Introduction
Low back pain (LBP) is a common condition; according
to the statistics, at least 70% of people will suffer from LBP at
some point in their life in western countries (1). The phenomenon of LBP is exacerbated
by the aging process. In addition to the influence on life quality
and self-care ability, LBP places a huge burden on the social
economy (2). In 2013 alone, the
cost of low back and neck pain for social healthcare in the USA was
$87.6 billion, the third highest among individual and public health
costs (3). The leading cause of
LBP is symptomatic intervertebral disc degeneration disease. The
peripheral annulus, inner nucleus pulposus and each end of the
cartilage endplate together form an intervertebral disc, and the
discs create vertebral bodies in conjunction with each other and
bring flexibility, cushioning and stress transmission (4). The disc is the biggest avascular
organ and has a hypoxic microenvironment in its normal state
(5). During degeneration, changes
in the material composition of the inner disc induce changes in its
mechanical properties. The loading force exceeding the endurance of
the intervertebral disc leads to the formation of annulus fissures;
subsequently, granulation tissue and new blood vessels grow into
the fissures, simultaneously carrying inflammatory cells for
infiltration (6). In adults, the
extent of the intervertebral disc nerve endings does not exceed
one-third of the outer annulus, and chronic LBP is often thought to
be caused by a compression of the nerve endings deep in the
intervertebral disc (7).
Stefanakis et al (8) found
that there was new vascular nerve ingrowth in annulus fissures.
Freemont et al (9)
presented a point of view that the blood vessels that grew into the
intervertebral disc produced nerve growth factor (NGF), and the
nociceptive fibers expressed a high-affinity NGF receptor that
followed the growth of the blood vessels into the degenerated
intervertebral disc. A study showed that neovascularization was
also one of the differences between painful degenerative discs and
asymptomatic degenerative intervertebral discs (10). Therefore, angiogenesis after
degeneration of the intervertebral disc is important in the
occurrence of LBP.
Stromal cell-derived factor 1 (SDF1), also known as
C-X-C family chemokine ligand 12 (CXCL12), was initially identified
as a pre-B-cell growth stimulating factor (11). CXCR4 is a
seven-transmembrane-spanning G protein-coupled receptor and was
identified first in peripheral blood leucocytes (12), binding to SDF1 specifically to form
the SDF1/CXCR4 axis. The SDF1/CXCR4 axis participates not only in
hematopoiesis (13), the immune
response (14) and organ
development (15), but also in
vascular remodeling or neovascularization (16). There are some reports that the
SDF1/CXCR4 axis is involved in angiogenesis in some repair
processes for tissue damage (17,18).
The formation of fissures under the condition of an unusual loading
force is a damage process, and the ingrowth of new blood vessels
and granulation tissue is considered a repair process; hence, it
was speculated that the SDF1/CXCR4 axis may be involved in this
pathological activity.
Whether the SDF1/CXCR4 axis plays a role in disc
angiogenesis and what role it plays is a key interest. The present
study sought to determine the influence of the SDF1/CXCR4 axis on
disc angiogenesis by regulating SDF1 expression in nucleus pulposus
cells (NPCs) and inhibiting superficial CXCR4 in vascular
endothelial cells (VECs) using a molecular compound. This may help
create a full understanding of disc degeneration.
Materials and methods
Cell isolation and culture
Degenerative disc tissues were obtained from the
Department of Orthopedics in The First Affiliated Hospital of
Chongqing Medical University (from July 2017 to March 2018), and
all 10 patients were diagnosed with disc degeneration diseases
(lumbar disc herniation, lumbar spinal stenosis or
spondylolisthesis). Informed consent was obtained from the donors,
and the experimental protocol was approved by the ethics committee
of Chongqing Medical University. All specimens was evaluated
according to the Pfirrmann classification (19) of preoperative lumbar MRI images,
and all specimens were grade III and above (Table I).
 | Table I.Specimens data for nucleus pulposus
cell isolation. |
Table I.
Specimens data for nucleus pulposus
cell isolation.
| Specimen no. | Sex | Age, years | Segment | Pfirrmann
grade |
|---|
| 1 | Male | 68 | L4-5 | IV |
| 2 | Female | 56 | L4-5 | IV |
| 3 | Female | 60 | L4-S1 | V |
| 4 | Female | 54 | L5-S1 | III |
| 5 | Male | 55 | L4-5 | IV |
| 6 | Female | 63 | L5-S1 | V |
| 7 | Male | 57 | L4-S1 | IV |
| 8 | Female | 60 | L5-S1 | IV |
| 9 | Female | 52 | L4-5 | III |
| 10 | Male | 60 | L4-5 | V |
Blood was flushed from disc tissues using PBS buffer
solution and the nucleus pulposus was carefully separated. Type II
collagenase (2%) was used to digest the nucleus pulposus tissue for
8 h at 37°C the primary NPC suspension was collected. The NPCs were
expanded in a monolayer culture with DMEM/F12 (HyClone; GE
Healthcare Life Sciences) containing 15% FBS (CellMax Life). The
NPCs were incubated at 37°C in a humid atmosphere with 5%
CO2. The medium was changed every 3 days, and the cells
were passaged when confluency reached 80%. The passage 1 (P1) cells
were chosen for subsequent experiments.
