Introduction
Cardiovascular risk factors promote the development
of atherosclerosis (AS) by inducing endothelial cell injury and
dysfunction; therefore, improving endothelial function may limit AS
(1). An increasing amount of
evidence indicates that endothelial progenitor cells (EPCs)
function in endogenous endothelial regeneration and repair of
damaged endothelium (2–4). In addition, inflammation is
associated with atherosclerotic disease (5) and inflammatory mechanisms may
contribute to EPC impairment in AS (6). Therefore, drugs protecting EPCs from
inflammatory damage may have potential in the treatment of AS.
Trans-3,4′,5-trihydroxystilbene, also termed
resveratrol (RES), is a natural polyphenol compound that is present
in grapes, red wine and other food products (7). Previous studies have reported that
RES induces a protective effect on the cardiovascular system
(8–10). It has been demonstrated that
elevated levels of tumor necrosis factor (TNF)-α in the blood are
associated with AS (11) and that
RES restores endothelial function in diabetes by suppressing
TNF-α-induced upregulation of NADPH oxidase and promoting the
phosphorylation of endothelial nitric oxide (NO) synthase (eNOS)
(12). However, the direct effects
and the underlying mechanisms of RES in AS are yet to be
elucidated.
Krüppel-like factors (KLFs) belong to a family of
transcription factors that are responsible for various biological
processes, including proliferation, development and apoptosis
(13). Among the members of the
KLF family, KLF2 functions in several signaling pathways associated
with cell migration, vasomotor function and hemostasis (14–16).
Previous studies have demonstrated that KLF2 regulates endothelial
thrombotic function and the expression of flow-responsive genes
induced by shear stress (14,17).
However, the expression of KLF2 is inhibited by TNF-α (18) and it remains to be determined
whether RES is able to inhibit TNF-α-induced inflammatory damage in
late EPCs via regulation of KLF2 functional pathways. The present
study aimed to determine the effects of RES on the functions of
late EPCs and the underlying mechanism of TNF-α-induced
inflammatory damage in late EPCs.
Materials and methods
Identification and culture of
EPCs
Late EPCs were cultured in vitro, as
previously described (19).
Initially, bone marrow was separated from one male and one female
Sprague-Dawley rats (150 to 175 g, 8 weeks; Weifang Medical
College, Weifang, China), which were housed under controlled
conditions (12-h light/dark cycle, 22°C, 60% humidity) with ad
libitum access to food and water and mononuclear cells were
separated by density gradient centrifugation under 4°C (700 × g for
20 min) with Histopaque®-1083 (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany). Subsequently, mononuclear cells were
inoculated on dishes precoated with 5% fibronectin (Sigma-Aldrich;
Merck KGaA,) and cultured in complete Endothelial Cell Growth
Medium-2 (CC-3202; Lonza Group, Ltd., Basel, Switzerland) at 37°C
and 5% CO2 in a humidified atmosphere for 4 days.
Subsequently, non-adherent cells were washed away with PBS. The
culture was maintained for an additional 7 days, during which the
medium was replaced every other day. EPC identification was
performed as described in our previous study (19) and EPCs at passages 3–5, considered
late EPCs, and 2×106 EPCs/well were seeded in 6-well
plates and used for subsequent experiments (20).
Late EPC cells were divided into three groups with
different treatments: TNF-α group, EPCs were cultured with TNF-α
(10 ng/ml; PeproTech, Inc., Rocky Hill, NJ, USA) at 37°C for 24 h;
RES + TNF-α group, EPCs were cultured with RES (20 µmol/l;
Sigma-Aldrich; Merck KGaA) at 37°C for 12 h prior to treatment with
TNF-α (10 ng/ml) at 37°C for 24 h; and dimethyl sulfoxide (DMSO)
group, EPCs were treated with the same volume of DMSO at 37°C for
24 h. The present study was approved by the Medical Ethics
Committee of Weifang Medical University (Weifang, China) and was
performed in accordance with the committee guidelines.
Cell proliferation assays
Late EPCs at the density of 10,000 cells/well were
seeded in triplicate in 96-well plates. Cell proliferation ability
was detected using a Cell-Light™
5-ethynyl-2′-deoxyuridine (EdU) DNA Cell Proliferation kit
(Guangzhou RiboBio Co., Ltd., Guangzhou, China) according to the
manufacturer's protocol. Results of the proliferation assay were
observed under Apollo® 567 reaction reagent at room
temperature for 30 min in dark and counterstained with DAPI at room
temperature for 15 min and images from five random fields/well were
captured at ×200 magnification. Image-Pro Plus 6.0 software (Media
Cybernetics, Inc., Rockville, MD, USA) was used to calculate the
percentage of EdU-positive cells of all cells.
EPC migration assay
Migration of late EPCs was measured using a modified
Boyden chamber assay, as previously described (19). Briefly, following pretreatment with
20 µmol/l RES for 12 h, EPCs were cultured with TNF-α (10 ng/ml)
for another 24 h in a serum-free EGM2 medium. A total of
4×104 EPCs were added in the upper insert of the chamber
in an EGM2 medium (Lonza Group, Ltd.,) and also EGM2 medium with
fetal bovine serum was added to the bottom chamber. Following
incubation for 8 h at 37°C in an incubator with 5% CO2,
the membrane in the upper chamber was washed gently with PBS and
non-migratory cells were scraped using cotton swabs. Subsequently,
EPCs were fixed with 4% paraformaldehyde at room temperature for 30
min and stained with DAPI at room temperature for 15 min, and the
number of migratory cells in the bottom chamber in six randomly
selected fields/well was calculated under a fluorescent microscope
(Leica Microsystems GmbH, Wetzlar, Germany; DMI 4000;
magnification, ×100).
EPC adhesion assay
The adhesion assay was performed as previously
described (21). Following
pretreatment with 20 µmol/l RES for 12 h, EPCs were stimulated with
10 ng/ml TNF-α for 24 h. Subsequently, EPCs (1×104) were
placed onto fibronectin-coated 12-well plates and incubated at 37°C
for 30 min. Following washing of non-adherent cells with PBS, the
adherent cells were counted independently in six random fields/well
under an inverted phase contrast microscope (Leica Microsystems
GmbH; DM 1400B; magnification, ×100).
Tube formation assay
The tube formation ability of EPCs on Matrigel (BD
Biosciences, San Jose, CA, USA) was detected. Initially, a 96-well
plate precoated with 100 µl Matrigel at 37°C for 1 h was prepared.
Subsequently, 100 µl late EPCs (2×104 cells/ml) were
seeded into each well and incubated for 6 h at 37°C. Images of
enclosed networks of tubes were captured in six randomly selected
fields under an inverted phase contrast microscope (Leica
Microsystems GmbH; DM 1400B; magnification, ×200) and the length of
complete tubes formed/unit area was quantified by Image-Pro Plus
6.0 software (Media Cybernetics, Inc.).
