Introduction
Parkinson's disease (PD) is the second most
prevalent neurodegenerative disease worldwide after Alzheimer's
disease (1). PD is characterized
by the loss of dopaminergic neurons in the substantia nigra pars
compacta (SNpc) and decreased dopamine levels in the striatum of
the basal ganglia (2). Although
decades of research have seen advancements in the field, the
precise mechanisms underlying the pathogenesis of PD remains to be
fully elucidated (3). However,
studies conducted over the last decades, including age,
epidemiological, environmental toxins, genetic, immune dysfunction
and postmortem studies, have contributed significantly to the
understanding of the PD pathogenesis (4–6).
Understanding these mechanisms may provide us the future
disease-modifying strategies.
In recent decades, peripheral inflammation has been
considered to increases the deleterious effect of CNS inflammation
on the nigrostriatal dopaminergic cells (7,8), and
the peripheral immunity has been recognized to increase the central
inflammation in neurodegenerative processes (9). Considering the deleterious role of
peripheral inflammation in PD development, immunomodulation as a
neuroprotective and therapeutic strategy is thought to be a novel
method for Parkinson's disease (10). Thus, it is may be a new therapeutic
approach to investigate PD from an immunological perspective to
alleviate the disease.
Treatment of PD has attracted big interest because
the prevention of PD is still a challenge for physicians. Previous
studies have reported that green tea polyphenol,
(−)-epigallocatechin-3-gallate (EGCG), prevented
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced loss of
dopaminergic neurons in the substantia nigra, which was concomitant
with a depletion in striatal dopamine and tyrosine hydroxylase (TH)
protein levels (11). Another
study demonstrated that the protective effects of EGCG in the MPTP
mouse model of PD was via inhibiting neuronal nitric oxide synthase
in the substantia nigra (12).
Moreover, EGCG has immunomodulatory effects in many disease models,
including nerve system disease (13–15).
Therefore, the present study investigated the neuroprotective
effect of EGCG and the peripheral immune response changes in the
MPTP-induced PD mouse model, which will hopefully identify the
possible targets of EGCG in PD.
Materials and methods
Ethics statement
All animal experiments were performed in strict
accordance with the recommendations in the Guide for the Care and
Use of Laboratory Animals of the National Institutes of Health
(Bethesda, MD, USA). The animal protocols were approved by the
Committee on the Ethics of Animal Experiments of the Dalian Medical
University (Dalian, China).
Animals and treatment
C57BL/6J mice (6–8 weeks old, male, weighing 16–25
g) purchased from the Experimental Animal Center of Dalian Medical
University (Dalian, China; SPF level) were used for the present
study. Mice were maintained at a constant temperature of 20–22°C
under a 12 h light/dark cycle of artificial light and had free
access to food and water. The 20 mice were randomly divided into
four groups with five per group: i) The control group, ii) the MPTP
(30 mg/kg/day) group, iii) the MPTP+EGCG (MTPT dose of 30
mg/kg/day; EGCG dose of 25 mg/kg/day) group; and iv) the MPTP+EGCG
(MTPT dose of 30 mg/kg/day; EGCG dose of 50 mg/kg/day) group. The
subchronic method was used to establish the MPTP-induced PD mouse
model (16). MPTP groups were