Human umbilical vein endothelial cells (cat. no.
hy926; Shanghai Cell Bank, Chinese Academy of Sciences) were
cultured with RPMI-1640 (HyClone; GE Healthcare Life Sciences)
containing 10% FBS. The medium wa8s changed every 3 days, and the
cells were passaged when confluency reached 80%. To inhibit CXCR4,
VECs were exposed to a final concentration of 1 µM AMD3100 (Selleck
Chemicals) for 1 h at 37°C.
Virus transfection of NPCs
Upregulating SDF1 adenovirus
transfection
A total of 105 NPCs were plated in
six-well plates overnight, then adenovirus-SDF1 (ADV-SDF1; Shanghai
GenePharma Co., Ltd.) was added to the wells [multiplicity of
infection (MOI)=100], and adenovirus-negative control (ADV-NC;
Shanghai GenePharma Co., Ltd.) was added in the same way for the
control group. The plate was gently shaken and incubated in a 37°C
incubator; ~12 h later, the medium was changed and culture of the
NPCs continued.
Downregulating SDF1 lentivirus
transfection
A total of 105 NPCs were plated in
six-well plates overnight, and then, lentivirus-SDF1-RNA
interference (RNAi; LV-SDF1-RNAi) [sequence of small interfering
(si)RNA: 5′-GTGCATTGACCCGAAGCTAAA-3′; Shanghai GeneChem Co., Ltd.]
was added to the wells (MOI=10) in combination with polybrene at a
final concentration of 5 µg/ml; lentivirus-negative control (LV-NC;
Shanghai GeneChem Co., Ltd.) was added in the same way for the
control group. The plate was gently shaken and incubated in a 37°C
incubator for 6 h, and then the medium was changed. After
transfection for 24–48 h, the mRNA expression was determined by
reverse transcription quantitative (RT-q)PCR; after transfection
for 48–72 h, protein expression was determined by western
blotting.
RT-qPCR
The NPC medium was discarded, and the NPCs were
washed three times using a PBS solution. Total RNA was extracted by
TRIzol® (Thermo Fisher Scientific, Inc.). The
complementary DNA (cDNA) was reverse transcribed using a RevertAid
First Stand cDNA Synthesis kit (Thermo Fisher Scientific, Inc.)
following the manufacturer's protocol as follows: 25°C for 5 min,
42°C for 60 min and 70°C for 5 min. The cDNA was subsequently used
to perform qPCR with SYBR Select Master Mix (Thermo Fisher
Scientific, Inc.) in an ABI-7500 Real-Time PCR system (ABI; Thermo
Fisher Scientific, Inc.). The final reaction volume was 10 µl and
consisted of 1 µl cDNA, 1 µl primer (5 µM/l), 3 µl nuclease-free
water and 5 µl SYBR master mix. SDF1 oligonucleotide primers
(forward, 5′-TCAGCCTGAGCTACAGATGCC-3′; reverse,
5′-TCTGAAGGGCACAGTTTGGAG-3′; synthesized by Takara Bio, Inc.) and
GAPDH oligonucleotide primers (forward,
5′-CGGAGTCAACGGATTCGGTCGTAT-3′; reverse,
5′-AGCCTTCTCCATGGTGGTGAAGAC-3′; synthesized by Takara Bio, Inc.)
were used. The thermal cycling was performed as follows: 50°C for 2
min, 95°C for 2 min, 40 cycles of 95°C for 3 sec and 60°C for 30
sec. The quantification cycle (Cq) value was normalized to GAPDH
(20). All samples were analyzed
in triplicate.
