Introduction
Airway inflammatory diseases, such as chronic
obstructive pulmonary disease (COPD) and asthma, are major causes
of morbidity and mortality in patients, and place a substantial
burden on healthcare systems (1).
Bacterial infection, cigarette smoking and air pollutants are
implicated in the onset and progression of lung inflammation
(2). Lipopolysaccharides (LPS) is
the major outer surface membrane component of Gram-negative
bacteria and a biologically active component present in cigarette
smoke (3). Thus, LPS-mediated
inflammatory response is a major lung inflammation source from
exposure to both gram-negative bacterial infection and cigarette
smoke. During this inflammatory response, macrophages first help in
endocytosis of bacterium debris, followed by generation of
inflammatory cytokines and expansion of the local inflammatory
response (4,5). In addition, the respiratory
epithelium has an active role in the airway defense through the
production of cytoprotective mucus and through coordinating local
inflammation responses by producing proinflammatory cytokines.
These cytokines, however, also result in bronchial maladaptations,
including pulmonary dysfunction, increased mucin production and
protease-antiprotease imbalance (6–8).
It has been reported that proinflammatory cytokine
production is induced by many stimuli through the mitogen-activated
protein kinase (MAPK), nuclear factor (NF)-κB or Janus kinase
(JAK)/signal transducer and activator of transcription (STAT3)
signaling cascades. For instance, LPS-induced MAPK and JAK/STAT3
activation results in activation of the downstream transcriptional
factors NF-κB, activator protein (AP)-1, peroxisome
proliferator-activated receptor (PPAR) and STAT3, which mediate the
transcription and translation of proinflammatory genes (9–11).
In the present study, in order to investigate the mechanisms of LPS
induced-inflammatory response in airway epithelial and macrophage
cells, the effects of LPS exposure on the expression of interleukin
(IL)-6, IL-8, IL-10, tumor necrosis factor (TNF)-α, matrix
metallopeptidase (MMP)-9 and tissue inhibitor of metalloproteinases
(TIMP)-1 were examined in human airway epithelial H292 cells and
macrophage THP-1 cells. Subsequently, inflammation-related
transcription factors and intracellular signaling pathways that may
be involved in LPS-induced pro-inflammation cytokine production
were explored.
Materials and methods
Cell culture
H292 human lung mucoepidermoid carcinoma cells and
THP-1 human monocytic cells were obtained from the American Type
Culture Collection (Manassas, VA, USA). The cells were maintained
in RPMI-1640 (Beijing Solarbio Science and Technology Co., Ltd.,
Beijing, China) supplemented with 10% fetal calf serum (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA), penicillin (100
U/ml) and streptomycin (100 mg/ml), and incubated at 37°C with 5%
CO2. All experiments were performed with exponentially
growing cells. For differentiation to a macrophage phenotype, THP-1
cells were adjusted to the desired concentrations for each
experiment and incubated with 20 µM phorbol myristate acetate (PMA)
diluted in complete culture medium for 24 h. Then, the cells were
washed with serum-free RPMI-1640 medium prior to each
experiment.
Cell viability assay
For the cell viability assay, H292 (2×104
cells/well) and THP-1 cells (1×104 cells/well,
pretreated with 20 µM PMA) were plated in sextuplicate 96-well
plates for 24 h. The cells were then stimulated with the indicated
concentrations of LPS for 6, 12, 24 and 48 h. Normal complete
RPMI-1640 media without LPS was used as a negative control.
Subsequently, MTT solution was added for 4 h, following which the
supernatants were removed and 150 µl dimethyl sulfoxide was added
to each well to dissolve the formazan crystals. The absorbance was
measured at 570 nm.
Cytokine analysis
H292 (2×104 cells/well) and THP-1 cells
(1×104 cells/well, pretreated with 20 µM PMA) were
seeded in triplicate into 96-well tissue culture plates for 24 h.
The cells were then stimulated with 1, 2 or 2.5 µg/ml of LPS for 6,
12, 24 and 48 h. The levels of human cytokines were measured in the
collected supernatants with human IL-6 (cat. no. EK0410), IL-8
(cat. no. EK0413), IL-10 (cat. no. EK0416), TNF-α (cat. no.
EK0525), MMP-9 (cat. no. EK0465) and TIMP-1 (cat. no. EK0520) ELISA
kits (Boster Biological Technology, Ltd., Wuhan, China), according
to the manufacturer's protocols. For the inhibitor experiments,
H292 and THP-1 cells were pretreated with pyrrolidine
dithiocarbamate (PDTC), SP00125 or stattic (all Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) for 4 h, followed by exposure to
LPS for 48 h, prior to measuring IL-8 and TNF-α levels in the cell
culture supernatants with ELISA kits, as aforementioned.
Electrophoretic mobility shift assay
(EMSA)
Nuclear protein was extracted from LPS-stimulated
H292 and THP-1 cells using a total nuclear protein extraction kit
(Beyotime Institute of Biotechnology, Shanghai, China), according
to the manufacturer's protocol. Protein concentration was measured
with the bicinchoninic acid (BCA) method.
