Introduction
Differentiated thyroid carcinoma (DTC) accounts for
~86% of all thyroid cancers, which are divided into papillary
thyroid and follicular thyroid type carcinomas (1). The treatment efficiency of thyroid
carcinoma has been substantially improved by thyroidectomy or
131iodine (131I) radiotherapy; however, the
10-year-survival rate for certain patients was reported to be 10%
(2). The particular target of
radioiodine in thyroid disease is sodium iodide symporter, which is
an intrinsic plasma membrane protein that mediates active iodide
transport into the thyroid gland (3). Adjuvant therapy in the form of
radioactive iodine is often administered as a means of reducing the
risk of tumor recurrence and to facilitate future cancer
surveillance (4). However, certain
side effects have been reported, such as neck pain or swelling,
headache and vertigo, or insomnia (5). Regardless of whether patients respond
to 131I, it is difficult to obtain a satisfactory
therapeutic effect with 131I treatment alone. Therefore,
treatments with increased efficacy for thyroid carcinoma are
required (6).
The transcription factor nuclear factor (NF)-κB was
first discovered in 1986 as a nuclear factor which binds to the
enhancer element of the immunoglobulin kappa light-chain of
activated B cells (thereby coining the abbreviation NF-κB)
(7,8). Later on, several studies demonstrated
that NF-κB is a ubiquitously expressed transcription factor that
mediates signal-induced expression of numerous genes involved in
different biological processes, including immune responses,
inflammation, cell growth and survival (9,10).
NF-κB is normally sequestered in the cytoplasm as an inactive
complex via physical association with inhibitory proteins, termed
IκBs. In response to immune and stress stimuli, NF-κB associated
proteins become activated via two major signaling pathways, the
canonical and noncanonical pathways, and translocate to the nucleus
to exert transcriptional functions (11–13).
In recent years, in-depth research concerning thyroid carcinoma has
revealed that nuclear factor-κB (NF-κB) is closely associated with
the occurrence and development of thyroid carcinoma (14). Additionally, it has been reported
that radiotherapy and chemotherapy may induce the production of
NF-κB within thyroid carcinomas, leading to resistance (15,16).
Bauerle et al (17)
demonstrated that the inhibition of NF-κB activity may promote the
apoptosis of thyroid carcinoma cells; however, the association
between the expression of NF-κB and 131I radiation
therapy in DTC remains unclear. Therefore, the present study aimed
to investigate the effects of NF-κB on the uptake of
131I and apoptosis in thyroid carcinoma cells.
Materials and methods
Cell culture
The TPC-1 and BCPAP human papillary thyroid
carcinoma cell lines were obtained from the Chinese Academy of
Sciences (Shanghai, China). Dulbecco's modified Eagle's medium was
obtained from Gibco (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and was supplemented with 5% fetal bovine serum
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), 1% (w/v)
penicillin (final concentration: 100 IU/ml) and streptomycin (100
IU/ml), and 1 mU/ml thyroid-stimulating hormone. Penicillin and
streptomycin were purchased from Sigma-Aldrich (Merck KGaA). The
conditions of cell culture were 37°C and 5% CO2
(18).
Cell transfection and RNA interference
(RNAi)
According to the mRNA sequence of the NF-κB gene of
humans (GenBank accession number, X61498), the sequence of RNAi was
designed. The mRNA sequences were selected with GC contents of 50%
and lengths of 29 bp; the aforementioned sequences and the mRNA of
other genes in the Human Genome Database (19) were analyzed with NCBI-BLAST
(20) (https://blast.ncbi.nlm.nih.gov/Blast.cgi) to exclude
the presence of homologous sequences; an NF-κB-RNAi sequence in
accordance with the conditions was selected, some of the base
sequences were mutated randomly, following exclusion of the
homologous sequences with other genes and finally the sequences
were selected as the negative control scramble-RNAi. Sequences are
presented in Table I, which were
synthesized by Shanghai Shenggong Biology Engineering Technology
Service, Ltd. (Shanghai, China).
 | Table I.Nucleic acid sequences for RNAi. |
Table I.
