Introduction
Salivary adenoid cystic carcinoma (SACC) is a
malignant tumor that occurs in the large and small salivary glands,
accounting for ~7.5–10% of all salivary malignancies (1). SACC is characterized by its unique
biological features, including slow but aggressive growth, high
incidence of distant metastasis and a high recurrence rate
(2–4). Molecular alterations or biomarkers
associated with the development and progression of SACC, in
addition to the therapeutic efficacy, are poorly characterized
(5). The mortality rate associated
with SACC remains high and lung metastasis is the major cause of
death (6). As SACC is not
sensitive to radiotherapy and chemotherapy, and recurrence is
common following surgery, the discovery of appropriate therapeutic
targets for the treatment of SACC is required.
MicroRNAs (miRNAs/miRs) belong to a class of
non-coding single-stranded RNA molecules that have a length of ~22
nucleotides and are encoded by endogenous genes (7). miRNAs participate in the regulation
of numerous biological processes by binding to the base sequence of
the 3′-untranslated region in target mRNA, which leads to mRNA
degradation or translational repression (8). miRNAs are considered to be important
regulators of diverse cellular processes. In addition, they may act
as oncogenes or anti-oncogenes by altering various cellular
pathways (9). Certain miRNAs are
abnormally expressed in various cancer types compared with the
corresponding normal tissues, including head and neck cancer,
breast, lung, colon and stomach cancer, and leukemia and lymphoma
(10–12).
Of the various miRNAs that are reported to be
involved in cancer development, miR-21 has recently been reported
to be overexpressed in SACC (13).
In addition, miR-21 upregulation has been observed in various types
of malignant tumors compared with their adjacent normal tissue, and
accumulating evidence indicates that it may be involved in the
regulation of the initiation and development of cancer, which
involves cell proliferation, invasion, migration and apoptosis
(14,15). Furthermore, miR-21 acts as an
important regulator in several signaling pathways, including
RAS/MEK/extracellular signal-regulated kinase, Wnt/β-catenin and
phosphatase and tensin homolog deleted on chromosome ten
(PTEN)/phosphatidylinositol 3-kinase/AKT, or by targeting
programmed cell death 4 (PDCD4), PTEN and TIMP metallopeptidase
inhibitor 3 (16,17). However, the role of miR-21 in SACC
and the mechanisms underlying its regulation of this malignant
tumor remain unclear.
Therefore, the current study aimed to elucidate the
role of miR-21 in the pathological and physiological processes of
SACC, and to investigate the underlying mechanisms. Small
interfering (si)RNA was employed to reduce miR-21 expression in
SACC cells to investigate its role in proliferation, apoptosis,
migration and invasion.
Materials and methods
Cell culture and transfection
Human salivary adenoid cystic carcinoma cell lines
with high metastatic potential (SACC-LM) and low metastatic
potential (SACC-83) were provided by the Laboratory of Oral and
Maxillofacial Surgery, Ninth People's Hospital, School of Medicine,
Shanghai Jiao Tong University. Cells were cultured in RPMI-1640
(Hyclone; GE Healthcare Life Sciences, Logan, UT, USA) culture
medium containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), 100 U/ml penicillin and 100
µg/ml streptomycin at 37°C in a humidified incubator with 5%
CO2. Upon 70–80% confluence, using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) as transfection medium, miR-21 inhibitor/siRNA
(Shanghai GenePharma Co., Ltd., Shanghai, China) or negative
control siRNA (Shanghai GenePharma Co., Ltd.) were transfected into
SACC-LM cells, according to the manufacturer's protocol. The
sequences of miR-21 siRNA and negative control siRNA were
5′-UCAACAUCAGUCUGAUAAGCUA-3′ and 5′-CAGUACUUUUGUGUAGUACAA-3′,
respectively. Non-transfected cells were used as a blank control.
The final concentration of siRNA was 60 nM. At 5 h after
transfection, the culture medium was replaced with fresh RPMI-1640
containing 10% FBS and transfection efficiency was determined using
fluorescent images.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
miRanda (http://34.236.212.39/microrna/home.do), miRBase
(http://www.mirbase.org/) and TargetScan (release
7.1) were searched to predicted the target genes of miR-21. At 48 h
after transfection, total RNA and miRNA were extracted using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). cDNA was
obtained using HiScript Reverse Transcriptase (Vazyme Biotech Co.,
Ltd., Nanjing, China) including 3.575 µg RNA, 2 µl Oligo (dT) 15 or
2 µl hsa-miR-21-5p loop, 4 µl dNTP, 4 µl Hiscript Buffer, 1 µl
Reverse Transcriptase and 0.5 µl RNase inhibitor, according to the
following protocol: 25°C for 5 min, 50°C for 15 sec, 85°C for 5 min
and 4°C for 10 min. mRNA levels of miR-21, PDCD4, PTEN and B-cell
lymphoma (Bcl)-2 were detected by qPCR using a SYBR Premix Ex Taq
II kit (Takara Bio, Inc., Otsu, Japan) on a 7900HT Fast Real-Time
PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc.).
Based on the sequence of miR-21, a stem-loop primer was designed by
Primer Premier 5.0 (Premier Biosoft International, Palo Alto, CA,
USA) as follows:
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCAACATC-3′,
5′-TGCGCTAGCTTATCAGACTGA-3′ (forward), 5′-CCAGTGCAGGGTCCGAGGTATT-3′
(reverse). U6 small nuclear RNA was used as a reference gene, with
the primer 5′-CGCTTCGGCAGCACATATAC-3′ (forward);
5′-AAATATGGAACGCTTCACGA-3′ (reverse). The following other primer
sequences were used: PDCD4, 5′-AACCAGTCC-AAAGGGAAGG-3′ (forward)
and 5′-ACATCCACCTCCTCCACATC-3′ (reverse); PTEN,
5′-CACGACGGGAAGACAAGTTC-3′ (forward) and 5′-TCTGCACGCTCTATACTGCA-3′
(reverse); and Bcl-2, 5′-AGCCTGAGAGCAACCCAAT-3′ (forward) and
5′-AGCGACGAGAGAAGTCATCC-3′ (reverse). GAPDH was used as an internal
reference, the sequences were 5′-TCAAGAAGGTGGTGAAGCAGG-3′
(forward); 5′-TCAAAGGTGGAGGAGTGGGT-3′ (reverse). The PCR reaction
was performed as follows: 95°C for 30 sec, followed by 40 cycles at
95°C for 5 sec and 60°C for 30 sec. All RT-qPCR experiments were
performed in triplicate and repeated three times. Each gene
expression was quantified using the 2−ΔΔCq method
(18), and their expression was
compared between cells transfected with negative control and miR-21
siRNA.