H292 and differentiated THP-1 cells were treated
with LPS for 24 h. The DNA binding activities of AP-1, PPAR, NF-κB
and STAT3 in the nuclear extracts were assessed by EMSA using the
EMSA kit (Beyotime Institute of Biotechnology, Haimen, China),
according to the manufacturer's protocol. The sequences of the
oligonucleotides used were as follows: AP1, forward
5′-CGCTTGATGACTCAGCCGGAA-3′ and reverse
3′-GCGAACTACTGAGTCGGCCTT-5′; AP-1 mutated probe, forward
5′-CGCTTGATGACTTGGCCGGAA-3′ and reverse
3′-GCGAACTACTGAACCGGCCTT-5′; PPAR, forward
5′-CAAAACTAGGTCAAAGGTCA-3′ and reverse 3′-GTTTTGATCCAGTTTCCAGT-5′;
PPAR mutated probe, forward 5′-CAAAACTAGCACAAAGCACA-3′,
3′-GTTTTGATCGTGTTTCGTGT-5′ and reverse STAT3, forward
5′-GATCCTTCTGGGAATTCCTAGATC-3′ and reverse
3′-CTAGGAAGACCCTTAAGGATCTAG-5′; STAT3 mutated probe, forward
5′-GATCCTTCTGGGCCGTCCTAGATC-3′ and reverse
3′-CTAGGAAGACCCGGCAGGATCTAG-5′; NF-κB, forward
5′-AGTTGAGGGGACTTTCCCAGGC-3′ and reverse
3′-TCAACTCCCCTGAAAGGGTCCG-5′; NF-κB mutated probe, forward
5′-AGTTGAGGCGACTTTCCCAGGC-3′ and reverse
3′-TCAACTCCGCTGAAAGGGTCCG-5′. Briefly, 10 µg nuclear extract was
added to 20 fmol biotin-labeled oligonucleotides, 1X binding
buffer, 2.5% Glycerol, 5 mM MgCl2, 0.1 mM EDTA and 0.05%
NP-40 at 24°C for 40 min. The reactions were analyzed by
electrophoresis on 5% polyacrylamide gels in 0.5X TBE buffer at 180
V for 35 min, and were transferred to positively-charged nylon
membranes and cross-linked in a Stratagene cross-linker (Agilent
Technologies, Inc., Santa Clara, CA, USA). The signals were
visualized using chemiluminescent detection (Beyotime Institute of
Biotechnology) on X-ray film.
Western blotting assay
H292 cells and differentiated THP-1 cells were
stimulated with 1 µg/ml LPS for 24 h. Cells were washed with
ice-cold PBS and then lysed with radio-immunoprecipitation assay
lysis buffer (Beijing Solarbio Science & Technology Co., Ltd.).
Protein concentration was determined using a BCA protein assay.
Samples containing 30 µg of protein were mixed with SDS sample
buffer and boiled. Protein was separated by 10% SDS-polyacrylamide
gel electrophoresis prior to being transferred electrophoretically
into polyvinylidene fluoride membrane. The membranes were blocked
for 1.5 h at room temperature with 5% nonfat milk and then
incubated with primary antibodies at 4°C overnight followed by
incubation with horseradish peroxidase (HRP)-conjugated secondary
antibodies for 1 h at 22°C. The bands were visualized by film
exposure following incubation with enhanced chemiluminescence
reagent (Beijing Solarbio Science & Technology Co., Ltd.).
Antibodies used were as follows: Anti-TLR4 (Proteintech Group,
Inc., Chicago, IL, USA; cat. no. 19811-1-AP; 1:500), anti-inhibitor
of κBα (IκBα; Proteintech Group, Inc.; cat. no. 10268-1-AP; 1:500),
anti-p65 (Proteintech Group, Inc.; cat. no. 10745-1-AP; 1:500),
anti-p38 (Proteintech Group, Inc.; cat. no. 14064-1-AP; 1:500),
anti-Jun proto-oncogene AP-1 transcription factor subunit (JUN;
Proteintech Group, Inc.; cat. no. 10024-2-AP; 1:500), anti-JAK1
(Proteintech Group, Inc.; cat. no. 66466-1-Ig; 1:500), anti-JAK2
(Proteintech Group, Inc.; cat. no. 17670-1-AP; 1:500), anti-β-actin
rabbit polyclonal antibody (Proteintech Group, Inc.; cat. no.
20536-1-AP; 1:4,000), anti-p-STAT3 (Bioworld Technology, Inc., St
Louis Park, MN, USA; cat. no. BS4181; 1:800), anti-p-p65 (Bioworld
Technology, Inc.; cat. no. BS5088, 1:600), anti-p-p38 (Bioworld
Technology, Inc.; cat. no. 4522; 1:500), HRP-conjugated Affinipure
goat anti-mouse IgG(H+L) (Proteintech Group, Inc.; cat. no.
SA00001-1; 1:5,000) and HRP-conjugated Affinipure Goat Anti-Rabbit
IgG(H+L) (Proteintech Group, Inc.; cat. no. SA00001-2;
1:5,000).
Statistical analysis
Data were presented as mean ± standard error.
Results were analyzed by one-way analysis of variance followed by
Tukey's range test using the SPSS 17.0 software package (SPSS,
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results and Discussion
Effect of LPS exposure on cell
viability
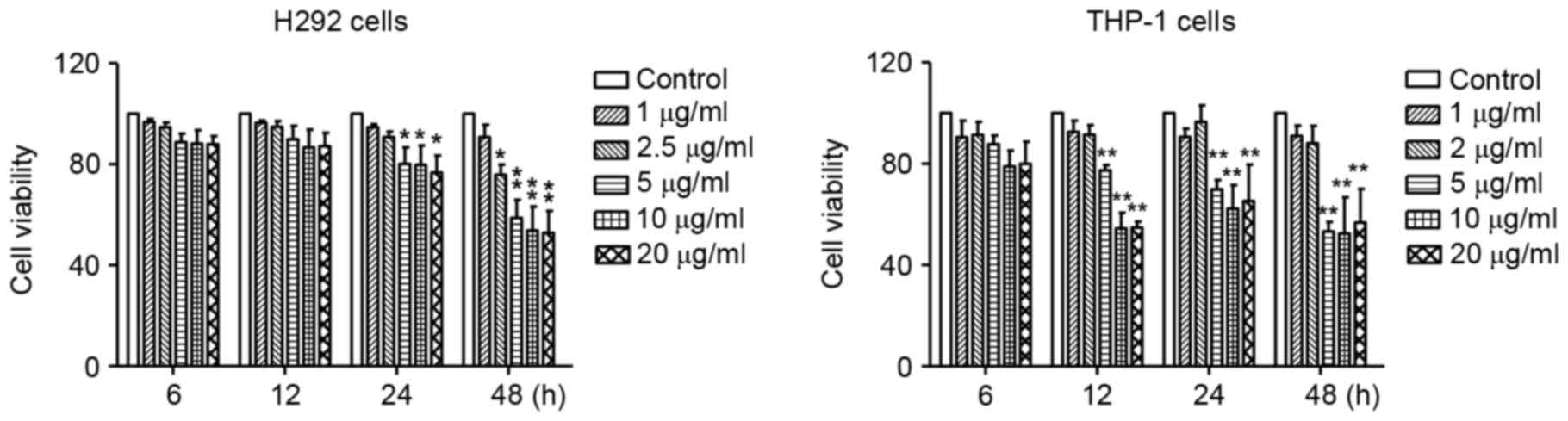
First, the cytotoxic effects of LPS were determined
by MTT assay in human lung mucoepidermoid carcinoma H292 cells and
monocyte THP-1 cells treated with various concentrations of LPS for
6, 12, 24 and 48 h. As illustrated in Fig. 1, compared with the untreated
controls, no significant change was observed in the viability of
H292 cells treated with 1 and 2.5 µg/ml LPS, and the viability of
THP-1 cells treated with 1 and 2 µg/ml LPS, indicating that LPS was
not cytotoxic at these concentrations. Higher concentration (5–20
µg/ml) of LPS were significantly cytotoxic to both H292 and THP-1
cells (Fig. 1). Therefore, the
concentrations of 1–2.5 and 1–2 µg/ml were selected for H292 and
THP-1 cells, respectively, in all subsequent experiments.