administered intraperitoneal injections of MPTP-HCl (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) in saline once daily at a dosage of
30 mg/kg/day for 5 consecutive days. The remaining groups were
administered intraperitoneal injections of saline. EGCG (≥97%) was
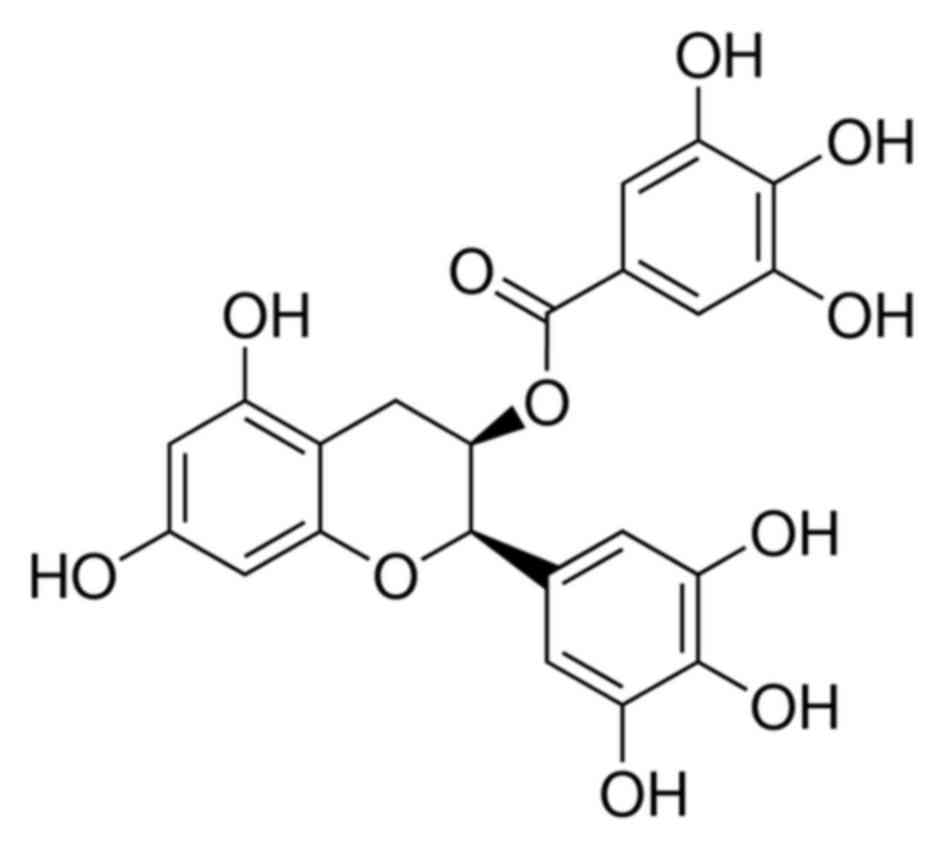
purchased from Sigma-Aldrich; Merck KGaA, the chemical structure of
EGCG is presented in Fig. 1. The
doses of EGCG (25 and 50 mg/kg) was chosen according to the
previous article (17). EGCG in
water was administered from 1 day prior to MPTP treatment to the
day 20 after MPTP injection with gavage.
Behavior test
The mice in the PD groups performed the ‘pole test’
to assess motor coordination 1 day prior to MPTP injection and on
the 5, 10, 15 and 20th day following the last MPTP injection. Mice
were trained 3 days before MPTP injection (18). The ‘pole test’ consisted of a
gauze-taped pole (50 cm high, 1 cm in diameter) with a small cork
ball at the top. Mice were placed with their head facing upwards
immediately below the ball. Two times were recorded: The time it
took for the mouse to turn completely downward (T-turn) and the
time it took to descend to the floor (T-total), with a cut-off
limit of 60 sec. The test was performed 3 times at 10 min
intervals, and the average time was recorded.
Blood sample preparation
Peripheral blood was drawn following the last day
treatment of each group. All of the mice were anesthetized and
0.5–0.6 ml peripheral blood was drawn through the angular vein.
Serum was separated from the whole blood at 4°C by centrifugation
at 400 × g for 10 min, subpackaged in an EP tube and stored at
−80°C until processed for ELISA analysis. Peripheral blood
mononuclear cells (PBMCs) were isolated with deionized water as a
lysate, incubated with fluorescently labeled antibodies against
CD3+, CD4+ and/or CD8+ T cells
(cat. nos. 100309, 100405, and 100707; BioLegend, Inc., San Diego,
CA, USA) according to the manufacturer's instructions, washed twice
with 0.01 M PBS (pH=7.4) and prepared for flow cytometric
analysis.
Brain tissue preparation
Brain tissue was dissected from mice from each group
following treatment. All of the mice were anesthetized and rapidly
perfused through the aorta with saline for 10 min, followed by 4°C
precooled 4% paraformaldehyde for 10 min. The mice were then
decapitated, and their brains were rapidly removed and post-fixed
by immersion in 4% paraformaldehye at 4°C for 12 h. Finally, the
brain was sequentially dehydrated with 20 and 30% sucrose in 0.1 M
PBS for immunofluorescence.
Immunofluorescence
Brain sections (20 µm thick) were cut at −20°C
following dehydration with 20 and 30% sucrose in 0.1 M PBS, and
were then mounted on glass slides. The sections were microwaved
twice and then cooled at room temperature for 30 min. The sections
were then rinsed 3 times and incubated in 0.3% Triton X-100 for 30
min. After washing in PBS, the sections were blocked for 30 min at
37°C with 10% normal goat serum (cat. no. SL039; 1:10; Beijing
Solarbio Science & Technology, Co., Ltd., Beijing, China). The
sections were then incubated with mouse anti-tyrosine hydroxylase
(cat. no. 22941; 1:4,000; ImmunoStar, Inc., Hudson, WI, USA) at 4°C
for 16–24 h. With overnight incubation, the sections were rinsed
and incubated in the dark for 2 h with
tetramethylrhodamine-conjugated goat anti-mouse (cat. no.
115-025-003; 1:500; Jackson ImmunoResearch Laboratories, Inc., West
Grove, PA, USA). After washing, the slides were coverslipped.