Cell proliferation and viability
assay
A Cell Counting Kit (CCK)-8 assay was performed to
measure cell proliferation and viability. SACC-LM cells were seeded
at a density of 8×103 cells/well in 96-well plates. When
the cells reached 70–80% confluence, they were transfected with
miR-21 inhibitor or negative control and incubated for 24, 48, 72
or 96 h. Subsequently, 10 µl CCK-8 reagent (Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was added into each well of
the plate and cells were incubated at 37°C for 30 min. Absorbance
at 460 nm was measured using a microplate reader (BioTek
Instruments, Inc., Winooski, VT, USA).
Wound healing assay
A total of 5×105 SACC-LM cells per well
were seeded into 6-well plates and cultured until 80% confluence.
At 48 h after transfection, a wound was created in the center of
the layer of confluent cells adhering to the bottom of the well
using a 100 µl pipette tip. Plates were gently washed three times
with PBS to remove the cell debris, and serum-free RPMI-1640 medium
was added. The scratch width was measured by photo recordings at 0
and 24 h after the wound was created using an inverted optical
microscope at ×100 magnification (Olympus Corporation, Tokyo,
Japan).
Matrigel invasion assay
Cell invasion was detected by a Matrigel invasion
assay. Diluted Matrigel (100 µl; BD Biosciences, Franklin Lakes,
NJ, USA) was added to the upper chamber of each well of a 24-well
Transwell plate (Corning Incorporated, Corning, NY, USA). At 48 h
after transfection, 1×105 SACC-LM cells in 200 µl
serum-free RPMI-1640 medium were seeded in the upper chamber, while
500 µl RPMI-1640 containing 10% FBS was added to the lower chamber.
After 24 h incubation at 37°C, residual cells in the upper chamber
were carefully removed with a cotton swab, while cells that had
invaded to the lower chamber were fixed using 4% paraformaldehyde
for 20 min and stained with 0.1% crystal violet for 15 min at room
temperature. Cells on the bottom surface were quantified by
counting five random fields per well under the light microscope at
a magnification of ×100 using Image-Pro Plus version 6.0 (Media
Cybernetics, Inc., Rockville, MD, USA), and the mean number of
cells passing through the chamber calculated.
Flow cytometry
At 48 h after transfection, transfected and
non-transfected SACC-LM cells were harvested by trypsinization and
washed twice in cold PBS. Subsequently, 1.0×105 cells
were suspended in 500 µl binding buffer containing 5 µl annexin
V-FITC (Nanjing KeyGen Biotech Co., Ltd.) and 5 µl propidium iodide
(PI; all from Nanjing KeyGen Biotech Co., Ltd., Nanjing, China),
gently mixed with a pipette and incubated at room temperature for
15 min. The apoptotic rate was obtained from the percentage of
cells that were double-stained with annexin V-FITC and PI, as
detected by a flow cytometer (Facs Canto II; BD Biosciences) using
BD FACSDiva version 8.0.1 software.
Western blot analysis
Then, 48 h after transfection, non-transfected and
transfected SACC-LM cells were trypsinized, collected and
centrifuged at 671 × g for 5 min at room temperature. Total protein
was extracted using protein lysis solution containing RIPA buffer
and 1% phenyl methane sulfonyl fluoride (PMSF; Beyotime Institute
of Biotechnology, Jiangsu, China). Protein concentrations were
measured using a BCA Protein assay kit (Beyotime Institute of
Biotechnology). Denatured protein samples (40 µg) were separated by
10% SDS-PAGE and subsequently transferred to polyvinylidene
fluoride membranes. After blocking in 5% nonfat dry milk for 2 h at
room temperature, the corresponding membranes were incubated with
antibodies against PDCD4 (ab80590; 1:7,000; Abcam, Cambridge, UK),
Bcl-2 (ab32124; 1:1,000; Abcam), PTEN (ab32199; 1:1,000; Abcam),
Snail family transcriptional repressor 1 (Snail1; BZ06198; 1:500;
Bioworld Technology, Inc., St. Louis Park, MN, USA) and E-cadherin
(BS1098; 1:500; Bioworld Technology, Inc.) at 4°C overnight. GAPDH
(BZ00672; 1:7,000; Bioworld Technology, Inc.) was used as an
internal control. Subsequently, the membranes were incubated with
goat anti-rabbit polyclonal IgG secondary antibodies (ZB-2010;
1:500; OriGene Technologies, Inc., Beijing, China) for 2 h at room
temperature. Membranes were washed in washing buffer three times
and PDCD4, PTEN, Bcl-2, Snail1, E-cadherin and GAPDH protein bands
were visualized using a BCIP/NBT Alkaline Phosphatase Color
Development kit (P0321; Beyotime Institute of Biotechnology).
Western blot results were normalized to the expression of GAPDH and
analyzed using ImageJ version 1.49 software (National Institutes of
Health, Bethesda, MD, USA).
Statistical analysis
Data were analyzed using SPSS statistical software
version 16.0 (SPSS, Inc., Chicago, IL, USA). Results are presented
as the mean ± standard deviation. Comparisons between two groups
were performed using Student's t-test, while comparisons among
multiple groups were performed using one-way analysis of variance
followed by LSD test. P<0.05 was considered to indicate a
statistically significant difference.