Western blotting
The lysis of NPCs was performed for total protein
with a RIPA lysis buffer (Beyotime Institute of Biotechnology). The
concentrations of every group were tested using a BCA protein
quantitation kit (Beyotime Institute of Biotechnology). Proteins
from cell lysates were separated via 12% SDS-PAGE (Beyotime
Institute of Biotechnology) and 40 µg protein was loaded per lane,
then transferred to polyvinylidene fluoride membranes (EMD
Millipore), which were blocked using 5% skimmed milk for 1 h at
room temperature. The membranes were incubated at 4°C overnight
with primary antibodies against SDF1 (1:1,000; ab9797, Abcam) and
GAPDH (1:3,000; 10494-1-AP, Proteintech Group, Inc.). After being
washed in a TBST solution, the membranes were incubated with
horseradish peroxidase-conjugated secondary antibodies (1:1,000;
A0208, Beyotime Institute of Biotechnology) for 1 h at 37°C. The
protein bands were detected using an ECL kit (Wanleibio Co., Ltd.)
and a chemiluminescence imaging system (Fusion SOLO S, VILBER).
Protein expression was quantified using EvolutionCapt SL6 software
(v16.0.8.0, Vilber Lourmat Sté).
Immunofluorescence
The coverslips for each condition were taken out of
24-well plates. The NPCs were fixed with 4% paraformaldehyde for 15
min and permeabilized with 0.1% Triton X-100 for 3 min at room
temperature. Then, NPCs were blocked with goat serum
(Sigma-Aldrich; Merck KGaA). Subsequently, NPCs were incubated with
SDF1 primary antibody (1:200; ab9797, Abcam), type II collagen
primary antibody (1:200; ab185430, Abcam) and aggrecan primary
antibody (1:200; ab36861, Abcam) at 4°C overnight. The next day,
the slides were washed with PBS solution three times and incubated
with FITC or Cy3 conjugated secondary antibodies (A0562 and A0521,
Beyotime Institute of Biotechnology)for 1 h at 37°C. Cell nuclei
were stained with DAPI (Beyotime Institute of Biotechnology)for 3
min at room temperature. Images were gathered using a fluorescence
microscope (magnification, ×200; Leica Microsystems, Inc.).
NPC conditioned medium (CM)
preparation
According to the literature (21), NPCs from the SDF1 upregulation
(Up), degeneration (D) and SDF1 downregulation (Down) groups were
prepared and counted. For each group, 5×105 cells were
seeded in a T25 flask, to which 4 ml DMEM/F12 medium with 10% FBS
was added. These NPCs were incubated at 37°C for 3 days. Then, the
culture supernatant was obtained and centrifuged at 12,880 × g for
10 mins at 4°C. The supernatant collected from these NPCs was
defined as their CM. The CM of the NPCs was stored at −80°C and
used within 2 weeks.
Cell counting kit-8 (CCK-8) assay
A total of 3,000 VECs were seeded in 96-well plates
overnight. The mixed medium was composed of NPC CM and complete
medium, at a ratio of 6:4. From each group of NPCs, 100 µl mixed
medium was added to the relevant wells. After 24 h of incubation,
10 µl CCK-8 solution (Selleck Chemicals) was added to each well.
After an additional 1 h incubation at 37°C, the absorbance was
measured spectrophotometrically at 450 nm. Cell viability was
calculated according to the formula: Cell viability (%)=[AX
group-Ablank group]/[AD
group-Ablank group]x100.
Transwell cell migration assay
From each group, 5×104 NPCs were seeded
in 24-well plates and each well was filled with 500 µl medium
containing 10% FBS. A total of 104 VECs were seeded in a
Transwell filter (Corning, Inc.) with 100 µl of a medium containing
5% FBS. The two sets of cells were cultured separately overnight.
Filters were placed into the relevant wells the next day, keeping
the membrane of the filter immersed in the medium of the lower
chamber. After 12 h of co-culture, the upper filters were removed
and the cells of the upper membrane surface were removed with a
cotton swab. The filters were fixed in 4% paraformaldehyde solution
for 15 mins at room temperature. After washing with PBS solution
three times, the filters were immersed in 0.1% crystal violet
solution for 10 mins at room temperature. The excess crystal violet
was rinsed with PBS solution. The images were captured using an
inverted light microscope (Leica Microsystems GmbH). A total of
five random fields of view in each group were acquired and analyzed
using ImageJ software (v1.51; National Institutes of Health).
Tube formation assay
The serum starved VECs were prepared one night in
advance. Matrigel (BD Biosciences) was melted on ice, and 50 µl of
Matrigel was placed into the wells of a 96-well plate, ensuring
that the Matrigel was homogeneous. The Matrigel was polymerized at
37°C for 30 min. The VECs were acquired and resuspended using a
mixed medium (NPC CM:complete medium with 10% FBS at a ratio of
6:4). A total of 2×104 VECs in 100 µl mixed medium were
placed onto the Matrigel coated wells and incubated at 37°C for
another 4 h. The tube structure was formed and images were captured
using an inverted light microscope (Leica Microsystems GmbH). A
total of five random fields of view in each group were acquired and
analyzed using ImageJ software (v1.51, National Institutes of
Health).
