Introduction
Stroke is the leading cause of mortality and
permanent disabilities worldwide (1). Patients with ischemic brain injury
suffer from strokes and the prognosis is poor. Thus, the current
treatment of strokes requires further improvement.
Previous studies have demonstrated that decreased
quantities of microvessels, reduced collateral circulation and
damaged neovascularization were associated with the poor prognosis
of stroke (2). As a type of adult
stem cell, endothelial progenitor cells (EPCs) are pivotal in
keeping the endothelium intact and for vascular homeostasis during
the process of angiogenesis (3,4).
These functions allow EPCs to engage in vascular repair. Increasing
evidence indicates that the quantity of EPCs decreased in stroke
patients (5–7). Furthermore, studies demonstrate that
infusion of EPCs attenuated tissue injury, promoted angiogenesis
and improved functional recovery of ischemia organs, such as the
heart and brain (8,9). These findings provide the theoretical
basis for treatment of EPCs.
microRNAs (miRNA) are a type of non-coding RNA
(length, 20–25 bp) and >30% of human genes are under
post-transcriptional regulation of miRNAs (10). The post-transcription regulatory
effect of miRNAs provides a novel strategy for gene expression
regulation and influences cellular events (11). A previous study indicated that
miRNA is involved in angiogenesis and miR-126 modification enhances
the function of EPCs, so as to improve therapeutic efficacy
(12).
Thus, the present study evaluated the expression
level and function of miR-126 in EPCs, and demonstrated the
miR-126-associated mechanism using a cell migration model in
vitro and a middle cerebral artery occlusion model in
vivo.
Materials and methods
Extraction, culture and identification
of EPCs
A total of 15 healthy male mice, aged 8–10 weeks and
weighing 20–25 g were purchased from Shanghai SLAC laboratory
Animal Co., Ltd., Shanghai, China. The mice were housed in a
sterilized and specific pathogen free environment with a
temperature ~25°C, 12-h light/dark cycle and free access to
sterilized food and water. All procedures involving the mice were
approved by the Animal Ethics Committee of the Weifang People's
Hospital.
As reported previously (13), EPCs were extracted from mouse bone
marrow. Thighbone and shinbone were harvested from the leg of mice
after removal of muscle from entire leg for the extraction of
mononuclear cells using the density gradient method. Extracted
cells (1×106) were cultured in 24-well plates with
Endothelial Basal Medium-2 (EBM-2; Clonetics Corporation, San
Diego, CA, USA) at 37°C under 5% CO2 for 3 days.
Non-adherent cells were discarded after 3 days of culture.
Dil-acetylated-low density lipoprotein (Dil-Ac-LDL) and Ulex
europaeus agglutinin 1 (UEA-1) immunofluorescent staining (at
37°C for 1 h) were performed under a fluorescent microscope (Leica
Microsystems GmbH, Wetzlar, Germany). Meanwhile, cluster of
differentiation (CD)34, CD133 and vascular endothelial growth
factor receptor 2 (VEGFR2) were also measured by flow cytometry (BD
FACSCalibur, BD Biosciences, Franklin Lakes, NJ, USA) using
FITC-conjugated anti-CD34 antibody, PE-conjugated anti-CD133
antibody and APC-conjugated anti-VEGFR2 antibody.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA was extracted from bone marrow-derived EPCs
using TRIzol reagent (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). Reverse transcription was performed using M-MLV First-Strand
Synthesis reagent (Thermo Fisher Scientific, Inc.), and mRNA
expression levels of stromal cell-derived factor 1 (SDF-1) and
miR-126 were examined using the SYBR method (SYBR-Green
Quantitative RT-qPCR kit; QR0100; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). qPCR was performed using an iCycler
IQ® RT-PCR Detection System (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). PCR reaction was performed under the condition
of 95°C for 2 min, followed by 40 cycles of 95°C for 5 sec and 60°C
for 15 sec, and finally 72°C for 10 sec. The expression level of
SDF-1 was assessed with GAPDH serving as the internal control, and
miR-126 with U6 RNA as the internal control. The data was
quantified using 2−ΔΔCq method (14). The primer sequences are presented
in Table I.
 | Table I.Primer sequences of the three
genes. |
Table I.
Primer sequences of the three
genes.
|
| Primer (5′ to
3′) |
|---|
|
|
|
|---|
| Gene | Forward | Reverse |
|---|
| Stromal cell-derived
factor 1 |
GTGGTCGTGCTGGTCCTC |
CACACTTGTCTGTTGTTGTTCTTC |
| GAPDH |
ACTCCCACTCTTCCACCTTC |
CACCACCCTGTTGCTGTAG |
| U6 |
CTCGCTTCGGCAGCACA |
AACGCTTCACGAATTTGCGT |
Cell transfection
Bone marrow-derived EPCs or bEnd3 cells (mouse brain
endothelial cell line; American Type Culture Collection, Manassas,
VA, USA) (5×104 cells) were cultured in 6-well plates.
Scramble and angomir-126 were transfected into the cells using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocols. A 48-h culture was
maintained at 37°C and then transfected cells collected by
centrifugation at 125 × g for 5 min at room temperature.