Images were taken using a fluorescence microscope (Leica DM4000B,
Leica, Wetzlar, Germany). Positive cells were measured using image
analysis software (ImageJ, version, 1.46; National Institutes of
Health, Bethesda, MD, USA).
Flow cytometric analysis
Following centrifugation at 4°C and 400 × g for 10
min, each sample was resuspended in 500 µl of 0.01 M PBS (pH=7.4 at
a density of 1×106 cells/100 µl) and analyzed using a
FAC Scan flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
PBMCs were stained at room temperature for 30 min with
PE-Cy5-conjugated anti-CD3+ (cat. no. 100309; BioLegend,
Inc.), fluorescein isothiocyanate-conjugated anti-CD4+
(cat. no. 100405; BioLegend, Inc.), and phycoerythrin-conjugated
anti-CD8+ (cat. no. 100707; BioLegend, Inc.) antibodies.
The data analysis was performed using CellQuest software (version
5.1; BD Biosciences), and the results are expressed as the
percentage of cells in a gated T cell region. CD4+ and
CD8+ T cells were gated on CD3+ T cells.
ELISA analysis
Concentrations of tumor necrosis factor (TNF)-α and
interleukin (IL)-6 in the blood serum were determined by ELISA
using commercially available kits (cat. nos. 410-TRNC-050/CF and
406-ML-005/CF; R&D Systems, Inc., Minneapolis, MN, USA).
Standard, control and test samples were added to each well and
incubated for the appropriate times with associated primary
antibodies and conjugates, in accordance with the manufacturer's
protocol (R&D Systems, Inc.). After washing four times, the
substrate solution was added to each well, and the reactions were
incubated for 30 min at room temperature in the dark. Finally, the
reactions were terminated by the addition of stop solution, and the
optical density of each well was determined at a wavelength of 450
nm using a microplate reader (AD340; BioTek Instruments, Inc.,
Winooski, VT, USA).
Statistical analysis
All quantitative data were analyzed using SPSS
software (version, 17.0; SPSS, Inc., Chicago, IL, USA). The results
are expressed as the means ± standard error of the mean. The data
were analyzed using a one-way analysis of variance, followed by
Tukey's multiple comparison test. P<0.05 was considered to
indicate a statistically significant difference. All statistical
analyses in this study were performed using Prism 5 for Windows,
version 5.01 (GraphPad Software, Inc., La Jolla, CA, USA).
Results
Effects of EGCG on behavior in the
‘pole test’
The motor dysfunction in the MPTP-induced PD model
was used to study the dopaminergic neuron degeneration and the
motor activity. The authors used the ‘pole test’ to assess whether
the MPTP-induced PD model was successful for bradykinesia. As
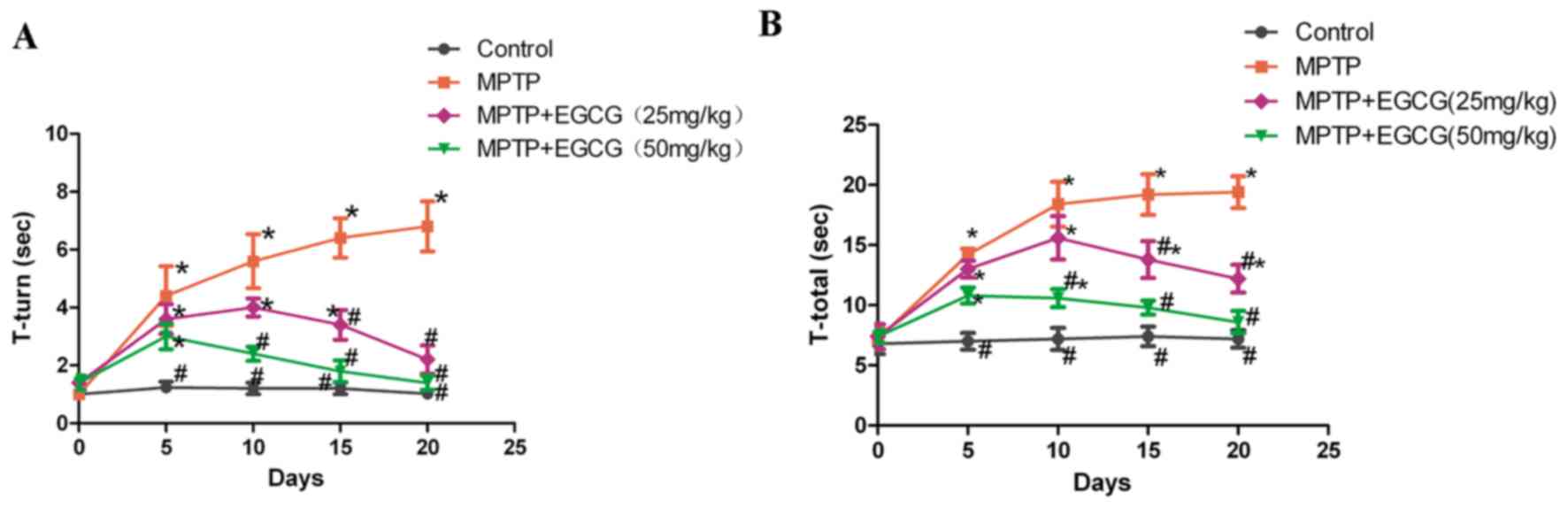
presented in Fig. 2, the effect of
EGCG on the behavior of mice was compared to the non-treated
groups. Time of ‘T-turn’ and ‘T-total’ significantly increased
following given MPTP (P<0.05) and were reduced with
administration of EGCG (P<0.05).
