Introduction
Diabetic nephropathy (DN) is a major cause of
end-stage renal failure and is one of the direct causes of
mortality in diabetic patients (1,2). In
clinical terms, DN patients are commonly characterized by the
development of proteinuria, specifically albuminuria, with a
subsequent reduction in the glomerular filtration rate (2). Multitude pathological changes in the
kidneys contribute to the progressive decline in renal function,
including macrophages in filtration, extracellular matrix (ECM)
accumulation in multiple renal cells, glomerular and tubular
hypertrophy, endothelial dysfunction, mesangial cell expansion and
podocyte injury (3).
Identification of molecular changes associated with the
pathophysiology of DN is imperative for the development of
therapeutic strategies.
The loss of size-selective and/or charge-selective
properties of the glomerular filtration barrier are reported to be
responsible for the observed proteinuria in glomerular diseases.
The glomerular filtration barrier is composed of three layers: A
glomerular basement membrane (GBM), a fenestrated endothelial layer
and podocyte foot processes that are connected by a slit diaphragm.
Diaphragm-associated proteins podocin and nephrin (4) are crucial proteins in the complex of
molecules that assemble and reinforce the slit diaphragm and serve
central roles in maintaining the size-selective barrier (5,6).
Advanced glycation end-products (AGEs) are
biochemical end-products of nonenzymatic glycation (7). AGEs have been suggested to serve a
vital role in the development of DN due to the fact that drugs that
either break the AGE crosslink or inhibit AGE formation have been
demonstrated to exhibit a protective effect on experimental DN
(8). The structure and function of
matrix tissue proteins can be altered by AGE modification, which
thus stimulates a variety of cellular responses via a specific
cell-surface receptor on glomerular cells (9). Importantly, the specific cellular
receptor for AGE (RAGE) that is upregulated and activated by AGE
(10), has been demonstrated to
serve an important role in the development of DN (11). Therefore, the AGE-RAGE interaction
likely serves an important role in the pathogenesis of DN (12–14).
GSPE is derived from grape seeds and possesses
potent properties, such as radical scavenging, anti-inflammation,
antioxidant, anti-tumor and cardiovascular protection activity
(15). Previous studies
demonstrate that GSPE can act in the prevention of multiple
diseases such as cataracts, gastric ulcer, atherosclerosis,
myocardial ischemia-reperfusion and especially diabetes (16–18).
However, the function of GSPE on DN has been rarely studied. The
aim of the current study was to determine the effect of GSPE on the
streptozotocin-induced diabetic rat model and to explore the
underlying mechanisms of its activity.
Materials and methods
Ethics statement
The current study was approved by the Animal Ethics
Committee of Shandong University (Jinan, Shandong, China).
Animals
A total of 60 male Wistar rats (lot no.
SCXL20030004; 6–7 weeks old; weight, 180–200 g) were obtained from
the Shandong University Experimental Animal Center (Jinan,
Shandong, China). These rats were housed in cages and allowed free
access to food and tap water. They were maintained on a 12:12 h
dark/light cycle, with a room temperature of 22±1.5°C and a
humidity of 55±5%. A total of 24 rats were selected randomly and
divided into 2 groups (n=12 in each group), and the control group
(C group) was treated with a dosage of 250
mg·kg−1·day−1 GSPE (CT group) (lot no.
G050412; Tianjin Jianfeng Natural Product R&D Co., Ltd.,
Tianjin, China) (14). The
remaining 36 rats were injected with a dose of 55
mg·kg−1 STZ (injected into the tail veins). After five
days, rats with blood glucose levels higher than 16.7 mmol/l were
considered as being diabetic, and six rats were excluded for
failure to meet this criterion. One week after STZ injection, the
diabetic rats were then divided randomly into 2 groups (n=15 in
each group), a diabetic group without any treatment (diabetes
mellitus; DM; group), and another diabetic group that was treated
with a dosage of 250 mg·kg−1·day−1 GSPE (T
group). The GSPE was intragastrically administrated in normal
saline solution for 24 weeks. At the end of the experiments, the
animals were fasted overnight for 18 h and then anesthetized with
intraperitoneal injection of sodium pentobarbitone (60 mg/kg;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and sacrificed.
Kidneys were perfused with ice-cold PBS and then were stored at
−80°C.
Assessment of renal function
At the end of the experiments, the fasting plasma
glucose (FPG), 24 h urinary albumin excretion, hemoglobin A1c
(HbA1C), urinary creatinine (Ucr), serum creatinine
(Scr), Ccr = Ucr/ScrxV (V:ml/min, urine per minute), and the ratio
of kidney weight to bodyweight and serum AGEs was measured for each
rat. Scr and Ucr were measured using an automatic biochemical
analysis instrument (DVI 1650; Bayer AG, Leverkusen, Germany).
HbA1C was measured by high-pressure liquid phase methods
(ADAMS A1c HA-8180; Arkray, Inc., Kyoto, Japan). The level of AGEs
in serum was measured by the enzyme-linked immunosorbent assay
(ELISA) method (19) (Hitachi 850;
Hitachi, Tokyo, Japan). To collect urine samples, rats were placed
in individual metabolic cages for 24 h prior to sacrificing. ELISA
(Nephrat II; Exocell, Philadelphia, PA, USA) was used to determine
24 h urinary albumin excretion levels.
Light microscope examination
For the light microscopic examination, the kidneys
were immersion-fixed, embedded and two sections (4 µm thickness)
were stained with periodic acid-Schiff (PAS) reagent. For electron
microscope examination, the renal cortex was cut into small pieces
and pre-fixed in glutaraldehyde, post-fixed in buffered sodium
tetroxide for 1 h, and then embedded in EPON 812 embedding resin
(Electron Microscopy Sciences, Fort Washington, PA, USA). The
PAS-positive area present in the mesangial region excluding
cellular elements indicated mesangial matrix expansion. By using
Leica QWin version 3 image analysis software (Leica Microsystems
GmbH, Wetzlar, Germany), the percentage of the PAS-positive area in
the glomerulus was analyzed. A total of 10 glomeruli, randomly
selected in the two slides from the rats (a minimum of 8 rats in
each group), were evaluated by two investigators blinded of the
origins of the slides.