Quantification of NO
The total amount of NO in the culture supernatant of
EPCs from three groups were, respectively measured using a Nitric
Oxide (NO) assay kit (cat. no. A012-1; Nanjing Jiancheng
Bioengineering Institute, Nanjing, China) based on the Griess
reaction. The total nitrite level was measured to assess NO levels
at 550 nm according to the manufacturer's protocol. As NO is easily
transformed to nitrite and nitrate in vivo, therefore the
total nitrite and nitrate level may represent the NO level. In the
presence of nitrate reductase in the kit that was used, nitrate is
reduced to nitrite via reduced NADPH and then nitrite level was
examined by the colorimetric assay kit.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
PrimeScript™ RT Master Mix (RR036A; Takara Biotechnology
Co., Ltd., Dalian, China) was used to synthesize cDNA at 37°C for
15 min. Subsequently, qPCR was performed using SYBR Premix Ex
Taq™ (RR420A; Takara Biotechnology Co., Ltd.) using the
synthesized cDNA as a template. The thermal cycling conditions were
as follows: 30 sec at 95°C for pre-denaturation, 40 cycles for 15
sec at 95°C for denaturation, 1 min at 59°C for annealing, and 10
sec at 80°C for elongation. The following gene-specific
oligonucleotide sequences were used: GAPDH,
5′-GGCACAGTCAAGGCTGAGAATG-3′ (forward) and
5′-ATGGTGGTGAAGACGCCAGTA-3′ (reverse); KLF2,
5′-TCGCACCTAAAGGCGCATC-3′ (forward) and 5′-TAGTGGCGGGTAAGCTCGTC-3′
(reverse); monocyte chemoattractant protein-1 (MCP-1),
5′-CTATGCAGGTCTCTGTCACGCTTC-3′ (forward) and
5′-CAGCCGACTCATTGGGATCA-3′ (reverse); and intercellular adhesion
molecule-1 (ICAM-1), 5′-GCTTCTGCCACCATCACTGTGTA-3′ (forward) and
5′-ATGAGGTTCTTGCCCACCTG-3′ (reverse). RT-qPCR was performed using a
LightCycler 480 Instrument II (Roche Diagnostics GmbH, Mannheim,
Germany). Amplification of GAPDH was performed as a control for
normalization. Fold changes in gene expression were calculated
using 2−∆∆Cq method (22).
Western blotting
Late EPC cells were divided into six groups with
different treatments: TNF-α group, DMSO group, 5RES (R) + TNF-α (T)
group, 10R + T group, 20R + T group and 50R + T group. The density
of EPCs in those six groups was 2×106 cell/well. In 5R +
T, 10R + T, 20R + T and 50R + T groups, EPCs were pretreated with
5, 10, 20, and 50 µmol/l RES at 37°C for 12 h, respectively, and
then cultured with TNF-α (10 ng/ml) at 37°C for 24 h. In TNF-α and
DMSO group, EPCs were cultured with 10 ng/ml TNF-α or the same
volume of DMSO at 37°C for 24 h, respectively.
Following the various treatments, late EPCs were
lysed using radioimmunoprecipitation assay lysis Solution Enhanced
(C1053; Applygen Technologies Inc., Beijing, China) and the lysates
were isolated by centrifugation at 12,000 × g and 4°C for 5 min.