Effects of EGCG on tyrosine
hydroxylase (TH)-positive cells
The TH-positive neurons in the SNpc region, which
are the phenotypic marker for dopaminergic neurons, were used to
estimate the neuronal protection of EGCG in the MPTP-induced mouse
model. The number of TH-positive neurons in the SNpc region was
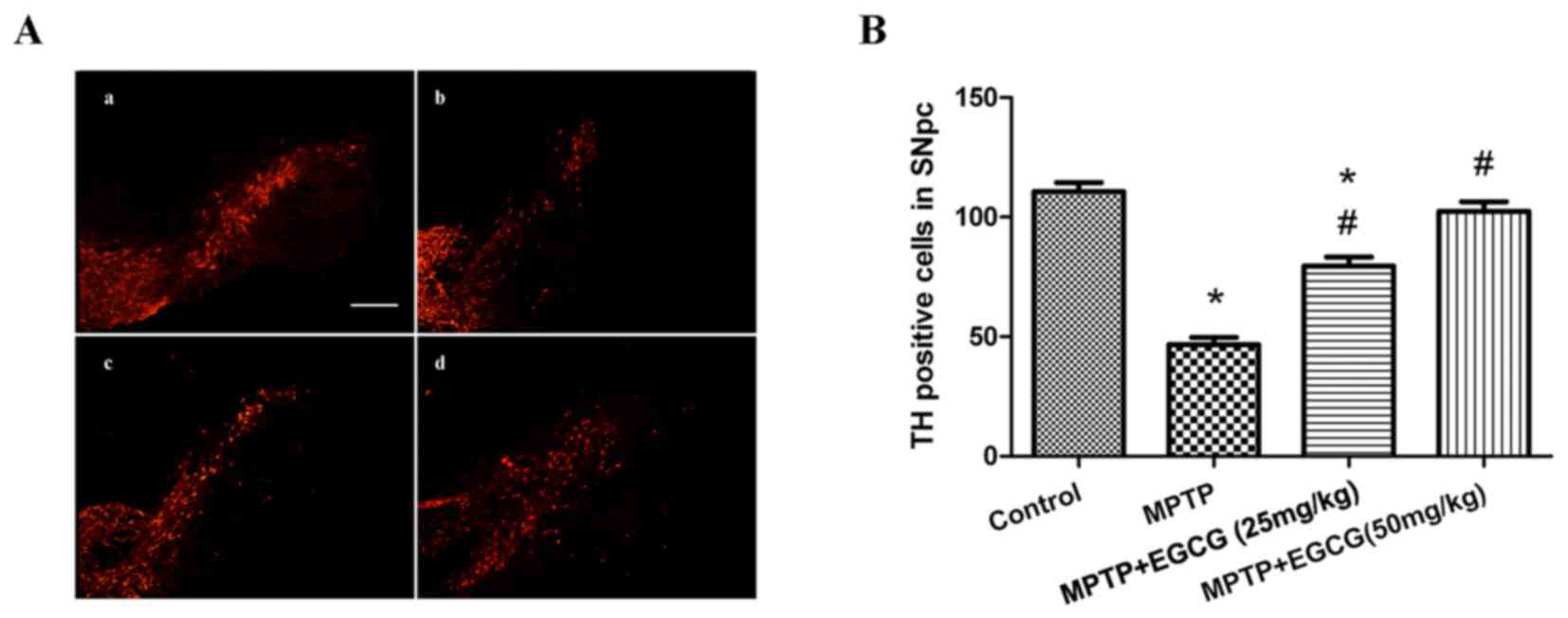
determined by counting the number of TH-positive cells. As
demonstrated in Fig. 3,
MPTP-treated mice exhibited a significant reduction in the number
of TH-positive neurons compared with the control group (P<0.05).