Electron microscope examination
For transmission electron microscopic examination, a
JEOL JSM 1011 microscope (JEOL, Ltd., Tokyo, Japan), was used to
photograph images covering one or two glomerular cross-sections.
The images at ×15,000 magnification were used to measure the GBM
length and the number of slit pores. To determine the FPW (foot
process width), the slit pore length, and the GBM thickness, the
images at ×50,000 or ×100,000 magnification were used according to
published methods (20–22).
Reverse transcription-polymerase chain
reaction (RT-PCR)
The total RNA was extracted from the renal cortex
using TRIzol reagent (Thermo Fisher Scientific Inc., Waltham, MA,
USA). Primers for RAGE, nephrin, podocin, and
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were designed and
synthesized (Huada Gene Center, Beijing, China) based on the
published sequence. The primers were as follows: RAGE forward,
5′-GCTCTGACCGAAGCGTGA-3′ and reverse, 5′-CCTTCAGGCTCAACCAACAG-3′,
240 bp product; nephrin forward, 5′-ACCAAGTCCAGTCGCCCAG-3′ and
reverse, 5′-ATACCAGCCTCACCCGAGTCCC-3′, 306 bp product; podocin
forward, 5′-GTGTCCAAAGCCATCCAGTT-3′ and reverse,
5′-GTCTTTGTGCCTCAGCTTCC-3′, 232 bp product; and GAP DH forward,
5′-GAGGGGCCATCCACAGTCTTCTG-3′ and reverse,
5′-CCCTTCATTGACCTCAACTACATGGT-3′. The Titan One Tube RT-PCR kit
(Boehringer-Mannheim, Shanghai, China) was used to amplify a total
of 0.5 µg RNA. Agarose gel electrophoresis was used to separate
products and ethidium bromide staining was used for visualization.
A Tanon-1000 Gel Image System (Tanon, Shanghai, China) was used to
digitize bands and GAPDH was used as the control gene based on
previous studies (5,23).
Western blotting
Western blot analysis was performed as previously
described (24). Renal tissues
were homogenized with ice-cold lysis buffer and were centrifuged at
10,000 × g for 15 min at 4°C. Equal amounts (100 µg) of protein
were loaded and separated by 10% SDS-PAGE. Separated proteins were
then transferred onto nitrocellulose paper (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). A sample of total protein from the lysate
(20 µg) was separated and blotted. The membranes were incubated
with the following primary antibodies: Anti-RAGE (1:250, cat no.
MAB1179; R&D Systems, Inc., Indianapolis, IN, USA),
anti-nephrin (1:1,000, cat no. ab58968; Abcam, Cambridge, UK),
anti-podocin (1:1,000, cat no. sc-21009; Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA) and anti-β-actin (1:500, cat no. ab8226;
Abcam) overnight at 4°C, and then reacted with anti-rabbit IgG
horseradish peroxidase-conjugated secondary antibodies (1:5,000,
cat no. sc-2004; Santa Cruz Biotechnology, Inc.) at room
temperature for 1.5 h. Signal detection was performed via exposure
of the blot to enhanced diaminobenzidenecolor reagents (OriGene
Technologies, Inc., Rockville, MD, USA) for 5 min. Quantification
of the luminosity of each identified protein band was performed
using Adobe Photoshop software (AdobePhotoshop 7.0; Adobe Systems,
Inc., San Jose, CA, USA).
Immunohistochemical study
Renal tissue sections (4 µm) were subjected to
immunohistochemical staining based on the published method with
specific antibodies anti-RAGE (1:100, ab37647; Abcam), anti-nephrin
(1:500, ab58968; Abcam), and anti-podocin (1:400, H-130, sc-21009;
Santa Cruz Biotechnology, Inc.) (25). Color was developed by incubating
with diaminobenzidine and counterstaining with hematoxylin.
Controls were obtained by replacing the primary antibody with PBS.
Semi quantitative analysis of the percentage of positive staining
area in the glomeruli and tubules was evaluated using a computer
imaging analysis system (Image-Pro Plus software version 4·5; Media
Cybernetics, Inc., Rockville, MD, USA). The brown areas were judged
as positive.
Statistical analysis
Data were presented as the mean ± standard
deviation. One way analysis of variance or the Mann-Whitney U test
was performed to compare the differences between groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
Changes of renal function and general
parameters
There were four treatment groups, the control group
(C group) that received no treatment, rats treated with
250·mg·kg−1·day−1 GSPE (CT group), rats that
received STZ injection to induce diabetes, however no additional
treatment (DM group) and rats that were diabetic but received
250·mg·kg−1·day−1 GSPE (T group). Rats in
each group were observed from 1 week to 24 weeks. At the start of
the experiment, there were 12 rats in each of the C and CT groups
and 15 rats in each of the T and DM groups. At the end of the
experiment, there were 10, 10, 8, and 12 surviving rats in the C,
CT, DM and T groups, respectively. As presented in Table I, the FPG, HbA1C, 24 h
urinary albumin excretion, Ccr and ratio of kidney weight to body
weight in the DM group were significantly increased compared with
the C group (P<0.01). The diabetic rats treated with GSPE (T
group) had markedly reduced Ccr, urinary albumin excretion
(P<0.01), and ratio of kidney weight to body weight than that of
the DM group. However, no significant differences in FPG and
HbA1C were identified between the DM and the T
groups.
 | Table I.General data of the rats. |
Table I.
