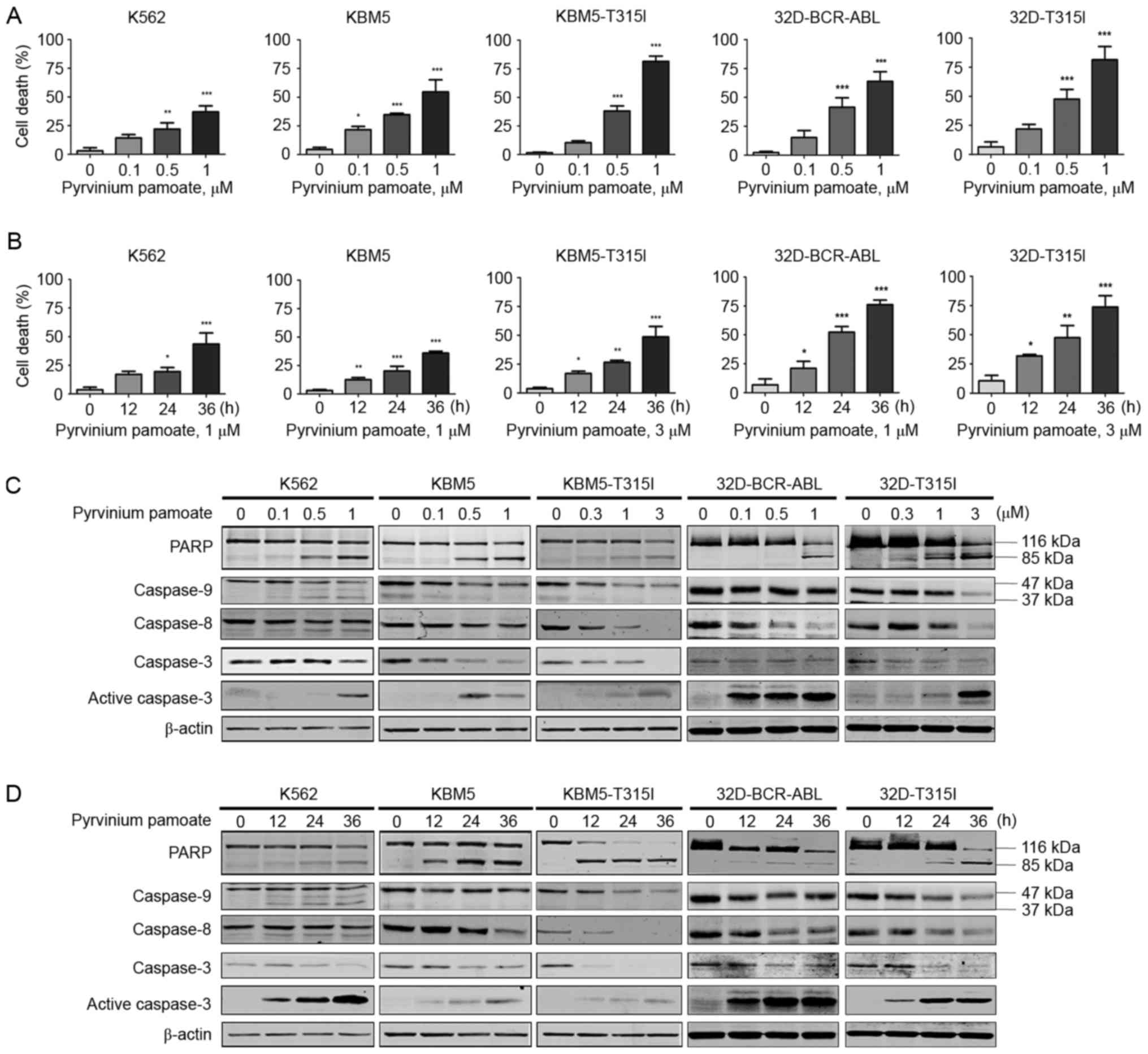 | Figure 2.Pyrvinium pamoate induced apoptosis in
CML cells with T315I BCR-ABL. (A and B) Cell death was evaluated by
trypan blue exclusion assay. (A) K562, KBM5 and 32D-BCR-ABL cells
were treated with pyrvinium pamoate (0, 0.1, 0.5 and 1 µM) for 36
h, KBM5-T315I and 32D-T315I cells were treated with pyrvinium
pamoate (0, 0.3, 1 and 3 µM) for 36 h. (B) K562, KBM5 and
32D-BCR-ABL cells were treated with 1 µM pyrvinium pamoate for 0,
12, 24 and 36 h; KBM5-T315I and 32D-T315I cells were treated with 3
µM pyrvinium pamoate for 0, 12, 24 and 36 h. Trypan blue exclusion
assay was used to measure the cell death of the five cell lines.