Results
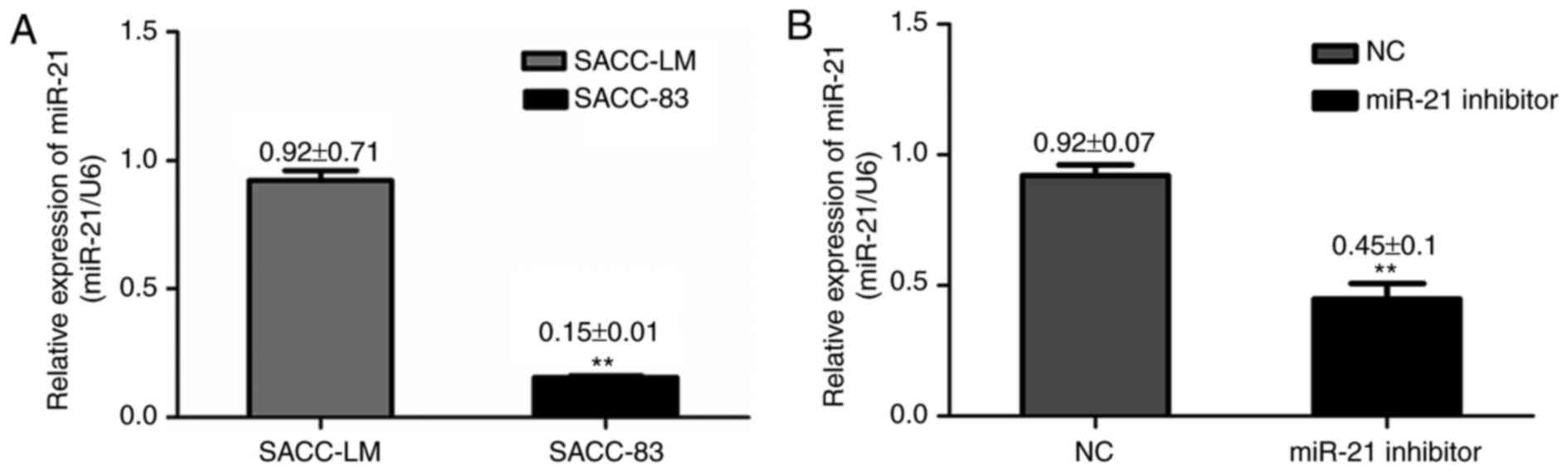
miR-21 is overexpressed in SACC cell
lines and may be associated with tumor metastasis
Previous reports have indicated that miR-21
expression in SACC tumor tissue samples was significantly higher
compared with normal tissue samples (13). In order to investigate the
potential role of miR-21 in SACC, the present study measured miR-21
expression in two SACC cell lines (SACC-83 and SACC-LM). RT-qPCR
results demonstrated that miR-21 expression was significantly
higher in SACC-LM cells, which have a high metastatic potential,
compared with SACC-83 cells, which have a low metastatic potential
(Fig. 1A), indicating a potential
role of miR-21 in SACC cells metastatic ability.
miR-21 downregulation inhibits tumor
proliferation and induces apoptosis
To investigate the effect of miR-21 on cell growth,
cells were transfected with miR-21 siRNA or negative control siRNA.
RT-qPCR analysis revealed that miR-21 expression was reduced in
SACC-LM cells transfected with miR-21 siRNA, compared with cells
transfected with negative control siRNA (Fig. 1B). Flow cytometry and CCK-8 assays
were performed to determine the effect of miR-21 silencing on cell
apoptosis and proliferation in SACC-LM cells, respectively. CCK-8
detection was performed at five time points (0, 24, 48, 72 and 96
h). The apoptotic rate in the miR-21 siRNA, negative control siRNA
and blank control groups was 14.5±0.6, 8.43±0.25 and 8.5±1.249%,
respectively. In addition, miR-21 siRNA-transfected cells exhibited
a significant increase in annexin V staining compared with the two
other groups (P<0.01; Fig. 2A and
B). As demonstrated in Fig.
2C, miR-21 silencing triggered a decreased cell viability
(P<0.05) compared with cells transfected with negative control
siRNA, and viability was decreased in a time-dependent manner.
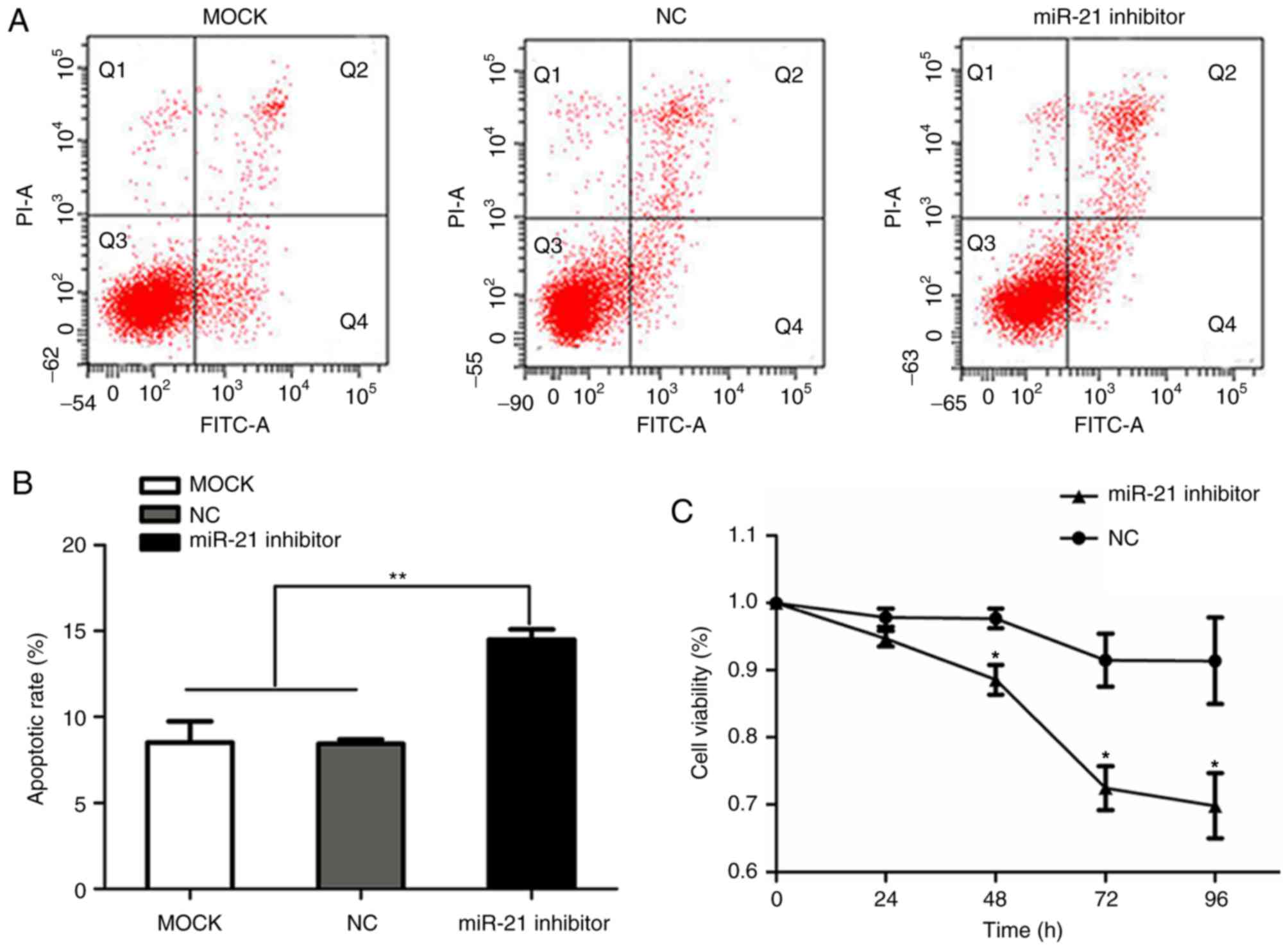 | Figure 2.Effect of miR-21 siRNA on SACC-LM
proliferation and apoptosis. (A) Representative flow cytometry
plots for blank control cells, and cells transfected with NC and
miR-21 siRNA. Cell apoptosis was measured by annexin V-FITC/PI
staining and flow cytometry (apoptotic cells refer to the cells in
Q2 and Q4 quadrants). (B) miR-21 siRNA transfection significantly
increased SACC-LM apoptosis compared with the control groups.