The proteins concentration were determined by bicinchoninic acid
(BCA) method (Pierce™ BCA Protein Assay kit, Thermo
Fisher Scientific, Inc.). Then, total proteins with equal quality
(40 µg/lane) from each group were separated by 12% (weight/volume)
polyacrylamide gels. Prior to incubation with the primary antibody
against KLF2 (cat. no. SAB2108684; 1:500; Sigma-Aldrich; Merck
KGaA) overnight at 4°C, the separated proteins were transferred to
polyvinylidene fluoride membranes and non-specific binding proteins
were blocked with 5% bovine serum albumin (cat. no. ST-023;
Beyotime Institute of Biotechnology, Jiangsu, China)/0.01 mol/l
TBS-Tween-20 (TBST) for 60 min at room temperature. β-actin (cat.
no. AF0003; 1:1,000; Beyotime Institute of Biotechnology) was
selected as the control and incubated overnight at 4°C.
Subsequently, membranes were washed with TBST and incubated with
horseradish peroxidase-conjugated secondary antibody (cat. no.
SC-2357; 1:1,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
at room temperature for 60 min. Immunoreactive bands were
visualized by Amersham enhanced chemiluminescence (ECL) western
blotting detection reagent (GE Healthcare Life Sciences, Little
Chalfont, UK). Finally, a densitometric analysis for the western
blot was performed by GelPro Analyser 4.0 software (Media
Cybernetics, Inc., Rockville, MD, USA).
Statistical analysis
Data are presented as the mean ± standard error of
the mean. Statistical analysis was performed by one-way analysis of
variance and Tukey-Kramer post-hoc test was applied for multiple
comparisons. All the experiments were repeated three times. All
statistical analyses were performed using SPSS 16.0 software (SPSS,
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Effects of RES on viability of EPCs
treated with TNF-α
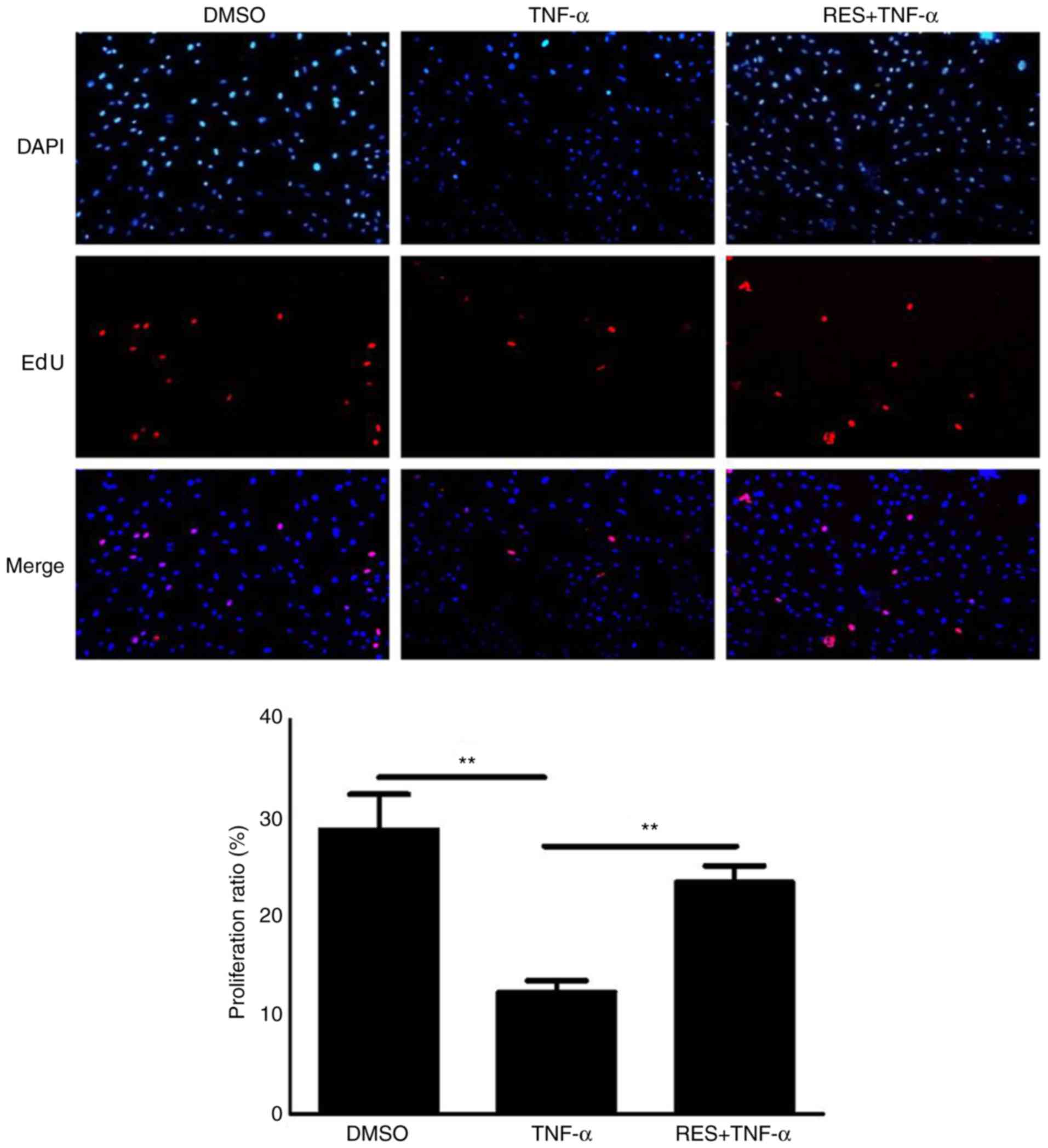
Based on the results of the EdU assay, the
proliferative potential of EPCs treated with TNF-α (10 ng/ml) was
markedly reduced compared with the DMSO group (P<0.01; Fig. 1). However, cell viability in the
RES + TNF-α group was significantly increased compared with the
TNF-α group (P<0.01; Fig.
1).
Effects of RES on the migration of
EPCs
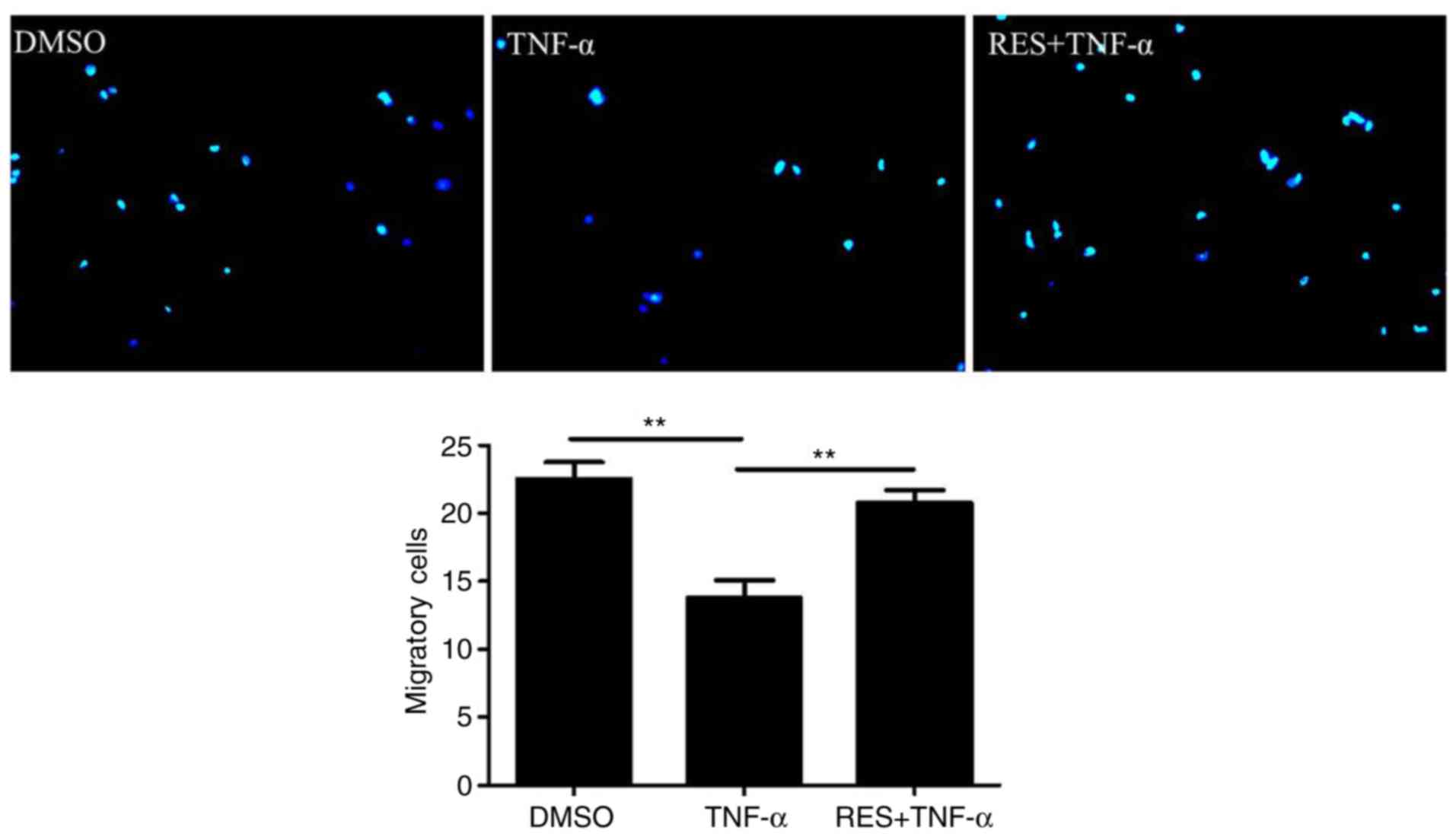
To determine the effect of RES on the migration of
EPCs, a migration assay was conducted. The results indicated that
TNF-α markedly inhibited the migration of EPCs (TNF-α group vs.
DMSO group, 13.75±2.63 vs. 22.5±2.52 cells/high-power field,
respectively; P<0.01; Fig. 2).
However, RES significantly enhanced the migration ability of EPCs
in TNF-α-treated cells; the number of migratory EPCs in the RES +
TNF-α group was markedly increased compared with the TNF-α group
(20.75±1.89 vs. 13.75±2.63 cells/high-power field, respectively;
P<0.01; Fig. 2).