General data of the rats.
| Group | n | FPG (mmol/l) | HbA1C
(%) | Ccr (ml/min) | Urinary albumin
excretion 24 h (mg) | Kidney weight/body
weight (×10−3) |
|---|
| C | 10 |
6.98±0.86 |
5.60±0.73 |
1.98±0.32 |
10.22±1.67 |
3.34±0.18 |
| CT | 10 |
6.60±0.81 |
5.56±0.71 |
2.11±0.74 |
11.37±1.08 |
3.24±0.18 |
| DM | 8 |
23.04±0.93a |
11.41±2.14a |
5.11±1.14a |
24.96±2.56a |
5.48±0.17a |
| T | 12 |
21.22±1.12a |
8.71±3.54a |
2.80±1.23b,c |
15.15±3.27a,c |
4.30±0.17a,c |
Changes in serum AGE levels
As presented in Table
II, animals that were injected with STZ exhibited a
significantly higher level of AGEs in serum than that in the C
group (P<0.01). Diabetic rats that were treated with GSPE (T
group) had significantly lower AGEs than the level in the DM group
(P<0.01).
| Table II.Alterations in serum AGE levels in
rats. |
Table II.
Alterations in serum AGE levels in
rats.
| Group | n | AGEs |
|---|
| C | 10 | 0.0182±0.0005 |
| CT | 10 | 0.0221±0.0063 |
| DM | 8 |
0.0351±0.0021a |
| T | 12 |
0.0276±0.0054b |
Renal morphological changes
The PAS-staining results in the glomeruli of each
group are presented in Fig. 1A.
Quantitative analysis of the percentage of the PAS-positive area in
the glomeruli is summarized in Fig.
1B. The ECM accumulation was significantly higher in the
glomeruli of the DM group than that of the C group (P<0.01), and
diabetic rats treated with GSPE (T group) had a significantly lower
ECM accumulation in the glomeruli than that of the DM group
(P<0.01).
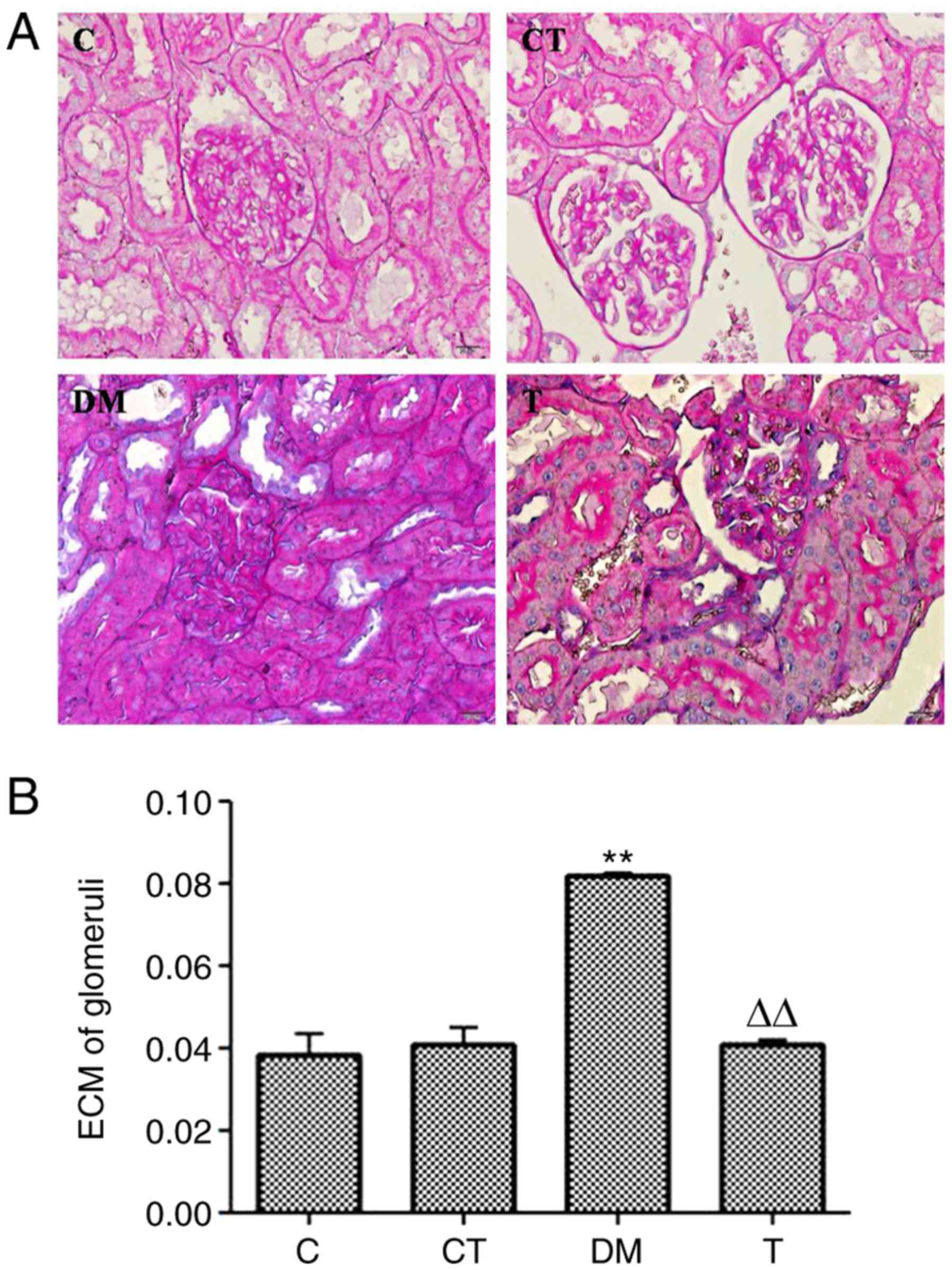 | Figure 1.(A) Light microscopic images of renal
morphological changes in rats by PAS (PAS, magnification ×400). ECM
accumulation in DN rat kidneys by PAS staining. C, normal kidney;
CT, normal rats treated with GSPE (250
mg·kg−1·day−1); DM, DM rats; T, DM rats
treated with GSPE (250 mg·kg−1·day−1). (B)
Analysis of ECM in glomeruli by PAS staining. Higher ECM
accumulation was observed in the kidneys of DM group rats compared
with that of the C group (**P<0.01 vs. C) Lower ECM accumulation
was observed in the kidneys of the T group rats compared with the
DM group (∆∆P<0.01 vs. DM). ECM, extracellular
matrix; DN, diabetic nephropathy; PAS, periodic acid-Schiff; DM,
diabetes mellitus; GSPE, grape seed proanthocyanidin extracts. |
Electron microscopic observations
The foot processes of glomeruli were broader and
flatter in the DM group than that in the other groups. It was also
identified that the mean FPW was significantly greater in diabetic
glomeruli as compared with control glomeruli (P<0.01). The mean
slit pore length was significantly shorter in diabetic glomeruli
than that of the control glomeruli (P<0.01). Additionally, there
were significantly fewer slit pores (per 100 µm) of GBM in diabetic
glomeruli than in the control glomeruli (P<0.01). The GBM was
significantly thicker in diabetic glomeruli as compared with
control glomeruli (P<0.01). However, these changes were
significantly reversed by GSPE treatment (Fig. 2 and Table III).