**P<0.01, as indicated. (C) Proliferation of SACC-LM cells at 0,
24, 48, 72 and 96 h after transfection with NC or miR-21 siRNA was
measured by a Cell Counting Kit-8 assay. *P<0.05, as indicated.
miR, microRNA; siRNA, small interfering RNA; SACC, salivary adenoid
cystic carcinoma; NC, negative control; FITC, fluorescein
isothiocyanate; PI, propidium iodide; MOCK, blank control; miR-21
inhibitor, siRNA targeting miR-21. |
miR-21 promotes SACC migration and
invasion
Wound healing migration and Matrigel invasion assays
were performed to investigate the effects of miR-21 on cell
migration and invasion, respectively. Based on the wound healing
assay results, transfection with miR-21 siRNA significantly reduced
the migratory ability of SACC-LM cells, compared with cells
transfected with negative control siRNA (Fig. 3A and B). In the Matrigel invasion
assay, miR-21 silencing in SACC-LM cells reduced the number of
cells crossing the membrane, compared with the negative control
siRNA group, with means of 675 and 269 cells per random microscopic
field, respectively (Fig. 3C and
D). Furthermore, the protein expression of
metastasis-associated markers was determined by western blot
analysis, and the results demonstrated that Snail1 protein
expression was decreased, while E-cadherin protein expression was
increased, in the miR-21 siRNA group compared with blank control
and negative control siRNA groups (Fig. 3E and F). These results indicate
that miR-21 may promote cell invasion and migration in SACC-LM
cells, which may be associated with the high potential for
metastasis of this cell line.
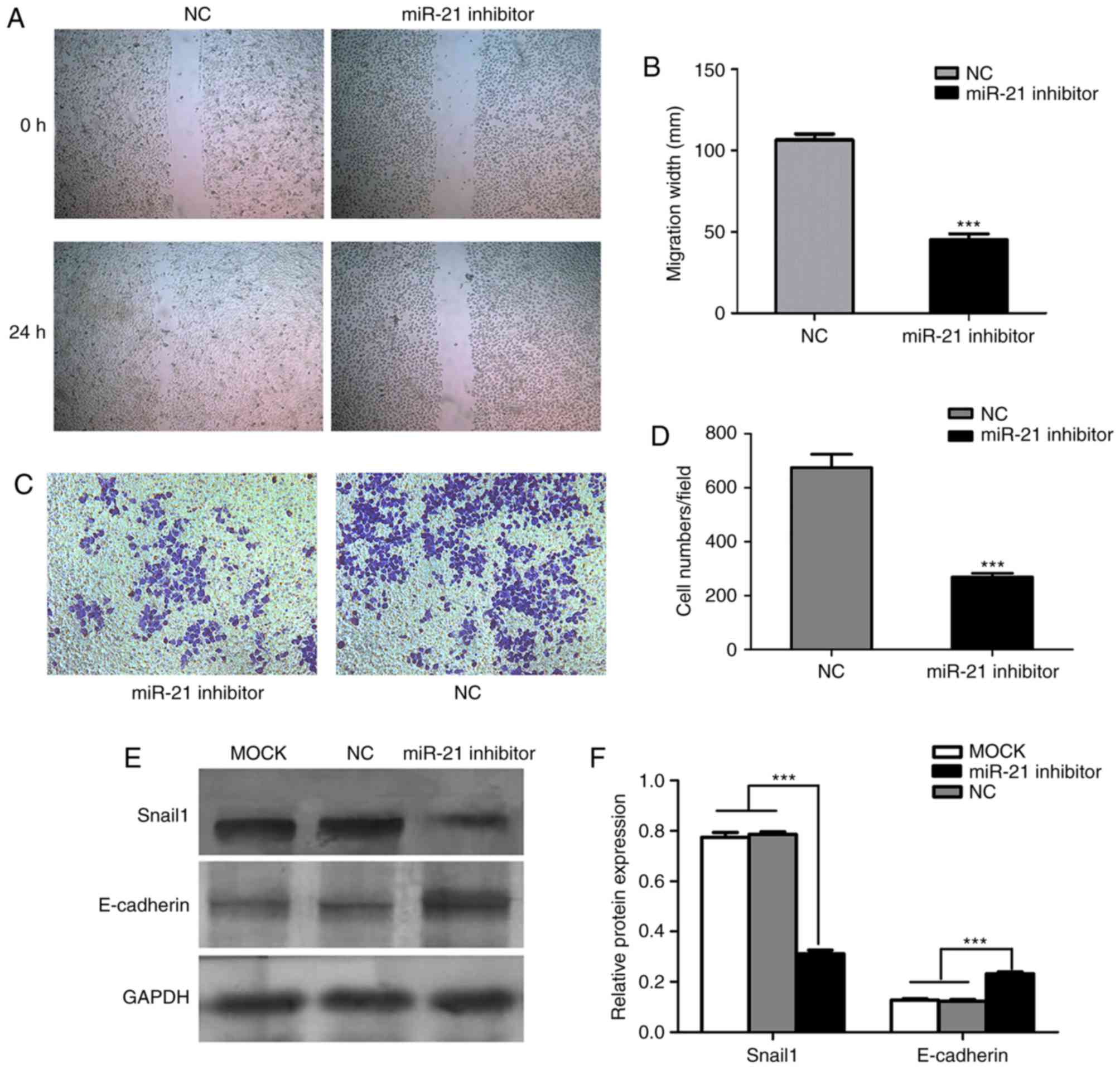 | Figure 3.Effects of miR-21 on SACC-LM migration
and invasion. (A) SACC-LM cell migration was measured by a wound
healing assay following transfection with NC or miR-21 siRNA.