Nucleic acid sequences for RNAi.
| RNAi | Sequence |
|---|
| NF-κB |
5′-CTTCCTTGTCTTCCACCAGAGGGTAATAG-3′ |
| Scramble |
5′-TCTCGTCTTATCTCCGGACCAGAGAGTAT-3′ |
Once thawed, the TPC-1 and BCPAP cells were cultured
to the logarithmic growth phase and digested with trypsin for
counting. The cells were diluted with fresh medium and were
inoculated into a 96-well plate at a density of
5×104/well for 24 h at 37°C. Targeted RNA (final
concentration 50 mM) or control scramble RNA (50 mM) was
transfected into the cells using a Liposome INTERFER in
Transfection kit (Polyplus-transfection SA, Illkirch, France)
according to the manufacturer's protocol. After 6 h, medium was
replaced with normal culture medium containing serum. After 48 h
continuous incubation, cells were collected for further
analysis.
131I uptake
experiments
To determine whether the inhibition of NF-κB
expression affects the 131I uptake of thyroid carcinoma
cells, TCP-1 and BCPAP cells were cultured to the logarithmic
growth phase and were seeded into a 6-well plate
(5×105/well). Into each well, 1 ml fresh culture medium
containing Na131I (37 kBq) (Sigma-Aldrich; Merck KGaA)
was added. After 1 h, the culture medium was removed and the cells
were collected and treated with trypsin following two washes with
PBS. Cell radiation was measured using a γ-ray counter. The effect
of NF-κB expression on the uptake of 131I was
investigated (21).
Detection of cell viability by MTT
assay
TPC-1 and BCPAP cell lines were transfected with
NF-κB-RNAi or scramble-RNAi sequences and cultured to the
logarithmic growth phase prior to digestion with trypsin. Following
centrifugation at 500 × g for 5 min at 4°C, the supernatant was
discarded and cultured with the cell suspension. A total of 6 µl
cell suspension was treated with 6 µl trypan blue solution;
following mixing, 10 µl solution was applied dropwise onto the
cover between cell counting plate. The cells were counted under a
light microscope (magnification ×40) selecting 6 random fields.
These two cell groups were divided into four groups: A,
scramble-RNAi group; B, scramble-RNAi + 131I group; C,
NF-κB-RNAi group; and D, NF-κB-RNAi + 131I group. Groups
A and C were suspended in fresh and normal culture medium; however,
groups B and D were suspended in fresh culture medium containing 20
MBq/ml 131I (Institute of Isotopes Co., Ltd., Budapest,
Hungary). The four groups of cell suspensions were plated in
24-well plates with 6,000 cells per well, and were cultured in a
CO2 incubation box for 24 h at 37°C. To each well, 70 µl
MTT solution was added and cells were cultured in a CO2
incubation box for 3 h at 37°C. Dimethyl sulfoxide was added
following the removal of the supernatant and detection was
performed at 570 nm (22).
Determination of the protein
expression of NF-κB and apoptosis-associated proteins by western
blot analysis
Cells of the four aforementioned groups were
cultured for 24 h at 37°C. Following collection, the cells were
pyrolyzed with radioimmunoprecipitation assay lysis buffer (Thermo
Fisher Scientific, Inc.) and total protein was extracted followed
by quantification by Pierce bicinchoninic acid Protein Assay kit
(Thermo Fisher Scientific, Inc.). Protein (20 µg/lane) was loaded
into a SDS-PAGE (15% separation gel and 5% concentration gel).
Proteins were transferred to polyvinylidene difluoride membranes by
an electrophoretic transfer method following electrophoresis. The
membrane was blocked with 5% non-fat milk powder for 1 h at 37°C;
following washing with TBS with 0.1% Tween-20 (TBST), the membrane
was incubated with a solution of antibodies, including anti-human
NF-κB (catalogue number #8242; Cell Signaling Technology, Danvers,
MA, USA; 1:1,000 dilution), anti human X-linked inhibitor of
apoptosis (XIAP) (catalogue number #2045; Cell Signaling
Technology; 1:1,000 dilution), anti-human cellular inhibitor of
apoptosis protein 1 (cIAP1) (catalogue number #4952; Cell Signaling
Technology; (1:1,000 dilution), anti-human caspase-3 (catalogue
number #9662; Cell Signaling Technology; 1:1,000 dilution) and
anti-human β-actin (catalogue number #4970; Cell Signaling
Technology; 1:1,000 dilution) primary antibodies, at 4°C overnight.