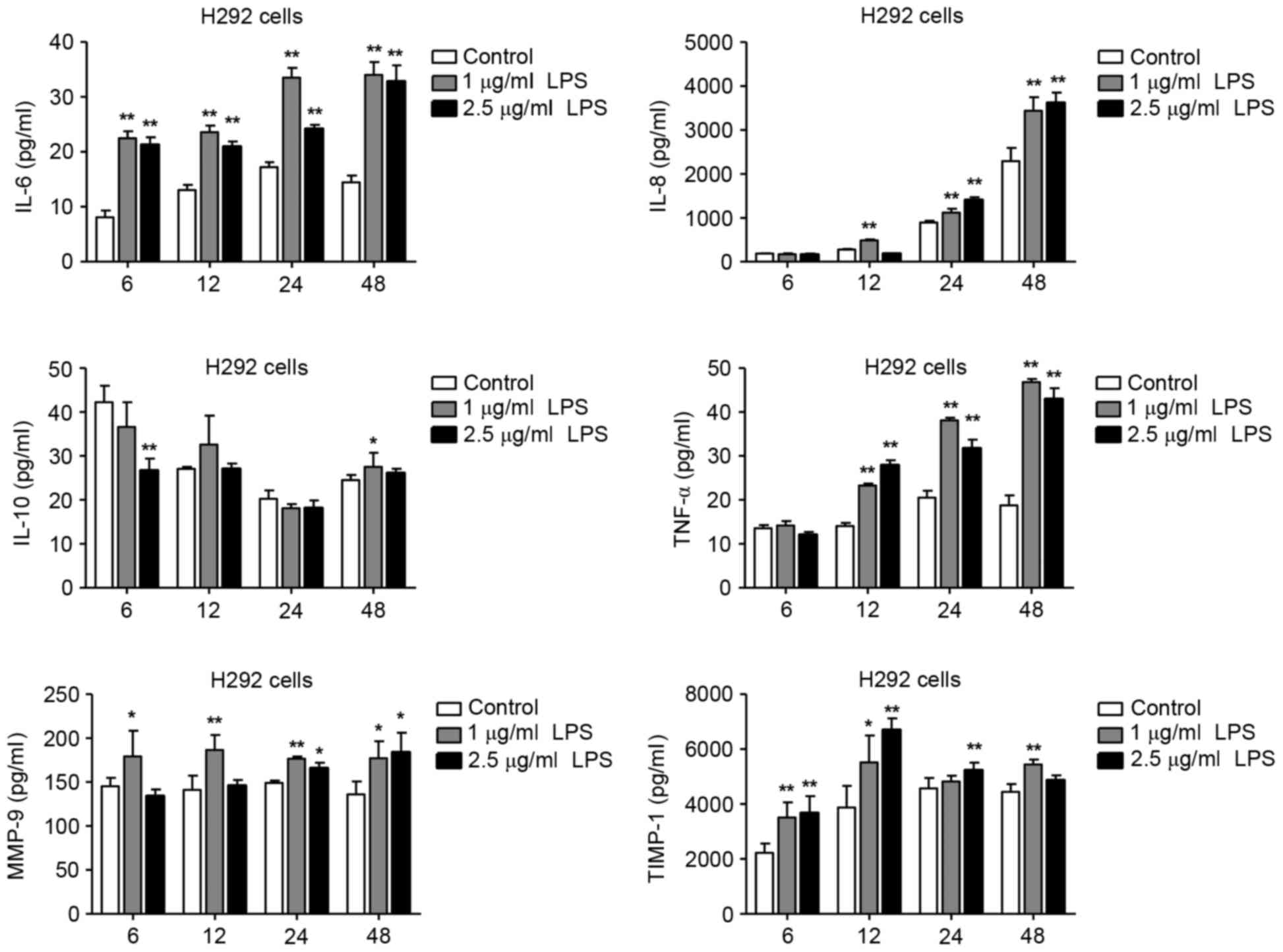
Levels of IL-6, −8, −10, TNF-α, MMP-9
and TIMP-1 in LPS-exposed H292 and THP-1 cells
LPS is a potent proinflammatory activator of
monocytes and macrophages in vivo and in vitro
(3,12). In the present study, H292 and
differentiated THP-1 cells were exposed to different concentrations
of LPS for 6, 12, 24 and 48 h, then the expression levels of IL-6,
−8, −10, TNF-α, MMP-9 and TIMP-1 were detected by ELISA. Compared
with untreated H292 cells, treatment with 1 and 2.5 µg/ml LPS
resulted in increased levels of IL-6, −8, TNF-α, MMP-9 and TIMP-1
(Fig. 2). IL-10 levels in the
LPS-treated H292 cells, however, were not significantly altered
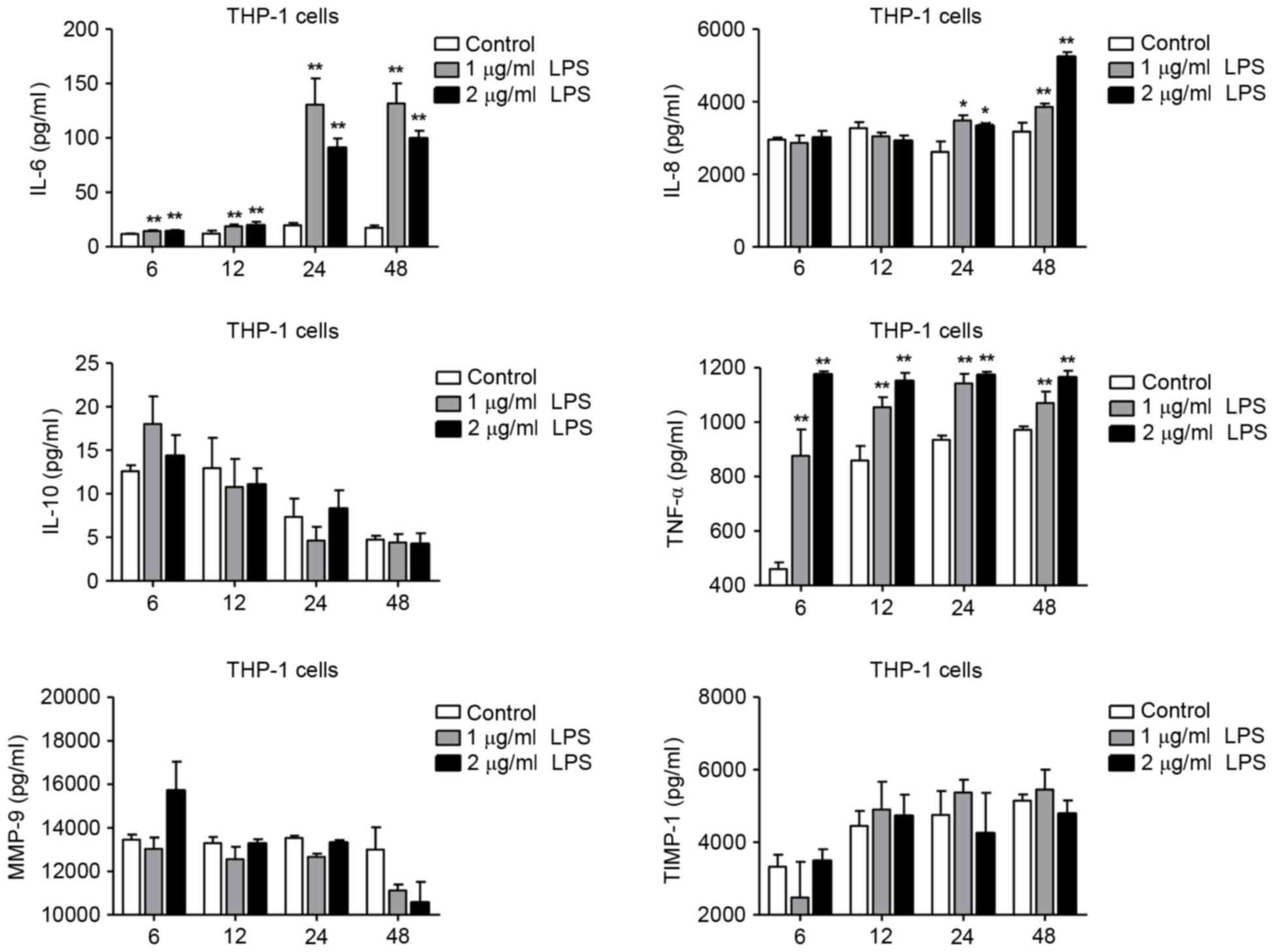
(Fig. 2). In differentiated THP-1
cells, stimulation with 1 and 2 µg/ml LPS resulted in an increase
of IL-6, −8 and TNF-α levels compared with control (Fig. 3). However, the expression levels of
IL-10, MMP-9 and TIMP-1 were not significantly altered in THP-1
cells following LPS stimulation (Fig.
3). These results demonstrated that LPS could induce IL-6, −8,
and TNF-α expression in both the H292 and THP-1 cells. In addition,
LPS treatment significantly increased the expression levels of
MMP-9 and TIMP-1 in H292 cells, but not in differentiated THP-1
cells. Taken together, these data suggest that LPS stimulation had
the different effects on cytokine expression profiles in H292 and
differentiated THP-1 cells, and this may be related to differences
in the physiological characteristics of these two cell types. H292
cells have characteristics similar to those of alveolar and
bronchiolar epithelial cells and are widely used in alveolar and