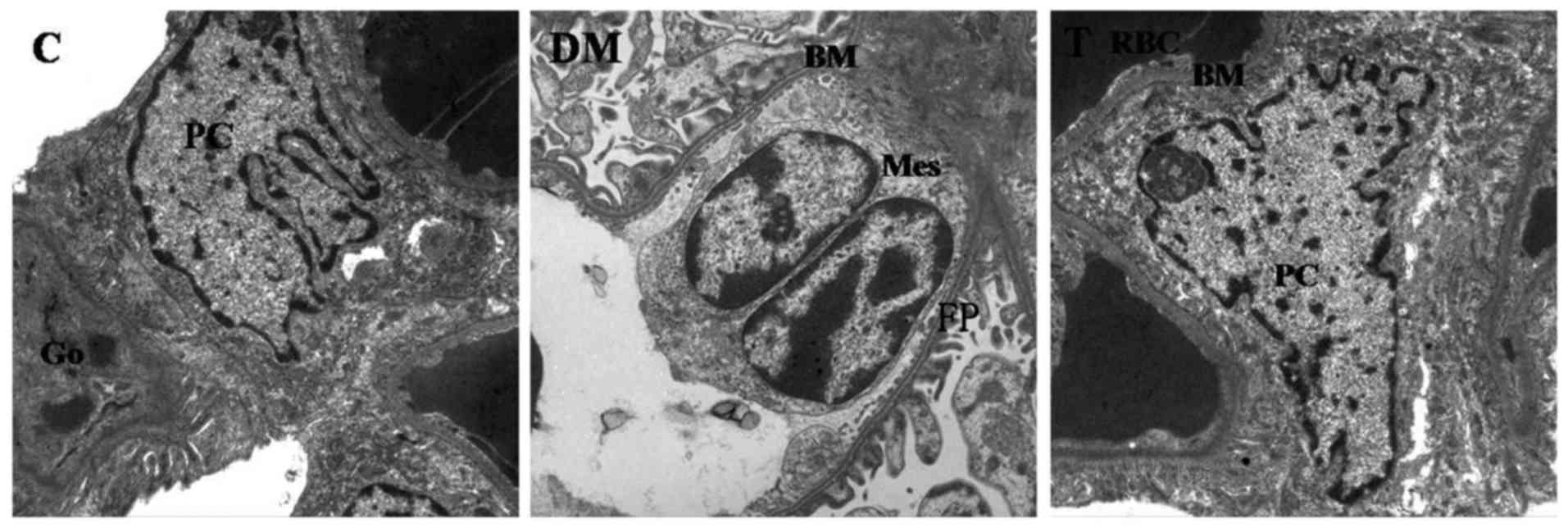 | Figure 2.Electron microscopic images of renal
microstructure changes in rats (magnification, ×15,000). GSPE
treatment decreased STZ-induced renal ultrastructural
abnormalities. (Original magnification ×15,000.) GSPE, grape seed
proanthocyanidin extracts; BM, basement membrane; PC, podocyte;
End, endothelial cell; FP, foot processes; Mes, mesangial cell; Go,
golgi apparatus; RBC, red blood cell. |
| Table III.Electron microscopic findings in the
four groups. |
Table III.
Electron microscopic findings in the
four groups.
| Parameter | C | DM | T |
|---|
| n | 10 | 8 | 12 |
| Foot process width
(nm) | 296.2±10.5 |
404.5±20.7a |
353.9±18.4b |
| Slit pore length
(nm) | 32.5±2.6 |
26.2±3.2a |
30.6±3.4b |
| Number of slit
pores (per 100 µm) | 256.3±13.2 |
181.5±12.1a |
239.2±13.5b |
| GBM thickness
(nm) | 165.8±20.5 |
