Introduction
Epilepsy is a serious, long-term and debilitating
brain disorder, identifiable by paroxysmal bursts of activity
within cortical neurons, which affects approximately 1% of the
world population (1,2). Although epilepsy frequently develops
in childhood, its symptoms may last a lifetime (3). Epilepsy is characterized by a
predisposition to recurring episodes of seizures and abnormally
synchronized neuronal discharges with the potential to disrupt the
function of the brain region from which they originate, or through
which they pass (1). Despite
currently available antiepileptic drugs and surgeries for focal
epilepsy being available, both clinical treatments have some side
effects, including headache, dizziness, fatigue and at axia.
Additionally, >20% of all patients with epilepsy continue to
have the symptoms following treatment (4,5).
Therefore, the identification of new biological markers for
treatment against epilepsy is of great importance. In recent years,
there has been a number of studies investigating molecular causes
of epilepsy (6–9). A previous study suggested the
involvement of microRNAs (miRNAs) in the lesion of epilepsy
(6). miRNAs are single-stranded
molecules, between 18–24 nucleotides in length, that block the
expression of protein-coding genes at the post transcriptional
level by directing translational repression or mRNA
destabilization, or a combination of the two (7). Many post transcriptional
inflammation-associated miRNAs, such as the brain-enriched miRNA
(miR)-146a, have been previously identified to be involved in the
regulation of inflammatory responses in epileptic rats and adult
patients (8,9).
The nuclear factor-κB (NF-κB) pathway is an
important intracellular signaling pathway involved in the early
stress response (10–12). A previous study suggested that
NF-κB acts as a key point of convergence for multiple stress
signals, including intracellular Ca2+ changes,
pro-inflammatory cytokines and oxidative stress (10). The pathway is also involved in the
modulation of neuronal excitability and seizure susceptibility
(13). Additionally, downstream
events activated by NF-κB, such as inflammation and oxidative
stress, are also known to be activated by seizure activity
(14,15).
The present study induced an epilepsy animal model
using the lithium-pilocarpine method. The expression of miR-146a
was detected using reverse transcription-semi-quantitative
polymerase chain reaction (RT-sqPCR). The expression levels of
pro-inflammatory cytokines, including interleukin (IL)-1β, IL-6 and
tumor necrosis factor-α (TNF-α), were determined using an ELISA.
Additionally, the protein expression levels of P-glycoprotein
(P-gp), B-cell lymphoma-2 (Bcl-2)/Bcl2-associated X (Bax) and
phosphorylated (p)-P65/P65 were quantified using western blot
analysis. The findings of the present study may aid the evaluation
of miR-146a regulation as a potential new anti-epilepsy therapy,
and suggests that miR-146a antagonist treatment of epilepsy may be
associated with NF-κB pathway.
Materials and methods
Animals
A total of 72 adult male Sprague-Dawley rats
(Weitonglihua Biomart, Beijing, China), weighing 180–220 g, 6–8
weeks were housed in a temperature (24±0.5°C) and 12-h light-dark
cycle with a 50–60% humidity and had free access to food and water.
Prior to the experiments, the animals were allowed to habituate to
the housing facilities for 1 week. The present study was conducted
with approval from the Animal Ethics Committee of the affiliated
Yantai Yuhuangding Hospital of Qingdao University (Yantai,
China).
Epilepsy induction
In order to induce status epilepticus (SE), rats
were intraperitoneally (i.p.) injected 127 mg/kg with lithium
chloride (CAS: 7447-41-8; Santa Cruz Biotechnology, Inc., Dallas,
TX, USA), followed 20 h later by anatropine (Tianjin Kaitong
Chemical Reagent Co., Ltd., Tianjin, China) i.p. (0.1 mg/kg)
injection and 30 min later by pilocarpine hydrochloride (cat. no.
P6503-5G, Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) treatment
(40 mg/kg; i.p.). SE was characterized by continuous limbic
seizures 30 min after the last treatment. The severity of seizures
was evaluated using the Racine scale (16): 0, No response; 1, motor arrest and
twitching vibrissae; 2, chewing and head bobbing; 3, forelimb
clonus; 4, forelimb clonus and rearing; and 5, rearing and falling.
Rats exhibiting continuous seizures at stage V of the Racine scale
were used for the subsequent experiments, were then divided into
two sections. Section I, consisted of two groups: i) 8 Naive rats
were used as control; and ii) 40 SE rats were used to evaluate the
expression of miR-146a. Section II, consisted of three groups: i) 8
naive rats were used as control; ii) SE model group n=8; and iii) 1
nmol miR-146a inhibitor-treated (Guangzhou RiboBio Co., Ltd.,
Guangzhou, China) group (IN group). The rats belonging to the IN
group were treated with 1 nmol miR-146a inhibitor 2 h
post-pilocarpine injection.