Cell migration experiment
The cell migration experiment was performed via
Transwell assay. Transfected cells (2×104) were cultured
in the upper compartment of a Transwell plate. SDF-1 (100 ng/ml) or
C-X-C chemokine receptor type 7 (CXCR7; 100 ng/ml) neutralizing
antibody (MAB7167; R&D Systems, Inc., Minneapolis, MN, USA) was
added to the lower compartment of the Transwell plate. Cell
abundance was assessed using a fluorescence microscope
(magnification, ×40; 5 randomly selected visual fields).
Untransfected EPCs served as the control.
Western blot analysis
Western blotting was performed to examine the
expression level of CXCR-7 in the cells. Cells were lysed with RIPA
Lysis buffer (Thermo Fisher Scientific, Inc.) and protein was
determined by Pierce BCA Protein Assay kit (Thermo Fisher
Scientific, Inc.). Then, 10% SDS-PAGE gel electrophoresis was
performed with 20 µg protein being loaded per lane, and proteins
were transferred onto polyvinylidene difluoride (PVDF) membranes at
a constant voltage setting of 25 V for 2 h. The PVDF membranes were
blocked with skimmed milk (5%) and then incubated with CXCR7
antibody (PA3-069; 1:3,000; Thermo Fisher Scientific, Inc.) at 25°C
for 2 h or GAPDH antibody (ab37168; 1:3,000; Abcam) followed by
addition of HRP-conjugated goat anti-rabbit secondary antibody
(65–6120; 1:5,000; Thermo Fisher Scientific, Inc.). GAPDH served as
the internal control. Specific protein bands were examined by
chemiluminescence (Pierce ECL Western Blotting Substrate; Thermo
Fisher Scientific, Inc.).
Reporter gene assay
EPCs were cultured in 96-well plates at 37°C for 3
days. Co-transfection was performed with 20 ng angomir-126, 5 pmol
negative control and 100 ng pLUC SDF-1 3′-untranslated region (UTR)
wild type (WT) or mutant after 24 h of culture. Transfection was
performed with Lipofectamine 2000. The reporter gene assay was
performed using a Luciferase assay system (Promega Corporation,
Madison, WI, USA) according to the manufacturer's instructions.
Animal experiments
The animal experiments were approved by the Animal
Ethics Committee of the Weifang People's Hospital (Shandong,
China). The mice were randomly assigned to three groups (5 mice per
group), including a control group, EPC group and an EPC + miR-126
group. Middle cerebral artery occlusion was performed according to
a previously reported protocol (15). In the EPC and EPC + miR-126 groups,
EPCs (2×105) were infused via intravenous injection into
the tail after 2 h of occlusion. Isovolumetric phosphate-buffered
saline (PBS) was infused in the control group mice.
Neurological scoring
Then, 5-mark scoring was performed according to
previously reported protocol (16). Scoring was as follows: 0, normal
neurological function; 1, bending of contralateral trunk and
foreleg as the leg was lifted; 2, torsion of contralateral trunk as
leg was lifted, while normal at resting-state; 3, torsion of
contralateral trunk at rest; 4, loss of autonomic movement.
Examination of microvessel
density
Mice were sacrificed by cervical dislocation on day
2 and 7 after transfection. As reported previously (5), Fluoro-Jade and CD31 staining were
performed on cerebral sections. Microvessel density was assessed
using ImageJ software version 1.44 (National Institutes of
Health-Bethesda, MD, USA).
Statistical analysis
SPSS software version 17.0 (SPSS, Inc., Chicago, IL,
USA) was used for data processing. Measurement data are presented
as normal distribution to mean ± standard deviation. Student's
t-test and analysis of variance (one-way for comparison among
different groups, two-way for comparison among groups at different
time points) were performed to establish the statistical
significance and P<0.05 was considered to indicate a
statistically significant difference.
Results
Identification of EPCs
As presented in Fig. 1A
and B, EPCs were roughly circular and formed a cell colony
following 3 days of culture. EPCs were fusiform after 7 days of
culture. Flow cytometry indicated that CD34, CD133 and VEGFR2 were
expressed in EPCs (Fig. 1C).
Immunofluorescent staining indicated that Dil-ac-LDL and UEA-1 were
expressed in EPCs (Fig. 1D).