270.2±29.8a |
190.3±22.3b |
Renal expression of RAGE, nephrin, and
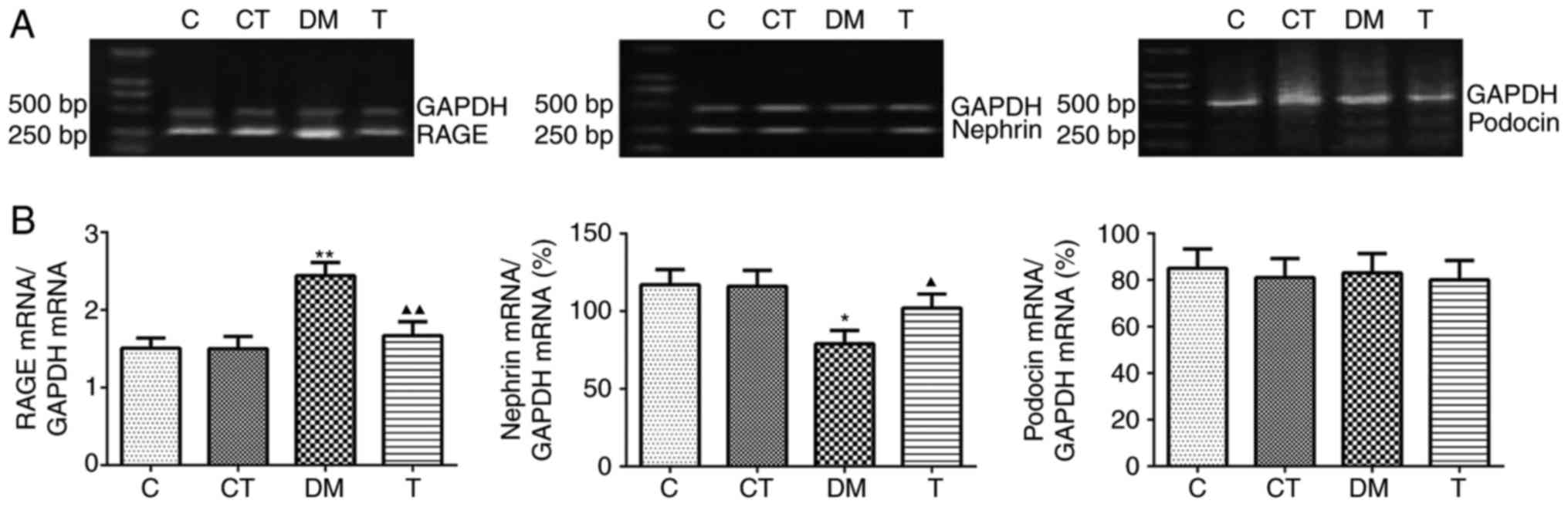
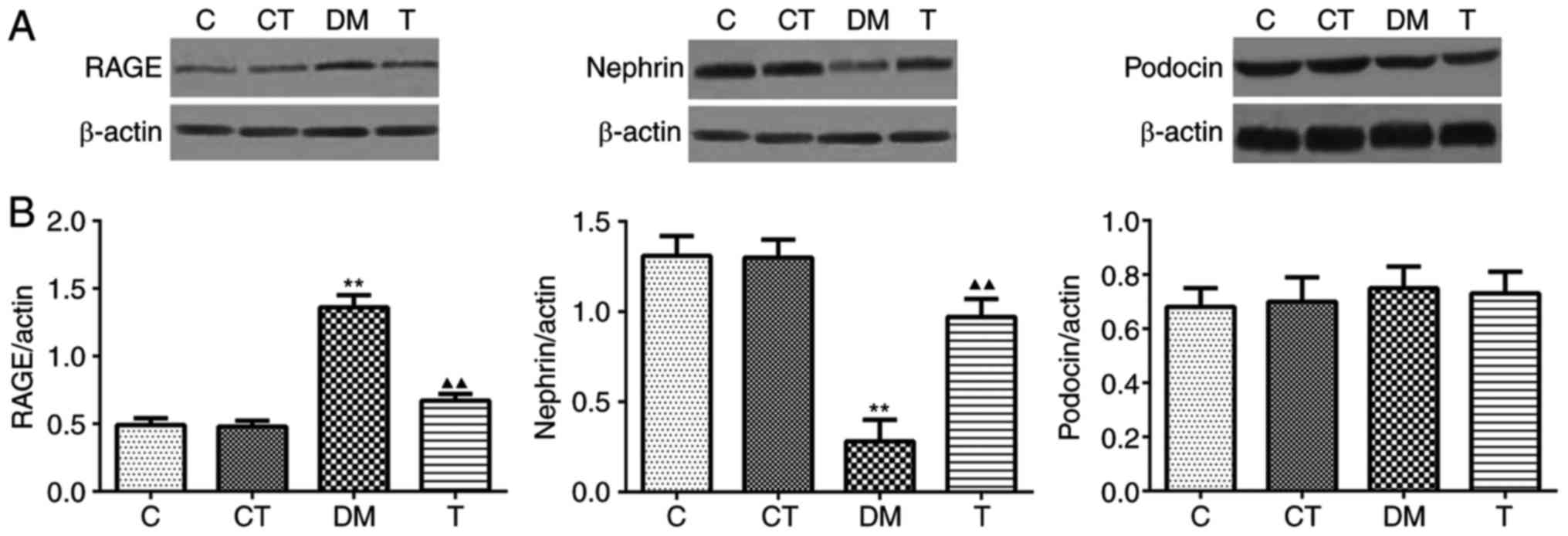
podocin by RT-PCR and western blotting
In the kidney cortex of DM rats, the mRNA and
protein expression of RAGE were markedly increased compared with
the levels in the C group (P<0.01), and GSPE treatment
significantly decreased the expression of RAGE (P<0.01)
(Figs. 3 and 4). A significantly decreased expression
of nephrin mRNA and protein was observed in the DM group as
compared with the C group (P<0.05 and P<0.01, respectively),
and GSPE treatment significantly increased the mRNA expression and
protein level of nephrin (P<0.05 and P<0.01, respectively)
(Figs. 3 and 4). However, no significant differences
were identified in the mRNA or protein expression of podocin among
the four groups (P>0.05) (Figs.
3 and 4).