Magnification, ×100. (B) Quantification of SACC-LM cell migration
at 24 h after the wound was created in NC and miR-21
siRNA-transfected cells. ***P<0.001 vs. NC group. (C) Cell
invasion was measured by a Matrigel invasion assay. Magnification,
×100. (D) Quantification of NC and miR-21 siRNA-transfected SACC-LM
cells crossing the Matrigel-coated membrane following incubation
for 24 h. ***P<0.001 vs. NC group. (E) Western blotting was
performed to investigate Snail1 and E-cadherin protein expression
in blank control, and NC and miR-21 siRNA-transfected SACC-LM
cells. Representative bands are presented. (F) Snail1 protein
expression was significantly decreased, while E-cadherin protein
expression was increased, following transfection of SACC-LM cells
with miR-21 siRNA, compared with the control groups. Protein
expression was normalized to GAPDH expression. ***P<0.001, as
indicated. miR, microRNA; SACC, salivary adenoid cystic carcinoma;
NC, negative control; siRNA, small interfering RNA; Snail1, snail
family transcriptional repressor 1; miR-21 inhibitor, siRNA
targeting miR-21; MOCK, blank control. |
PDCD4, PTEN and Bcl-2 are target genes
of miR-21 in SACC
To identify potential target genes of miR-21,
several databases were employed, including miRanda, miRBase and
TargetScan. The search results indicated that PDCD4, PTEN and Bcl-2
are predicted miR-21 target genes. Therefore, RT-qPCR was performed
to further verify the accuracy of the above result. The results
demonstrated that PDCD4 and PTEN mRNA levels were upregulated,
while Bcl-2 mRNA levels were downregulated, following transfection
of SACC-LM cells with miR-21 siRNA, compared with cells transfected
with negative control siRNA (Fig.
4A). Therefore, these results demonstrated a potential role for
miR-21 in the regulation of the expression of PDCD4, PTEN and
Bcl-2.
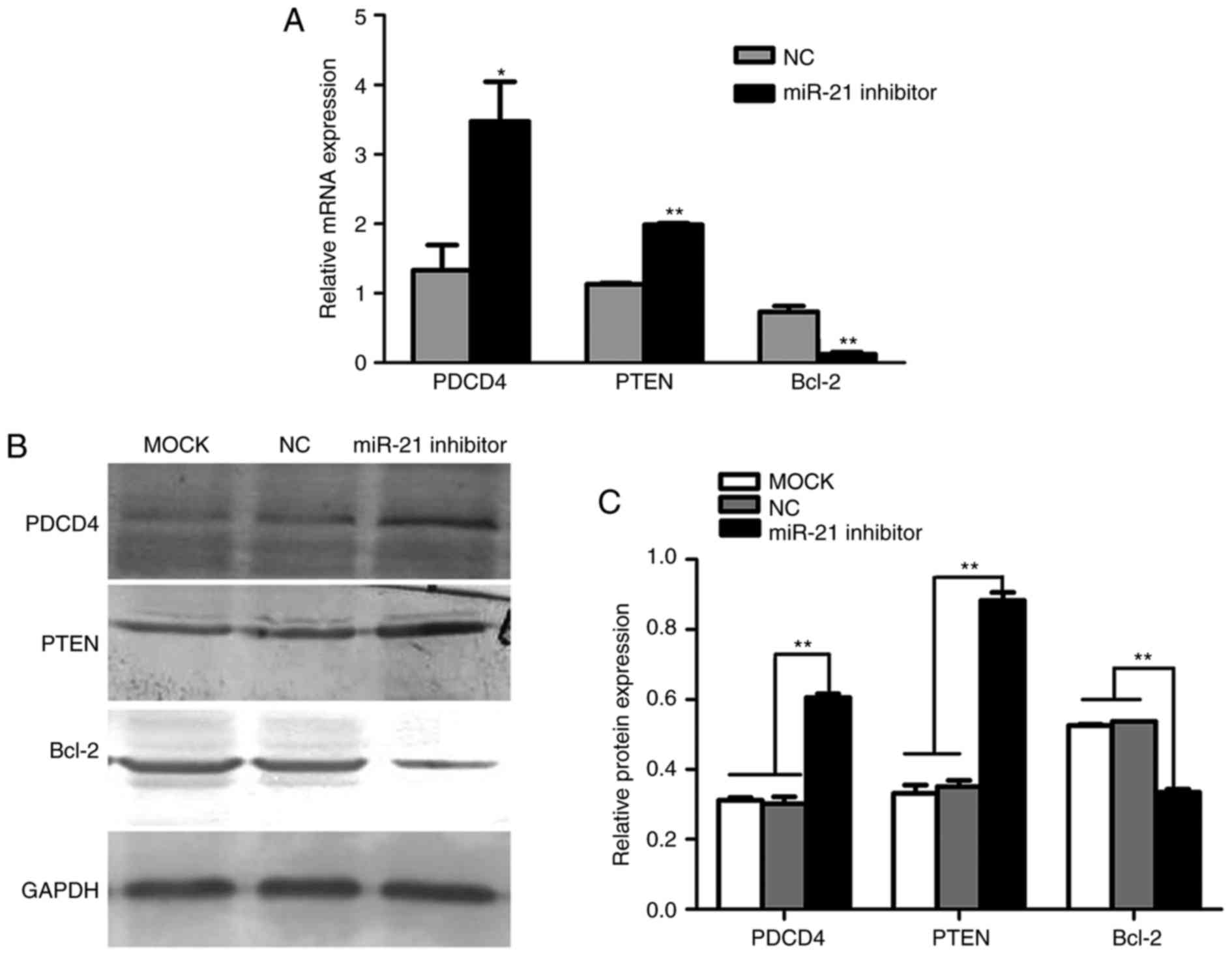 | Figure 4.Effects of miR-21 on PDCD4, PTEN and
Bcl-2 mRNA and protein expression. (A) Reverse
transcription-quantitative polymerase chain reaction results
demonstrated that PDCD4 and PTEN mRNA were significantly increased,
while Bcl-2 mRNA was decreased, when endogenous miR-21 was silenced
using miR-21-targeting siRNA. *P<0.05 and **P<0.01 vs. NC
siRNA group. (B) Protein levels of PDCD4, PTEN and Bcl-2 were
detected by western blotting. Representative bands are presented.