*P<0.05, **P<0.01, ***P<0.001, one-way analysis of
variance with post hoc intergroup comparison by the Tukey's test.
(C and D) Dose- and time-course western blotting analysis of PARP
cleavage, caspase-3, −8, and −9 in the whole-cell lysates of CML
cells were shown. β-actin was used as a loading control. CML,
chronic myeloid leukemia; PARP, poly adenosine diphosphate-ribose
polymerase. |
Pyrvinium pamoate effectively
inhibited the Wnt/β-catenin signaling in CML cells harboring T315I
BCR-ABL
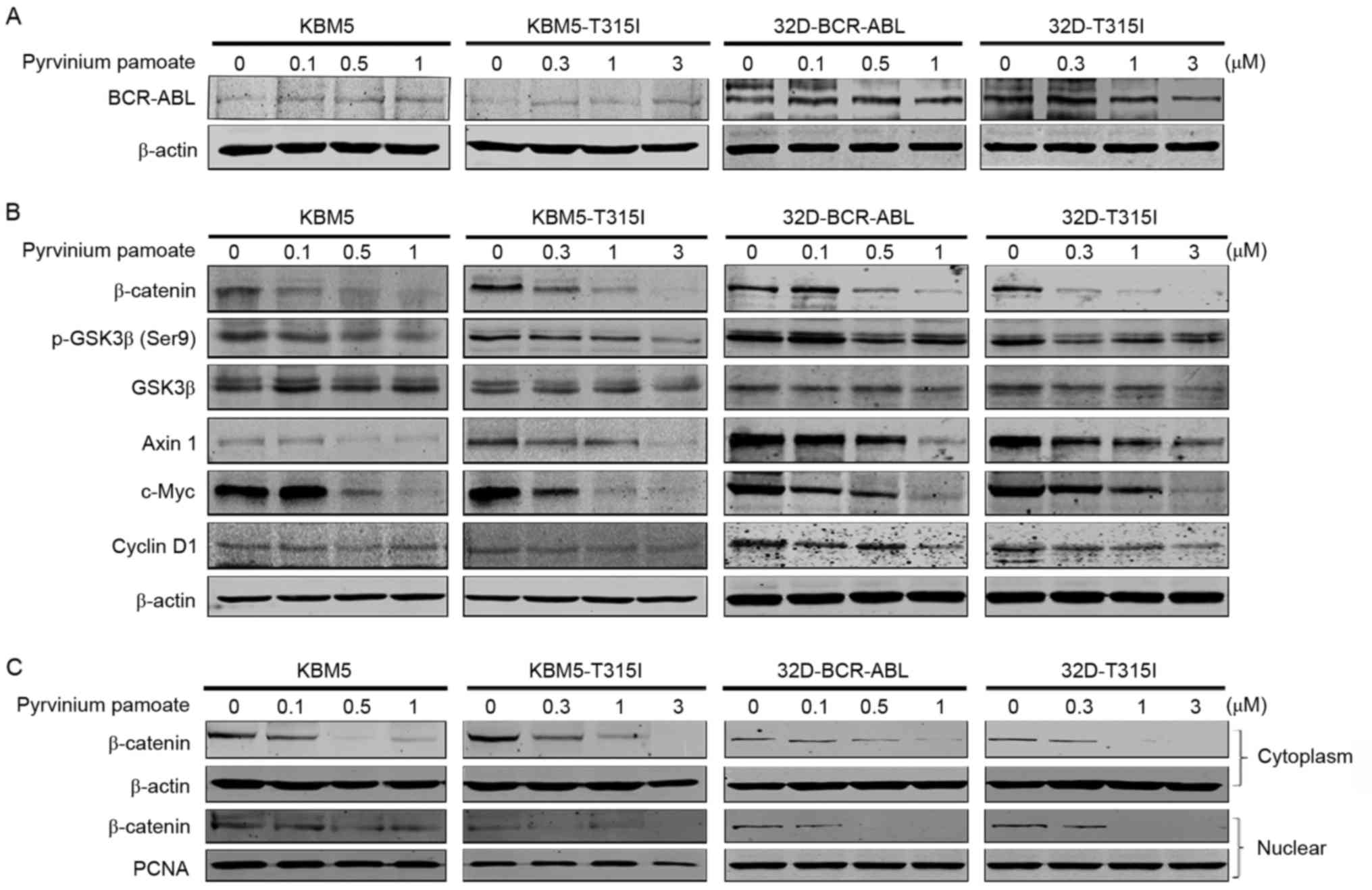
It was investigated whether pyrvinium pamoate
affected BCR-ABL expression, and western blotting analysis
indicated no change in BCR-ABL expression in the CML cells treated
with pyrvinium pamoate (Fig. 4A),
suggesting that the antineoplastic effect of pyrvinium pamoate on
CML cells is BCR-ABL independent. Previous reports have
demonstrated that pyrvinium pamoate inhibits Wnt/β-catenin
signaling pathway in breast and lung cancer (13,27).
It was therefore investigated whether pyrvinium pamoate inhibited
the Wnt/β-catenin signaling in CML cells. The levels of β-catenin
were markedly decreased in two pairs of CML cells (KBM5 and
KBM5-T315I cells or 32D-BCR-ABL and 32D-T315I cells)(Fig. 4B). Subsequently, the upstream and
downstream proteins of the Wnt/β-catenin signaling pathway were
measured. The results indicated that the levels of p-GSK3β (Ser9),
cyclin D1 and c-Myc were decreased (Fig. 4B), suggesting that pyrvinium
pamoate may block the activation of canonical Wnt/β-catenin
signaling.
Due to the fact that nuclear translocation of