RES improves the adhesion of EPCs
impaired by TNF-α in vitro
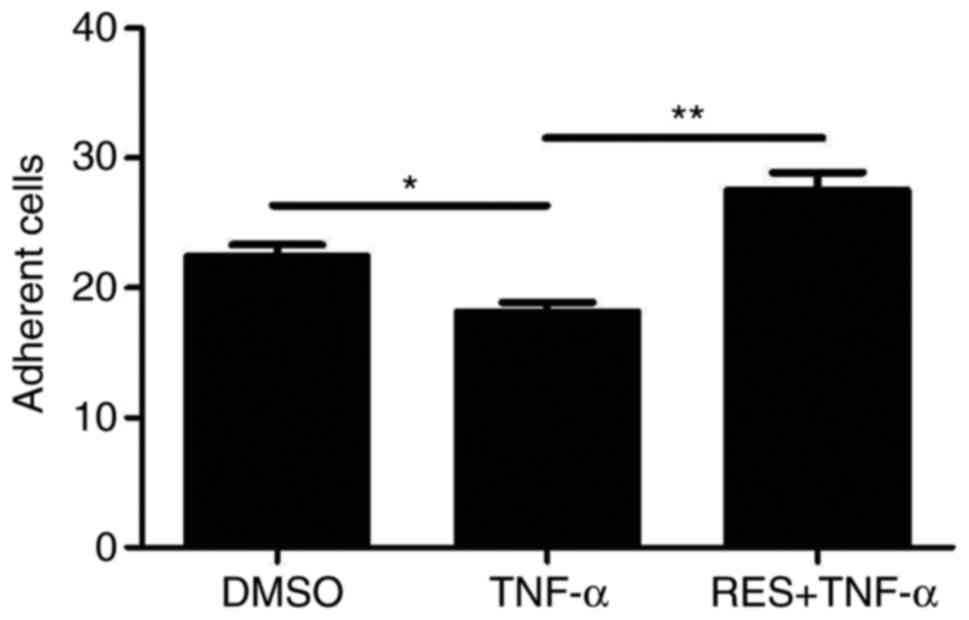
The present study also aimed to determine whether
RES affects the adhesion of late EPCs treated with TNF-α. The
results demonstrated that the adhesion potential of EPCs treated
with TNF-α was significantly lower compared with the DMSO group
(18.17±1.72 vs. 22.43±2.37, respectively; P<0.05; Fig. 3). However, the adhesion capacity of
EPCs was markedly elevated in the group that was pre-exposed to RES
(RES + TNF-α group) compared with the TNF-α only group (27.5±3.93
vs. 18.17±1.72, respectively; P<0.01; Fig. 3).
Effect of RES on the vasculogenesis of
EPCs
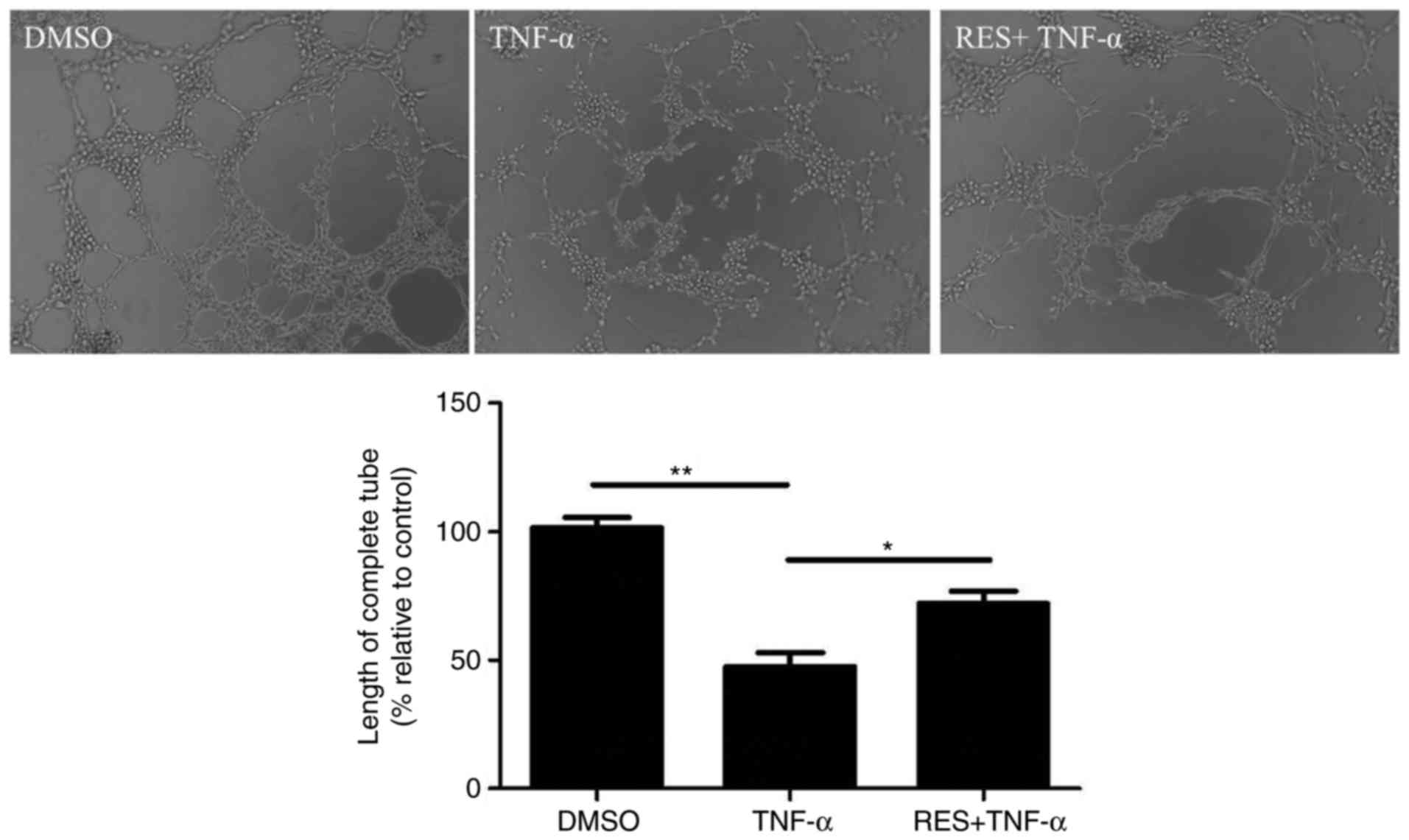
The neovascularization capacity of EPCs was
investigated using a vasculogenesis assay. TNF-α markedly inhibited
the length of tubules by EPCs compared with the DMSO control
(P<0.01; Fig. 4). By contrast,
tubule length increased in response to pretreatment with RES
compared with the treatment with TNF-α only (P<0.05; Fig. 4).