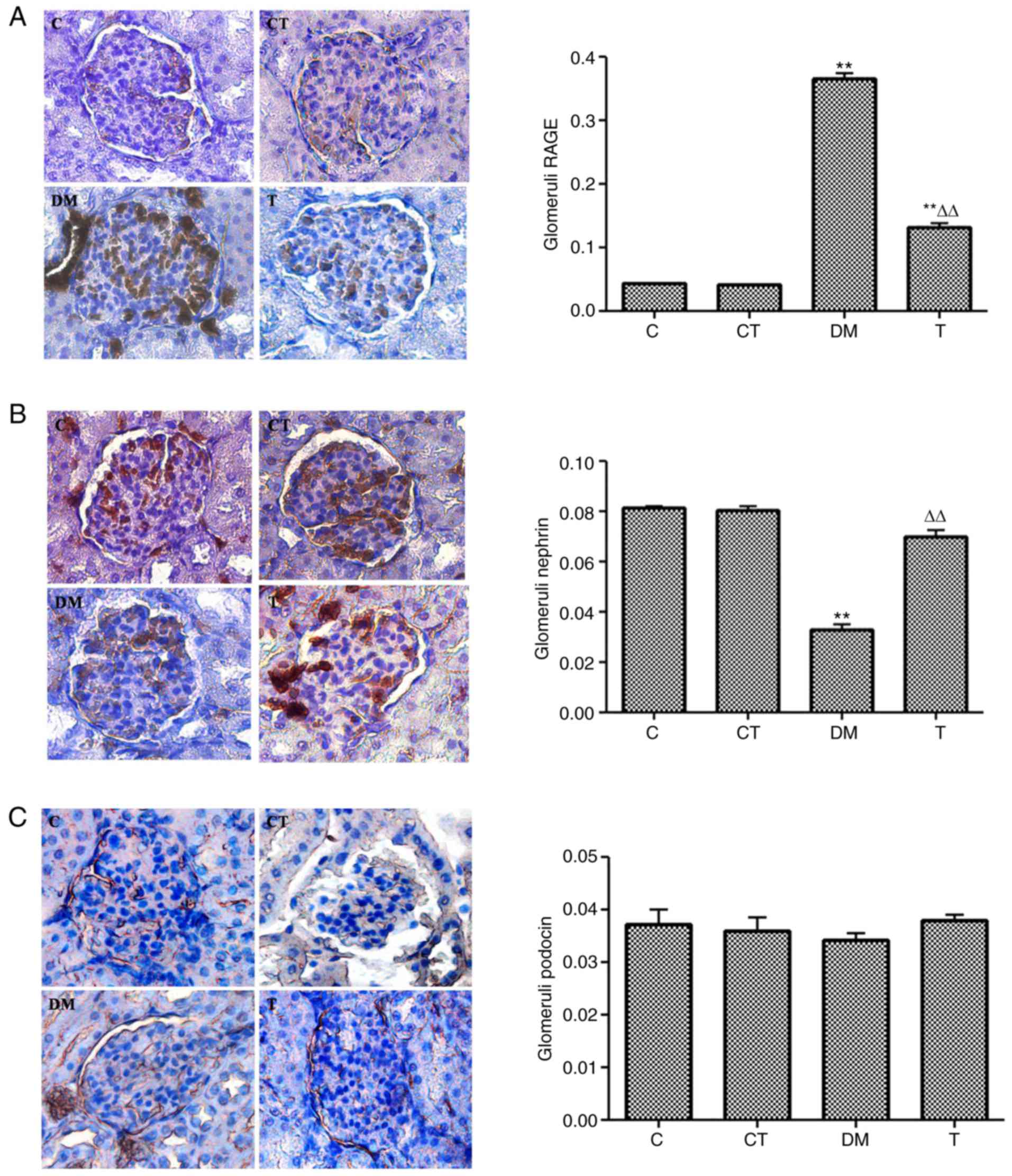
Immunohistochemical staining for RAGE,
nephrin, and podocin protein in glomeruli
Immunohistochemical staining indicated the
localization of signals for RAGE, nephrin and podocin predominantly
in the glomeruli. The intensity and area of RAGE staining were
significantly increased in the glomeruli of diabetic rats as
compared with the C group, and GSPE treatment significantly
reversed this effect (Fig. 5A).
The intensity and area of nephrin staining were significantly
decreased in the glomeruli of diabetic rats compared with the C
group, and GSPE treatment also significantly reversed these effects
(Fig. 5B). No differences were
observed in the intensity and area of podocin staining in the
glomeruli for the four groups (Fig.
5C).
Discussion
The incidence of diabetic nephropathy is increasing
worldwide, and identification of associated molecular changes is
important for prevention and treatment (1,2).
Among the three intrinsic cells in the glomerulus, podocytes have
special physiological functions and cytobiological traits (26). Podocyte damage occurs in the early
stage of DN, and this injury to podocytes can cause proteinuria,
and accelerate the development of DN (27). The slit diaphragm bridges adjacent
foot processes derived from different podocytes and functions as
the ultimate molecular size filter. Nephrin serves an important
role in the pathogenesis of proteinuria in DN as a
podocyte-associated protein that can comprise the slit diaphragm
(28,29). Additionally, proteinuria is closely
associated with ultrastructural changes in podocytes in DN. Reduced
slit pore length and increased foot process width have been
demonstrated in animal models of diabetes in addition to in
patients with diabetes (30,31).
Berg et al (30) also
identified a strong correlation between FPW and urinary albumin
excretion inpatients with type 1 diabetes.
In the present study, the HbA1C, 24 h
urinary albumin excretion, Ccr, and ratio of kidney weight to body
weight in DM group were significantly higher than in the C group.
The electron microscopic analysis demonstrated that the foot
processes of glomeruli in the DM group were broader and flatter
than that of the other groups. In diabetic glomeruli, the FPW was
significantly greater, the mean slit pore length was significantly
shorter, the number of slit pores (per 100 µm) of GBM was
significantly fewer and the GBM was significantly thicker as
compared with control glomeruli. Notably, the current study
observed that GSPE treatment significantly reversed these
morphological changes. Additionally, markedly reduced Ccr, urinary
albumin excretion and ratio of kidney weight to body weight were
observed in the T group relative to that of the DM group. These
results suggested that GSPE can decrease proteinuria and aid in the
maintenance podocyte microstructure.
In order to explore the underlying mechanism of GSPE
action, the effects on the podocyte-associated molecules nephrin
and podocin were examined. The results indicated that expression of
nephrin was significantly decreased, however the expression of
podocin was unchanged in diabetic rats, which was also observed in
a previous study (32). GSPE
treatment significantly increased the expression of nephrin in the
diabetic rats. These results suggested that the mechanism by which
GSPE decreases proteinuria may be associated with the improved
expression of nephrin.
AGEs have been documented to serve an important role
in stimulating growth factor and cytokine synthesis, which
contributes to the pathogenesis of DN (33). RAGE-dependent mechanisms are likely
to be responsible for the AGE-induced tissue damage and
dysfunction. In vivo study in RAGE-overexpressing diabetic
mice observed that interaction between AGEs and their receptor is
important in the development of DN (24). AGE-RAGE interactions may lead to
diabetic vascular derangement, or trigger and promote the
development of renal lesions (24). Additionally, a previous study
suggested that, in a mouse model of DN, inactivation of the RAGE
gene results in significant suppression of kidney changes,
including increased glomerular cell number, advanced
glomerulosclerosis, kidney enlargement, mesangial expansion,
increased albuminuria, and increased serum creatinine compared with
wild-type diabetic mice (34).