β-catenin is required for its functions (i.e., to activate T-cell
factor/lymphoid enhancer factor), the effects of pyrvinium pamoate
on the subcellular distribution of β-catenin were measured. The
results indicated that the levels of β-catenin in nuclear and
cytosolic fractions were reduced by pyrvinium pamoate in a
dose-dependent manner (Fig.
4C).
Discussion
The present study aimed to investigate the
anti-tumor activity of pyrvinium pamoate, an anthelminthic drug, in
CML cells harboring the T315I mutation. The results demonstrated
that this agent is equally effective in inhibiting proliferation
and inducing apoptosis in CML cells harboring either wild-type or
T315I BCR-ABL. Due to the fact that pyrvinium pamoate is an
FDA-approved drug, the results of the present study may lead to a
clinical trial for imatinib-resistant patients with T315I
BCR-ABL.
Pyrvinium pamoate is a quinolone-derived cyanine
dye, and the FDA approved its anthelmintic property in the 1950s
(28). This agent has been
previously investigated in solid tumors and hematopoietic
malignancy (8–14). It was identified that pyrvinium
pamoate potently inhibited the viability and induced apoptosis in
both wild-type and T315I BCR-ABL-expressing cells at low nanomolar
concentrations. The results were consistent with the report by
Xiang et al (16) regarding
the effect of pyrvinium pamoate on CML cells with wild-type
BCR-ABL. Of note, pyrvinium pamoate was reported to be active
against primary CD34+ blast cells from 8 patients with
CML who harbored BCR-ABL point mutations (E255K, E453K and E255V).