RT-sqPCR
Animals were sacrificed following deep anesthesia
with an i.p. injection of 10% chloral hydrate (500 mg/kg) at day 1,
3, 7, 14 and 30 following-SE induction. Following decapitation, the
brain was removed as previously described (17) for RNA isolation. Total RNA was
extracted with TRIzol reagent (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) and both the concentration and the purity
of RNA were then determined at 260/280 nm using a NanoDrop
spectrophotometer. Isolated RNA (1 µg) was then subjected to cDNA
synthesis. cDNA synthesis was conducted in a 14 µl reaction buffer,
containing 1 µl reverse transcriptase (50 U) and 1 µl ologo (dT)
primer, according to manufacturer's protocol (Takara Biotechnology
Co., Ltd., Dalian, China). The following temperature protocol was
used for the reverse transcription: 37°C for 15 min, 85°C for 5 sec
and 4°C until subsequent use for qPCR. The sequences of the primers
for qPCR wereas follows: miRNA-146a, forward (F),
5′-CAGTGCGTGTCGTGGAGT-3′ and reverse (R),
5′-GGGTGAGAACTGAATTCCA-3′; U6 F, 5′-GCTTCGGCAGCACATATACTAAAAT-3′
and R, 5′-CGCTTCACGAATTTGCGTGTCAT-3′. Reaction conditions were as
follow: 95°C for 30 sec, followed by 35 cycles of 95°C for 10 sec,
60°C for 15 sec and 72°C for 15 sec. Each 20 µl reaction system
consisted of 2 µl cDNA, 10 µl SYBR Premix Ex Taq II (Sangong
Biotech Co., Ltd., Shanghai, China) and 10 µmol/l of both forward
and reverse primers (Sangong Biotech Co., Ltd.). U6 was used to
normalize the mRNA. The results were analyzed on an 1% agarose gel
(Sangong Biotech Co, Ltd.) with ethidium bromide and the relative
intensity of the bands was determined using Image J software
(National Institutes of Health, Bethesda, MD, USA).
Histopathological analysis
The hippocampus was dehydrated in increasing
concentrations of ethanol, rinsed with Histoclear (National
Diagnostics, Atlanta, GA, USA), embedded in paraffin and then cut
on a microtome into 5-µm thick slices. Following dewaxing, slides
were boiled for 48 h at 45°C and then for 1 h at 60°C. The sections
were stained with hematoxylin and eosin for 10 and 5 min
respectively at room temperature. All stained sections were then
assessed under a light microscope (magnification, ×400).
Western blot analysis
The brain tissue samples were homogenized in lysis
buffer (20 mMTris, 1% Triton-X-100, 0.05% SDS, 5 mg of sodium
deoxycholate, 150 mMNaCl and 1 mM PMSF) containing a mixture of
protease and phosphatase inhibitors. The protein concentrations
were then determined using a Bicinchoninic acid Protein Assay
reagent kit (Beyotime Institute of Biotechnology, Shanghai, China).
Equal quantity of protein (40 µg/lane) were separated by 10%
SDS-PAGE and then transferred to polyvinylidenedifluoride
membranes. The nonspecific binding of antibodies was blocked with
5% non-fat dried milk in PBS, then incubated with the following
primary antibodies: Rabbit monoclonal Bcl-2 (1:500; cat. no. 3498,
Cell Signaling Technology, Inc., Danvers, MA, USA), rabbit
monoclonal Bax (1:500; cat. no. 5023; Cell Signaling Technology,
Inc.), rabbit monoclonal P-gp (1:500; cat. no. ab170964; Abcam,
Cambridge, UK), rabbit monoclonal p-P65 (1:500; cat. no. 3033; Cell
Signaling Technology, Inc.), rabbit monoclonal P65 (1:500; cat. no.
59674; Cell Signaling Technology, Inc.) at 4°C overnight. Following
incubation with the horseradish peroxidase-conjugated goat
anti-rabbit immunoglobulin G secondary antibody (1:5,000; cat. no.
SC-2004; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) at 4°C
for 1 h and developed with electrochemiluminescence (ECL) Western
Blotting Substrate (BD Biosciences, Franklin Lakes, NJ, USA). Data
was analyzed using Quantity One 1D image analysis software (version
4.4; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Enzyme-linked immunosorbent assay
(ELISA)
ELISA kits for rat IL-1β (cat. no. PI303; Beyotime
Institute of Biotechnology), TNF-α (cat. no. PT516; Beyotime
Institute of Biotechnology) and IL-6 (cat. no. PI328; Beyotime
Institute of Biotechnology) were used to determine cytokine levels
in cerebrospinal fluid (CSF) following the manufacturer's
protocols. The plates were quantified at 450 nm using amicroplate
reader.
Statistical analysis
Data are expressed as mean ± standard deviation.
Statistical differences were evaluated using SPSS version 19.0 (IBM
Corporation, Armonk, NY, USA). Statistical analysis was performed
using one-way analysis of variance. P<0.05 was considered to
indicate a statistically significant difference.