(C) PDCD4 and PTEN protein expression was significantly increased,
while Bcl-2 protein expression was decreased, when endogenous
miR-21 was silenced using miR-21 siRNA in SACC-LM cells. Protein
expression was normalized to GAPDH protein expression. **P<0.01,
as indicated. miR, microRNA; PDCD4, programmed cell death 4; PTEN,
phosphatase and tensin homolog deleted on chromosome ten; Bcl-2,
B-cell lymphoma-2; siRNA, small interfering RNA; SACC, salivary
adenoid cystic carcinoma; NC, negative control; miR-21 inhibitor,
siRNA targeting miR-21; MOCK, blank control. |
miR-21 regulates tumor growth and
metastasis by targeting PDCD4, PTEN and Bcl-2 in SACC
As PDCD4, PTEN and Bcl-2 are involved in migration,
invasion and apoptosis and they are miR-21 target genes, western
blot analysis was performed to further determine whether miR-21
regulates the protein expression of these genes in SACC-LM cells.
The results demonstrated that transfection of SACC-LM cells with
miR-21 siRNA led to a significant increase in PDCD4 and PTEN
protein expression, and markedly reduced Bcl-2 protein expression,
compared with blank control and negative control siRNA groups
(Fig. 4B and C). These results
indicate that miR-21 levels were negatively associated with PDCD4
and PTEN expression, and positively associated with Bcl-2
expression. Therefore, miR-21 may reduce apoptosis, and increase
cell proliferation and metastasis, by regulating the expression of
PDCD4, PTEN and Bcl-2 in SACC.
Discussion
SACC is a common malignant tumor of the head and
neck that has a distinct potential for invasion and distant
migration. At present, few studies have investigated the molecular
markers that are associated with SACC treatment and prognosis
(19,20). In recent years, an association
between various miRNAs and tumor development has been established
and substantial progress has been made concerning the biological
function of miRNAs and their clinical relevance in cancer (21–24)
Previous studies employed miRNA microarrays and RT-qPCR to
determine miRNA expression profiles in SACC cell lines with
different metastatic potentials (13,25).
These reports demonstrated that miR-4487, miR-4430, miR-486-3p,
miR-5191, miR-21 and miR-18a may be involved in the progression of
SACC. Notably, miR-21 is a key factor in multiple
malignancy-associated processes; it triggers the inflammatory
response leading to tumor growth and metastasis, and promotes the
interaction between tumor cells and the microenvironment, therefore
promoting tumor occurrence and development (26–28).
However, the underlying molecular mechanism of miR-21 action
requires further investigation.
In the present study, miR-21 expression was
significantly upregulated in the SACC-LM cell line (high metastatic
potential) compared with the SACC-83 cell line (low metastatic
potential), which indicated that the SACC-LM cell line is more
suitable for the investigation of miR-21. Downregulation of miR-21
expression using siRNA led to significantly reduced cell migration
and invasion, and increased apoptosis, while cell viability was
inhibited to a certain extent, compared with negative control
siRNA-transfected cells. Overall, these results indicated that
miR-21 may have various roles in tumor development and may be
considered an important oncomiR in SACC.
By searching several databases, PDCD4, PTEN and
Bcl-2 were considered to be potential targets of miR-21. PDCD4 is a
novel tumor suppressor gene that was recently discovered and has
been reported to be involved in the regulation of tumor development
and metastasis (29). PDCD4 is
frequently downregulated in various tumor tissues and cells, which
is associated with increased cell proliferation, migration and
invasion, and decreased apoptosis (30,31).
Previous studies have demonstrated that PDCD4 expression was
negatively associated with miR-21 levels in gastric cancer
(32,33). Bcl-2, as an anti-apoptotic gene, is
able to repress various apoptotic death events. A previous report
demonstrated that miR-21 inhibited apoptosis through regulation of
Bcl-2 in a breast cancer mouse model (34), while Shi et al (35) reported that miR-21 overexpression
upregulated Bcl-2 expression in human glioblastoma U-87MG cells.
The results of the present study demonstrated that transfection of
SACC-LM cells with miR-21 siRNA resulted in a higher apoptotic
rate, which may be due to the downregulation of Bcl-2 and
upregulation of PDCD4 at mRNA and protein levels. Therefore, a
negative association was observed between miR-21 and PDCD4 protein
levels in SACC-LM cells, while a positive association was observed
between miR-21 and Bcl-2 expression. These results indicate that
miR-21 may suppress cell apoptosis by regulating the expression of
PDCD4 and Bcl-2.
PTEN, which is an established tumor suppressor gene,
has important roles in various tumor processes, including cell
growth, apoptosis, adhesion, migration and infiltration (36,37).
Genetic inactivation of PTEN is frequently observed in
glioblastoma, endometrial cancer and prostate cancer, and reduced
PTEN expression has been reported in various other cancer types,
including lung and breast cancer (37–39).
Liu et al (40) reported
that the expression of PTEN was reduced in >80% of solid SACC
cases, and PTEN was also reported to be closely associated with the
prognosis and lymph node metastasis of salivary gland cancer
(41). In the current study,
miR-21 downregulation led to increased PTEN expression, and PTEN
protein expression was negatively associated with cell migration
and invasion in SACC-LM cells. Epithelial-mesenchymal transition
(EMT) is a basic process of embryonic development that involves the
separation and migration of epithelial cells from epithelial tissue
to other locations, and is the foundation for normal development,
wound healing and malignant epithelial tumors (42). Among various EMT regulatory
factors, Snail1 is considered to be a key factor in tumor
metastasis associated with EMT by directly repressing the
expression of the epithelial cell marker, E-cadherin (43). In the present study, western blot
analysis confirmed that downregulation of miR-21 resulted in
decreased protein expression of Snail1, which may therefore lead to
reduced cell migration and invasion, in SACC-LM cells. Furthermore,
E-cadherin protein levels were significantly increased following
miR-21 silencing, compared with the control groups. These data
indicate that miR-21 may control PTEN and Snail1 protein expression
to further regulate the metastasis and invasion of SACC cells.