Subsequently, the membrane was washed with TBST and incubated with
horseradish-peroxidase-conjugated anti-rabbit immunoglobulin G
secondary antibody (catalogue number #47074; Cell Signaling
Technology; 1:2,000 dilution) at room temperature for 1 h.
Following washing with TBST, 3,3′-diaminoenzidine was applied to
the membrane for 10 min to allow any color to develop; distilled
water was added to terminate this reaction. The western blotting
image was analyzed and the integral gray value was detected by a
gel image-forming analysis system. The relative expression level
was quantified using Image J software version 1.48 (National
Institutes of Health, Bethesda, MD, USA).
Statistical analysis
Data were analyzed by SPSS 19 statistical software
(IBM Corp., Armonk, NY, USA). Measurement data are presented as the
mean ± standard deviation. One-way analysis of variance with
Newman-Keuls multiple comparison post-hoc analysis was performed
for comparison of the difference. P<0.05 was considered to
indicate a statistically significant difference.
Results
Results of RNAi
An experiment with RNAi was performed by designing
specific oligonucleotide sequences. Cells transfected with the
negative control scramble sequence were defined as the control.
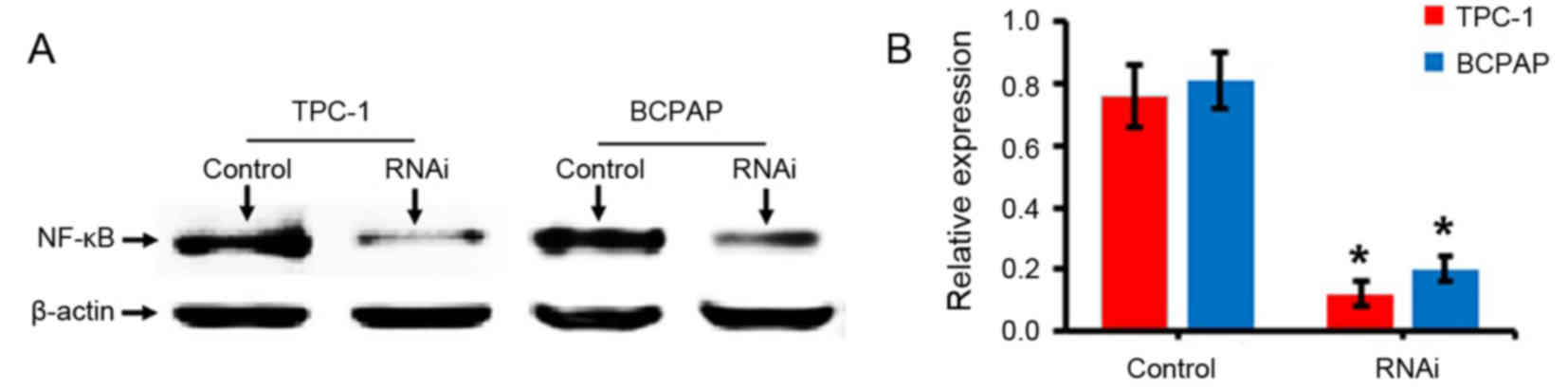
Western blotting results demonstrated that the expression levels of
NF-κB (p65) protein in TPC-1 and BCPAP cells following the
transfection of NF-κB-RNAi were significantly decreased compared
with the control group (P<0.05; Fig. 1), indicating that the cells were
successfully transfected with the NF-κB-RNAi oligonucleotide
sequence.