However, when MPTP was combined with EGCG treatment, the number of
TH-positive neurons significantly increased compared with that in
the MPTP-treated group (P<0.05).
Effects of EGCG on
CD3+CD4+ and CD3+CD8+ T
cells in the peripheral blood
To evaluate the adaptive immunity changes in
MPTP-treated mice and the therapeutic effect of EGCG, the authors
analyzed CD3+ T cells, CD3+CD4+ T
cells, and CD3+CD8+ T cells to represent the
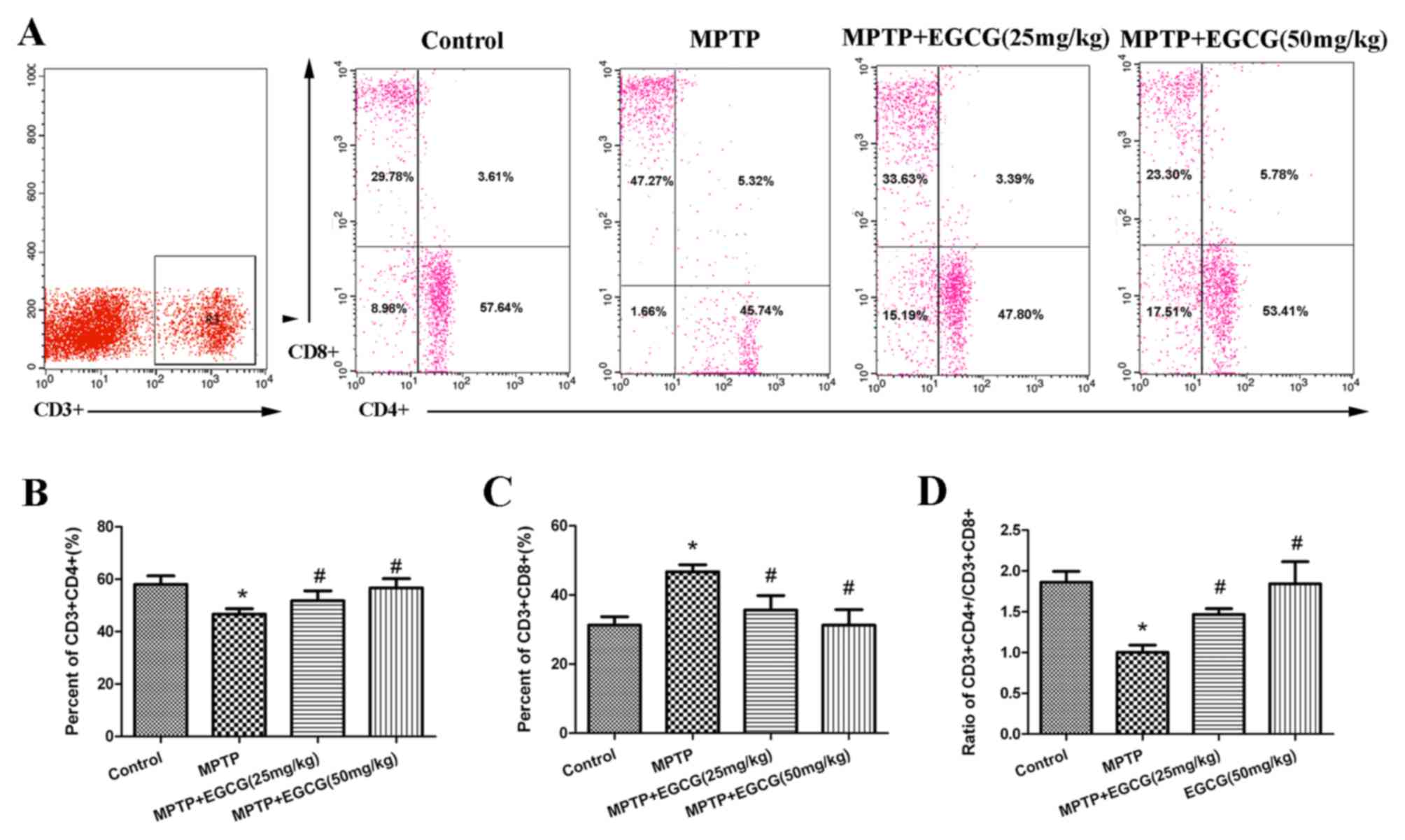
level of adaptive immunity. The results are presented in Fig. 4. The percentage of
CD3+CD4+ T cells was lower (P<0.05;
Fig. 4B) and the percentage of
CD3+CD8+ T cells was higher (P<0.05;
Fig. 4C) in MPTP mice than in
controls. The ratio of CD3+CD4+ to
CD3+CD8+ T cells was lower (P<0.05) in
MPTP mice than in controls (Fig.
4D). Whereas, with the EGCG treatment, the results were
reversed.