In conclusion, the results of the current study
indicated that miR-21 may be considered as a potential prognostic
biomarker for SACC and may have potential as a therapeutic agent
for the treatment of SACC.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 31060167 and
81160242), the ‘Qing Lan Project’ of the Jiangsu Higher Education
Institutions Young and Middle-aged Academic Leaders Funded Project
(grant no. 51934) and the Outstanding Talent Fund Project of Xuzhou
Medical College (grant no. 2013012). The authors thank the
Laboratory of Oral and Maxillofacial Surgery, Ninth People's
Hospital, School of Medicine, Shanghai Jiao Tong University
(Shanghai, China) for providing the SACC-83 and SACC-LM cell
lines.
References
|
1
|
Liu X, Zhang W, Guo H, Yue J and Zhuo S:
miR-98 functions as a tumor suppressor in salivary adenoid cystic
carcinomas. Onco Targets Ther. 9:1777–1786. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tang Y, Liang X, Zheng M, Zhu Z, Zhu G,
Yang J and Chen Y: Expression of c-kit and Slug correlates with
invasion and metastasis of salivary adenoid cystic carcinoma. Oral
Oncol. 46:311–316. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
He JF, Ge MH, Zhu X, Chen C, Tan Z, Li YN
and Gu ZY: Expression of RUNX3 in salivary adenoid cystic
carcinoma: Implications for tumor progression and prognosis. Cancer
Sci. 99:1334–1340. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hu K, Gan YH, Li SL, Gao Y, Wu DC, Wang CY
and Yu GY: Relationship of activated extracellular signal-regulated
kinase 1/2 with lung metastasis in salivary adenoid cystic
carcinoma. Oncology Rep. 21:137–143. 2009.
|
|
5
|
Qian X, Kaufmann AM, Chen C, Tzamalis G,
Hofmann VM, Keilholz U, Hummel M and Albers AE: Prevalence and
associated survival of high-risk HPV-related adenoid cystic
carcinoma of the salivary glands. Int J Oncol. 49:803–811. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li S, Zhang X, Zhou Z and Huang Z, Liu L
and Huang Z: Downregulation of nucleophosmin expression inhibited
proliferation and induced apoptosis in salivary gland adenoid
cystic carcinoma. J Oral Pathol Med. 46:175–181. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sun L, Liu B, Lin Z, Yao Y, Chen Y, Li Y,
Chen J, Yu D, Tang Z, Wang B, et al: miR-320a acts as a prognostic
factor and Inhibits metastasis of salivary adenoid cystic carcinoma
by targeting ITGB3. Mol Cancer. 14:962015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dong YL, Zhou L, Li YL, Xiao K and Weng
XS: Establishment and assessment of rat models of
glucocorticoid-induced osteonecrosis. Zhongguo Yi Xue Ke Xue Yuan
Xue Bao. 37:152–156. 2015.PubMed/NCBI
|
|
9
|
Mirzaei H, Gholamin S, Shahidsales S,
Sahebkar A, Jaafari MR, Mirzaei HR, Hassanian SM and Avan A:
MicroRNAs as potential diagnostic and prognostic biomarkers in
melanoma. Eur J Cancer. 53:25–32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yu F, Yao H, Zhu P, Zhang X, Pan Q, Gong
C, Huang Y, Hu X, Su F, Lieberman J and Song E: let-7 regulates
self renewal and tumorigenicity of breast cancer cells. Cell.
131:1109–1123. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Couzin J: Cancer biology. A new cancer
player takes the stage. Science. 310:766–767. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu MX, Zhou KC and Cao Y: MCRS1
overexpression, which is specifically inhibited by miR-129*,
promotes the epithelial-mesenchymal transition and metastasis in
non-small cell lung cancer. Mol Cancer. 13:2452014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jiang LH, Ge MH, Hou XX, Cao J, Hu SS, Lu
XX, Han J, Wu YC, Liu X, Zhu X, et al: miR-21 regulates tumor
progression through the miR-21-PDCD4-Stat3 pathway in human
salivary adenoid cystic carcinoma. Lab Invest. 95:1398–1408. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Martin del Campo SE, Latchana N, Levine
KM, Grignol VP, Fairchild ET, Jaime-Ramirez AC, Dao TV, Karpa VI,
Carson M, Ganju A, et al: miR-21 enhances melanoma invasiveness via
inhibition of tissue inhibitor of metalloproteinases 3 expression:
In vivo effects of miR-21 inhibitor. PLoS One. 10:e01159192015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Leone E, Morelli E, Di Martino MT, Amodio
N, Foresta U, Gullà A, Rossi M, Neri A, Giordano A, Munshi NC, et
al: Targeting miR-21 inhibits in vitro and in vivo multiple myeloma
cell growth. Clin Cancer Res. 19:2096–2106. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu Y, Kanwar SS, Patel BB, Oh PS, Nautiyal
J, Sarkar FH and Majumdar AP: MicroRNA-21 induces stemness by
downregulating transforming growth factor beta receptor 2 (TGFβR2)
in colon cancer cells. Carcinogenesis. 33:68–76. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang BG, Li JF, Yu BQ, Zhu ZG, Liu BY and
Yan M: microRNA-21 promotes tumor proliferation and invasion in
gastric cancer by targeting PTEN. Oncol Rep. 27:1019–1026. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen W, Liu BY, Zhang X, Zhao XG, Cao G,
Dong Z and Zhang SL: Identification of differentially expressed
genes in salivary adenoid cystic carcinoma cells associated with
metastasis. Arch Med Sci. 12:881–888. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Feng X, Matsuo K, Zhang T, Hu Y, Mays AC,
Browne JD, Zhou X and Sullivan CA: MicroRNA profiling and target
genes related to metastasis of salivary adenoid cystic carcinoma.