Effects of NF-κB expression on thyroid
carcinoma cell uptake of 131I
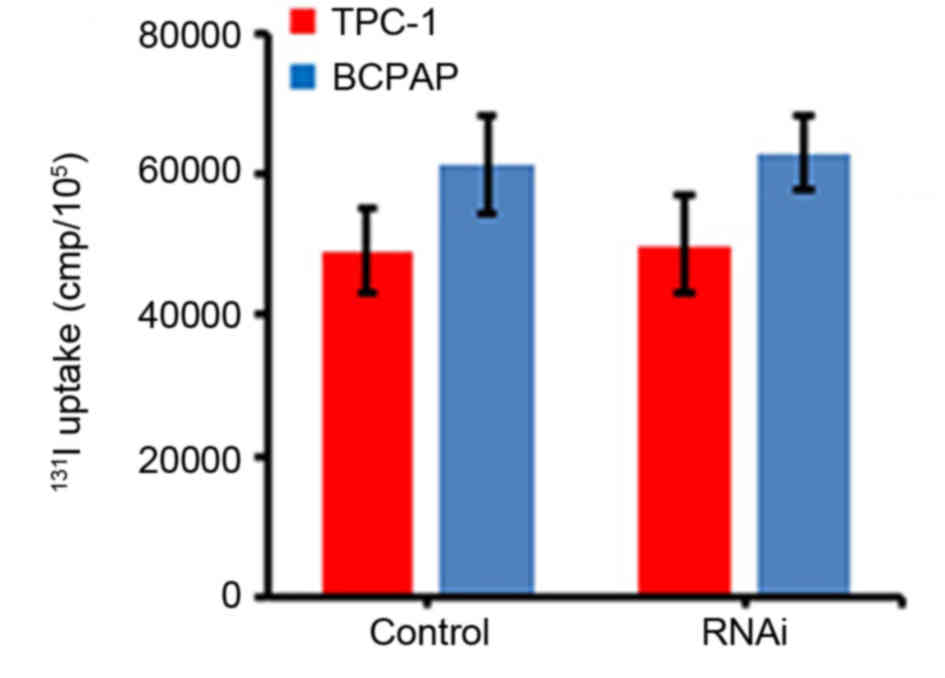
The effects of NF-κB expression on the
131I uptake of thyroid carcinoma cells were evaluated by
an 131I uptake experiment. The results demonstrated that
the uptake of 131I by TPC-1 and BCPAP cells was not
significantly altered following the inhibition of NF-κB expression
using RNAi, compared with the cells transfected with scramble RNAi
(P>0.05; Fig. 2).
Detection of alterations in cell
viability by MTT assays
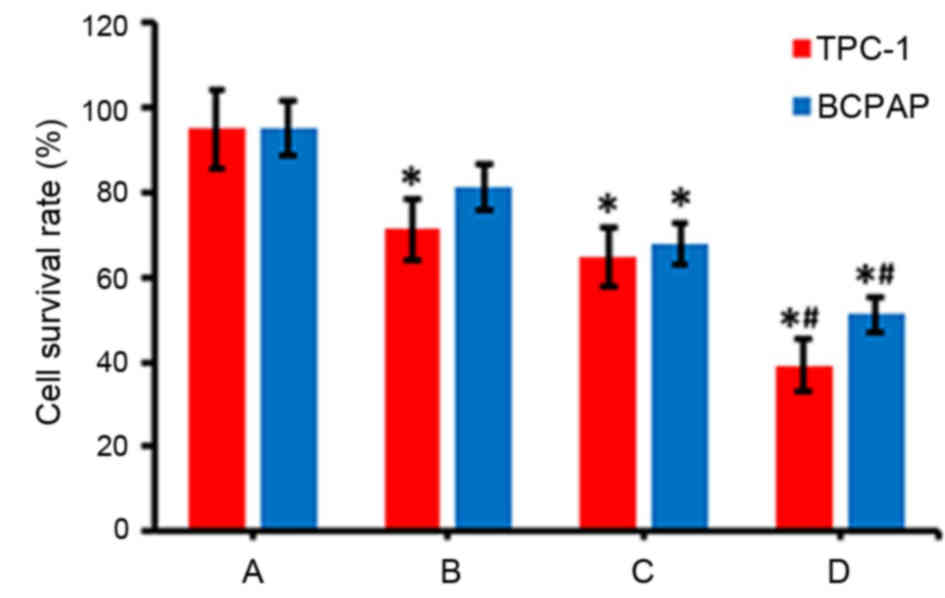
The viability of TPC-1 and BCPAP cells was detected
via an MTT assay following various treatments. As presented in
Fig. 3, the radioactive effects of
131I (group B) or interference and inhibition of NF-κB
expression (group C) decreased the survival rate of cells, compared
with the control group (group A; P<0.05). Furthermore, when
NF-κB inhibition and 131I radiation was combined (group
D), the survival rate of thyroid carcinoma cells was markedly lower
compared with groups B and C (Fig.
3).