bronchiolar epithelial models, and the gene expression profile of
H292 cells is similar to that of primary nasal epithelial cells
from healthy human controls (13).
THP-1 cells, a human leukemia-derived monocytic cell line, is the
most commonly used cellular model for investigating human
macrophage function (12).
Effect of LPS treatment on NF-κB,
AP-1, PPAR and STAT3 DNA-binding activity
The transcription factors NF-κB, AP-1, PPAR and
STAT3 are the main signal transduction molecules activated in
response to LPS-induced inflammatory response (12,14).
For example, NF-κB translocates to the nucleus and promotes the
transcription of proinflammatory genes, including the biologically
active cytokine TNF-α and proinflammatory interleukins in activated
macrophages (14). Thus, the
activation of NF-kB, AP-1, PPAR and STAT3 was investigated in H292
and THP-1 cells stimulated with LPS by EMSA. As illustrated in
Fig. 4, treatment of H292 cells
with 1or 2.5 µg/ml LPS for 24 h markedly increased the NF-κB and
AP-1 binding activities, but had no effect on the PPAR and STAT3
binding activities. Treatment of differentiated THP-1 cells with 1
and 2 µg/ml LPS for 24 h markedly increased the NF-κB and STAT3
binding activities, slightly increased the AP-1 binding activity,
but had no effect on PPAR (Fig.
5). These data indicated that LPS-induced cytokine expression
in H292 cells likely resulted from the activation of the NF-κB and
AP-1 transcription factors. Similarly, LPS-mediated cytokine
expression in THP-1 cells was associated with the activation of the
NF-κB, AP-1 and STAT3 transcription factors.