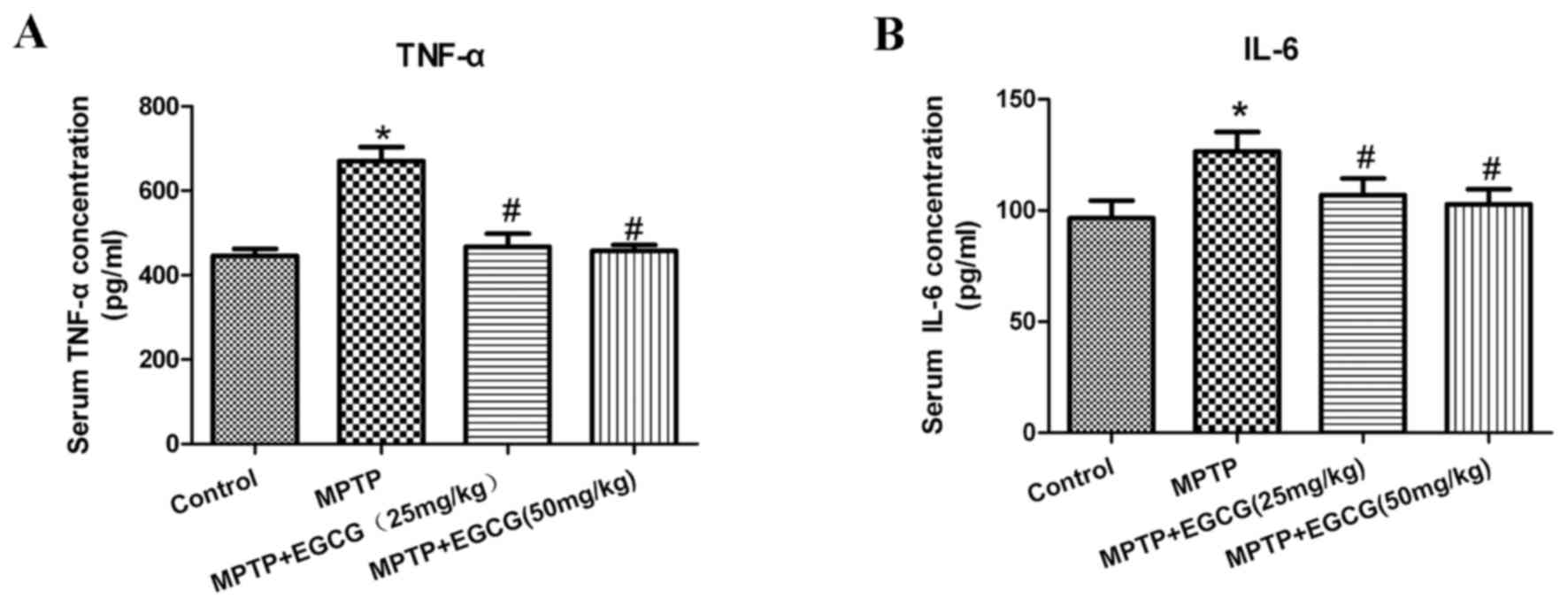
Effects of EGCG on TNF-α and IL-6 in
the serum
To further assess the effects of EGCG combined with
MPTP on the secretion of proinflammatory cytokines in PD mice, the
serum levels of TNF-α and IL-6 were tested. As presented in
(Table I) and (Fig. 5), the levels of TNF-α and IL-6
significantly increased in the MPTP mice compared with the control
mice (P<0.05). With EGCG treatment, the levels of TNF-α and IL-6
significantly decreased compared with the MPTP mice
(P<0.05).
 | Table I.Concentrations of TNF-α and IL-6 in
blood serum (pg/ml). |
Table I.
Concentrations of TNF-α and IL-6 in
blood serum (pg/ml).
| Group | n | TNF-α | IL-6 |
|---|
| Control | 8 | 445.35±17.13 |
96.50±7.89 |
| MPTP | 8 |
670.34±33.06a |
126.55±8.74a |
| MPTP+EGCG (25
mg/kg) | 8 |
456.83±31.81b |
108.12±7.18b |
| MPTP+EGCG (50
mg/kg) | 8 |
447.57±14.30b |
102.83±6.69b |
Discussion
Previous studies have indicated that peripheral
inflammation occurs in PD and accelerates disease progression
(19,20). Some nonsteroidal anti-inflammatory
drugs have been suggested to protect against PD progression and
have been associated with a lower PD risk (21). In the last decade, some of these
herbal medicines have been considered to be therapeutic agents in
PD models via their modulation of certain factors implicated in PD
pathogenesis (22–24). EGCG is the most widely studied
catechin in green tea and has been indicated to regulate immune
function (13). Although EGCG has
long been used to improve immune system function (25), the peripheral immunomodulatory
effect of EGCG has not been studied in an MPTP-induced PD mouse
model.