Expression of apoptosis-associated
proteins in thyroid carcinoma cells
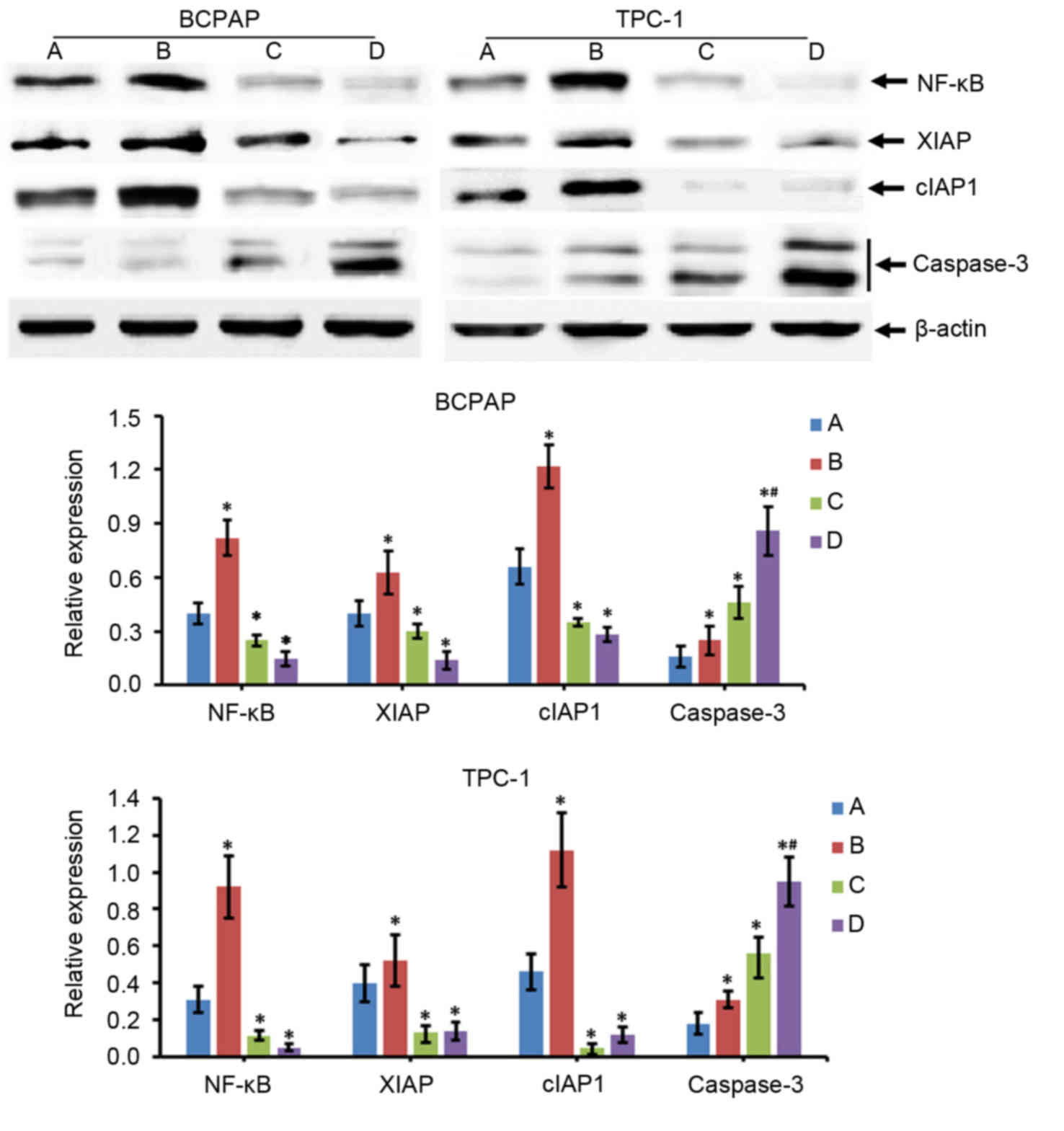
Alterations in the expression of
apoptosis-associated proteins were determined by western blotting.
The results of the present study revealed that compared with the
control group (group A), the expression levels of NF-κB, XIAP,
cIAP1 and caspase-3 in the 131I treatment group (group
B) were significantly increased (P<0.05; Fig. 4). However, the protein expression
levels of NF-κB, XIAP and cIAP1 were decreased in the NF-κB
interference group (group C) compared with in group A (P<0.05),
while levels of caspase-3 were increased (P<0.05; Fig. 4). The results for the
131I treatment + NF-κB interference group (group D) were
similar to those in group C. In addition, compared with group C,
caspase-3 expression levels were increased significantly in group D
(P<0.05), but the alterations in NF-κB, XIAP and cIAP1
expression levels between groups C and D were not significantly
different (P>0.05; Fig. 4).