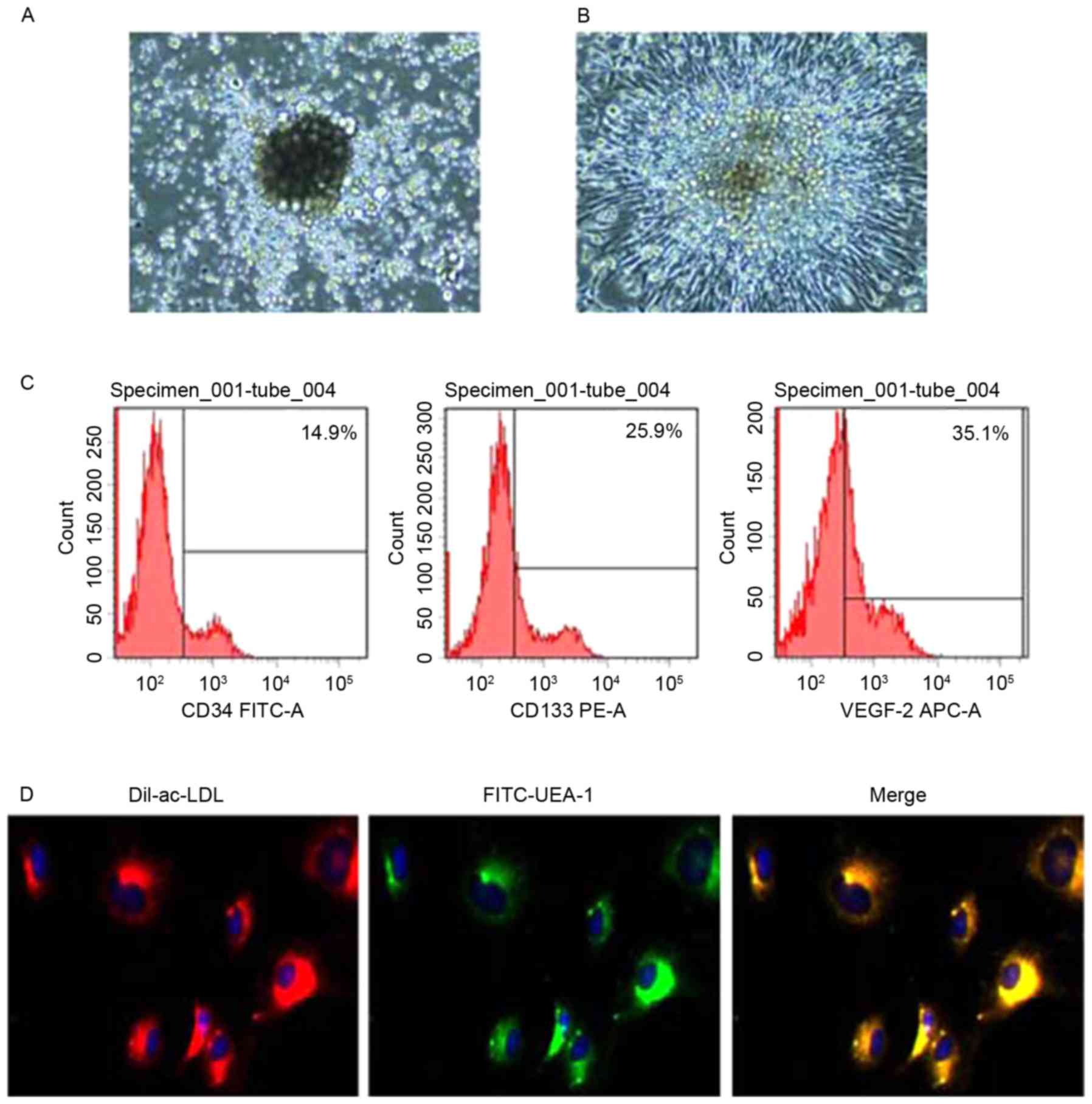 | Figure 1.Identification of endothelial
progenitor cells. Cellular morphology on days (A) 3 and (B) 7 of
culturing under a phase contrast microscope (magnification, ×40).
EPCs were roughly circular and formed a cell colony at day 3
culture. EPCs were fusiform at day 7 culture. (C) Expression levels
of CD34, CD133 and VEGFR2. (D) Expression levels of Dil-ac-LDL and
UEA-1 (magnification, ×40). Positive expressions of Dil-ac-LDL and
UEA-1 in cultured EPCs, indicating the cultured cells were EPCs.
CD, cluster of differentiation; FITC, fluorescein isothiocyanate;
PE, phycoerythrin; VEGFR2, vascular endothelial growth factor
receptor 2; APC, allophycocyanin; Ac-LDL, acetylated-low density
lipoprotein; UEA-1, Ulex europaeus agglutinin 1. |