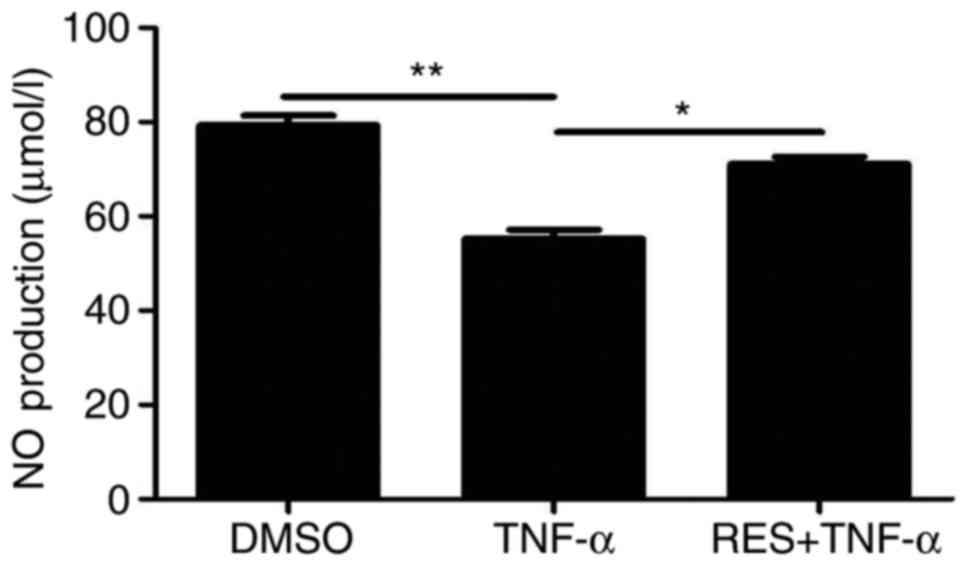
Effect of RES on the levels of NO
NO secretion by EPCs was detected using a
colorimetric assay kit in all three groups. NO secretion by EPCs
was reduced in the TNF-α group compared with the DMSO group
(P<0.01; Fig. 5). Conversely, a
significant increase in NO production was observed in the RES +
TNF-α group compared with the TNF-α group (P<0.05; Fig. 5). Therefore, RES alleviated the
TNF-α-induced inhibition of NO release in EPCs.
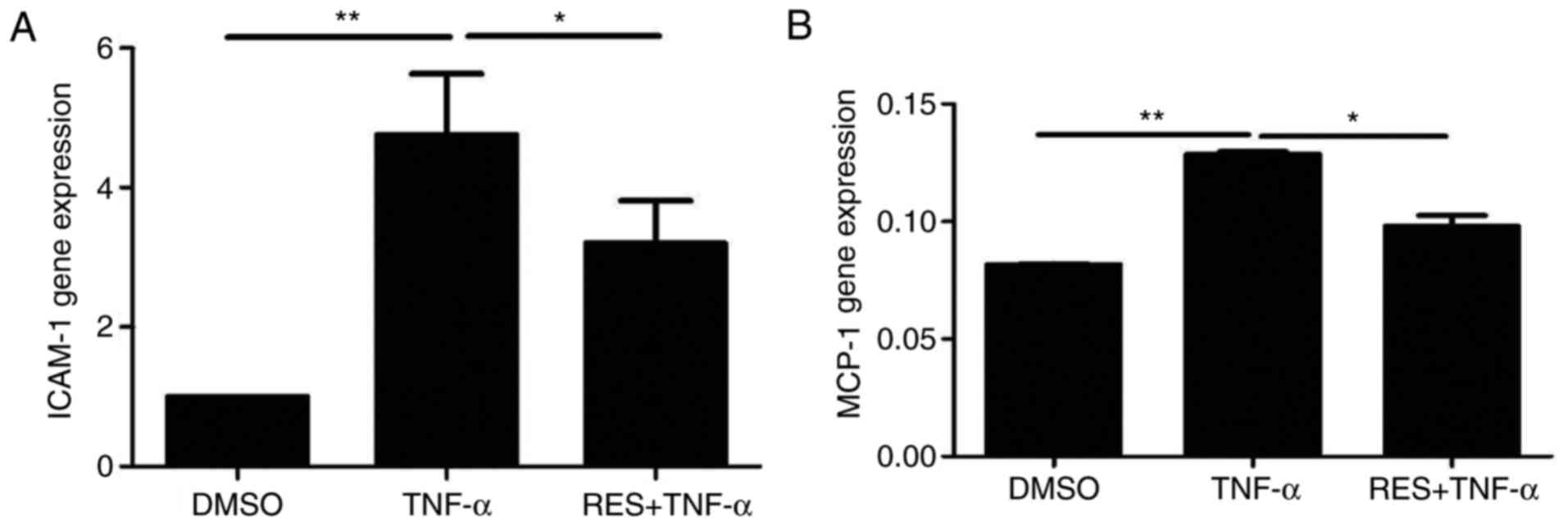
Effect of RES on the expression of the
inflammatory molecules ICAM-1 and MCP-1 in EPCs treated with
TNF-α
The mRNA levels of ICAM-1 and MCP-1 in the TNF-α
group were significantly increased compared with the DMSO group
(P<0.01; Fig. 6). EPCs
co-cultured with RES and TNF-α demonstrated markedly decreased mRNA
expression of ICAM-1 (Fig. 6A) and
MCP-1 (Fig. 6B) compared with the
respective TNF-α groups (P<0.05). The above results indicated
that RES may inhibit TNF-α-induced increases in the expression of
inflammatory molecules, including ICAM-1 and MCP-1, in late
EPCs.
Effect of RES on KLF2 mRNA and protein
expression in EPCs treated with TNF-α
The mRNA levels of KLF2 were significantly reduced
in late EPCs treated with TNF-α compared with EPCs treated with
DMSO (P<0.01; Fig. 7A).
However, KLF2 mRNA expression in the RES + TNF-α group was markedly
enhanced compared with the TNF-α only group (P<0.05; Fig. 7A). Subsequently, the protein levels
of KLF2 were determined in EPCs pretreated with RES at different
concentrations (5, 10, 20 and 50 µmol/l). The results demonstrated
that TNF-α inhibited KLF2 protein expression (Fig. 7B). However, the protein levels of
KLF2 were increased in a dose-dependent manner when EPCs were
treated with RES at concentrations between 5 and 20 µmol/l, and
KLF2 protein expression was decreased in EPCs following treatment
with 50 µmol/l RES compared with 20 µmol/l RES group (Fig. 7C). Therefore, these results
indicated that 20 µmol/l RES was the optimal concentration to
stimulate KLF2 protein expression in late EPCs.