Statistical analysis
All data are presented as the mean ± SD. All
experiments were repeated three times. SPSS 19.0 (IBM Corp.) was
used for the statistical analysis. Single variable multigroup data
were analyzed by a one-way ANOVA; two variable multigroup data were
analyzed by a two-way ANOVA and a Tukey's post hoc test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Culture and identification of primary
NPCs
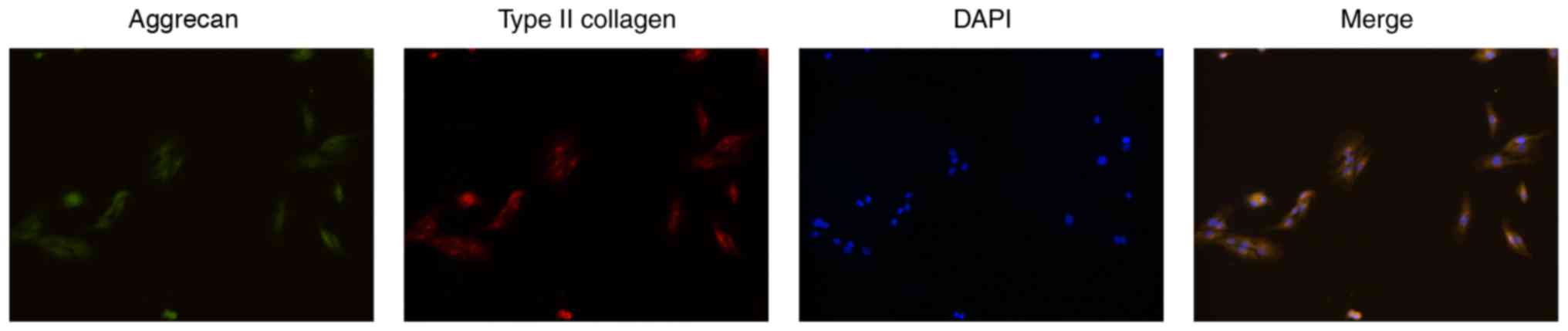
The primary NPCs were obtained and monolayer
cultured in vitro. Firstly, the phenotype of the NPCs was
identified by cell immunofluorescence of type II collagen and
aggrecan (22). The acquired
fluorescent images showed that the cells were polygonal and
simultaneously appeared as having green fluorescence (aggrecan) and
red fluorescence (type II collagen), still showing a
chondrocyte-like phenotype, and were identified as NPCs (Fig. 1). The P1 generation cells were used
for subsequent experiments to keep the phenotype to the greatest
extent.
Upregulation and downregulation of
SDF1 expression in NPCs
To explore the effect of the different expression of
SDF1 in NPCs on VECs, it was first necessary to obtain NPCs with
different expression levels of SDF1. The expression of SDF1 in NPCs
was regulated by virus transfection and identified using RT-qPCR,
western blotting and cellular immunofluorescence technology. The
NPCs not treated with the virus were defined as the D group. After