Discussion
Thyroid carcinoma is a common type of malignant
tumor in humans and in recent years, due alterations in human
lifestyle and living environments, the incidence of this disease
has been increasing annually. Research concerning the molecular
biology of thyroid carcinoma has indicated that combining
thyroidectomy with radioactive 131I may improve the
efficacy of treatment for this disease. Additionally, β-ray therapy
and radiotherapy in vitro have exhibited lethal effects on
cancer cells (23); however,
certain patients with thyroid carcinoma have been reported to
exhibit resistance to 131I radiation (24). A study reported that NF-κB was
associated with the occurrence, development and treatment, as well
as the resistance, of various types of malignant tumor, such as
thyroid carcinoma (25).
Researchers have also demonstrated that the inhibition of NF-κB
activity may induce cancer cell apoptosis (26). In the present study, the use of
RNAi technology to inhibit the expression of NF-κB was
investigated, followed by analysis of the effects of NF-κB on the
131I uptake by thyroid carcinoma cells. In addition, the
levels of apoptosis were investigated. The results of the present
study revealed that decreased expression levels of NF-κB exhibited
no significant effects on the ability of thyroid carcinoma cells to
uptake 131I; however, NF-κB inhibition may enhance the
function of 131I-induced apoptosis of cancer cells.
Various studies have demonstrated that NF-κB may
inhibit the apoptosis of various types of cancer cells as NF-κB
serves as a nuclear factor that regulates the expression of various
cell apoptosis-inhibiting genes at the transcriptional level,
including XIAP, cIAP1and B-cell lymphoma-extra-large (27–29).
XIAP and cIAP1 belong to the family of inhibitor of apoptosis (IAP)
proteins, which constitute a highly conserved family of endogenous
anti-apoptotic factors that suppress apoptosis by inhibiting
caspase activity (30). IAP family
proteins within the mitochondrial pathway are able to bind to
caspase-9 precursors and interfere with their processing.
Additionally, the caspase activation and recruitment domain of the
IAP family protein structure binds to apoptotic
peptidase-activating factor 1 to interfere with the activation of
caspases (31). Finally, XIAP is
reported to inhibit the activation of caspase-3 and caspase-7 via
the baculovirus inhibitor of apoptosis protein repeat domain to
inhibit apoptosis (32). In the
present study, the protein expression of XIAP, cIAP1 and caspase-3
in cells was detected by western blotting following various
treatments, and the results demonstrated that the inhibition of
NF-κB was associated with decreased expression levels of XIAP and
cIAP, and increased expression levels of caspase-3, compared with
the control group. However, although 131I exposure
increased the expression levels of the anti-apoptotic genes NF-κB,
XIAP and cIAP, cell viability was also reduced following
131I exposure, indicating that other factors may be
involved, which may include microRNA-100 and retinoblastoma 1
serine phosphates from human chromosome 3, as reported by a
previous study (33). In addition,
although the inhibition of NF-κB mediated by RNAi markedly
suppressed the protein expression of XIAP and cIAP, and increased
caspase-3 expression, NF-κB inhibition did not reduce cell
viability compared with control group, which may be due to the
function of other factors that may compensate the effect of NF-κB
inhibition on cell proliferation, including certain cell cycle
proteins, retinoblastoma protein or cyclin-dependent kinases
(34).
The balance between apoptosis-inducing effects and
anti-apoptotic effects of cells is important for normal cell
function; however, when this balance is disrupted, cells may become
cancerous. NF-κB is reported to increase the transcriptional level
of the anti-apoptotic genes, which may promote the malignant
transformation of cells (35).
Furthermore, NF-κB has been demonstrated to regulate cyclin D1 and
promote cell proliferation, and also promote the expression of the
proto-oncogene, H-ras, leading to the promotion of cell
carcinogenesis (36). In the
course of cancer treatment, the activity of NF-κB has been reported
to be closely associated with the resistance of cancer cells to
radiotherapy and chemotherapy. Therefore, the effects of
radiotherapy and chemotherapy treatments may be improved by
inhibiting the activity of NF-κB in a particular manner (37).