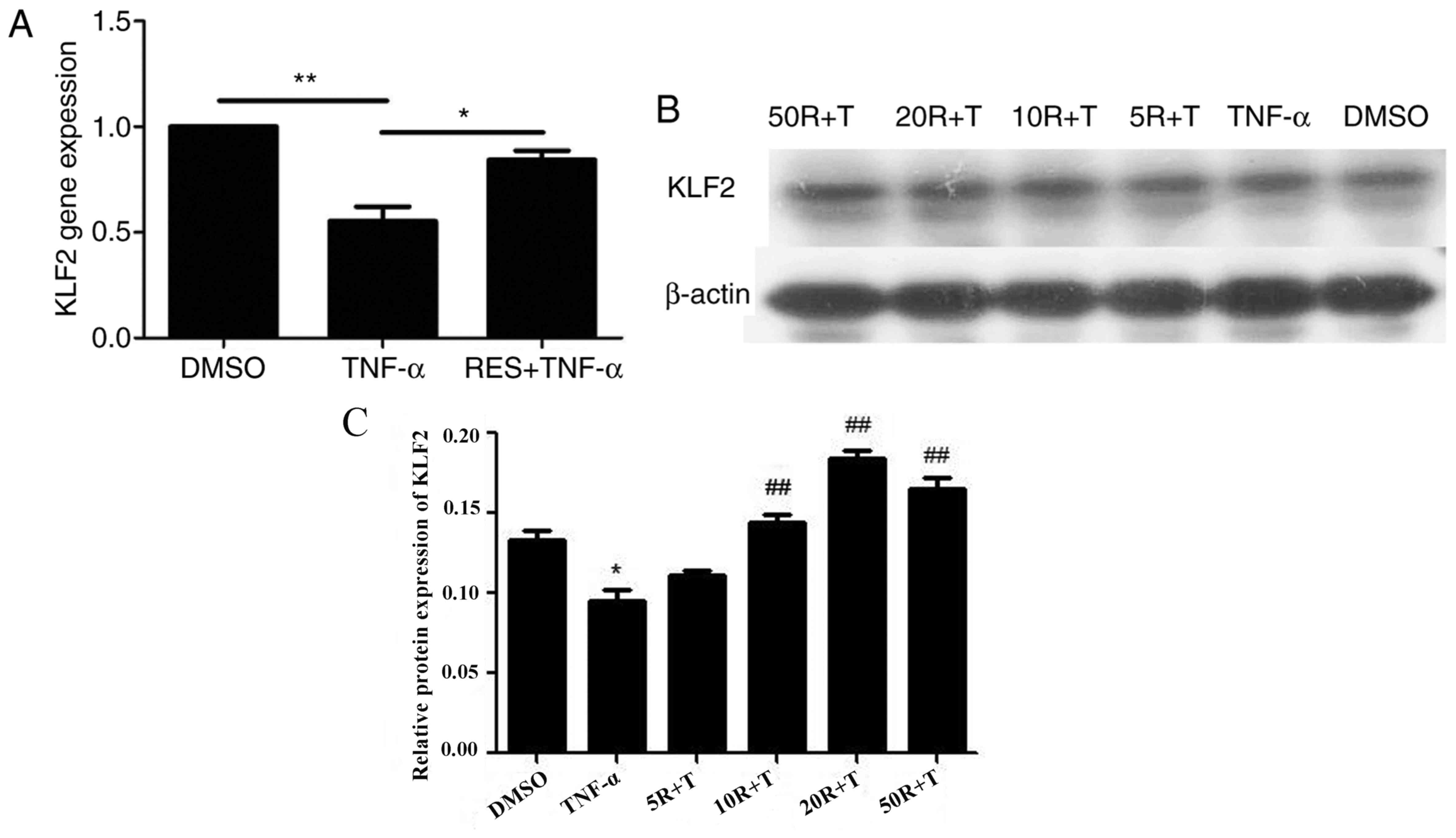 | Figure 7.Effect of RES on KLF2 mRNA and
protein expression in EPCs. (A) KLF2 mRNA expression was determined
by reverse transcription-quantitative polymerase chain reaction.
(B) KLF2 protein expression was determined by western blot
analysis. For western blotting, EPCs were pretreated with RES at
concentrations of 5, 10, 20 and 50 µmol/l for 12 h, and
subsequently, EPCs were cultured with 10 ng/ml TNF-α for 24 h.
β-actin was analyzed as a loading control. (C) Densitometric
analysis for the western blot was performed by GelPro Analyser 4.0
software. EPCs treated with DMSO served as the blank group.
*P<0.05 and **P<0.01, as indicated. In C, *P<0.05 vs. DMSO
groups, ##P<0.01 vs. TNF-α groups. RES, resveratrol;
KLF2, Krüppel-like factor-2; EPCs, endothelial progenitor cells;
DMSO, dimethyl sulfoxide; TNF-α, tumor necrosis factor-α; R,
resveratrol; T, tumor necrosis factor-α. |
Discussion
EPCs can promote plaque angiogenesis and restore
endothelial function to serve a protective role in atherosclerotic
disease (23). The biological
properties of EPCs have clinical implications for coronary
atherosclerotic disease, since cardiovascular risk factors can
negatively influence the number and function of EPCs (23,24).
A number of studies have demonstrated that RES delayed the
senescence of EPCs by augmenting telomerase activity to maintain
the appropriate levels and function of EPCs (25–27).
The present study aimed to determine the effect of RES on the
function of EPCs treated with TNF-α and the underlying
anti-inflammatory mechanisms, in addition to the potentially
protective role of RES in atherosclerotic disease.
In the current study, the effects of RES on the
proliferation, migration, adhesion, tube formation ability and NO
production of EPCs were evaluated following treatment with TNF-α.
The results demonstrated that RES attenuated TNF-α-induced damage
to the functioning of EPCs. These results are consistent with a
previous study by Wang et al (28), which demonstrated that RES may
promote the proliferation, adhesion and migration of EPCs in a
dose- and time-dependent manner. The aforementioned study also
indicated that RES increased the expression of vascular endothelial
growth factor to further induce vasculogenesis (28).
The results of the present study indicated that RES
promoted NO production and KLF2 expression in EPCs, and it has been
previously reported that endothelial NO release may protect cells
against AS (29). NO is
synthesized by inducible nitric oxide synthase and eNOS, where eNOS
acts as an anti-atherosclerotic factor (30). It has been previously demonstrated
that RES increased the expression of eNOS in endothelial cells
in vitro (31,32), which was mediated by the activation
of sirtuin 1 (SIRT1) and subsequent activation of the transcription
factor KLF2 (33). The
KLF2-dependent signaling pathway has been reported to enhance the
enzymatic activity of dimethylarginine dimethylaminohydrolase II,
which leads to the degradation of asymmetric dimethylarginine, an
endogenous inhibitor of eNOS (9,34).
Therefore, we hypothesized that RES may increase NO production in
EPCs via upregulation of KLF2.