transfection with the adenovirus carrying the SDF1 sequence, total
RNA and protein were obtained. The PCR results showed that the
relative mRNA expression of SDF1 in the Up group was significantly
upregulated compared with the D and negative control (NC) groups
(Fig. 2A), and the western
blotting results showed that the expression of SDF1 in the Up group
was significantly upregulated compared with the D and NC groups
(Fig. 2C and D). The cells of the
adenovirus transfected group were defined as the Up group. After
transfection with a lentivirus carrying siRNA sequences, total RNA
and protein were obtained. The qPCR results showed that the
relative mRNA expression of SDF1 in the Down group was
significantly downregulated compared with the D and NC groups
(Fig. 2B), and the western
blotting results showed that the expression of SDF1 in the Down
group was significantly downregulated compared with the D and NC
groups (Fig. 2E and F). The cells
of the lentivirus transfection group were defined as the Down
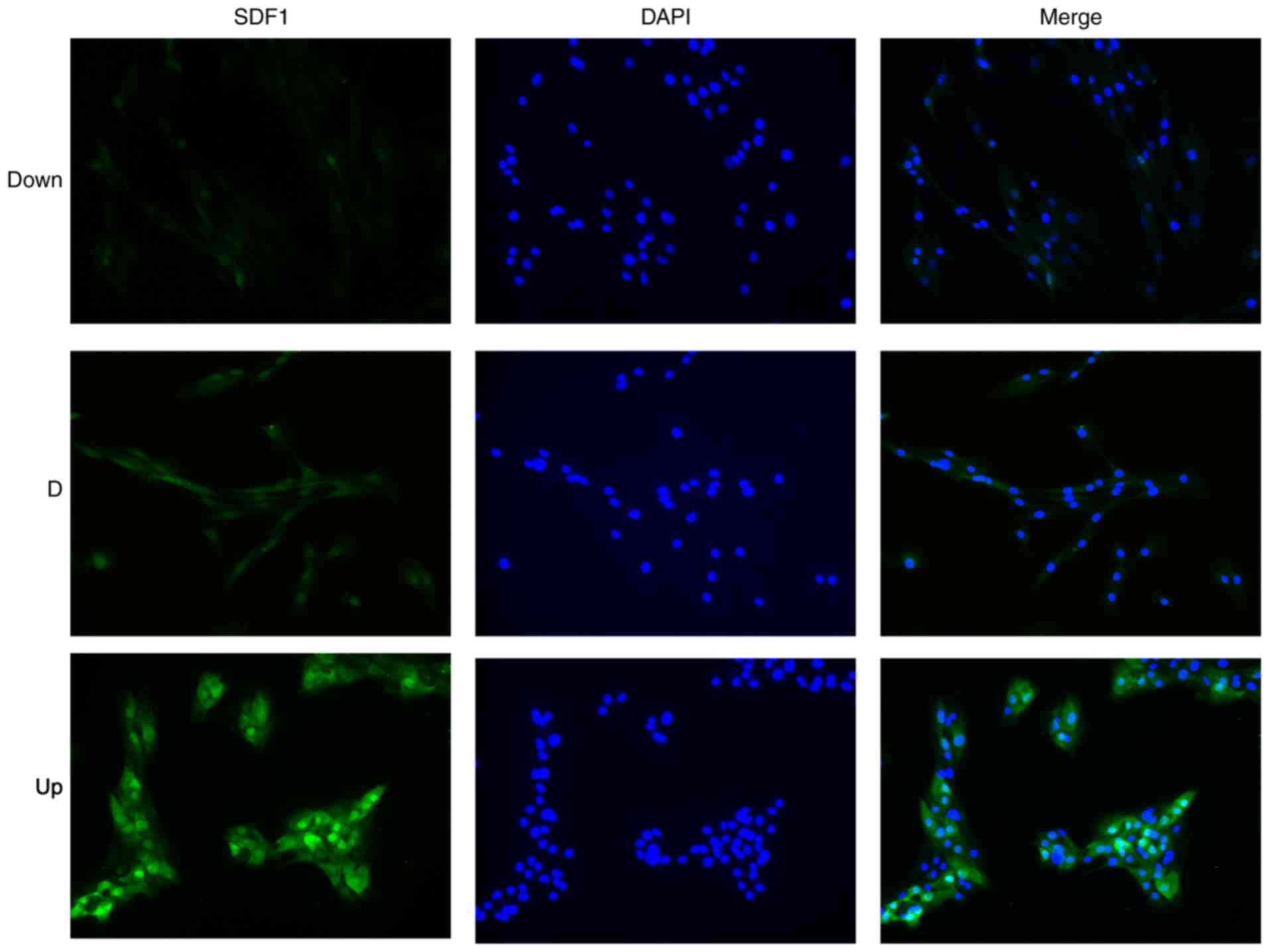
group. Later, using cellular immunofluorescence technology, the
expression of SDF1 (green) in the three groups of NPCs could be
visualized (Fig. 3). The Up group
showed the strongest green fluorescence, the D group showed medium
green fluorescence, and the Down group showed the weakest green
fluorescence. Finally, groups of NPCs with different expression
levels of SDF1 were successfully obtained for subsequent
experiments.