Results
RT-sqPCR
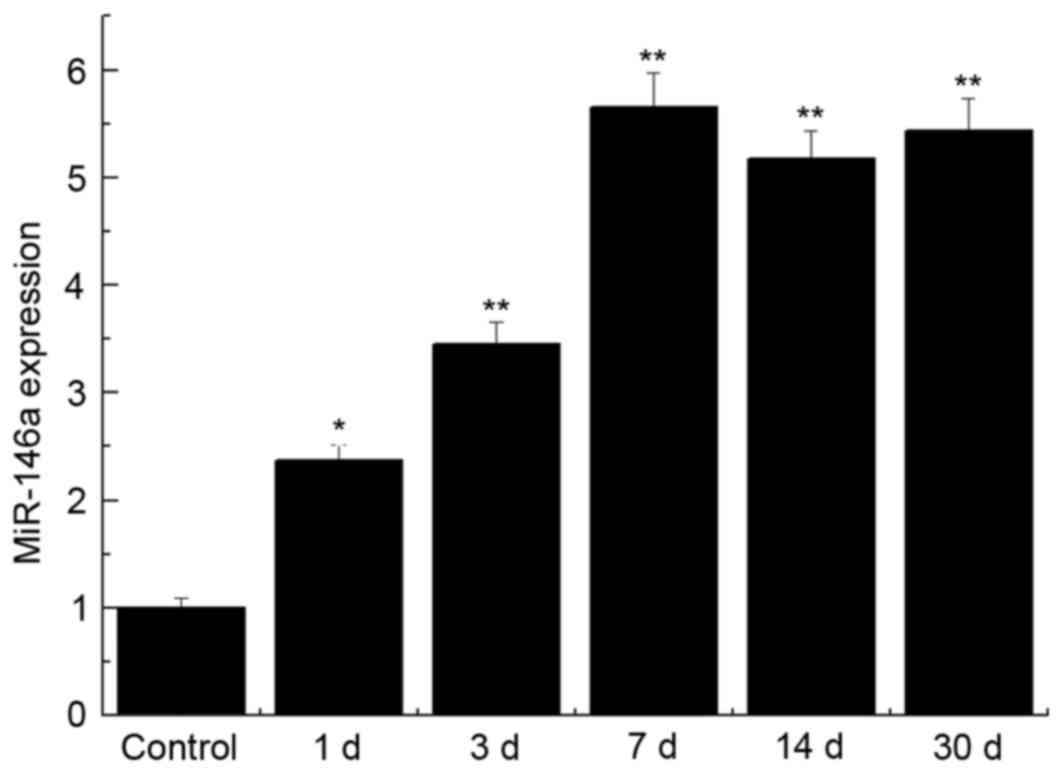
miR-146a expression levels were significantly
upregulated in the brain tissue of the SE group when compared with
control group at day 1, 3, 7, 14 and 30 (P<0.05; P<0.01;
Fig. 1). The upregulation of
miR-146a expression reached a maximum at the day 7 following
pilocarpine treatment (P<0.01). The rats were treated with the
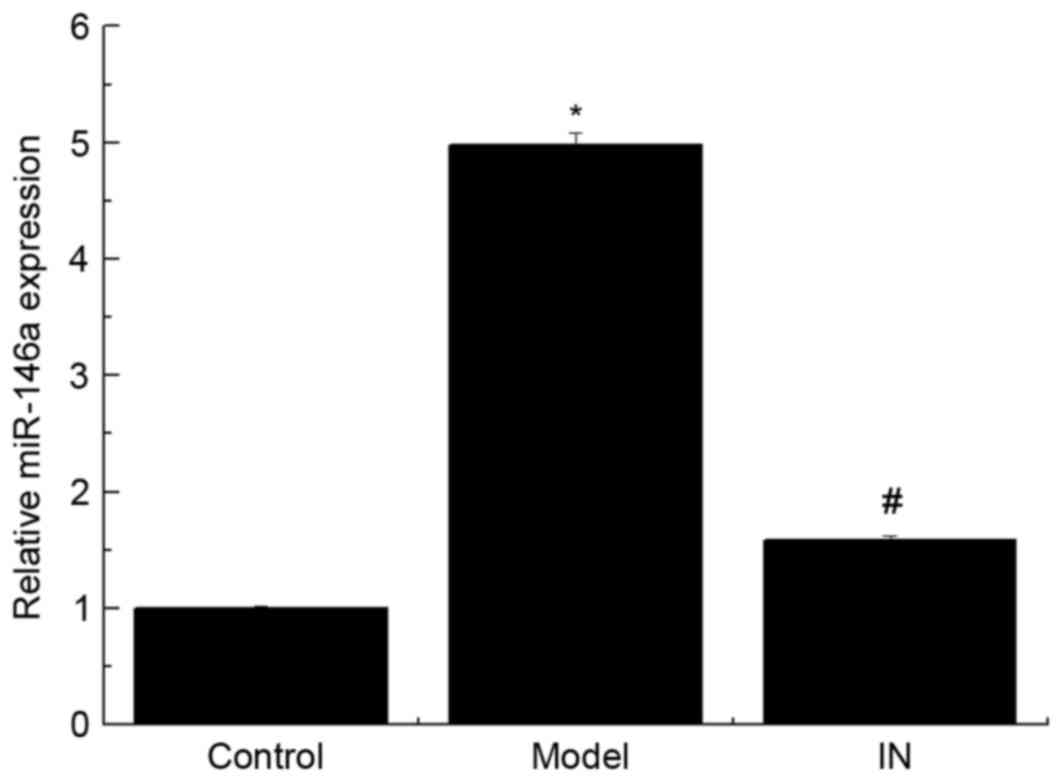
miR-146a inhibitor, the expression of miR-146a was significantly
reduced (Fig. 2; P<0.01).
However, no significant difference between the control group and
the IN group was identified (P>0.05).