Piga et al (35) indicated that adhesion and migration
of human aortic endothelial cells were influenced by overexpression
of ICAM-1 and MCP-1. In addition, upregulation of ICAM-1 and MCP-1
was considered a marker for AS progression via TNF-α-induced
activation of the nuclear factor-κB (NF-κB) pathway (36). Therefore, in the present study, the
expression of ICAM-1 and MCP-1 was detected in EPCs following
inflammatory damage induced by TNF-α. Based on the results of the
present study, it may be hypothesized that TNF-α may promote the
expression of ICAM-1 and MCP-1 in EPCs, which may be responsible
for alterations in the function of EPCs during the development of
AS. Furthermore, the present study demonstrated that RES may
attenuate inflammatory damage of EPCs via downregulation of ICAM-1
and MCP-1. Similar to the results of the present study, Zhu et
al (37) demonstrated that RES
may reduce TNF-α-activated expression of MCP-1 via inhibition of
NF-κB transcriptional activity in adipocytes. Xiao et al
(38) indicated that RES inhibited
the expression of TNF-α-activated ICAM-1 and vascular cell adhesion
molecule-1 (VCAM-1) in endothelial cells. Additionally, several
studies have indicated that KLF2 may inhibit inflammation via
suppression of a number of adhesion molecules, including VCAM-1,
E-selectin and MCP-1 (39,40). Therefore, we hypothesized that RES
may downregulate the expression of ICAM-1 and MCP-1 in response to
proinflammatory stimuli by increasing the expression of KLF2.
KLF2 has been reported to be involved in the
prevention of the progression of AS via anti-inflammatory action
and the regulation of eNOS expression (41,42).
Based on the results of the present study and previous reports, we
hypothesized that RES may have potential in the development of
novel therapies against AS, and the anti-atherosclerotic effects of
RES may be attributed to the promotion of KLF2 expression and
subsequent inhibition of inflammation and promotion of NO
release.
In conclusion, the results of the present study
demonstrated that RES increased the proliferation, migration,
adhesion, tube formation ability and NO release of EPCs and
upregulated KLF2 expression following treatment with TNF-α, and
also inhibited the TNF-α-stimulated expression of ICAM-1 and MCP-1.
Therefore, these results indicate that RES may alter the growth
properties of EPCs treated with TNF-α by increasing NO production
and ameliorating inflammatory damage by reducing the expression
levels of inflammatory molecules, including ICAM-1 and MCP-1,
through regulation of KLF2-associated signaling pathways.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 31570941, 31270993
and 81700406), the Program for New Century Excellent Talents in
University (grant no. NCET-10-0922), the Natural Science Foundation
of Shandong Province (grant nos. ZR2013CQ032 and ZR2014JL018,
ZR2016CM20), Administration of Traditional Chinese Medicine of
Shandong Province (grants no. 2016WS0667 and 2015–239) and Shandong
Province Higher Educational Science and Technology Program (grants
no. J14LK59, J14LK12 and J15LK08).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
HC, HL and MC: Conception and design of the
research. XG, HY, XZ, XC and XL: Acquisition of data, analysis and
interpretation of data and statistical analysis. HC and HL:
Drafting the manuscript. MC: Revision of manuscript for important
intellectual content.
Ethics approval and consent to
participate
The present study was approved by the Medical Ethics
Committee of Weifang Medical University (Weifang, China) and was
performed in accordance with the committee guidelines.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hill JM, Zalos G, Halcox JP, Schenke WH,
Waclawiw MA, Quyyumi AA and Finkel T: Circulating endothelial
progenitor cells, vascular function, and cardiovascular risk. N
Engl J Med. 348:593–600. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schmidt-Lucke C, Rössig L, Fichtlscherer
S, Vasa M, Britten M, Kämper U, Dimmeler S and Zeiher AM: Reduced
number of circulating endothelial progenitor cells predicts future
cardiovascular events: Proof of concept for the clinical importance
of endogenous vascular repair. Circulation. 111:2981–2987. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zampetaki A, Kirton JP and Xu Q: Vascular
repair by endothelial progenitor cells. Cardiovasc Res. 78:413–421.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Miller-Kasprzak E and Jagodziński PP:
Endothelial progenitor cells as a new agent contributing to
vascular repair. Arch Immunol Ther Exp (Warsz). 55:247–259. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Paramo JA, Rodríguez JA and Orbe J:
Atherosclerosis in inflammatory diseases. Med Clin (Barc).
128:749–756. 2007.(In Spanish). PubMed/NCBI
|
|
6
|
Tousoulis D, Andreou I, Antoniades C,
Tentolouris C and Stefanadis C: Role of inflammation and oxidative
stress in endothelial progenitor cell function and mobilization:
Therapeutic implications for cardiovascular diseases.
Atherosclerosis. 201:236–247. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jang M, Cai L, Udeani GO, Slowing KV,
Thomas CF, Beecher CW, Fong HH, Farnsworth NR, Kinghorn AD, Mehta
RG, et al: Cancer chemopreventive activity of resveratrol, a
natural product derived from grapes. Science. 275:218–220. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baur JA and Sinclair DA: Therapeutic
potential of resveratrol: The in vivo evidence. Nat Rev Drug
Discov. 5:493–506. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schmitt CA, Heiss EH and Dirsch VM: Effect
of resveratrol on endothelial cell function: Molecular mechanisms.
Biofactors. 36:342–349. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang H, Yang YJ, Qian HY, Zhang Q, Xu H
and Li JJ: Resveratrol in cardiovascular disease: What is known
from current research? Heart Fail Rev. 17:437–448. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bruunsgaard H, Skinhøj P, Pedersen AN,
Schroll M and Pedersen BK: Ageing, tumour necrosis factor-alpha
(TNF-alpha) and atherosclerosis. Clin Exp Immunol. 121:255–260.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang H, Zhang J, Ungvari Z and Zhang C:
Resveratrol improves endothelial function: Role of TNF{alpha} and
vascular oxidative stress. Arterioscler Thromb Vasc Biol.