In conclusion, in the present study, the expression
of NF-κB in thyroid carcinoma cells was inhibited by RNAi the
effects of NF-κB, and the results demonstrated that although NF-κB
inhibition did not alter the uptake of 131I by DTC
cells, the inhibition of NF-κB expression may enhance
131I-induced apoptosis of DTC cells. These results may
provide a theoretical basis for improving the efficacy of
radioactive 131I in the treatment of thyroid carcinoma
and reducing the effective therapeutic dose of 131I in
the future.
References
|
1
|
Pellegriti G, Frasca F, Regalbuto C,
Squatrito S and Vigneri R: Worldwide increasing incidence of
thyroid cancer: Update on epidemiology and risk factors. J Cancer
Epidemiol. 2013:9652122013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Londero SC, Krogdahl A, Bastholt L,
Overgaard J, Trolle W, Pedersen HB, Bentzen J, Schytte S,
Christiansen P and Godballe C; Danish Thyroid Cancer Group, :
Papillary thyroid microcarcinoma in Denmark 1996–2008: A national
study of epidemiology and clinical significance. Thyroid.
23:1159–1164. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lin Y: Internal radiation therapy: A
neglected aspect of nuclear medicine in the molecular era. J Biomed
Res. 29:345–355. 2015.PubMed/NCBI
|
|
4
|
Pryma DA and Mandel SJ: Radioiodine
therapy for thyroid cancer in the era of risk stratification and
alternative targeted therapies. J Nucl Med. 55:1485–1491. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu L, Shan F, Li W and Lu H: Short-term
side effects after radioiodine treatment in patients with
differentiated thyroid cancer. Biomed Res Int. 2016:43767202016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hombach-Klonisch S, Natarajan S,
Thanasupawat T, Medapati M, Pathak A, Ghavami S and Klonisch T:
Mechanisms of therapeutic resistance in cancer (stem) cells with
emphasis on thyroid cancer cells. Front Endocrinol (Lausanne).
5:372014.PubMed/NCBI
|
|
7
|
Sen R and Baltimore D: Multiple nuclear
factors interact with the immunoglobulin enhancer sequences. Cell.
46:705–716. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sen R and Baltimore D: Inducibility of
kappa immunoglobulin enhancer-binding protein Nf-kappa B by a
posttranslational mechanism. Cell. 47:921–928. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vallabhapurapu S and Karin M: Regulation
and function of NF-kappaB transcription factors in the immune
system. Annu Rev Immunol. 27:693–733. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hayden MS and Ghosh S: NF-κB, the first
quarter-century: Remarkable progress and outstanding questions.
Genes Dev. 26:203–234. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang H and Sun SC: NF-kappaB in
inflammation and renal diseases. Cell Biosci. 5:632015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wertz IE and Dixit VM: Signaling to
NF-kappaB: Regulation by ubiquitination. Cold Spring Harb Perspect
Biol. 2:a0033502010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Oeckinghaus A, Hayden MS and Ghosh S:
Crosstalk in NF-κB signaling pathways. Nat Immunol. 12:695–708.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pozdeyev N, Berlinberg A, Zhou Q, Wuensch
K, Shibata H, Wood WM and Haugen BR: Targeting the NF-κB Pathway as
a combination therapy for advanced thyroid cancer. PLos One.
10:e01349012015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li F and Sethi G: Targeting transcription
factor NF-kappa B to overcome chemoresistance and radioresistance
in cancer therapy. Biochim Biophys Acta. 1805:167–180.
2010.PubMed/NCBI
|
|
16
|
Lin Y, Bai L, Chen WJ and Xu SL: The
NF-kappa B activation pathways, emerging molecular targets for
cancer prevention and therapy. Expert Opin Ther Tar. 14:45–55.