Histopathological analysis
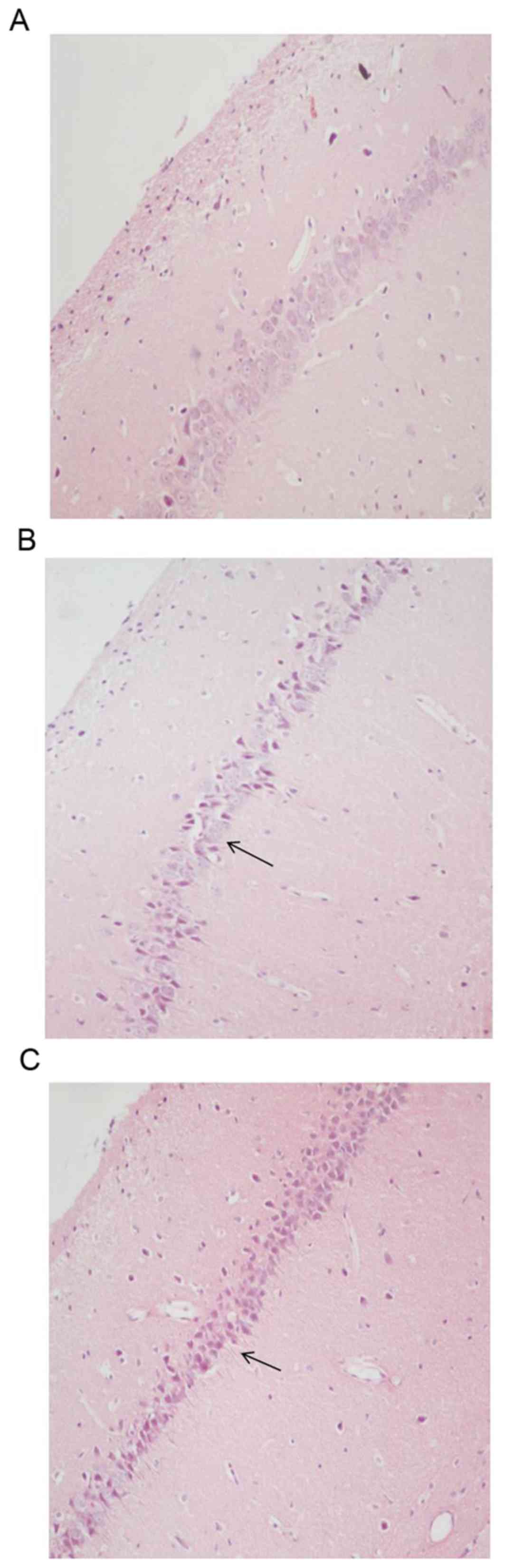
Examination of hippocampal CA3 and CA4 regions in
the control group revealed compactly arranged and healthy pyramidal
cells with clear nuclei, and intact cell membranes (Fig. 3A). Compared to the control group,
hematoxylin and eosin staining of the model group 7 days
post-pilocarpine treatment displayed numerous shrunken neurons,
neuronal cell loss, disordered tissue structure in hippocampal CA3
and CA4 are as and abnormal cell morphology (Fig. 3B). Rats treated with the miR-146a
inhibitor exhibited significantly reduced levels of neuronal cell
loss and other morphological signs of damage (Fig. 3C).
Western blot analysis
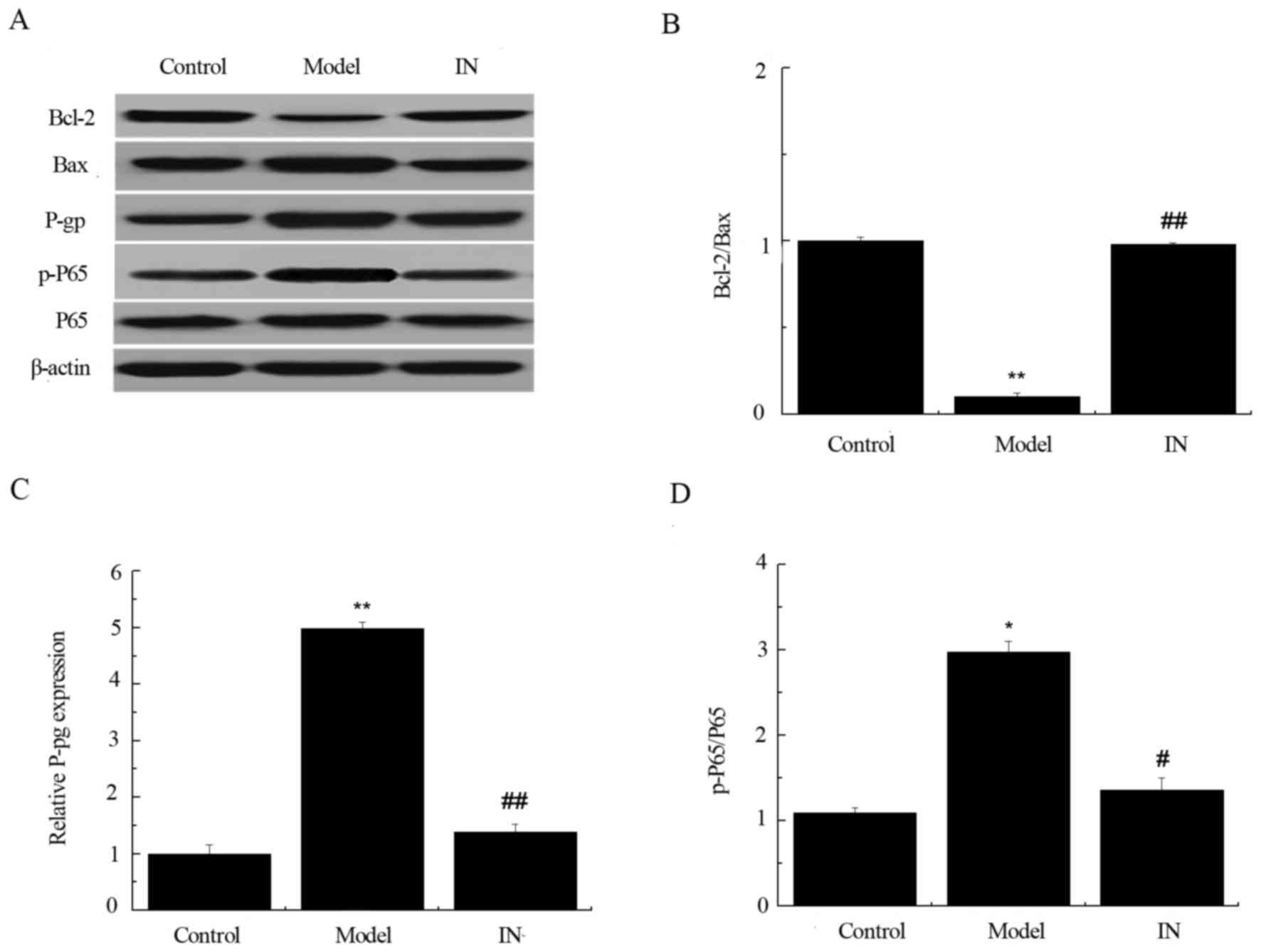
P-gp and p-P65/P65 expression was significantly
increased in the hippocampus of the model group compared with the