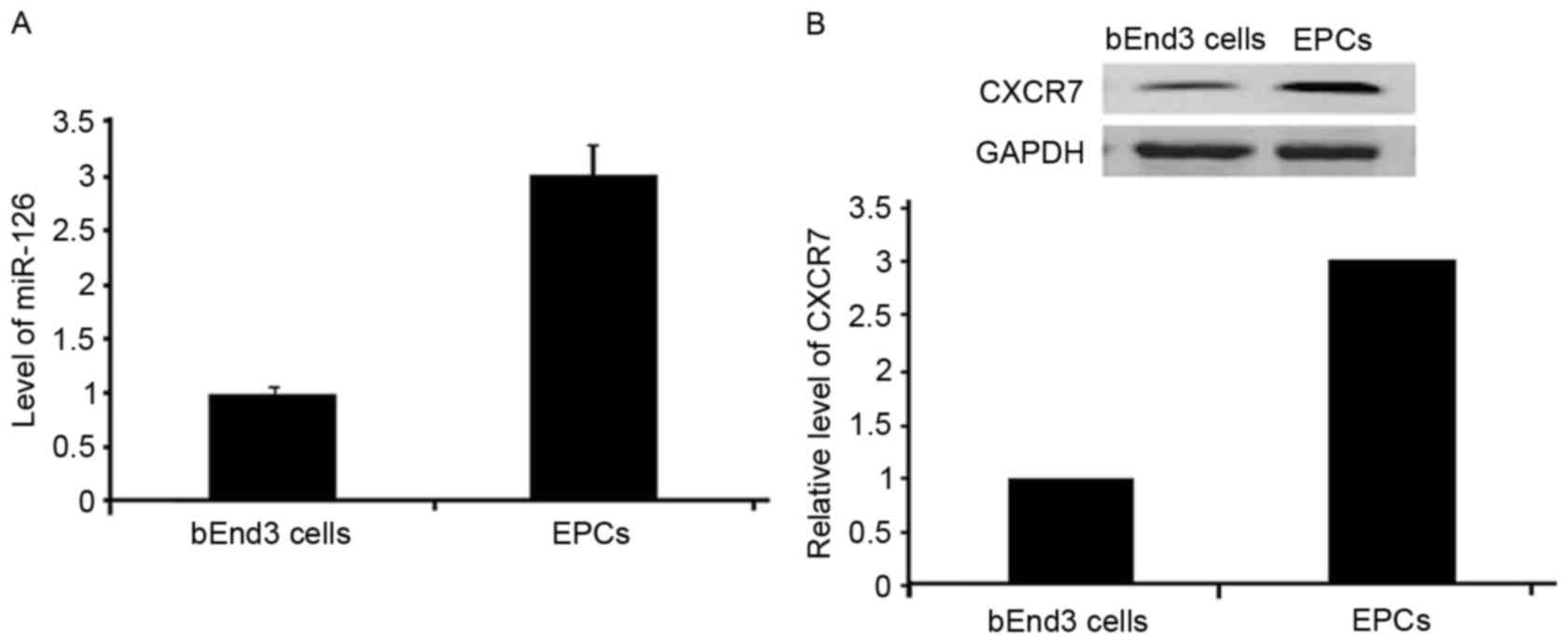
miR-126 and CXCR7 are expressed in
EPCs
Western blot analysis demonstrated CXCR7 protein
expression in EPCs. In addition, miR-126 and CXCR7 mRNA expression
was observed in EPCs, as verified by qPCR (Fig. 2).
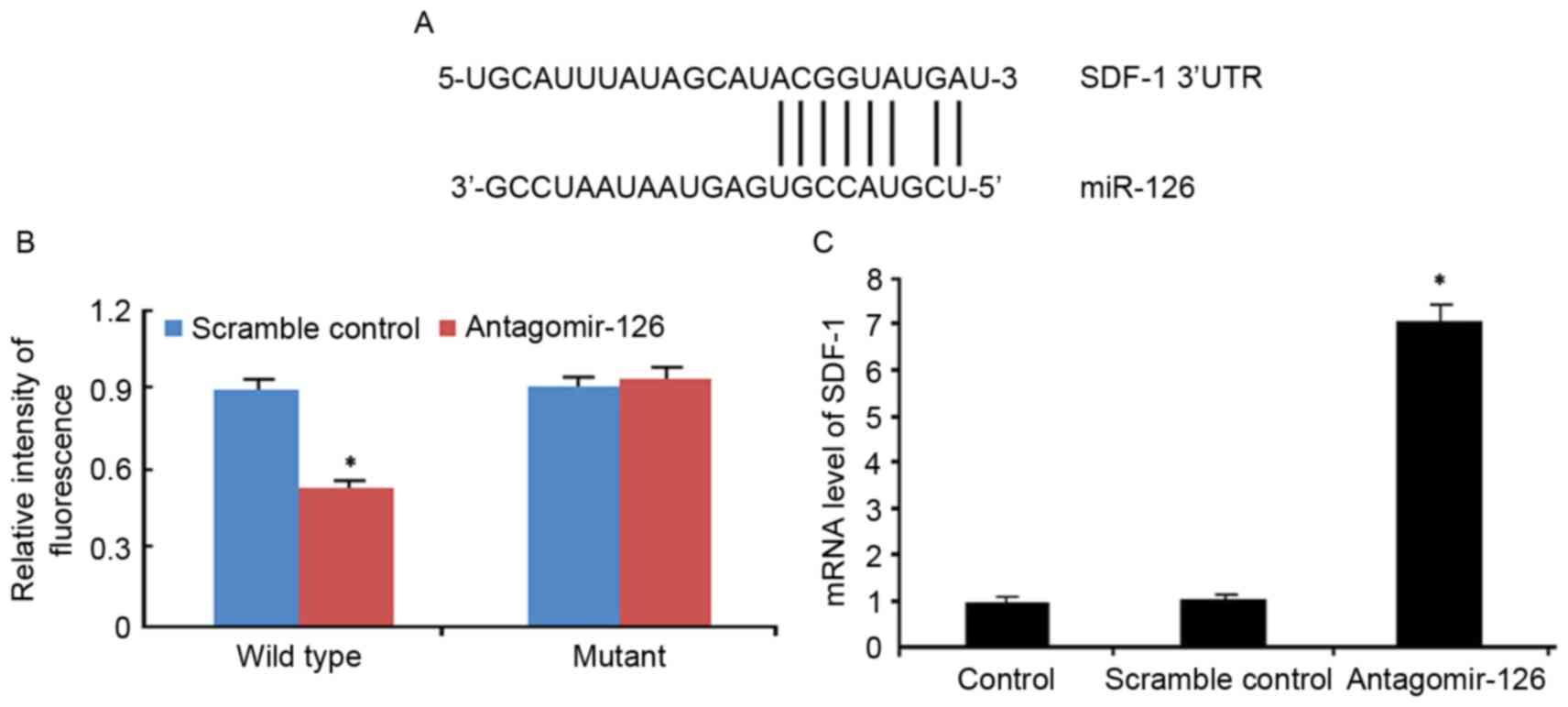
miR-126 regulates SDF-1 in EPCs
TargetScan software demonstrated that the 3′UTR
region of SDF-1 was complementary base paired with the sequence of
miR-126, indicating that SDF-1 is a target gene of miR-126
(Fig. 3A). The association between
SDF-1 and miR-126 was determined using a reporter gene assay and
RT-qPCR. The fluorescence intensity was decreased by 45% following
transfection of antagomir-126 into the WT, while no difference was
observed in the mutant (Fig. 3B).