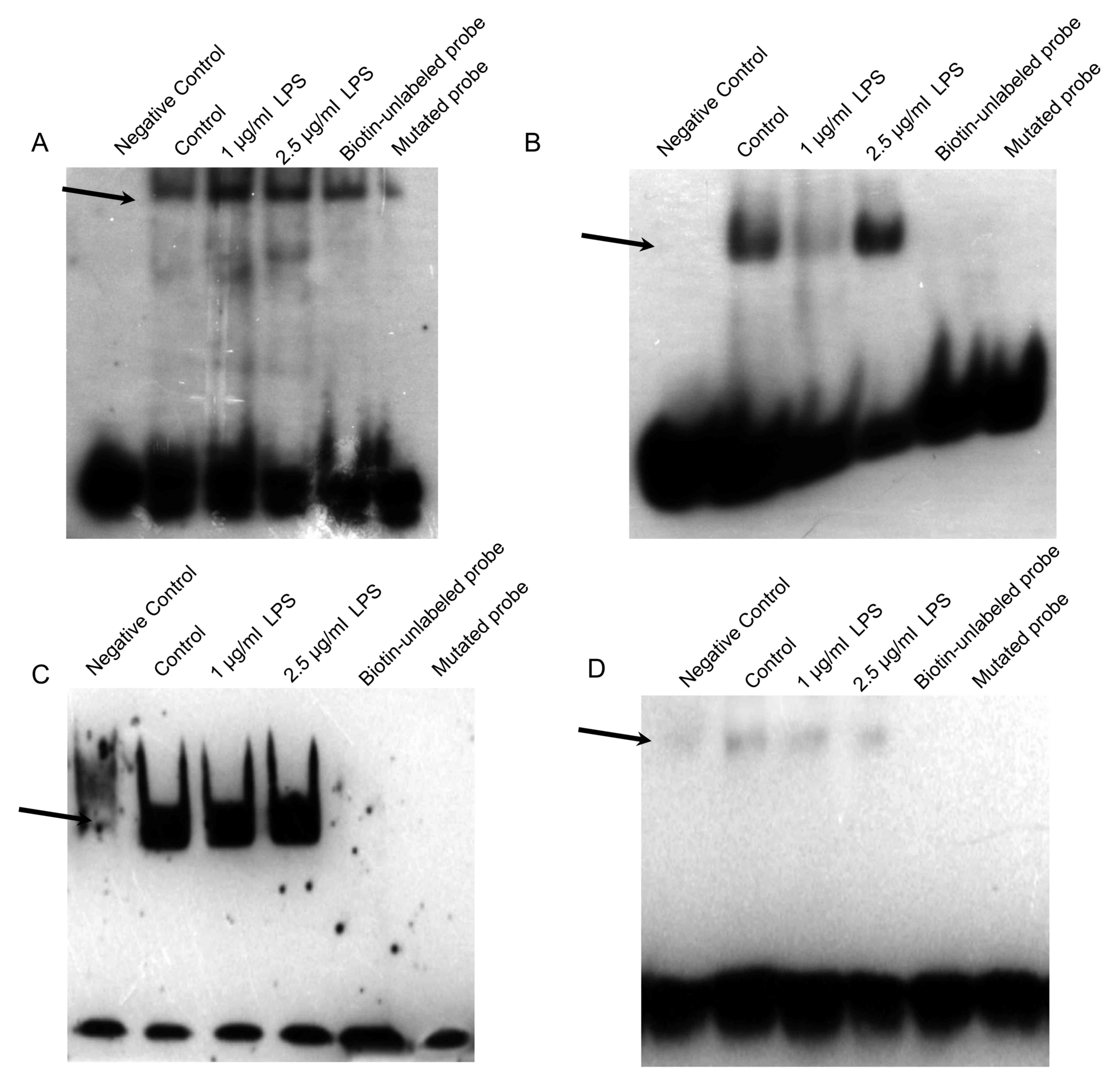 | Figure 4.Effects of LPS stimulation on NF-κB,
AP-1, PPAR and STAT3 DNA-binding activity in H292 cells. Nuclear
extracts were prepared from H292 cells and EMSA was performed by
incubation of extracts with the (A) NF-κB, (B) AP-1, (C) PPAR or
(D) STAT3 DNA-binding consensus sequences. Negative control, the
biotin-labelled DNA oligo probe run without any protein extract;
biotin-unlabeled protein, the biotin-unlabelled DNA oligo probe run
with protein extract; mutated probe, the biotin-labelled DNA oligo
probe with binding site mutation, run with protein extract. A total
of three separate experiments were performed. Representative gel
images are shown. LPS, lipopolysaccharide; NF-κB, nuclear
factor-κB; AP, activator protein; PPAR, peroxisome
proliferator-activated receptor; STAT, signal transducer and
activator of transcription. |
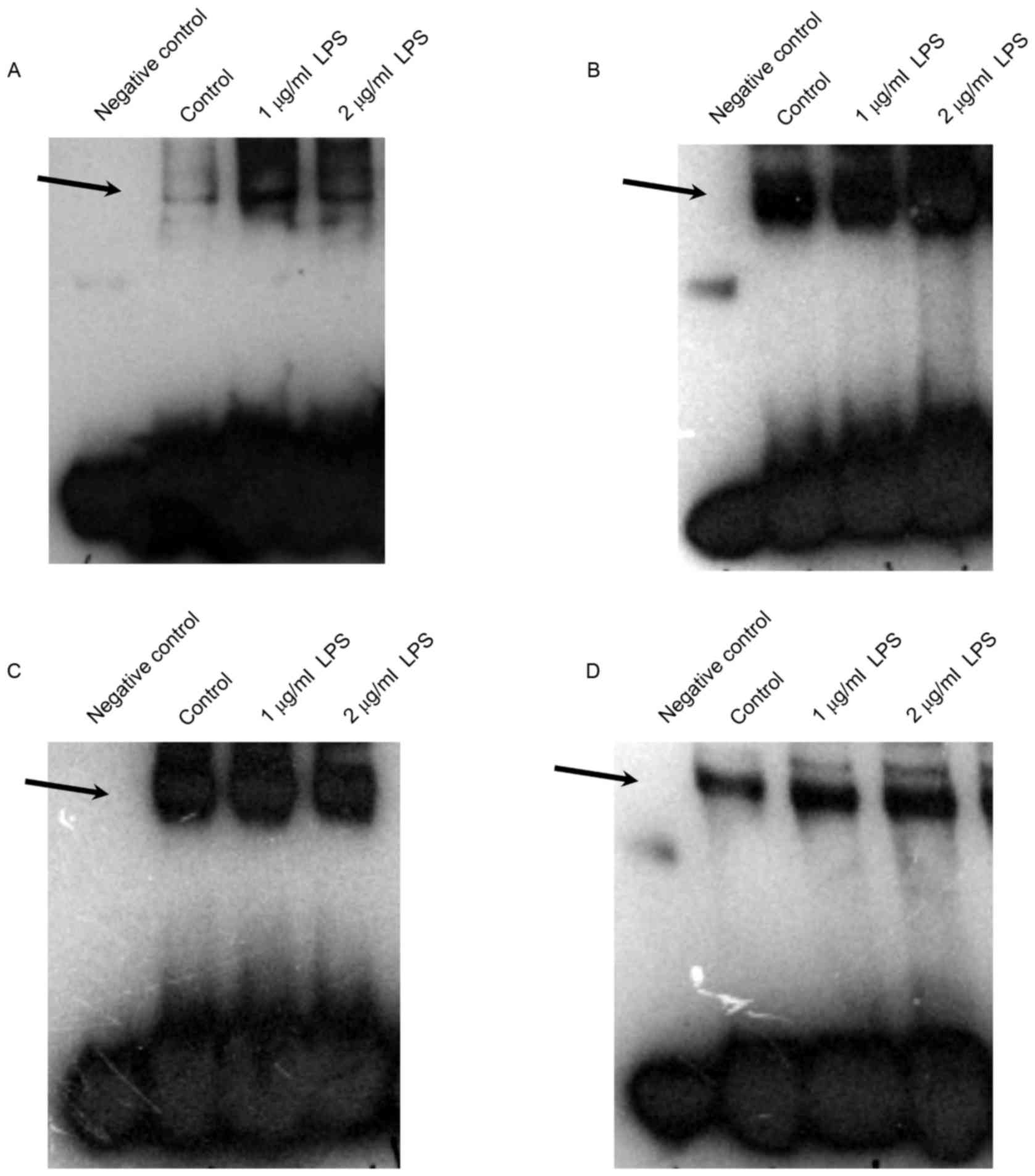 | Figure 5.Effects of LPS stimulation on NF-κB,
AP-1, PPAR and STAT3 DNA-binding activity in THP-1 cells. Nuclear
extracts were prepared from the THP-1 cells and EMSA was performed
by incubation of extracts with the (A) NF-κB, (B) AP-1, (C) PPAR
and (D) STAT3 DNA-binding consensus sequences. Negative control,
the biotin-labelled DNA oligo probe run without any protein
extract. A total of three separate experiments were performed.
Representative gel images are shown. LPS, lipopolysaccharide;
NF-κB, nuclear factor-κB; AP, activator protein; PPAR, peroxisome
proliferator-activated receptor; STAT, signal transducer and
activator of transcription. |
Effect of LPS exposure on the NF-κB,
MAPK and STAT3 signaling pathways
The activation of NF-κB signaling increases the IκB
phosphorylation or degradation, and subsequent NF-κB
transactivation, translocation and promoter binding (15). In addition, NF-κB activation in the
airways of allergen-challenged mice is attenuated by Toll-like
receptor (TLR) 2 or TLR4 gene deletion, suggesting that TLR2 or
TLR4 contribute to NF-κB signaling (16,17).
Similarly, the AP-1 and STAT3 promoter-binding activity are
regulated by MAPK and JAK/STAT signaling (18,19).
Therefore, it was hypothesized that LPS may induce NF-κB, MAPK and
JAK/STAT pathway activation, leading to NF-κB, AP-1 and STAT3
nuclear translocation and cytokine expression in the H292 and THP-1