control group (Fig. 4; P<0.01).
Compared with model group, the miR-146a inhibitor-treated group (IN
group) significantly downregulated P-gp and p-P65/P65 expression.
Conversely, the expression level ratio of Bcl-2/Baxin hippocampal
tissues in the model group was significantly lower when compared
withthe control group (P<0.01; Fig.
4B). The treatment of miR-146a antagonist significantly
reversed this as Bcl-2/Bax ratio was increased in the IN group when
compared with model group (P<0.01). These findings suggest that
the miR-146a inhibitor reduces the expression of P-gp and p-P65/P65
and increases the expression of Bcl-2/Bax in rats.
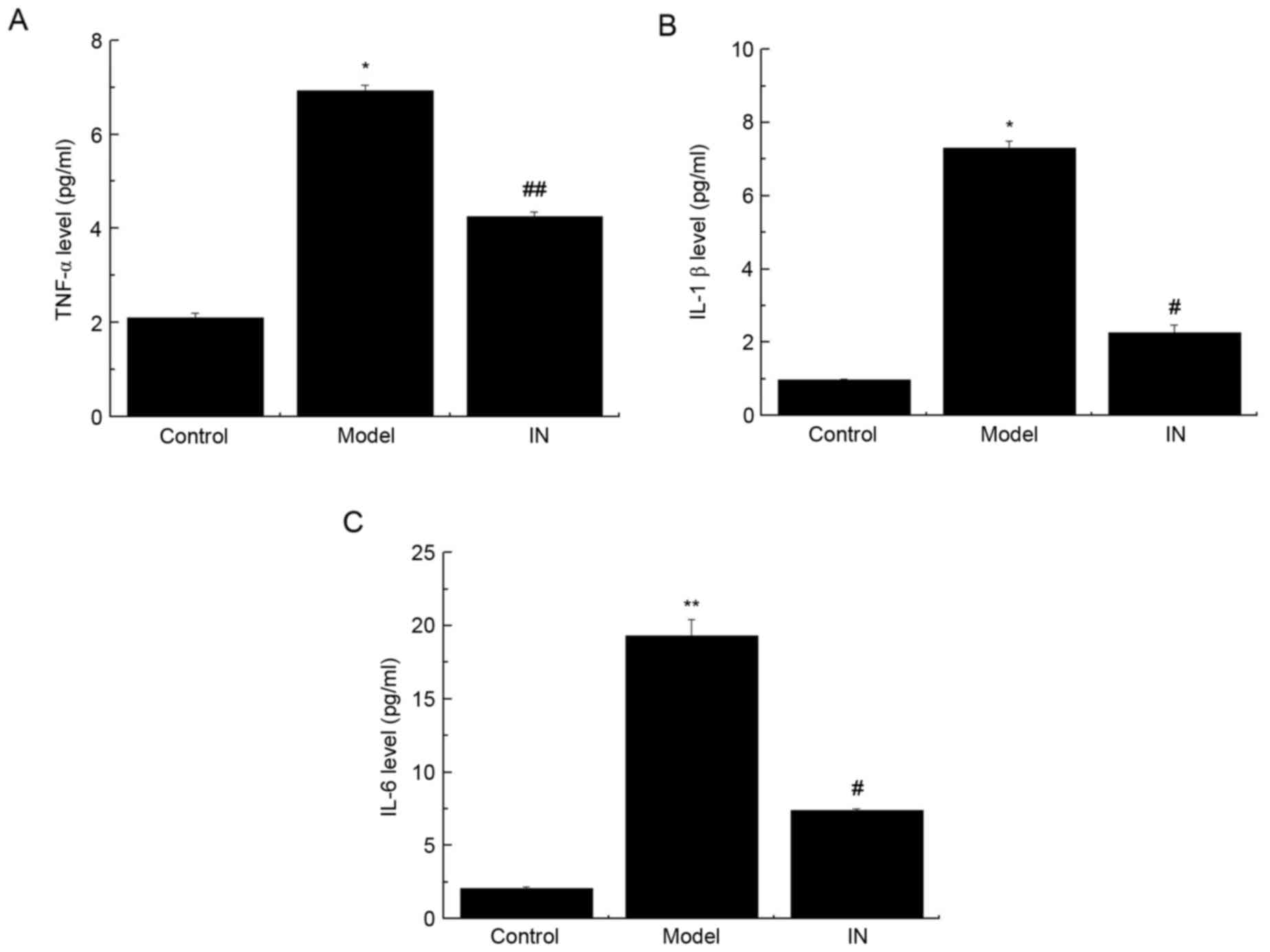
Enzyme-linked immunosorbent assay
(ELISA)
Previous studies have demonstrated that
pro-inflammatory cytokines are involved in SE (18–20).