29:1164–1171. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pearson R, Fleetwood J, Eaton S, Crossley
M and Bao S: Krüppel-like transcription factors: A functional
family. Int J Biochem Cell Biol. 40:1996–2001. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lin Z, Kumar A, SenBanerjee S,
Staniszewski K, Parmar K, Vaughan DE, Gimbrone MA Jr,
Balasubramanian V, García-Cardeña G and Jain MK: Kruppel-like
factor 2 (KLF2) regulates endothelial thrombotic function. Circ
Res. 96:e48–e57. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu J, Bohanan CS, Neumann JC and Lingrel
JB: KLF2 transcription factor modulates blood vessel maturation
through smooth muscle cell migration. J Biol Chem. 283:3942–3950.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sebzda E, Zou Z, Lee JS, Wang T and Kahn
ML: Transcription factor KLF2 regulates the migration of naive T
cells by restricting chemokine receptor expression patterns. Nat
Immunol. 9:292–300. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dekker RJ, van Thienen JV, Rohlena J, de
Jager SC, Elderkamp YW, Seppen J, de Vries CJ, Biessen EA, van
Berkel TJ, Pannekoek H and Horrevoets AJ: Endothelial KLF2 links
local arterial shear stress levels to the expression of vascular
tone-regulating genes. Am J Pathol. 167:609–618. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kumar A, Lin Z, SenBanerjee S and Jain MK:
Tumor necrosis factor alpha-mediated reduction of KLF2 is due to
inhibition of MEF2 by NF-kappaB and histone deacetylases. Mol Cell
Biol. 25:5893–5903. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li H, Zhang X, Guan X, Cui X, Wang Y, Chu
H and Cheng M: Advanced glycation end products impair the
migration, adhesion and secretion potentials of late endothelial
progenitor cells. Cardiovasc Diabetol. 11:462012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen YH, Lin SJ, Lin FY, Wu TC, Tsao CR,
Huang PH, Liu PL, Chen YL and Chen JW: High glucose impairs early
and late endothelial progenitor cells by modifying nitric
oxide-related but not oxidative stress-mediated mechanisms.
Diabetes. 56:1559–1568. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liang C, YR HT, He Z, Jiang Q, Wu J, Zhen
Y, Fan M and Wu Z: Rosiglitazone via upregulation of Akt/eNOS
pathways attenuates dysfunction of endothelial progenitor cells,
induced by advanced glycation end products. Br J Pharmacol.
158:1865–1873. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Werner N and Nickenig G: Clinical and
therapeutical implications of EPC biology in atherosclerosis. J
Cell Mol Med. 10:318–332. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ku IA, Imboden JB, Hsue PY and Ganz P:
Rheumatoid arthritis: Model of systemic inflammation driving
atherosclerosis. Circ J. 73:977–985. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xia L, Wang XX, Hu XS, Guo XG, Shang YP,
Chen HJ, Zeng CL, Zhang FR and Chen JZ: Resveratrol reduces
endothelial progenitor cells senescence through augmentation of
telomerase activity by Akt-dependent mechanisms. Br J Pharmacol.
155:387–394. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gu J, Wang CQ, Zhang DD, Fan HH, He B,
Wang BY and Huang DJ: Effect of resveratrol on reendothelialization
and neointimal formation in intimal injury model. Chin J
Arterioscler. 14:829–834. 2006.
|
|
27
|
Wang XB, Zhu L, Huang J, Yin YG, Kong XQ,
Rong QF, Shi AW and Cao KJ: Resveratrol-induced augmentation of
telomerase activity delays senescence of endothelial progenitor
cells. Chin Med J (Engl). 124:4310–4315. 2011.PubMed/NCBI
|
|
28
|
Wang XB, Huang J, Zou JG, Su EB, Shan QJ,
Yang ZJ and Cao KJ: Effects of resveratrol on number and activity
of endothelial progenitor cells from human peripheral blood. Clin
Exp Pharmacol Physiol. 34:1109–1115. 2007.PubMed/NCBI
|
|
29
|
Matthys KE and Bult H: Nitric oxide
function in atherosclerosis. Mediators Inflamm. 6:3–21. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Leifeld L, Fielenbach M, Dumoulin FL,
Speidel N, Sauerbruch T and Spengler U: Inducible nitric oxide
synthase (iNOS) and endothelial nitric oxide synthase (eNOS)
expression in fulminant hepatic failure. J Hepatol. 37:613–619.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Klinge CM, Blankenship KA, Risinger KE,
Bhatnagar S, Noisin EL, Sumanasekera WK, Zhao L, Brey DM and
Keynton RS: Resveratrol and estradiol rapidly activate MAPK
signaling through estrogen receptors alpha and beta in endothelial
cells. J Biol Chem. 280:7460–7468. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wallerath T, Deckert G, Ternes T, Anderson
H, Li H, Witte K and Förstermann U: Resveratrol, a polyphenolic
phytoalexin present in red wine, enhances expression and activity
of endothelial nitric oxide synthase. Circulation. 106:1652–1658.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gracia-Sancho J, Villarreal G Jr, Zhang Y
and García-Cardeña G: Activation of SIRT1 by resveratrol induces
KLF2 expression conferring an endothelial vasoprotective
phenotypee. Cardiovasc Res. 85:514–519. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Scalera F, Fulge B, Martens-Lobenhoffer J,
Heimburg A and Bode-Böger SM: Red wine decreases asymmetric
dimethylarginine via SIRT1 induction in human endothelial cells.
Biochem Biophys Res Commun. 390:703–709. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Piga R, Naito Y, Kokura S, Handa O and
Yoshikawa T: Short-term high glucose exposure induces
monocyte-endothelial cells adhesion and transmigration by
increasing VCAM-1 and MCP-1 expression in human aortic endothelial
cells. Atherosclerosis. 193:328–334. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang Y, Zhao X, Wang YS, Song SL, Liang H
and Ji AG: An extract from medical leech improve the function of
endothelial cells in vitro and attenuates atherosclerosis in ApoE
null mice by reducing macrophages in the lesions. Biochem Biophys
Res Commun. 455:119–125. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhu J, Yong W, Wu X, Yu Y, Lv J and Liu C,
Mao X, Zhu Y, Xu K, Han X and Liu C: Anti-inflammatory effect of
resveratrol on TNF-alpha-induced MCP-1 expression in adipocytes.
Biochem Biophys Res Commun. 369:471–477. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xiao J, Song J, Hodara V, Ford A, Wang XL,
Shi Q, Chen L and Vandeberg JL: Protective effects of resveratrol
on TNF-α-induced endothelial cytotoxicity in baboon femoral
arterial endothelial cells. J Diabetes Res 2013. 2013.doi:
10.1155/2013/185172.
|
|
39
|
Senbanerjee S, Lin Z, Atkins GB, Greif DM,
Rao RM, Kumar A, Feinberg MW, Chen Z, Simon DI, Luscinskas FW, et
al: KLF2 is a novel transcriptional regulator of endothelial
proinflammatory activation. J Exp Med. 199:1305–1315. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hampole AV, Mahabeleshwar GH, Sharma N and
Jain MK: Abstract 5546: Kruppel-like factor 2 (KLF)2 inhibits
macrophage pro-inflammatory activation. Circulation.
120:S1113–S1114. 2009.
|
|
41
|
Zhou XB and Yang LX: Krüppel-like factor 2
and atherosclerosis. Adv Cardiovasc Dis. 33:224–246. 2012.(In
Chinese).
|
|
42
|
Jia YJ and Li JJ: Research Progress of
KLF2 anti-atherosclerosis. Mol Cardiol China. 5:303–307. 2012.(In
Chinese).
|