In the present study, the authors established the PD
mouse model induced by MPTP, which could cause dopaminergic
neuronal loss in the SNpc and lead to motor deficits in mice
(26). Consistent with reports
before, the ‘pole test’ indicated that the motor function of the PD
mice was impaired and EGCG restored the motor dysfunction. The
authors further examined the number of TH-positive dopaminergic
neurons to assess the neuroprotective role of EGCG, and the results
indicated that the dopaminergic neurons of the PD mice reduced and
EGCG prevented the loss of the neurons. These results indicated
that EGCG exerted behavior restoration and protected dopaminergic
neurons from MPTP-induced degeneration.
Previous research has implicated peripheral
inflammation in neurodegenerative diseases. Increasing studies have
demonstrated that peripheral immune system activation exacerbates
the CNS inflammatory response and accelerates neurodegeneration in
PD (26). It has been reported
that, in peripheral blood, CD4+ T cells decreased and
CD8+ T cells increased were observed in mouse model
(27,28). In the current study, the ratio of
CD3+CD4+ to CD3+CD8+ T
cells decreased, which indicated altered T cell function in MPTP
mice, whereas with EGCG treatment, the results were reversed. These
results suggested that CD3+CD4+ to
CD3+CD8+ T cells altered in the MPTP-induced
PD mice, and EGCG successfully reversed this dysfunction.
Cytokines are small proteins that function in
inflammatory processes and in the regulation of the immune system
(29). In addition, the role of
proinflammatory cytokines in the serum of MPTP-treated mice was
investigated. Studies reported that elevated serum concentrations
of TNF-α and IL-6 correlated with an increased risk of PD (7,30),
which is consistent with a previous study (31). In the present study, the serum
concentrations of TNF-α and IL-6 were elevated in PD mice and
decreased in EGCG mice. These results suggested that EGCG could
reduce proinflammatory cytokines in the PD mouse model and may be
helpful in reducing the dopaminergic neurons death, and its
modulation may represent a new therapeutic approach for PD.
In conclusion, the authors demonstrated that the
neuroprotective and immunoprotective effects of EGCG in
MPTP-treated mice. These results indicated that EGCG could modulate
the peripheral inflammation and protect dopaminergic neurons loss
in MPTP PD model. However, the underlying molecular mechanism of
the immunological effects of EGCG remain unclear, future studies
will include the mechanisms responsible for immunomodulation of
EGCG.
References
|
1
|
Dauer W and Przedborski S: Parkinson's
disease: Mechanisms and models. Neuron. 39:889–909. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jenner P and Olanow CW: The pathogenesis
of cell death in Parkinson's disease. Neurology. 66 10 Suppl
4:S24–S36. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chau KY, Ching HL, Schapira AH and Cooper
JM: Relationship between alpha synuclein phosphorylation,
proteasomal inhibition and cell death: Relevance to Parkinson's
disease pathogenesis. J Neurochem. 110:1005–1013. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lees AJ, Hardy J and Revesz T: Parkinson's
disease. Lancet. 373:2055–2066. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Badger JL, Cordero-Llana O, Hartfield EM
and Wade-Martins R: Parkinson's disease in a dish-Using stem cells
as a molecular tool. Neuropharmacology. 76:88–96. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dexter DT and Jenner P: Parkinson disease:
From pathology to molecular disease mechanisms. Free Radic Biol
Med. 62:132–144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hernández-Romero MC, Delgado-Cortés MJ,
Sarmiento M, de Pablos RM, Espinosa-Oliva AM, Argüelles S, Bández
MJ, Villarán RF, Mauriño R, Santiago M, et al: Peripheral
inflammation increases the deleterious effect of CNS inflammation
on the nigrostriatal dopaminergic system. Neurotoxicology.
33:347–360. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen H, O'Reilly EJ, Schwarzschild MA and
Ascherio A: Peripheral inflammatory biomarkers and risk of
Parkinson's disease. Am J Epidemiol. 167:90–95. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Amor S and Woodroofe MN: Innate and
adaptive immune responses in neurodegeneration and repair.
Immunology. 141:287–291. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Olson KE and Gendelman HE:
Immunomodulation as a neuroprotective and therapeutic strategy for
Parkinson's disease. Curr Opin Pharmacol. 26:87–95. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Levites Y, Weinreb O, Maor G, Youdim MB
and Mandel S: Green tea polyphenol (−)-epigallocatechin-3-gallate
prevents N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced
dopaminergic neurodegeneration. J Neurochem. 78:1073–1082. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Choi JY, Park CS, Kim DJ, Cho MH, Jin BK,
Pie JE and Chung WG: Prevention of nitric oxide-mediated
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced Parkinson's
disease in mice by tea phenolic epigallocatechin 3-gallate.