Therefore, the current study aimed to investigate the alterations
in pro-inflammatory cytokine expression levels in CSF. As presented
in Fig. 5, significant increases
in TNF-α, IL-1β, and IL-6 expression levels in CSF of model rats
when compared with the control group were observed. However, the
addition of the miR-146a inhibitor reversed the effect of
pilocarpine treatment by preventing the increase in the expression
levels of TNF-α, IL-1β and IL-6 (Fig.
5; P<0.05). These findings suggest that the miR-146a
antagonist attenuates the expression of TNF-α, IL-1β and IL-6 in
the CSF of SE rats.
Discussion
miR-146a is a typical multifunctional miR and hasa
key role in several biological processes. miR-146a was upregulated
during epileptogenesis in an animal model and patients (4,21).
Aronica et al reported an altered expression pattern of
miR-146a in epileptic rats and in the temporal lobe of epilepsy
patients, and demonstrated high expression levels of miR-146a in
the latent and chronic stages of disease inthe rat model and human
tissues (22). The present study
investigated the dynamic expression of miR-146a in a
lithium-pilocarpine-induced epilepsy rat model and the possible use
of miR-146a as a target against epilepsy. The present findings
revealed a marked upregulation of miR-146a in the model group,
whereas rats treated with the miR-146a antagonist had reduced
miR-146 expression levels. This finding was consistent with that
reported by the Hu et al study (23).
The transcription factor, NF-κB is an important
regulator of immune and inflammatory processes, and has previously
been described to be involved in neuropathological processes,
including seizures and epilepsy (24,25).
Furthermore, a previous study demonstrated that NF-κB is activated
in response to convulsion stimulation in rodent models (26). When NF-κB is activated, it binds to
the promoter domain of the target genes through DNA NF-κB
sequences. A number of cytokines and chemokines are involved in
this pathway following NF-κB activation (27). Previous study has identified the
P-glycoprotein (P-gp) as a regulator of the NF-κB signaling pathway
at the blood-brain barrier (BBB) in a number of pathogenic
scenarios (28). It has previously
been suggested that antiepileptic drugs may use P-gp as an efflux
pump, which is located at the endothelial cell membrane in the
brain (29). The expression of
P-gp in the brain is increased under prolonged seizure conditions,
such as SE or frequent spontaneous seizures (30). In the present study, the expression
levels of NF-κB and P-gp were markedly increased in
lithium-pilocarpine-induced epilepsy; however, the addition of the
miR-146a inhibitor was able to block this effect. The findings
obtained by the current study are in line with those found in
previous studies. Experimental models and clinical studies have
confirmed that a prolonged seizure or SE may lead to neuronal death
in the brain (31). The underlying
mechanism by which neurons die following brain injury, such as a
stroke or SE, has revealed the involvement of apoptosis in neuronal
cell death (32). Apoptosis is
regulated by several signaling pathways, in which the Bcl-2 protein
family has an important role (33). Proteins of particular interest
within the Bcl-2 family are Bcl-2, an anti-apoptoticprote in which
may prevent cell death and Bax, a homolog of Bcl-2 which may
promote apoptosis (34). The
Bcl-2/Bax ratio has previously been used to determine the
anti-apoptosis capability (35).
Treatment with the miR-146a inhibitor may protect neurons from
lithium-pilocarpine-induced toxicity and reverse
lithium-pilocarpine-induced neuronal injury, partially through
inhibition of the aforementioned apoptotic pathways.
NF-κB is an important inflammatory regulator, which
is responsible for the activation of downstream pro-inflammatory
cytokines, including IL-1β, IL-6 and TNF-α, in SE models or
patients (36). Previous studies
have revealed that inflammation contributes to epileptogenesis
(18–20). Ravizza et al (18) identified that expression of IL-1β
was markedly induced in the acute stage 4 h following SE.
Furthermore, it has been revealed that seizures may allow
circulating proteins to enter the brain through the BBB (6). IL-6 is a cytokine that is upregulated
following different types of tissue trauma and inflammation. IL-6
and TNF-α levels were observed as being upregulated in the SE model
and in patients following a seizure (37,38).