The miR-126 angomir significantly reduced the expression level of
SDF-1, which was verified by RT-qPCR (Fig. 3C).
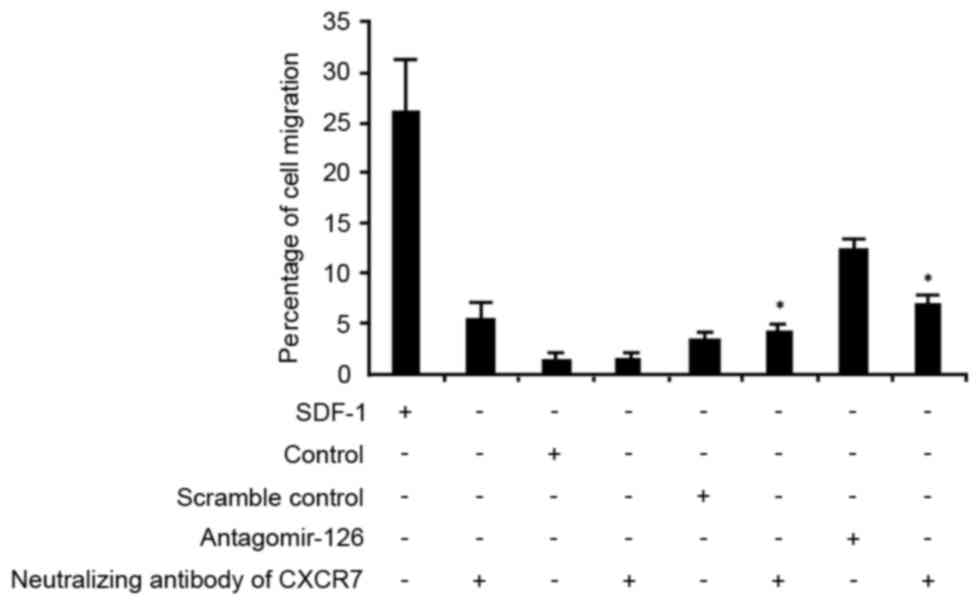
CXCR-7 inhibition abrogates miR-126
angomir-induced migration of EPCs
The Transwell assay demonstrated that inhibition of
CXCR7 repressed migration of the miR-126 angomir-transfected cells,
indicating that CXCR7 was involved in miR-126-induced migration of
EPCs (Fig. 4).
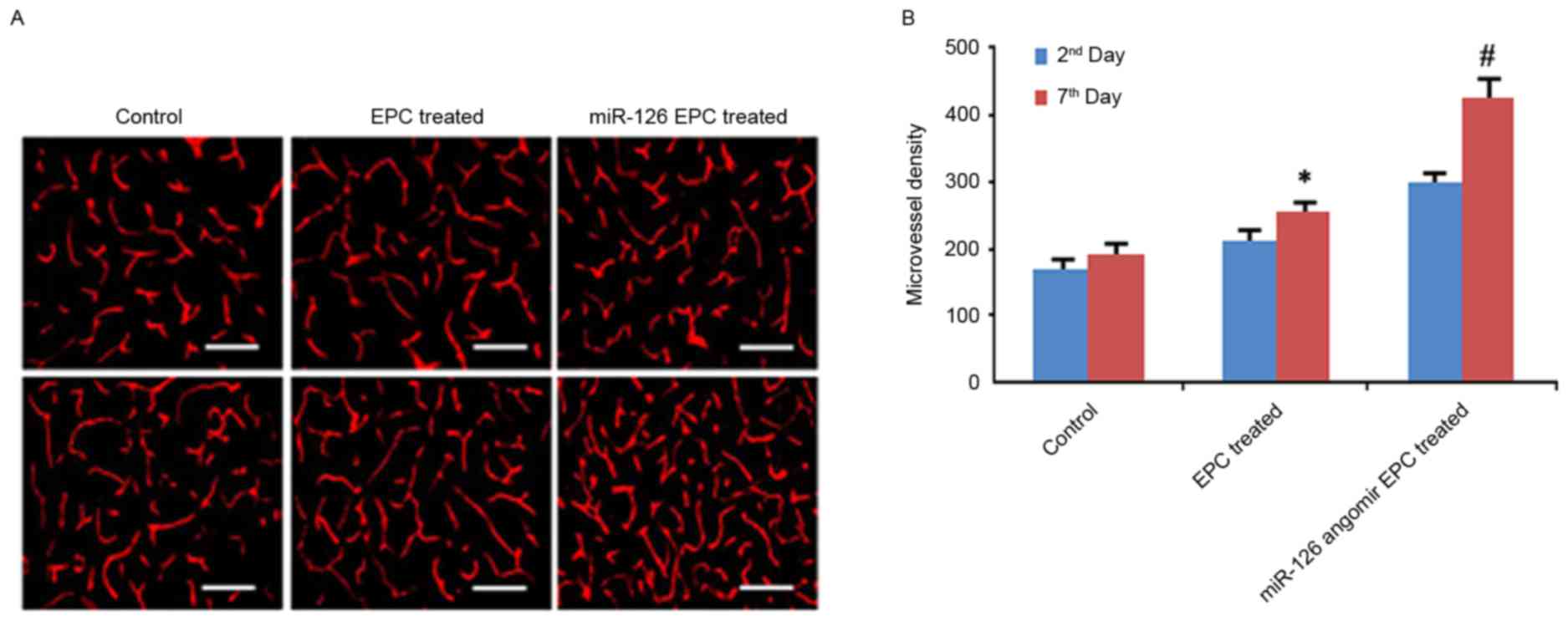
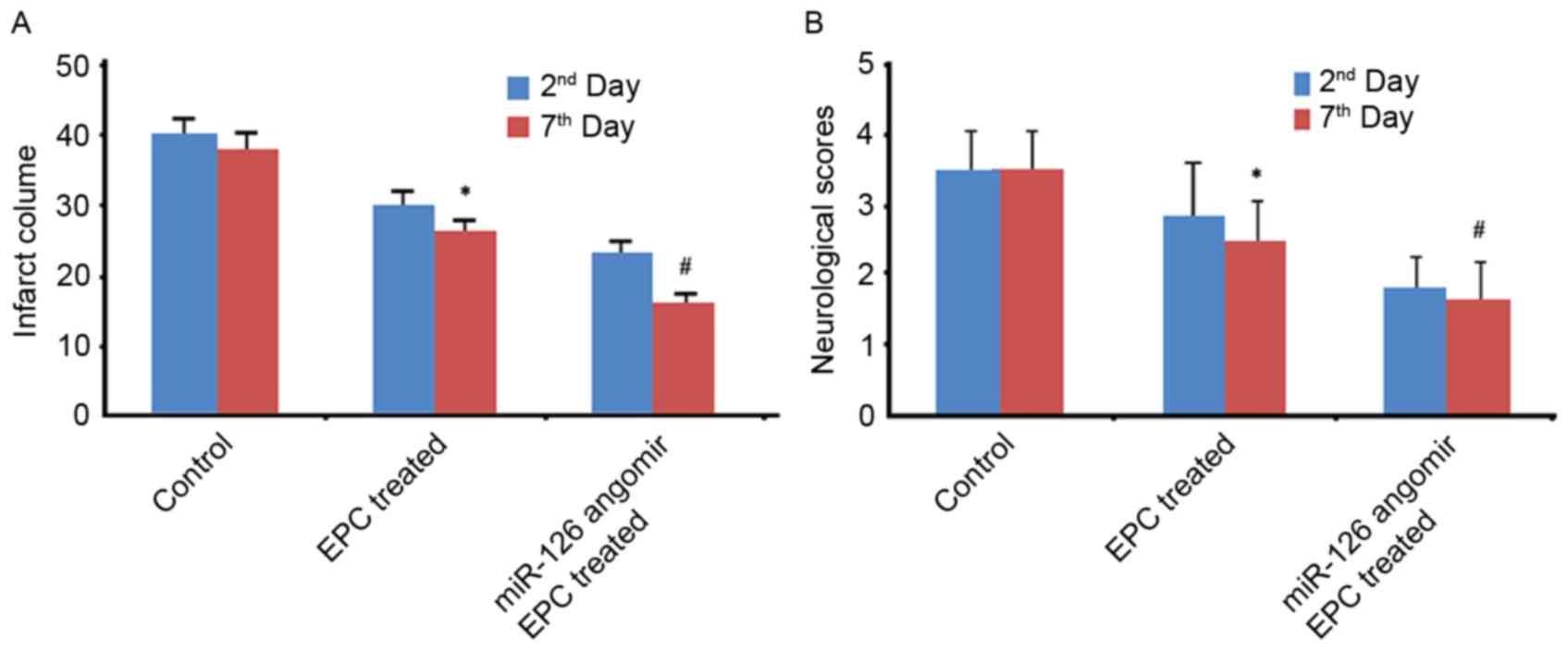
miR-126 angomir improves the efficacy
of EPC treatment in mice with middle cerebral artery occlusion
In the animal experiments, miR-126
angomir-transfected EPCs demonstrated significantly increased
microvessel densities at 2 and 7 days after middle cerebral artery
occlusion (Fig. 5A and B). In
addition, miR-126 angomir-transfected EPCs improved the prognosis
of neurological function, as verified by increased neurological
scores and decreased infarct volumes (Fig. 6).
Discussion
Antagomir, as a group of oligonucleotides, is a type
of chemically engineered single-strand mRNA inhibitor that
efficiently blocks miRNA regulation of target gene expression. The
antisense strand consists of two phosphorothioates at the 5′ end,
four phosphorothioates, four cholesterol groups at the 3′ end, with
2′-methoxy modification (17).
Previous studies identified that miR-126 influenced angiogenesis in
gene knock-out mice models or angomir treatment models, indicating
that miR-126 may have attenuated structural damage to vessels
(18–20). The present study demonstrated that
miR-126 angomir contributed to EPC treatment.