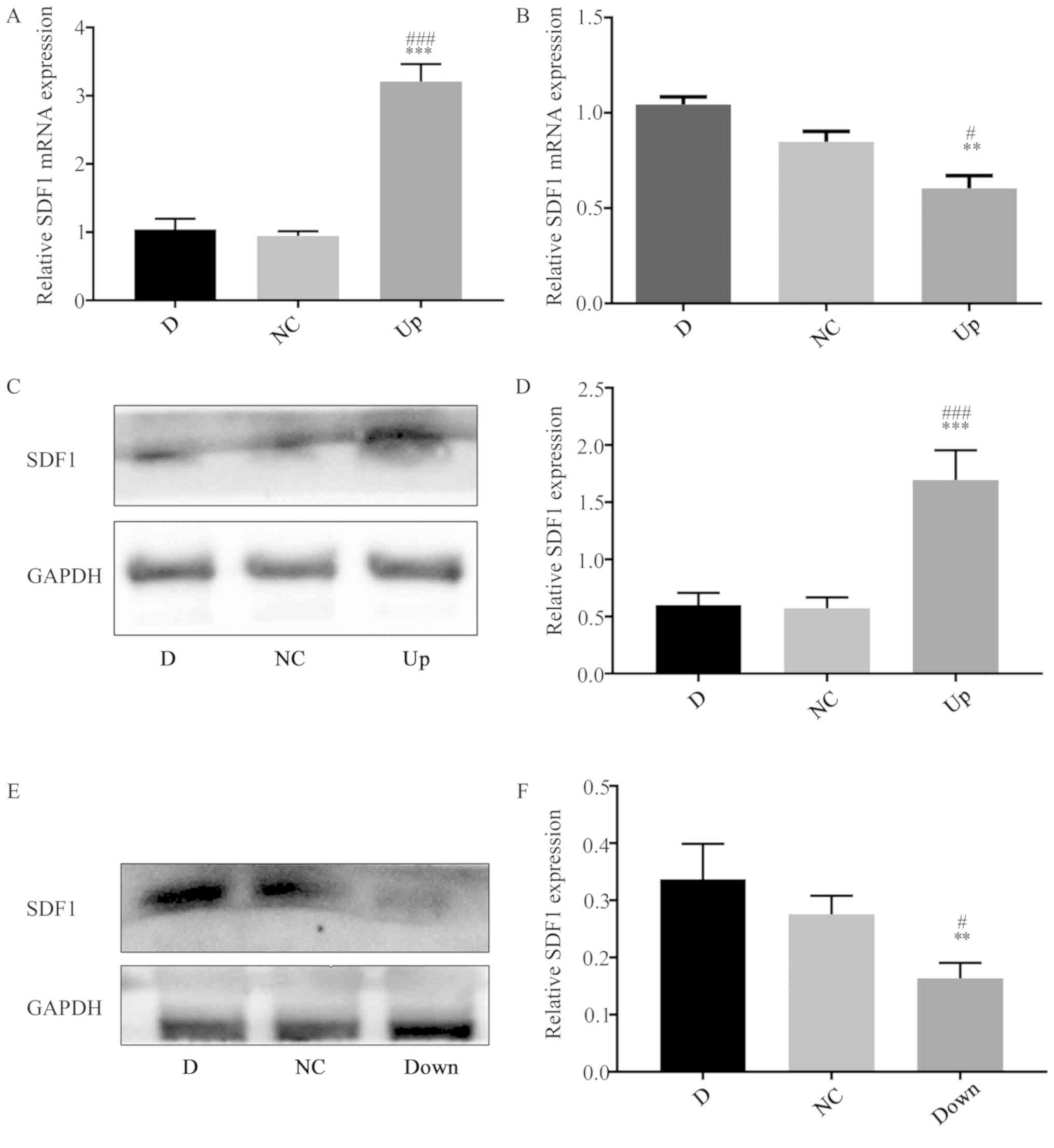 | Figure 2.Upregulation and downregulation of
SDF1 expression in nucleus pulposus cells. (A) RT-qPCR showing that
the mRNA expression level of SDF1 in the Up group was significantly
upregulated compared with the D and NC groups. (B) RT-qPCR showing
that the mRNA expression level of SDF1 in the Down group was
significantly downregulated compared with the D and NC groups. (C)
Western blot analysis showing the SDF1 expression level in the D,
NC and Up groups. (D) The statistical results of (C), showing that
SDF1 expression in the Up group was significantly increased
compared with the D and NC groups. (E) Western blot analysis
showing the SDF1 expression levels in the D, NC and Down groups.
(F) The statistical results of (E), showing that SDF1 expression in
the Down group was significantly decreased compared with the D and
NC groups. **P<0.01, ***P<0.001 vs. respective D group.
#P<0.05, ###P<0.001 vs. respective NC
group. D, degeneration; NC, negative control; Up, upregulation;
Down, downregulation; RT-qPCR, reverse transcription quantitative
PCR; SDF1, stromal cell derived factor 1. |
Effect of NPCs on VECs in a co-culture
system
To detect the effect of NPCs (or the CM of NPCs)
with different expression levels of SDF1 on VECs, a co-culture
system was employed. Later, VECs were pre-incubated with AMD3100 to
detect the effect of CXCR4 on the angiogenesis of VECs. The cell
viability of VECs among the three groups was significantly enhanced
as the expression of SDF1 increased; after pre-incubation with
AMD3100, the viability of VECs in the D and Up groups was
significantly decreased (Fig. 4).