The present study demonstrated an upregulation of TNF-α, IL-1β and
IL-6 in the model group when compared with the control group.
Additionally, it was determined that treatment with miR-146a may
significantly reduce the expression levels of pro-inflammatory
cytokines compared to those observed in the model group. The
findings of the present study therefore suggest that activation of
the cytokine network is associated with epilepsy.
The present study may not explicitly explain the
enacting mechanism of the miR-146a inhibitor; however, the current
findings may provide a novel the rapeutictarget for antiepileptic
therapy. Further investigation is required to establish whether the
change in expression of pro-inflammatory cytokines and the altered
activity of the NF-κB pathway occur via distinct or related
mechanisms in the SE model.
In conclusion, the use of miR-146a inhibitor for the
blocking of the pathogenic activation of the NF-κB pathway
contributing to epileptogenesis in the SE brain may have a
protective function. The current study demonstrated that there was
an upregulation of miR-146a expression in
lithium-pilocarpine-induced epilepsy. The current findings also
demonstrated an upregulation of miR-146a expression associated with
seizures. The expression patterns of pro-inflammatory cytokines,
including TNF-α, IL-1β and IL-6 and their regulator, NF-κB, suggest
an interactive relationship. The findings of the current study
support the proposal that pro-inflammatory cytokines and the NF-κB
pathway have a role in the pathogenesis of SE development.
Additionally, the findings of the present study suggest that
modulation of miR-146a expression by using an inhibitor may act asa
potential target for antiepileptic therapy.
References
|
1
|
Engel T and Henshall DC: Apoptosis, Bcl-2
family proteins and caspases: The ABCs of seizure-damage and
epileptogenesis? Int J Physiol Pathophysiol Pharmacol. 1:97–115.
2009.PubMed/NCBI
|
|
2
|
Cai Z, Li S, Li S, Song F, Zhang Z, Qi G,
Li T, Qiu J, Wan J, Sui H and Guo H: Antagonist targeting
microRNA-155 protects against lithium-pilocarpine-induced status
epilepticus in C57BL/6 mice by activating brain-derived
neurotrophic factor. Front Pharmacol. 7:1292016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang H, Zhou H and Wang N: Recent
advances in epilepsy management. Cell Biochem Biophys. 73:7–10.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Omran A, Peng J, Zhang C, Xiang QL, Xue J,
Gan N, Kong H and Yin F: Interleukin-1β and microRNA-146a in an
immature rat model and children with mesial temporal lobe epilepsy.
Epilepsia. 53:1215–1224. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stephen LJ, Kelly K, Mohanraj R and Brodie
MJ: Pharmacological outcomes in older people with newly diagnosed
epilepsy. Epilepsy Behav. 8:434–437. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li MM, Li XM, Zheng XP, Yu JT and Tan L:
MicroRNAs dysregulation in epilepsy. Brain Res. 1584:94–104. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aronica E, Fluiter K, Iyer A, Zurolo E,
Vreijling J, van Vliet EA, Baayen JC and Gorter JA: Expression
pattern of miR-146a, an inflammation-associated microRNA, in
experimental and human temporal lobe epilepsy. Eur J Neurosci.
31:1100–1107. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li X, Gibson G, Kim JS, Kroin J, Xu S, van
Wijnen AJ and Im HJ: MicroRNA-146a is linked to pain-related
pathophysiology of osteoarthritis. Gene. 480:34–41. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Miller JA, Kirkley KA, Padmanabhan R,
Liang LP, Raol YH, Patel M, Bialecki RA and Tjalkens RB: Repeated
exposure to low doses of kainic acid activates nuclear factor kappa
B (NF-κB) prior to seizure in transgenic NF-κB/EGFP reporter mice.
Neurotoxicology. 44:39–47. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liang Y and Wang X: Developing a new
perspective to study the health of survivors of Sichuan earthquakes
in China: A study on the effect of post-earthquake rescue policies
on survivors' health-related quality of life. Health Res Policy
Syst. 11:412013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liang Y and Lu P: Effect of occupational
mobility and health status on life satisfaction of Chinese
residents of different occupations: Logistic diagonal mobility
models analysis of cross-sectional data on eight Chinese provinces.
Int J Equity Health. 13:152014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lubin FD, Ren Y, Xu X and Anderson AE:
Nuclear factor-kappa B regulates seizure threshold and gene
transcription following convulsant stimulation. J Neurochem.
103:1381–1395. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vezzani A, Friedman A and Dingledine RJ:
The role of inflammation in epileptogenesis. Neuropharmacology.
69:16–24. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rowley S and Patel M: Mitochondrial
involvement and oxidative stress in temporal lobe epilepsy. Free
Radical Biol Med. 62:121–131. 2013. View Article : Google Scholar
|
|
16
|
Meffert MK and Baltimore D: Physiological
functions for brain NF-kappaB. Trends Neurosci. 28:37–43. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
da Costa AV, Calábria LK, Nascimento R,
Carvalho WJ, Goulart LR and Espindola FS: The
streptozotocin-induced rat model of diabetes mellitus evidences
significant reduction of myosin-Va expression in the brain. Metab
Brain Dis. 26:247–251. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ravizza T, Rizzi M, Perego C, Richichi C,
Velísková J, Moshé SL, De Simoni MG and Vezzani A: Inflammatory
response and glia activation in developing rat hippocampus after
status epilepticus. Epilepsia. 46 Suppl 5:113–117. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Patterson KP, Brennan GP, Curran M,
Kinney-Lang E, Dubé C, Rashid F, Ly C, Obenaus A and Baram TZ:
Rapid, coordinate inflammatory responses after experimental febrile
status epilepticus: Implications for epileptogenesis. eNeuro.