The results of the present study were in agreement with those from
the literature (16), suggesting
that this agent may be a promising potential therapy for patients
with CML who harbor wild-type and different point mutations of
BCR-ABL including T315I.
Although it has been used for more than 50 years,
the molecular mechanisms of pyrvinium pamoate remain unclear. It
has been reported that in different types of cancer that pyrvinium
pamoate may target different signaling pathways (8–11).
We observed that the inhibitory effects of pyrvinium pamoate on
mutant CML cells might be associated with its apoptotic action.
Apoptosis, which is programmed cell death, can occur via the cell
death receptor-mediated extrinsic pathway or the
mitochondria-mediated intrinsic pathway (29). Reduction of apoptosis is believed
to contribute to cancer development. In the present study, it was
observed that pyrvinium pamoate induced apoptosis in both the
wild-type and T315I BCR-ABL mutant CML cells (Figs. 2 and 3). The apoptosis may be caused by the
mitochondrial-dependent pathway, as evidenced by the release of AIF
and cytochrome c to the cytosol was increased following
treatment with pyrvinium pamoate (Fig.
3A).
Previous studies indicated that pyrvinium pamoate
could inhibit the growth and differentiation of different cancer
cells through the Hedgehog signaling pathway (14), or Wnt/β-catenin signaling pathway
(13,27,30).
Consistent with those results, decreased levels of β-catenin were
observed, and the upstream and downstream proteins of Wnt/β-catenin
pathway, including phospho-GSK3β (Ser 9) and c-Myc in CML cells
either with wild-type or T315I BCR-ABL mutations following
pyrvinium pamoate treatment (Fig.
4B). Thus, it is suggested that pyrvinium pamoate inhibits the
growth of CML cells by inhibiting the Wnt/β-catenin signaling
pathway. Taken together, it may be beneficial to investigate the
clinical efficacy of pyrvinium pamoate in treating patients with
CML bearing T315I BCR-ABL.
In summary, it was reported that pyrvinium pamoate
effectively inhibits the proliferation and induces apoptosis in the
wild-type or T315I BCR-ABL-expressing cells, suggesting that
pyrvinium pamoate is a promising agent that may be used for
treatment of patients with imatinib-resistant T315I BCR-ABL CML in
addition to imatinib-sensitive wild-type CML cells.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81025021,
U1301226 and 81373434 to Professor J.P.; grant nos. 81473247 and
81673451 to Dr Y.J.), Guangdong Natural Science Funds for
Distinguished Young Scholars (grant no. 2016A030306036 to Dr Y.J.),
the Natural Science Foundation of Guangdong Province (grant no.
2015A030312014 to Professor J.P.), and the Research Foundation of
Education Bureau of Guangdong Province, China (grant no. cxzd1103
to Professor J.P.).
Glossary
Abbreviations
Abbreviations:
|
CML
|
chronic myeloid leukemia
|
|
TKIs
|
tyrosine kinase inhibitors
|
|
COX II
|
cytochrome c oxidase subunit II
|
|
AIF
|
apoptosis-inducing factor
|
References
|
1
|
Rowley JD: Letter: A new consistent
chromosomal abnormality in chronic myelogenous leukaemia identified
by quinacrine fluorescence and Giemsa staining. Nature.