The migration ability was monitored by counting the number of
transmembrane VECs, and the analysis was conducted using ImageJ
software. As the expression of SDF1 increased, the number of
transmembrane VECs significantly increased among the three groups;
after preincubation with AMD3100, the number of transmembrane VECs
in the D and Up groups significantly decreased (Fig. 5). The tube formation ability of
VECs was monitored by measuring the total branching length, and
measurements were conducted using ImageJ software. As the
expression of SDF1 increased, the total branching length
significantly increased among the three groups; after
pre-incubation with AMD3100, the total branching length in the D
and Up groups significantly decreased (Fig. 6). It was also noted that the cell
viability, migration and tube formation ability of VECs in the Down
group decreased before and after incubation with AMD3100, but there
was no significant difference.
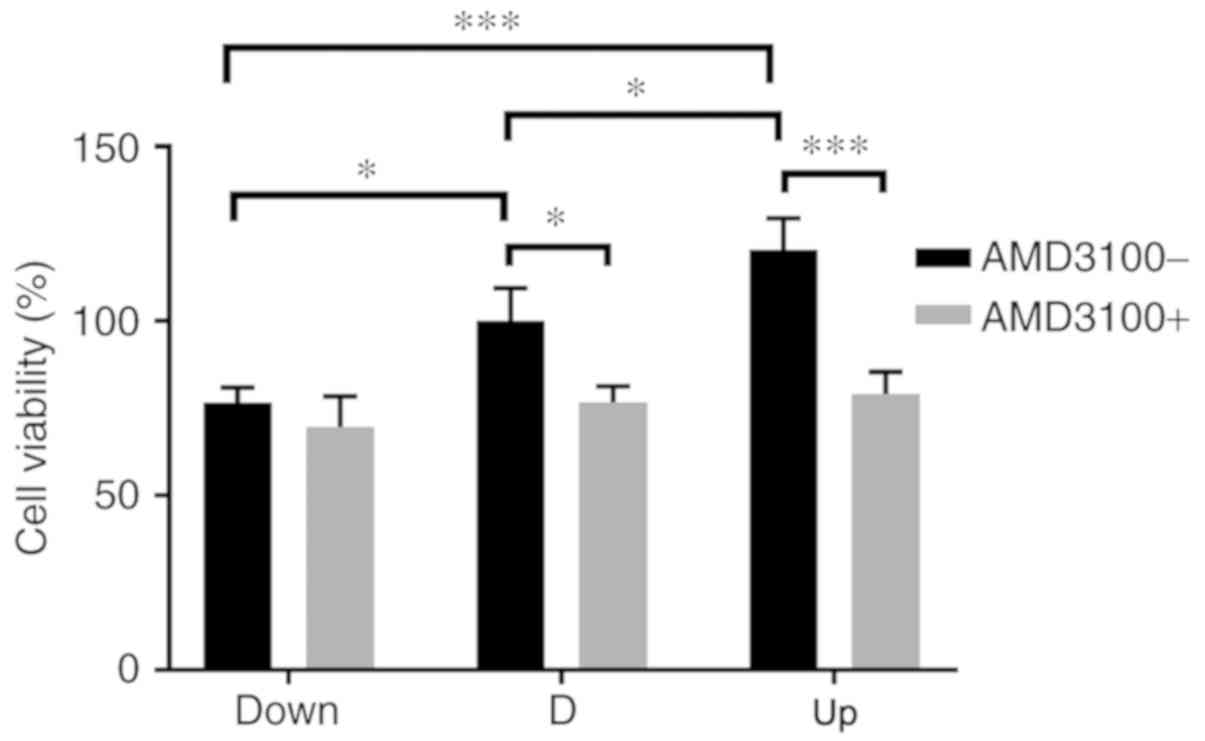 | Figure 4.Influence of nucleus pulposus cells on
the cell viability of VECs in the co-culture system. Before adding
AMD3100, the cell viability of VECs increased as stromal cell
derived factor 1 increased in the Down, D and Up groups, and there
were significant differences among the groups. After adding
AMD3100, the cell viability in the D and Up groups decreased
significantly. There was no significant difference in the Down
group before or after adding AMD3100. *P<0.05, ***P<0.001.