cells investigated in the present study.
In order to further examine the effects of LPS on
pathway activation in H292 and THP-1 cells, cells were treated with
1 µg/ml LPS and the activation of NF-κB and MAPK signaling pathways
in H292 cells, and the activation of NF-κB and JAK/STAT3 signaling
pathways in THP-1 cells, were tested by western blotting. As the
results demonstrated, LPS treatment for 24 h resulted in an obvious
increase in p65 and p38 phosphorylation, as well as a marked
increase in TLR4, IκBα, p65, p38 and JUN protein expression levels
in H292 cells (Fig. 6A). In the
differentiated THP-1 cells, LPS treatment for 24 h resulted in a
marked increase of STAT3 and p65 phosphorylation, as well as an
increase in TLR4, IκBα and Janus kinase (JAK) 1 protein expression
levels, while JAK2 expression was not altered (Fig. 6B). These results indicated that
NF-κB, MAPK and their downstream signals were activated in H292
cells, while NF-κB and STAT3 downstream signals were activated in
THP-1 cells following LPS exposure.
 | Figure 6.Effect of LPS stimulation on the
NF-κB, MAPK and STAT3 signaling pathways. (A) H292 and (B) THP-1
cells were treated with 1 µg/ml LPS for 24 h and protein expression
levels of the indicated signaling molecules were examined by
western blotting. Representative images are shown from three
independent repeats. LPS, lipopolysaccharide; NF-κB, nuclear
factor-κB; MAPK, mitogen-activated protein kinase; STAT, signal
transducer and activator of transcription; TLR, Toll-like receptor;
IκB, inhibitor of κB; P-, phosphorylated; p65, RELA proto-oncogene
NF-κB subunit; p38, mitogen-activated protein kinase 14; JUN, Jun
proto-oncogene AP-1 transcription factor subunit; JAK, Janus
kinase. |
LPS has been reported to bind to the TLR4 receptor,
and to induce activation of the NF-κB pathway which then results in
the release of proinflammatory cytokines (20). In the present study, LPS treatment
was demonstrated to increase TLR4 expression in both H292 and THP-1
cells, and to activate NF-κB and MAPK signaling in H292 cells and
NF-κB and STAT3 signaling in THP-1 cells. These findings suggested
that LPS may bind to TLR4 receptor and activate different pathways
in the human airway H292 and the human macrophage THP-1 cells.