2:0034–15. 2015. View Article : Google Scholar
|
|
20
|
Itoh K, Ishihara Y, Komori R, Nochi H,
Taniguchi R, Chiba Y, Ueno M, Takata-Tsuji F, Dohgu S and Kataoka
Y: Levetiracetam treatment influences blood-brain barrier failure
associated with angiogenesis and inflammatory responses in the
acute phase of epileptogenesis in post-status epilepticus mice.
Brain Res. 1652:1–13. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Faraoni I, Antonetti FR, Cardone J and
Bonmassar E: miR-155 gene: A typical multifunctional microRNA.
Biochim Biophys Acta. 1792:497–505. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Aronica E, van Vliet EA, Hendriksen E,
Troost D, Lopes da Silva FH and Gorter JA: Cystatin Ca cysteine
protease inhibitor, is persistently up-regulated in neurons and
glia in a rat model for mesial temporal lobe epilepsy. Eur J
Neurosci. 14:1485–1491. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hu K, Zhang C, Long L, Long X, Feng L, Li
Y and Xiao B: Expression profile of microRNAs in rat hippocampus
following lithium-pilocarpine-induced status epilepticus. Neurosci
Lett. 488:252–257. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mattson MP and Camandola S: NF-kappaB in
neuronal plasticity and neurodegenerative disorders. J Clin Invest.
107:247–254. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mattson MP and Meffert MK: Roles for
NF-kappaB in nerve cell survival, plasticity, and disease. Cell
Death Differ. 13:852–860. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu Z, Zhou D, Cheng G and Mattson MP:
Neuroprotective role for the p50 subunit of NF-kappaB in an
experimental model of Huntington's disease. J Mol Neurosci.
15:31–44. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu N, Di Q, Liu H, Hu Y, Jiang Y, Yan YK,
Zhang YF and Zhang YD: Nuclear factor-kappa B activity regulates
brain expression of P-glycoprotein in the kainic acid-induced
seizure rats. Mediat Inflamm. 2011:6706132011. View Article : Google Scholar
|
|
28
|
Zhang J, Zhang M, Sun B, Li Y, Xu P, Liu
C, Liu L and Liu X: Hyperammonemia enhances the function and
expression of P-glycoprotein and Mrp2 at the blood-brain barrier
through NF-κB. J Neurochem. 131:791–802. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lazarowski A, Czornyj L, Lubienieki F,
Girardi E, Vazquez S and D'Giano C: ABC transporters during
epilepsy and mechanisms underlying multidrug resistance in
refractory epilepsy. Epilepsia. 48 Suppl 5:S140–S149. 2007.
View Article : Google Scholar
|
|
30
|
Volk H, Potschka H and Löscher W:
Immunohistochemical localization of P-glycoprotein in rat brain and
detection of its increased expression by seizures are sensitive to
fixation and staining variables. J Histochem Cytochem. 53:517–531.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Thom M, Zhou JM, Martinian L and Sisodiya
S: Quantitative post-mortem study of the hippocampus in chronic
epilepsy: Seizures do not inecvitably cause neuronal loss. Brain.
128:1344–1357. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kondratyev A and Gale K: Intracerebral
injection of caspase-3 inhibitor prevents neuronal apoptosis after
kainic acid-evoked status epilepticus. Brain Res Mol Brain Res.
75:2216–224. 2000. View Article : Google Scholar
|
|
33
|
Heath-Engel HM, Chang NC and Shore GC: The
endoplasmic reticulum in apoptosis and autophagy: Role of the BCL-2
protein family. Oncogene. 27:6419–6433. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sanchez A, Tripathy D, Yin X, Luo J,
Martinez J and Grammas P: Pigment epithelium-derived factor (PEDF)
protects cortical neurons in vitro from oxidant injury by
activation of extracellular signal-regulated kinase (ERK) 1/2 and
induction of Bcl-2. Neurosci Res. 72:1–8. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zeren T, Inan S, Vatansever HS and Sayhan
S: Significance of apoptosis related proteins on malignant
transformation of ovarian tumors: A comparison between Bcl-1/Bax
ratio and p53 immunoreactivity. Acta Histochem. 116:1251–1258.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vezzani A and Granata T: Brain
inflammation in epilepsy: Experimental and clinical evidence.
Epilepsia. 46:1724–1743. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vezzani A and Baram TZ: New roles for
interleukin-1 Beta in the mechanisms of epilepsy. Epilepsy Curr.
7:45–50. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Peltola J, Palmio J, Korhonen L, Suhonen
J, Miettinen A, Hurme M, Lindholm D and Keränen T: Interleukin-6
and interleukin-1 receptor antagonist in cerebrospinal fluid from
patients with recent tonic-clonic seizures. Epilepsy Res.
41:205–211. 2000. View Article : Google Scholar : PubMed/NCBI
|