SDF-1, also termed CXCL12, has been implicated in
various types of disease, including numerous cancer types (21) and coronary artery disease (22). SDF-1 exerts a significant
regulatory role in various processes, such as morphogenesis,
angiogenesis and immune responses, and is considered to be a
potential target for drug development (23). Furthermore, CXCR7, also termed
atypical chemokine receptor 3 or G-protein coupled receptor 159, is
a high affinity receptor of CXCL12, and has recently been
identified to be involved in the growth, migration, chemotaxis,
adhesion and spreading of tumors (24). The current study elucidated that
miR-126 exerted a protective effect of EPCs during angiogenesis via
the SDF-1/CXCR7 signaling pathway. In addition, the neutralizing
antibody of CXCR7 blocked the migration-induced effect of the
miR-126 angomir on EPCs in vitro. Furthermore, infusion of
the miR-126 angomir-treated EPCs was more effective than routine
EPC treatment, which improved neurological scores, increased
microvessel density and decreased infarct volume.
Interactions between miR-126 and SDF-1 has been
demonstrated to increase elimination of endothelial apoptosis
bodies induced by miR-126 (25). A
potential mechanism is that miR-126 enhances the expression level
of SDF-1 via inhibiting regulators of G protein signalling
(25). The present study confirmed
that silencing of miR-126 was associated with changes of SDF-1
expression levels, indicating miR-126 as a potential biomarker for
clinical treatment. miRNAs have been associated with the occurrence
and progression of diseases (26).
Furthermore, circulating miRNAs influence tumorigenesis (27) and cardiovascular diseases (28,29).
Endothelial injury is a significant risk factor for patients with
cardiovascular diseases. miRNAs secreted by EPCs have been
demonstrated as cardiovascular disease markers in the clinical
setting (30). Clinical trials
have indicated that miR-126 expression levels decreased in acute
coronary syndrome patients and diabetes patients (31,32).
Thus, miR-126-associated signaling pathways have specific signals
for EPCs, and selective activation of miR-126 was significant
during EPC treatment.
In addition, miR-126 inhibits the expression of
SDF-1 and stimulates angiogenesis when vessels are injured.
Furthermore, downregulation of miR-126 inhibits angiogenesis in
aging EPCs or EPCs with function disorders. The current study
indicated that miR-126 improved the function of ECPs and
angiogenesis via the downregulation of SDF-1.
In conclusion, miR-126 indeed improved migration of
EPCs via the miR-126/SDF-1/CXCR7 signaling pathway. Furthermore,
the miR-126 angomir improved efficacy of EPC treatment, and may
present as a potential therapeutic target for cardiovascular
diseases. The main study limitation is that the EPC used in the
present study was from mouse, not human, whether this effect also
exists in human EPCs remains unclear and requires further
investigation.
References
|
1
|
Ma F, Morancho A, Montaner J and Rosell A:
Endothelial progenitor cells and revascularization following
stroke. Brain Res. 1623:150–159. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen J, Chen J, Chen S, Zhang C, Zhang L,
Xiao X, Das A, Zhao Y, Yuan B, Morris M, et al: Transfusion of
cxcr4-primed endothelial progenitor cells reduces cerebral ischemic
damage and promotes repair in db/db diabetic mice. PloS One.
7:e501052012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Werner N and Nickenig G: Influence of
cardiovascular risk factors on endothelial progenitor cells:
Limitations for therapy? Arterioscler Thromb Vasc Biol. 26:257–266.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Quirici N, Soligo D, Caneva L, Servida F,
Bossolasco P and Deliliers GL: Differentiation and expansion of
endothelial cells from human bone marrow cd133(+) cells. Br J
Haematol. 115:186–194. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen J, Chen S and Chen Y, Zhang C, Wang
J, Zhang W, Liu G, Zhao B and Chen Y: Circulating endothelial
progenitor cells and cellular membrane microparticles in db/db
diabetic mouse: Possible implications in cerebral ischemic damage.
Am J Physiol Endocrinol Metab. 301:E62–E71. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yue WS, Lau KK, Siu CW, Wang M, Yan GH,
Yiu KH and Tse HF: Impact of glycemic control on circulating
endothelial progenitor cells and arterial stiffness in patients
with type 2 diabetes mellitus. Cardiovasc Diabetol. 10:1132011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Miller-Kasprzak E and Jagodziński PP:
Endothelial progenitor cells as a new agent contributing to
vascular repair. Arch Immunol Ther Exp (Warsz). 55:247–259. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chong MS, Ng WK and Chan JK: Concise
review: Endothelial progenitor cells in regenerative medicine:
Applications and challenges. Stem Cells Transl Med. 5:530–538.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fan Y, Shen F, Frenzel T, Zhu W, Ye J, Liu
J, Chen Y, Su H, Young WL and Yang GY: Endothelial progenitor cell
transplantation improves long-term stroke outcome in mice. Ann
Neurol. 67:488–497. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chekulaeva M and Filipowicz W: Mechanisms
of miRNA-mediated post-transcriptional regulation in animal cells.