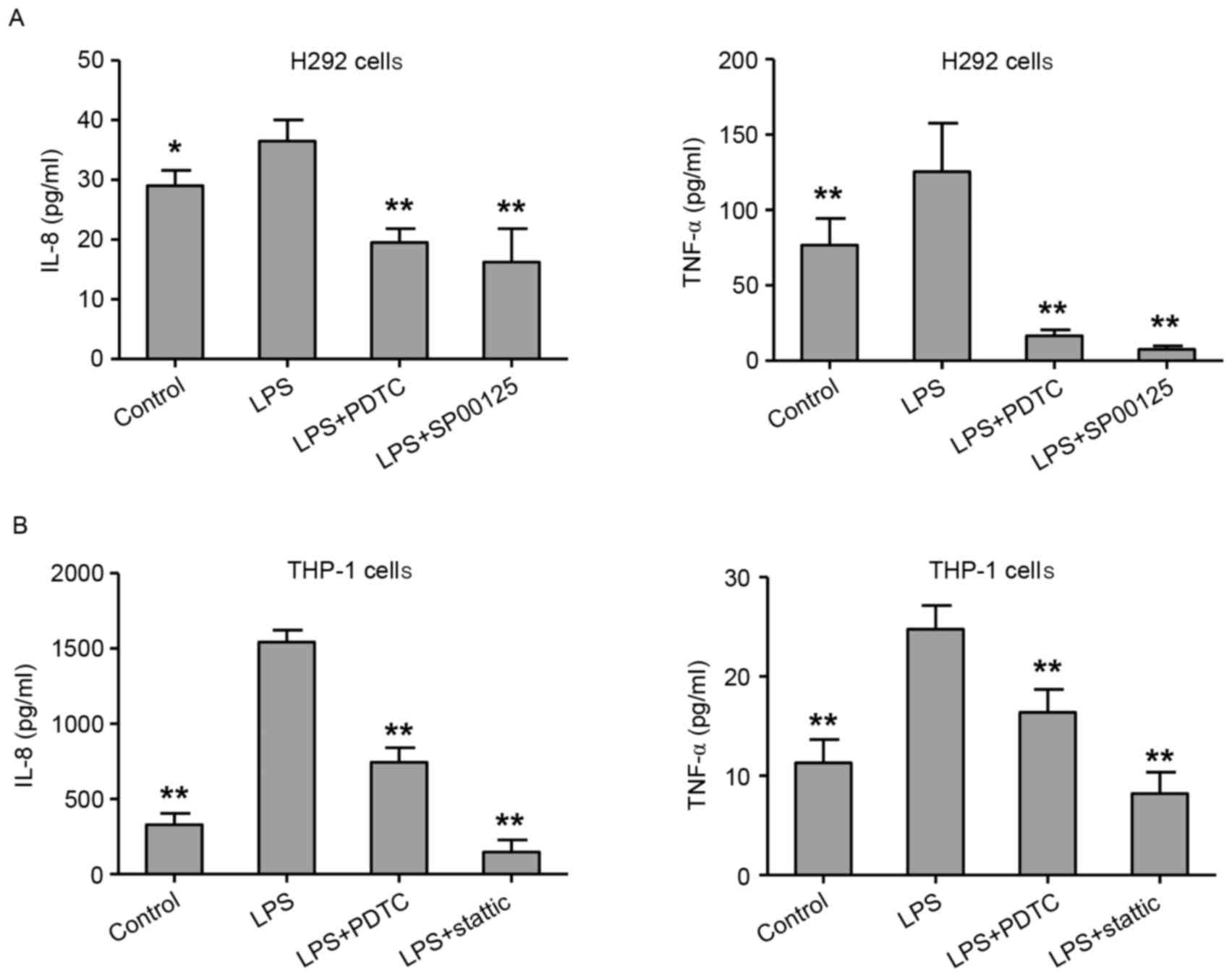
Treatment with NF-κB, STAT3 and AP-1
inhibitors reverses the LPS-induced cytokine expression
The role of the NF-κB, AP-1 or STAT3 pathway
activation in the LPS-mediated cytokine expression in H292 and
differentiated THP-1 cells was further explored. H292 and THP-1
cells were pretreated with PDTC (a NF-κB inhibitor; 100 µM),
SP00125 (an AP-1 inhibitor; 100 µM) or stattic (a STAT3 inhibitor;
100 µM) for 4 h, followed by exposure to LPS for 48 h. The results
demonstrated that treatment with PDTC or SP00125 significantly
inhibited the LPS-induced expression of IL-8 and TNF-α in H292
cells, compared with cells treated with LPS alone (Fig. 7A). Similarly, treatment with PDTC
or stattic significantly decreased the LPS-induced expression of
IL-8 and TNF-α in THP-1 cells, compared with cells treated with LPS
alone (Fig. 7B). These results
suggest that NF-κB, AP-1 or STAT3 signaling might serve critical
roles in LPS-mediated cytokine expression in lung epithelial cells
and macrophages.
In conclusion, LPS stimulation increased the
production of IL-6, IL-8, TNF-α, MMP-9 and TIMP-1 in H292 cells,
and it increased the expression of IL-6, IL-8, and TNF-α in THP-1
cells. In addition, LPS stimulation increased the DNA-binding
activation of NF-κB and AP-1 in H292 cells, and the DNA-binding
activation of NF-κB and STAT3 in THP-1 cells. NF-κB, MAPK and STAT3
downstream signaling was also activated by LPS stimulation.
Furthermore, NF-κB, AP-1 or STAT3 inhibitors significantly reversed
the LPS-mediated IL-8 and TNF-α induction in H292 and THP-1 cells.