2010. View Article : Google Scholar
|
|
17
|
Bauerle KT, Schweppe RE and Haugen BR:
Inhibition of nuclear factor-kappa B differentially affects thyroid
cancer cell growth, apoptosis, and invasion. Mol Cancer. 9:1172010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Phan T, Yu XM, Kunnimalaiyaan M and Chen
H: Antiproliferative effect of chrysin on anaplastic thyroid
cancer. J Surg Res. 170:84–88. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Letovsky SI, Cottingham RW, Porter CJ and
Li PW: GDB: The human genome database. Nucleic Acids Res. 26:94–99.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Camacho C, Coulouris G, Avagyan V, Ma N,
Papadopoulos J, Bealer K and Madden TL: BLAST+: Architecture and
applications. BMC Bioinformatics. 10:4212009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Flux GD, Haq M, Chittenden SJ, Buckley S,
Hindorf C, Newbold K and Harmer CL: A dose-effect correlation for
radioiodine ablation in differentiated thyroid cancer. Eur J Nucl
Med Mol Imaging. 37:270–275. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen GF, Xu SH, Renko K and Derwahl M:
Metformin inhibits growth of thyroid carcinoma cells, suppresses
self-renewal of derived cancer stem cells, and potentiates the
effect of chemotherapeutic agents. J Clin Endocr Metab.
97:E510–E520. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Verburg FA, Stokkel MP, Düren C,
Verkooijen RB, Mäder U, van Isselt JW, Marlowe RJ, Smit JW, Reiners
C and Luster M: No survival difference after successful (131)I
ablation between patients with initially low-risk and high-risk
differentiated thyroid cancer. Eur J Nucl Med Mol Imaging.
37:276–283. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim WG, Ryu JS, Kim EY, Lee JH, Baek JH,
Yoon JH, Hong SJ, Kim ES, Kim TY, Kim WB and Shong YK: Empiric
high-dose 131-iodine therapy lacks efficacy for treated papillary
thyroid cancer patients with detectable serum thyroglobulin, but
negative cervical sonography and 18F-fluorodeoxyglucose positron
emission tomography scan. J Clin Endocr Metab. 95:1169–1173. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pacifico F and Leonardi A: Role of
NF-kappa B in thyroid cancer. Mol Cell Endocrinol. 321:29–35. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lampiasi N, Azzolina A, Umezawa K,
Montalto G, McCubrey JA and Cervello M: The novel NF-κB inhibitor
DHMEQ synergizes with celecoxib to exert antitumor effects on human
liver cancer cells by a ROS-dependent mechanism. Cancer Lett.
322:35–44. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Galbán S and Duckett CS: XIAP as a
ubiquitin ligase in cellular signaling. Cell Death Differ.
17:54–60. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiang CY and Lin X: Regulation of NF-κB by
the CARD proteins. Immunol Rev. 246:141–153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Go HS, Seo JE, Kim KC, Han SM, Kim P, Kang
YS, Han SH, Shin CY and Ko KH: Valproic acid inhibits neural
progenitor cell death by activation of NF-κB signaling pathway and
up-regulation of Bcl-XL. J Biomed Sci. 18:482011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gyrd-Hansen M and Meier P: IAPs: From
caspase inhibitors to modulators of NF-kappa B, inflammation and
cancer. Nat Rev Cancer. 10:561–574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bratton SB and Salvesen GS: Regulation of
the Apaf-1-caspase-9 apoptosome. J Cell Sci. 123:3209–3214. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Altieri DC: Survivin and IAP proteins in
cell-death mechanisms. Biochem J. 430:199–205. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang SN, Deng BP, Zhang YY and Jiang NY:
Expression of miR-100 and RBSP3 in FTC-133 cells after exposure to
I-131. Nucl Med Commun. 35:932–938. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Duronio RJ and Xiong Y: Signaling pathways
that control cell proliferation. Cold Spring Harb Perspect Biol.
5:a0089042013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu M, Sakamaki T, Casimiro MC, Willmarth
NE, Quong AA, Ju X, Ojeifo J, Jiao X, Yeow WS, Katiyar S, et al:
The canonical NF-kappaB pathway governs mammary tumorigenesis in
transgenic mice and tumor stem cell expansion. Cancer Res.
70:10464–10473. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hinz M, Krappmann D, Eichten A, Heder A,
Scheidereit C and Strauss M: NF-kappaB function in growth control:
Regulation of cyclin D1 expression and G0/G1-to-S-phase transition.
Mol Cell Biol. 19:2690–2698. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li XY, Abdel-Mageed AB, Mondal D and
Kandil E: The nuclear factor Kappa-B signaling pathway as a
therapeutic target against thyroid cancers. Thyroid. 23:209–218.
2013. View Article : Google Scholar : PubMed/NCBI
|