243:290–293. 1973. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
McLaughlin J, Chianese E and Witte ON: In
vitro transformation of immature hematopoietic cells by the P210
BCR/ABL oncogene product of the philadelphia chromosome. Proc Natl
Acad Sci USA. 84:pp. 6558–6562. 1987; View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Slupianek A, Falinski R, Znojek P,
Stoklosa T, Flis S, Doneddu V, Pytel D, Synowiec E, Blasiak J,
Bellacosa A and Skorski T: BCR-ABL1 kinase inhibits uracil DNA
glycosylase UNG2 to enhance oxidative DNA damage and stimulate
genomic instability. Leukemia. 27:629–634. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim SJ, Jung KH, Yan HH, Son MK, Fang Z,
Ryu YL, Lee H, Lim JH, Suh JK, Kim J, et al: HS-543 induces
apoptosis of Imatinib-resistant chronic myelogenous leukemia with
T315I mutation. Oncotarget. 6:1507–1518. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bu Q, Cui L, Li J, Du X, Zou W, Ding K and
Pan J: SAHA and S116836, a novel tyrosine kinase inhibitor,
synergistically induce apoptosis in imatinib-resistant chronic
myelogenous leukemia cells. Cancer Biol Ther. 15:951–962. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Senior M: FDA halts then allows sales of
Ariad's leukemia medication. Nat Biotechnol. 32:9–11. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jung RC: Treatment of intestinal parasitic
disease. South Med J. 69:799–804. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lamb R, Ozsvari B, Lisanti CL, Tanowitz
HB, Howell A, Martinez-Outschoorn UE, Sotgia F and Lisanti MP:
Antibiotics that target mitochondria effectively eradicate cancer
stem cells, across multiple tumor types: Treating cancer like an
infectious disease. Oncotarget. 6:4569–4584. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu W, Lacerda L, Debeb BG, Atkinson RL,
Solley TN, Li L, Orton D, McMurray JS, Hang BI, Lee E, et al: The
antihelmintic drug pyrvinium pamoate targets aggressive breast
cancer. PLoS One. 8:e715082013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Deng L, Lei Y, Liu R, Li J, Yuan K, Li Y,
Chen Y, Liu Y, Lu Y, Edwards CK III, et al: Pyrvinium targets
autophagy addiction to promote cancer cell death. Cell Death Dis.
4:e6142013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Harada Y, Ishii I, Hatake K and Kasahara
T: Pyrvinium pamoate inhibits proliferation of
myeloma/erythroleukemia cells by suppressing mitochondrial
respiratory complex I and STAT3. Cancer Lett. 319:83–88. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu DH, Macdonald J, Liu G, Lee AS, Ly M,
Davis T, Ke N, Zhou D, Wong-Staal F and Li QX: Pyrvinium targets
the unfolded protein response to hypoglycemia and its anti-tumor
activity is enhanced by combination therapy. PLoS One. 3:e39512008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Thorne CA, Hanson AJ, Schneider J, Tahinci
E, Orton D, Cselenyi CS, Jernigan KK, Meyers KC, Hang BI, Waterson
AG, et al: Small-molecule inhibition of Wnt signaling through
activation of casein kinase 1α. Nat Chem Biol. 6:829–836. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li B, Fei DL, Flaveny CA, Dahmane N,
Baubet V, Wang Z, Bai F, Pei XH, Rodriguez-Blanco J, Hang B, et al:
Pyrvinium attenuates Hedgehog signaling downstream of smoothened.
Cancer Res. 74:4811–4821. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li B, Flaveny CA, Giambelli C, Fei DL, Han
L, Hang BI, Bai F, Pei XH, Nose V, Burlingame O, et al: Repurposing
the FDA-approved pinworm drug pyrvinium as a novel chemotherapeutic
agent for intestinal polyposis. PLoS One. 9:e1019692014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xiang W, Cheong JK, Ang SH, Teo B, Xu P,
Asari K, Sun WT, Than H, Bunte RM, Virshup DM and Chuah C:
Pyrvinium selectively targets blast phase-chronic myeloid leukemia
through inhibition of mitochondrial respiration. Oncotarget.
6:33769–33780. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang H, Trachootham D, Lu W, Carew J,
Giles FJ, Keating MJ, Arlinghaus RB and Huang P: Effective killing
of Gleevec-resistant CML cells with T315I mutation by a natural
compound PEITC through redox-mediated mechanism. Leukemia.