Importantly, a recent study indicated that blockade of RAGE using a
neutralizing antibody prevented the loss of nephrin expression
induced by glycated albumin (GA), suggesting the involvement of
this receptor in mediating the effect of GA on nephrin expression
(35). In the present study, in
diabetic rats, markedly higher AGEs and increased expression of
RAGE were observed in the DM group relative to the C group.
However, diabetic rats treated with GSPE had lower
serum AGEs and decreased expression of RAGE. The inhibition by GSPE
of AGEs formation may be associated with the antioxidative
activities of GSPE (17). The
oxidation system has been documented to serve an important role in
the formation of AGEs (8,33,35).
GSPE increases the renal antioxidative ability, decreases NO levels
and decreases the activity of NOS in kidney and serum (16). GSPE decreased RAGE mRNA and protein
expression in diabetic rats in the present study, which also
contributed to the improvement of renal function. It has been
reported that AGE can activate the RAGE gene through NF-κB and AGEs
accumulation (11). Thus, the
action of GSPE to decrease RAGE is associated with the GSPE's
inhibition of AGE formation. The in vitro experiments
suggest at least two possible mechanisms of nephrin alteration in
diabetic conditions, the first mediated by GA-RAGE interaction
involving the inhibition of nephrin gene transcription, and the
second associated with a rapid redistribution and shedding from the
cell surface that depends on Ang II stimulation of the cell
cytoskeleton (35,36). A previous study indicated that GA
inhibited nephrin synthesis through the engagement of the receptor
for AGEs, whereas angiotensin II acted on cytoskeleton
redistribution, inducing the shedding of nephrin in proteinuric
patients with diabetes (37). The
results are consistent with an inhibitory effect of GSPE on the
AGEs/RAGE system, which results in increased nephrin expression and
decreased proteinuria.
In summary, the results demonstrated that GSPE has a
protective effect on kidney in STZ-induced diabetic rats by
decreasing urinary albumin excretion and reversing renal
pathological damage. The underlying mechanism is associated with
the decreased expression of the AGEs/RAGE axis and the increased
expression of nephrin in DN rats. Based on these results, it is
suggested that GSPE may be a promising therapeutic approach for the
prevention and treatment of DN.
Acknowledgements
The authors would like to thank Dr Xueqing Gao from
the Institute of Medical Science (Jinan, China) for the technical
assistance, and Mr David Henry (World International English of
Jinan, USA) for revising the paper.
References
|
1
|
Collins AJ, Kasiske B, Herzog C, Chavers
B, Foley R, Gilbertson D, Grimm R, Liu J, Louis T, Manning W, et
al: United States Renal Data System 2005 Annual Data Report
Abstract. Am J Kidney Dis. 57:A82011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang Y, Xiao HQ, Wang Y, Yang ZS, Dai LJ
and Xu YC: Differential expression and therapeutic efficacy of
microRNA-346 in diabetic nephropathy mice. Exp Ther Med.
10:106–112. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bhatt K, Lanting LL, Jia Y, Yadav S, Reddy
MA, Magilnick N, Boldin M and Natarajan R: Anti-inflammatory role
of microRNA-146a in the pathogenesis of diabetic nephropathy. J Am
Soc Nephrol. 27:2277–2288. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wolf G and Ziyadeh FN: Cellular and
molecular mechanisms of proteinuria in diabetic nephropathy.
Nephron Physiol. 106:p26–p31. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim JJ, Li JJ, Jung DS, Kwak SJ, Ryu DR,
Yoo TH, Han SH, Choi HY, Kim HJ, Han DS and Kang SW: Differential
expression of nephrin according to glomerular size in early
diabetic kidney disease. J Am Soc Nephrol. 18:2303–2310. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Welsh GI and Saleem MA: Nephrin-signature
molecule of the glomerular podocyte? J Pathol. 220:328–337.
2009.
|
|
7
|
Daroux M, Prévost G, Maillard-Lefebvre H,
Gaxatte C, D'Agati VD, Schmidt AM and Boulanger E: Advanced
glycation end-products: Implications for diabetic and non-diabetic
nephropathies. Diabetes Metab. 36:1–10. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fukami K, Yamagishi S, Ueda S and Okuda S:
Role of AGEs in diabetic nephropathy. Curr Pharm Des. 14:946–952.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Park CH, Noh JS, Fujii H, Roh SS, Song YO,
Choi JS, Chung HY and Yokozawa T: Oligonol, a low-molecular-weight
polyphenol derived from lychee fruit, attenuates
gluco-lipotoxicity-mediated renal disorder in type 2 diabetic db/db
mice. Drug Discov Ther. 9:13–22. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Suzuki D, Toyoda M, Yamamoto N, Miyauchi
M, Katoh M, Kimura M, Maruyama M, Honma M, Umezono T and Yagame M:
Relationship between the expression of advanced glycation
end-products (AGE) and the receptor for AGE (RAGE) mRNA in diabetic
nephropathy. Intern Med. 45:435–441. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ramasamy R, Yan SF and Schmidt AM:
Receptor for AGE (RAGE): Signaling mechanisms in the pathogenesis
of diabetes and its complications. Ann N Y Acad Sci. 1243:88–102.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yamagishi S: Role of advanced glycation
end products (AGEs) and receptor for AGEs (RAGE) in vascular damage
in diabetes. Exp Gerontol. 46:217–224. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kaida Y, Fukami K, Matsui T, Higashimoto
Y, Nishino Y, Obara N, Nakayama Y, Ando R, Toyonaga M, Ueda S, et
al: DNA aptamer raised against AGEs blocks the progression of
experimental diabetic nephropathy. Diabetes. 62:3241–3250. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Thallas-Bonke V, Coughlan MT, Tan AL,
Harcourt BE, Morgan PE, Davies MJ, Bach LA, Cooper ME and Forbes
JM: Targeting the AGE-RAGE axis improves renal function in the
context of a healthy diet low in advanced glycation end-product
content. Nephrology (Carlton). 18:47–56. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhai O, Zhong N, Gao HQ, Li BY and Jiang
B: Grape seed proanthocyanidins extracts promote apolipoprotein A-I
mRNA expression in HepG2 cells under experimental sugar and
high-sugar conditions. Eur Rev Med Pharmacol Sci. 16:299–304.