The present study suggests that these signals might serve vital
roles in LPS-induced cytokine expression in human airway cells and
macrophages.
Acknowledgements
The authors of the present study would like to
acknowledge financial support from the National Natural Science
Fund of China (Influence and Long-Term Effects of Three Tiao-Bu
Fei-Shen Therapies in Rats with Chronic Obstructive Pulmonary
Disease on Regulation of Multidimensional Molecular Network; grant
no. 81130062), Outstanding TCM Academic Leader Program of Henan
Province (grant no. HNZYLJ201301001), and Scientific and
Technological Leading Talent Projects of Zhengzhou (grant no.
131PJRC659).
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Eisner MD, Anthonisen N, Coultas D,
Kuenzli N, Perez-Padilla R, Postma D, Romieu I, Silverman EK and
Balmes JR; Committee on Nonsmoking COPD, Environmental and
Occupational Health Assembly, : An official American Thoracic
Society public policy statement: Novel risk factors and the global
burden of chronic obstructive pulmonary disease. Am J Respir Crit
Care Med. 182:693–718. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dohrman A, Miyata S, Gallup M, Li JD,
Chapelin C, Coste A, Escudier E, Nadel J and Basbaum C: Mucin gene
(MUC 2 and MUC 5AC) upregulation by Gram-positive and Gram-negative
bacteria. Biochim Biophys Acta. 1406:251–259. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hasday JD, Bascom R, Costa JJ, Fitzgerald
T and Dubin W: Bacterial endotoxin is an active component of
cigarette smoke. Chest. 115:829–835. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li L, Wang Y, Gao W, Yuan C, Zhang S, Zhou
H, Huang M and Yao X: Klotho reduction in alveolar macrophages
contributes to cigarette smoke Extract-induced inflammation in
chronic obstructive pulmonary disease. J Biol Chem.
290:27890–27900. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tseng CT, Perrone LA, Zhu H, Makino S and
Peters CJ: Severe acute respiratory syndrome and the innate immune
responses: Modulation of effector cell function without productive
infection. J Immunol. 174:7977–7985. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ursini CL, Cavallo D, Fresegna AM, Ciervo
A, Maiello R, Tassone P, Buresti G, Casciardi S and Iavicoli S:
Evaluation of cytotoxic, genotoxic and inflammatory response in
human alveolar and bronchial epithelial cells exposed to titanium
dioxide nanoparticles. J Appl Toxicol. 34:1209–1219. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Speth JM, Bourdonnay E, Penke LR, Mancuso
P, Moore BB, Weinberg JB and Peters-Golden M: Alveolar epithelial
cell-derived prostaglandin E2 serves as a request signal for
macrophage secretion of suppressor of cytokine signaling 3 during
Innate Inflammation. J Immunol. 196:5112–5120. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li YJ, Yu CH, Li JB and Wu XY:
Andrographolide antagonizes cigarette smoke extract-induced
inflammatory response and oxidative stress in human alveolar
epithelial A549 cells through induction of microRNA-218. Exp Lung
Res. 39:463–471. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zeng KW, Wang S, Dong X, Jiang Y, Jin HW
and Tu PF: Sesquiterpene dimmer (DSF-27) inhibits the release of
neuroinflammatory mediators from microglia by targeting spleen
tyrosine kinase (Syk) and Janus kinase 2 (Jak2): Two major
non-receptor tyrosine signaling proteins involved in inflammatory
events. Toxicol Appl Pharmacol. 275:244–256. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Singh D, Siew L, Christensen J, Plumb J,
Clarke GW, Greenaway S, Perros-Huguet C, Clarke N, Kilty I and Tan
L: Oral and inhaled p38 MAPK inhibitors: Effects on inhaled LPS
challenge in healthy subjects. Eur J Clin Pharmacol. 71:1175–1184.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu F, Xu Y, Zhu L, Rao P, Wen J, Sang Y,
Shang F and Liu Y: Fasudil inhibits LPS-induced migration of
retinal microglial cells via regulating p38-MAPK signaling pathway.
Mol Vis. 22:836–846. 2016.PubMed/NCBI
|
|
12
|
Lund ME, To J, O'Brien BA and Donnelly S:
The choice of phorbol 12-myristate 13-acetate differentiation
protocol influences the response of THP-1 macrophages to a
pro-inflammatory stimulus. J Immunol Methods. 430:64–70. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vroling AB, Jonker MJ, Breit TM, Fokkens
WJ and van Drunen CM: Comparison of expression profiles induced by
dust mite in airway epithelia reveals a common pathway. Allergy.
63:461–467. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamamoto Y and Gaynor RB: Role of the
NF-kappaB pathway in the pathogenesis of human disease states. Curr
Mol Med. 1:287–296. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hart LA, Krishnan VL, Adcock IM, Barnes PJ
and Chung KF: Activation and localization of transcription factor,
nuclear factor-kappaB, in asthma. Am J Respir Crit Care Med.
158:1585–1592. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lam D, Ng N, Lee S, Batzer G and Horner
AA: Airway house dust extract exposures modify allergen-induced
airway hypersensitivity responses by TLR4-dependent and independent
pathways. J Immunol. 181:2925–2932. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li X, Chen Q, Chu C, You H, Jin M, Zhao X,
Zhu X, Zhou W and Ji W: Ovalbumin-induced experimental allergic
asthma is Toll-like receptor 2 dependent. Allergy Asthma Proc.
35:pp. e15–e20. 2014; View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yew-Booth L, Birrell MA, Lau MS, Baker K,
Jones V, Kilty I and Belvisi MG: JAK-STAT pathway activation in
COPD. Eur Respir J. 46:843–845. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Luff SA and Papoutsakis ET: Megakaryocytic
maturation in response to shear flow is mediated by the activator
protein 1 (AP-1) transcription factor via Mitogen-activated protein
kinase (MAPK) mechanotransduction. J Biol Chem. 291:7831–7843.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kagan JC and Medzhitov R:
Phosphoinositide-mediated adaptor recruitment controls Toll-like
receptor signaling. Cell. 125:943–955. 2006. View Article : Google Scholar : PubMed/NCBI
|