Curr Opin Cell Biol. 21:452–460. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Urbich C, Kuehbacher A and Dimmeler S:
Role of microRNAs in vascular diseases, inflammation, and
angiogenesis. Cardiovasc Res. 79:581–588. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Marrotte EJ, Chen DD, Hakim JS and Chen
AF: Manganese superoxide dismutase expression in endothelial
progenitor cells accelerates wound healing in diabetic mice. J Clin
Invest. 120:4207–4219. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen S, Li G, Zhang W, Wang J, Sigmund CD,
Olson JE and Chen Y: Ischemia-induced brain damage is enhanced in
human renin and angiotensinogen double-transgenic mice. Am J
Physiol Regul Integr Comp Physiol. 297:R1526–R1531. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Girouard H, Lessard A, Capone C, Milner TA
and Iadecola C: The neurovascular dysfunction induced by
angiotensin II in the mouse neocortex is sexually dimorphic. Am J
Physiol Heart Circ Physiol. 294:H156–H163. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Scherr M, Venturini L, Battmer K,
Schaller-Schoenitz M, Schaefer D, Dallmann I, Ganser A and Eder M:
Lentivirus-mediated antagomir expression for specific inhibition of
miRNA function. Nucleic Acids Res. 35:e1492007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sessa R, Seano G, di Blasio L, Gagliardi
PA, Isella C, Medico E, Cotelli F, Bussolino F and Primo L: The
miR-126 regulates angiopoietin-1 signaling and vessel maturation by
targeting p85β. Biochim Biophys Acta. 1823:1925–1935. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
van Solingen C, Seghers L, Bijkerk R,
Duijs JM, Roeten MK, van Oeveren-Rietdijk AM, Baelde HJ, Monge M,
Vos JB, de Boer HC, et al: Antagomir-mediated silencing of
endothelial cell specific microRNA-126 impairs ischemia-induced
angiogenesis. J Cell Mol Med. 13:1577–1585. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang S, Aurora AB, Johnson BA, Qi X,
McAnally J, Hill JA, Richardson JA, Bassel-Duby R and Olson EN: The
endothelial-specific microRNA miR-126 governs vascular integrity
and angiogenesis. Dev Cell. 15:261–271. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guo JC, Li J, Zhou L, Yang JY, Zhang ZG,
Liang ZY, Zhou WX, You L, Zhang TP and Zhao YP: CXCL12-CXCR7 axis
contributes to the invasive phenotype of pancreatic cancer.
Oncotarget. 7:62006–62018. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mega JL, Stitziel NO, Smith JG, Chasman
DI, Caulfield M, Devlin JJ, Nordio F, Hyde C, Cannon CP, Sacks F,
et al: Genetic risk, coronary heart disease events, and the
clinical benefit of statin therapy: An analysis of primary and
secondary prevention trials. Lancet. 385:2264–2271. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Neesse A and Ellenrieder V:
NEMO-CXCL12/CXCR4 axis: A novel vantage point for antifibrotic
therapies in chronic pancreatitis? Gut. 66:211–212. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun X, Cheng G, Hao M, Zheng J, Zhou X,
Zhang J, Taichman RS, Pienta KJ and Wang J: CXCL12/CXCR4/CXCR7
chemokine axis and cancer progression. Cancer Metastasis Rev.
29:709–22. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zernecke A, Bidzhekov K, Noels H,
Shagdarsuren E, Gan L, Denecke B, Hristov M, Köppel T, Jahantigh
MN, Lutgens E, et al: Delivery of microRNA-126 by apoptotic bodies
induces CXCL12-dependent vascular protection. Sci Signal.
2:ra812009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mitchell PS, Parkin RK, Kroh EM, Fritz BR,
Wyman SK, Pogosova-Agadjanyan EL, Peterson A, Noteboom J, O'Briant
KC, Allen A, et al: Circulating microRNAs as stable blood-based
markers for cancer detection. Proc Natl Acad Sci USA. 105:pp.
10513–10518. 2008; View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lawrie CH, Gal S, Dunlop HM, Pushkaran B,
Liggins AP, Pulford K, Banham AH, Pezzella F, Boultwood J,
Wainscoat JS, et al: Detection of elevated levels of
tumour-associated microRNAs in serum of patients with diffuse large
B-cell lymphoma. Br J Haematol. 141:672–675. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Corsten MF, Dennert R, Jochems S,
Kuznetsova T, Devaux Y, Hofstra L, Wagner DR, Staessen JA, Heymans
S and Schroen B: Circulating microRNA-208b and microRNA-499 reflect
myocardial damage in cardiovascular disease. Circ Cardiovasc Genet.
3:499–506. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ai J, Zhang R, Li Y, Pu J, Lu Y, Jiao J,
Li K, Yu B, Li Z, Wang R, et al: Circulating microRNA-1 as a
potential novel biomarker for acute myocardial infarction. Biochem
Biophys Res Commun. 391:73–77. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bronze-da-Rocha E: MicroRNAs expression
profiles in cardiovascular diseases. Biomed Res Int.
2014:9854082014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fichtlscherer S, De Rosa S, Fox H,
Schwietz T, Fischer A, Liebetrau C, Weber M, Hamm CW, Röxe T,
Müller-Ardogan M, et al: Circulating microRNAs in patients with
coronary artery disease. Circ Res. 107:677–684. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zampetaki A, Kiechl S, Drozdov I, Willeit
P, Mayr U, Prokopi M, Mayr A, Weger S, Oberhollenzer F, Bonora E,
et al: Plasma microRNA profiling reveals loss of endothelial
miR-126 and other microRNAs in type 2 diabetes. Circ Res.
107:810–817. 2010. View Article : Google Scholar : PubMed/NCBI
|