2012.PubMed/NCBI
|
|
16
|
Okudan N, Barışkaner H, Gökbel H, Sahin
AS, Belviranlı M and Baysal H: The effect of supplementation of
grape seed proanthocyanidin extract on vascular dysfunction in
experimental diabetes. J Med Food. 14:1298–1302. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mansouri E, Panahi M, Ghaffari MA and
Ghorbani A: Effects of grape seed proanthocyanidin extract on
oxidative stress induced by diabetes in rat kidney. Iran Biomed J.
15:100–106. 2011.PubMed/NCBI
|
|
18
|
Liang Y, Qiu J, Gao HQ and Li BY:
Protective effect of grape seed proanthocyanidins extracts on
reperfusion arrhythmia in rabbits. J Nutr Sci Vitaminol (Tokyo).
55:223–230. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Makita Z, Vlassara H, Cerami A and Bucala
R: Immunochemical detection of advanced glycosylation end products
in vivo. J Biol Chem. 267:5133–5138. 1992.PubMed/NCBI
|
|
20
|
Lahdenkari AT, Lounatmaa K, Patrakka J,
Holmberg C, Wartiovaara J, Kestilä M, Koskimies O and Jalanko H:
Podocytes are firmly attached to glomerular basement membrane in
kidneys with heavy proteinuria. J Am Soc Nephrol. 15:2611–2618.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Koop K, Eikmans M, Baelde HJ, Kawachi H,
De Heer E, Paul LC and Bruijn JA: Expression of podocyte-associated
molecules in acquired human kidney diseases. J Am Soc Nephrol.
14:2063–2071. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Osawa G, Kimmelstiel P and Seiling V:
Thickness of glomerular basement membranes. Am J Clin Pathol.
45:7–20. 1966. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lam S, Van der Geest RN, Verhagen NA, van
Nieuwenhoven FA, Blom IE, Aten J, Goldschmeding R, Daha MR and van
Kooten C: Connective tissue growth factor and igf-I are produced by
human renal fibroblasts and cooperate in the induction of collagen
production by high glucose. Diabetes. 52:2975–2983. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yamamoto Y, Kato I, Doi T, Yonekura H,
Ohashi S, Takeuchi M, Watanabe T, Yamagishi S, Sakurai S, Takasawa
S, et al: Development and prevention of advanced diabetic
nephropathy in RAGE-overexpressing mice. J Clin Invest.
108:261–268. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Toyoda M, Suzuki D, Umezono T, Uehara G,
Maruyama M, Honma M, Sakai T and Sakai H: Expression of human
nephrin mRNA in diabetic nephropathy. Nephrol Dial Transplant.
19:380–385. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Reddy GR, Kotlyarevska K, Ransom RF and
Menon RK: The podocyte and diabetes mellitus: Is the podocyte the
key to the origins of diabetic nephropathy? Curr Opin Nephrol
Hypertens. 17:32–36. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee EY, Kim GT, Hyun M, Kim S, Seok S,
Choi R, Lee MY and Chung CH: Peroxisome proliferator-activated
receptor-δ activation ameliorates albuminuria by preventing nephrin
loss and restoring podocyte integrity in type 2 diabetes. Nephrol
Dial Transplant. 27:4069–4079. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kobayashi R, Kamiie J, Yasuno K, Ogihara K
and Shirota K: Expression of nephrin, podocin, α-actinin-4 and
α3-integrin in canine renal glomeruli. J Comp Pathol. 145:220–225.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hussain S, Romio L, Saleem M, Mathieson P,
Serrano M, Moscat J, Diaz-Meco M, Scambler P and Koziell A: Nephrin
deficiency activates NF-kappaB and promotes glomerular injury. J Am
Soc Nephrol. 20:1733–1743. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Berg UB, Torbjörnsdotter TB, Jaremko G and
Thalme B: Kidney morphological changes in relation to long-term
renal function and metabolic control in adolescents with IDDM.
Diabetologia. 41:1047–1056. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mifsud SA, Allen TJ, Bertram JF, Hulthen
UL, Kelly DJ, Cooper ME, Wilkinson-Berka JL and Gilbert RE:
Podocyte foot process broadening in experimental diabetic
nephropathy: Amelioration with renin-angiotensin blockade.
Diabetologia. 44:878–882. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Schermer B and Benzing T: Lipid-protein
interactions along the slit diaphragm of podocytes. J Am Soc
Nephrol. 20:473–478. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tan AL, Forbes JM and Cooper ME: AGE, RAGE
and ROS in diabetic nephropathy. Semin Nephrol. 27:130–143. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Myint KM, Yamamoto Y, Doi T, Kato I,
Harashima A, Yonekura H, Watanabe T, Shinohara H, Takeuchi M,
Tsuneyama K, et al: RAGE control of diabetic nephropathy in a mouse
model: Effects of RAGE gene disruption and administration of
low-molecular weight heparin. Diabetes. 55:2510–2522. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Doublier S, Salvidio G, Lupia E,
Ruotsalainen V, Verzola D, Deferrari G and Camussi G: Nephrin
expression is reduced in human diabetic nephropathy: Evidence for a
distinct role for glycated albumin and angiotensin II. Diabetes.
52:1023–1030. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Miyata T and Izuhara Y: Inhibition of
advanced glycation end products: An implicit goal in clinical
medicine for the treatment of diabetic nephropathy? Ann N Y Acad
Sci. 1126:141–146. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cohen MP, Chen S, Ziyadeh FN, Shea E, Hud
EA, Lautenslager GT and Shearman CW: Evidence linking glycated
albumin to altered glomerular nephrin and VEGF expression,
proteinuria and diabetic nephropathy. Kidney Int. 68:1554–1561.
2005. View Article : Google Scholar : PubMed/NCBI
|