Neurotoxicology. 23:367–374. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kuo CL, Chen TS, Liou SY and Hsieh CC:
Immunomodulatory effects of EGCG fraction of green tea extract in
innate and adaptive immunity via T regulatory cells in murine
model. Immunopharmacol Immunotoxicol. 36:364–370. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yoneyama S, Kawai K, Tsuno NH, Okaji Y,
Asakage M, Tsuchiya T, Yamada J, Sunami E, Osada T, Kitayama J, et
al: Epigallocatechin gallate affects human dendritic cell
differentiation and maturation. J Allergy Clin Immunol.
121:209–214. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kuang X, Huang Y, Gu HF, Zu XY, Zou WY,
Song ZB and Guo QL: Effects of intrathecal epigallocatechin
gallate, an inhibitor of Toll-like receptor 4, on chronic
neuropathic pain in rats. Eur J Pharmacol. 676:51–56. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jackson-Lewis V and Przedborski S:
Protocol for the MPTP mouse model of Parkinson's disease. Nat
Protoc. 2:141–151. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wei IH, Wu YC, Wen CY and Shieh JY: Green
tea polyphenol (−)-epigallocatechin gallate attenuates the neuronal
NADPH-d/nNOS expression in the nodose ganglion of acute hypoxic
rats. Brain Res. 999:73–80. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sedelis M, Schwarting RK and Huston JP:
Behavioral phenotyping of the MPTP mouse model of Parkinson's
disease. Behav Brain Res. 125:109–125. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stone DK, Reynolds AD, Mosley RL and
Gendelman HE: Innate and adaptive immunity for the pathobiology of
Parkinson's disease. Antioxid Redox Signal. 11:2151–2166. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
McGeer PL and McGeer EG: Inflammation and
the degenerative diseases of aging. Ann N Y Acad Sci. 1035:104–116.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wahner AD, Bronstein JM, Bordelon YM and
Ritz B: Nonsteroidal anti-inflammatory drugs may protect against
Parkinson disease. Neurology. 69:1836–1842. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Choi JG, Kim HG, Kim MC, Yang WM, Huh Y,
Kim SY and Oh MS: Polygalae radix inhibits toxin-induced neuronal
death in the Parkinson's disease models. J Ethnopharmacol.
134:414–421. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hwang DS, Kim HG, Kwon HJ, Cho JH, Lee CH,
Lee JM, Jang JB, Kim YS, Lee KS and Oh MS: Dangguijakyak-san, a
medicinal herbal formula, protects dopaminergic neurons from
6-hydroxydopamine-induced neurotoxicity. J Ethnopharmacol.
133:934–939. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lu C, Zhang J, Shi X, Miao S, Bi L, Zhang
S, Yang Q, Zhou X, Zhang M, Xie Y, et al: Neuroprotective effects
of tetramethylpyrazine against dopaminergic neuron injury in a rat
model of Parkinson's disease induced by MPTP. Int J Biol Sci.
10:350–357. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Katiyar SK, Challa A, McCormick TS, Cooper
KD and Mukhtar H: Prevention of UVB-induced immunosuppression in
mice by the green tea polyphenol (−)-epigallocatechin-3-gallate may
be associated with alterations in IL-10 and IL-12 production.
Carcinogenesis. 20:2117–2124. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Goldberg NR, Haack AK, Lim NS, Janson OK
and Meshul CK: Dopaminergic and behavioral correlates of
progressive lesioning of the nigrostriatal pathway with
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Neuroscience.
180:256–271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Amor S, Peferoen LA, Vogel DY, Breur M,
van der Valk P, Baker D and van Noort JM: Inflammation in
neurodegenerative diseases-an update. Immunology. 142:151–166.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bas J, Calopa M, Mestre M, Molleví DG,
Cutillas B, Ambrosio S and Buendia E: Lymphocyte populations in
Parkinson's disease and in rat models of parkinsonism. J
Neuroimmunol. 113:146–152. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dinarello CA: Historical insights into
cytokines. Eur J Immunol. 37 Suppl 1:S34–S45. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Koziorowski D, Tomasiuk R, Szlufik S and
Friedman A: Inflammatory cytokines and NT-proCNP in Parkinson's
disease patients. Cytokine. 60:762–766. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dobbs RJ, Charlett A, Purkiss AG, Dobbs
SM, Weller C and Peterson DW: Association of circulating TNF-alpha
and IL-6 with ageing and parkinsonism. Acta Neurol Scand.
100:34–41. 1999. View Article : Google Scholar : PubMed/NCBI
|