22:1191–1199. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu Z, Jin Y, Qiu L, Lai Y and Pan J:
Celastrol, a novel HSP90 inhibitor, depletes Bcr-Abl and induces
apoptosis in imatinib-resistant chronic myelogenous leukemia cells
harboring T315I mutation. Cancer Lett. 290:182–191. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qiu L, Wang XD, Yu BH, Lu RZ, Ge F, Wang
XL, Chen LJ, Han BH, Zhan ZM, Zhang BL and Ma J: Effect of a novel
tyrosine kinase inhibitor HHGV678 on growth inhibition of Bcr-Abl
wild type and IM-resistant cell lines in vitro. Zhongguo Shi Yan
Xue Ye Xue Za Zhi. 16:1039–1043. 2008.(In Chinese). PubMed/NCBI
|
|
20
|
Pan J, Quintas-Cardama A, Kantarjian HM,
Akin C, Manshouri T, Lamb P, Cortes JE, Tefferi A, Giles FJ and
Verstovsek S: EXEL-0862, a novel tyrosine kinase inhibitor, induces
apoptosis in vitro and ex vivo in human mast cells expressing the
KIT D816V mutation. Blood. 109:315–322. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jin Y, Chen Q, Shi X, Lu Z, Cheng C, Lai
Y, Zheng Q and Pan J: Activity of triptolide against human mast
cells harboring the kinase domain mutant KIT. Cancer Sci.
100:1335–1343. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jin Y, Lu Z, Ding K, Li J, Du X, Chen C,
Sun X, Wu Y, Zhou J and Pan J: Antineoplastic mechanisms of
niclosamide in acute myelogenous leukemia stem cells: Inactivation
of the NF-kappaB pathway and generation of reactive oxygen species.
Cancer Res. 70:2516–2527. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lu B, Li J, Pan J, Huang B, Liu J and
Zheng D: Everolimus enhances the cytotoxicity of bendamustine in
multiple myeloma cells through a network of pro-apoptotic and
cell-cycle-progression regulatory proteins. Acta Biochim Biophys
Sin (Shanghai). 45:683–691. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shi X, Jin Y, Cheng C, Zhang H, Zou W,
Zheng Q, Lu Z, Chen Q, Lai Y and Pan J: Triptolide inhibits Bcr-Abl
transcription and induces apoptosis in STI571-resistant chronic
myelogenous leukemia cells harboring T315I mutation. Clin Cancer
Res. 15:1686–1697. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pan J, Xu G and Yeung SC: Cytochrome c
release is upstream to activation of caspase-9, caspase-8, and
caspase-3 in the enhanced apoptosis of anaplastic thyroid cancer
cells induced by manumycin and paclitaxel. J Clin Endocrinol Metab.
86:4731–4740. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Blonska M, Shambharkar PB, Kobayashi M,
Zhang D, Sakurai H, Su B and Lin X: TAK1 is recruited to the tumor
necrosis factor-alpha (TNF-alpha) receptor 1 complex in a
receptor-interacting protein (RIP)-dependent manner and cooperates
with MEKK3 leading to NF-kappaB activation. J Biol Chem.
280:43056–43063. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang X, Lou Y, Zheng X, Wang H, Sun J,
Dong Q and Han B: Wnt blockers inhibit the proliferation of lung
cancer stem cells. Drug Des Devel Ther. 9:2399–2407.
2015.PubMed/NCBI
|
|
28
|
Beck JW, Saavedra D, Antell GJ and Tejeiro
B: The treatment of pinworm infections in humans (enterobiasis)
with pyrvinium chloride and pyrvinium pamoate. Am J Trop Med Hyg.
8:349–352. 1959. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu MD, Xiong SJ, Tan F and Liu Y:
Physcion 8-O-β-glucopyranoside induces mitochondria-dependent
apoptosis of human oral squamous cell carcinoma cells via
suppressing survivin expression. Acta Pharmacol Sin. 37:687–697.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Basu D, Reyes-Mugica M and Rebbaa A: Role
of the beta catenin destruction complex in mediating
chemotherapy-induced senescence-associated secretory phenotype.
PLoS One. 7:e521882012. View Article : Google Scholar : PubMed/NCBI
|