Anticancer Res. 37:3473–3481. 2017.PubMed/NCBI
|
|
21
|
Cai L, Chen Q, Fang S, Lian M and Cai M:
MicroRNA-329 inhibits cell proliferation and tumor growth while
facilitates apoptosis via negative regulation of KDM1A in gastric
cancer. J Cell Biochem. Nov 11–2017. View Article : Google Scholar
|
|
22
|
Rammer M, Webersinke G, Haitchi-Petnehazy
S, Maier E, Hackl H, Charoentong P, Malli T, Steinmair M, Petzer AL
and Rumpold H: MicroRNAs and their role for T stage determination
and lymph node metastasis in early colon carcinoma. Clin Exp
Metastasis. Nov 13–2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fu LL, Li CJ, Xu Y, Li LY, Zhou X, Li DD,
Chen SX, Wang FG, Zhang XY and Zheng LW: Role of lncRNAs as novel
biomarkers and therapeutic targets in ovarian cancer. Crit Rev
Eukaryot Gene Expr. 27:183–195. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takahasi K, Iinuma H, Wada K, Minezaki S,
Kawamura S, Kainuma M, Ikede Y, Shibuya M, Miura F and Sano K:
Usefulness of exosome-encapsulated microRNA-451a as a minimally
invasive biomarker for prediction of recurrence and prognosis in
pancreatic ductal adenocarcinoma. J Hepatobiliary Pancreat Sci. Nov
11–2017. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen W, Zhao X, Dong Z, Cao G and Zhang S:
Identification of microRNA profiles in salivary adenoid cystic
carcinoma cells during metastatic progression. Oncol Lett.
7:2029–2034. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shi GH, Ye DW, Yao XD, Zhang SL, Dai B,
Zhang HL, Shen YJ, Zhu Y, Zhu YP, Xiao WJ and Ma CG: Involvement of
microRNA-21 in mediating chemo-resistance to docetaxel in
androgen-independent prostate cancer PC3 cells. Acta Pharmacol Sin.
31:867–873. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alder H, Taccioli C, Chen H, Jiang Y,
Smalley KJ, Fadda P, Ozer HG, Huebner K, Farber JL, Croce CM and
Fong LY: Dysregulation of miR-31 and miR-21 induced by zinc
deficiency promotes esophageal cancer. Carcinogenesis.
33:1736–1744. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Connolly EC, Van Doorslaer K, Rogler LE
and Rogler CE: Overexpression of miR-21 promotes an in vitro
metastatic phenotype by targeting the tumor suppressor RHOB. Mol
Cancer Res. 8:691–700. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liao J, Liu R, Shi YJ, Yin LH and Pu YP:
Exosome-shuttling microRNA-21 promotes cell migration and
invasion-targeting PDCD4 in esophageal cancer. Int J Oncol.
48:2567–2579. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ma G, Zhang H, Dong M, Zheng X, Ozaki I,
Matsuhashi S and Guo K: Downregulation of programmed cell death 4
(PDCD4) in tumorigenesis and progression of human digestive tract
cancers. Tumor Biol. 34:3879–3885. 2013. View Article : Google Scholar
|
|
31
|
Wang D, Guo S, Han SY, Xu N, Guo JY and
Sun Q: Distinct roles of different fragments of PDCD4 in regulating
the metastatic behavior of B16 melanoma cells. Int J Oncol.
42:1725–1733. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Motoyama K, Inoue H, Mimori K, Tanaka F,
Kojima K, Uetake H, Sugihara K and Mori M: Clinicopathological and
prognostic significance of PDCD4 and microRNA-21 in human gastric
cancer. Int J Oncol. 36:1089–1095. 2010.PubMed/NCBI
|
|
33
|
Cao Z, Yoon JH, Nam SW, Lee JY and Park
WS: PDCD4 expression inversely correlated with miR-21 levels in
gastric cancers. J Cancer Res Clin Oncol. 138:611–619. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Asangani IA, Rasheed SA, Nikolova DA,
Leupold JH, Colburn NH, Post S and Allgayer H: MicroRNA-21 (miR-21)
post-transcriptionally downregulates tumor suppressor Pdcd4 and
stimulates invasion, intravasation and metastasis in colorectal
cancer. Oncogene. 27:2128–2136. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shi L, Chen JA, Yang JA, Pan TH, Zhang SG
and Wang ZM: miR-21 protected human glioblastoma U87MG cells from
chemotherapeutic drug temozolomide induced apoptosis by decreasing
Bax/Bcl-2 ratio and caspase-3 activity. Brain Res. 1352:255–264.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sizemore GM, Balakrishnan S, Hammer AM,
Thies KA, Trimboli AJ, Wallace JA, Sizemore ST, Kladney RD, Woelke
SA, Yu L, et al: Stromal PTEN inhibits the expansion of mammary
epithelial stem cells through Jagged-1. Oncogene. 36:2297–2308.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Upton DH, Walters KA, Allavena RE, Jimenez
M, Desai R, Handelsman DJ and Allan CM: Global or granulosa
cell-specific pten mutations in combination with elevated fsh
levels fail to cause ovarian tumours in mice. Horm Cancer.
7:316–326. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Krohn A, Diedler T, Burkhardt L, Mayer PS,
De Silva C, Meyer-Kornblum M, Kötschau D, Tennstedt P, Huang J,
Gerhäuser C, et al: Genomic deletion of PTEN is associated with
tumor progression and early PSA recurrence in ERG fusion-positive
and fusion-negative prostate cancer. Am J Pathol. 181:401–412.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xue X, Liu Y, Wang Y, Meng M, Wang K, Zang
X, Zhao S, Sun X, Cui L, Pan L and Liu S: miR-21 and miR-155
promote non-small cell lung cancer progression by downregulating
SOCS1, SOCS6, and PTEN. Oncotarget. 7:84508–84519. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu H, Du L, Wang R, Wei C, Liu B, Zhu L,
Liu P, Liu Q, Li J, Lu SL and Xiao J: High frequency of loss of
PTEN expression in human solid salivary adenoid cystic carcinoma
and its implication for targeted therapy. Oncotarget.
6:11477–11491. 2015.PubMed/NCBI
|
|
41
|
Ettl T, Gosau M, Brockhoff G,
Schwarz-Furlan S, Agaimy A, Reichert TE, Rohrmeier C, Zenk J and
Iro H: Predictors of cervical lymph node metastasis in salivary
gland cancer. Head Neck. 36:517–523. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang X, Zhang T, Yang K, Zhang M and Wang
K: miR-486-5p suppresses prostate cancer metastasis by targeting
Snail and regulating epithelial-mesenchymal transition. OncoTargets
Ther. 9:6909–6914. 2016. View Article : Google Scholar
|
|
43
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|