VECs, vascular endothelial cells; Down, downregulation; D,
degeneration; Up, upregulation; AMD3100-, without adding AMD3100
into the co-culture system; AMD3100+, adding AMD3100 into the
co-culture system. |
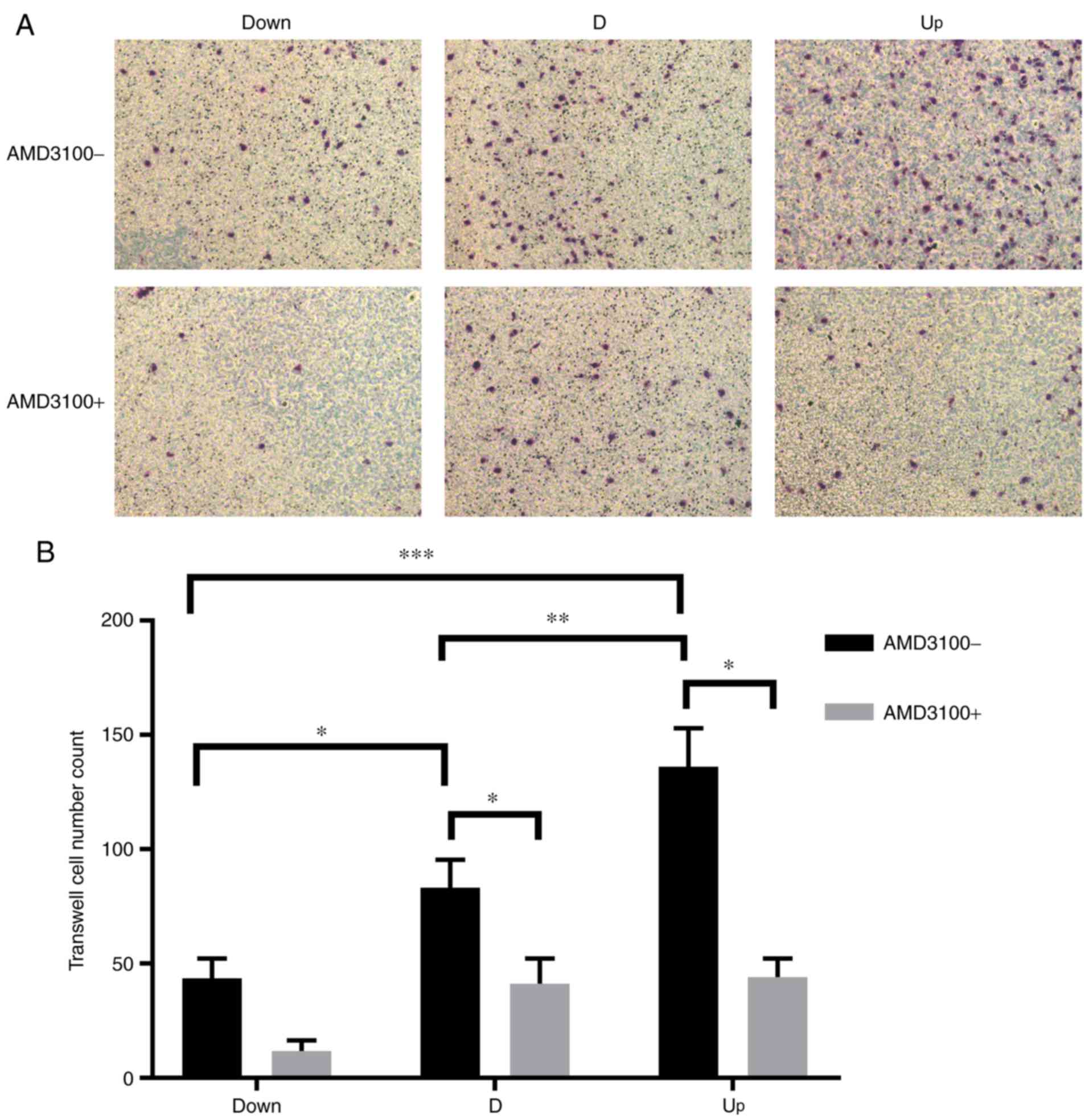 | Figure 5.Influence of nucleus pulposus cells on
the migration of VECs in the co-culture system. (A) In the
co-culture system, before adding AMD3100, the number of
transmembrane VECs increased as stromal cell derived factor
increased in the Down, D and Up groups. After adding AMD3100, the
number of transmembrane VECs decreased in all three groups.
Magnification, ×200. (B) Statistical results, showing that there
was a significant difference in the number of transmembrane VECs
between each group before adding AMD3100. After adding AMD3100, the
number of transmembrane VECs in the D and Up groups decreased
significantly. There was no significant difference in the Down
group before or after adding AMD3100. *P<0.05, **P<0.01,
***P<0.001. VECs, vascular endothelial cells; Down,
downregulation; D, degeneration; Up, upregulation; AMD3100-,
without adding AMD3100 into the co-culture system; AMD3100+, adding
AMD3100 into the co-culture system. |
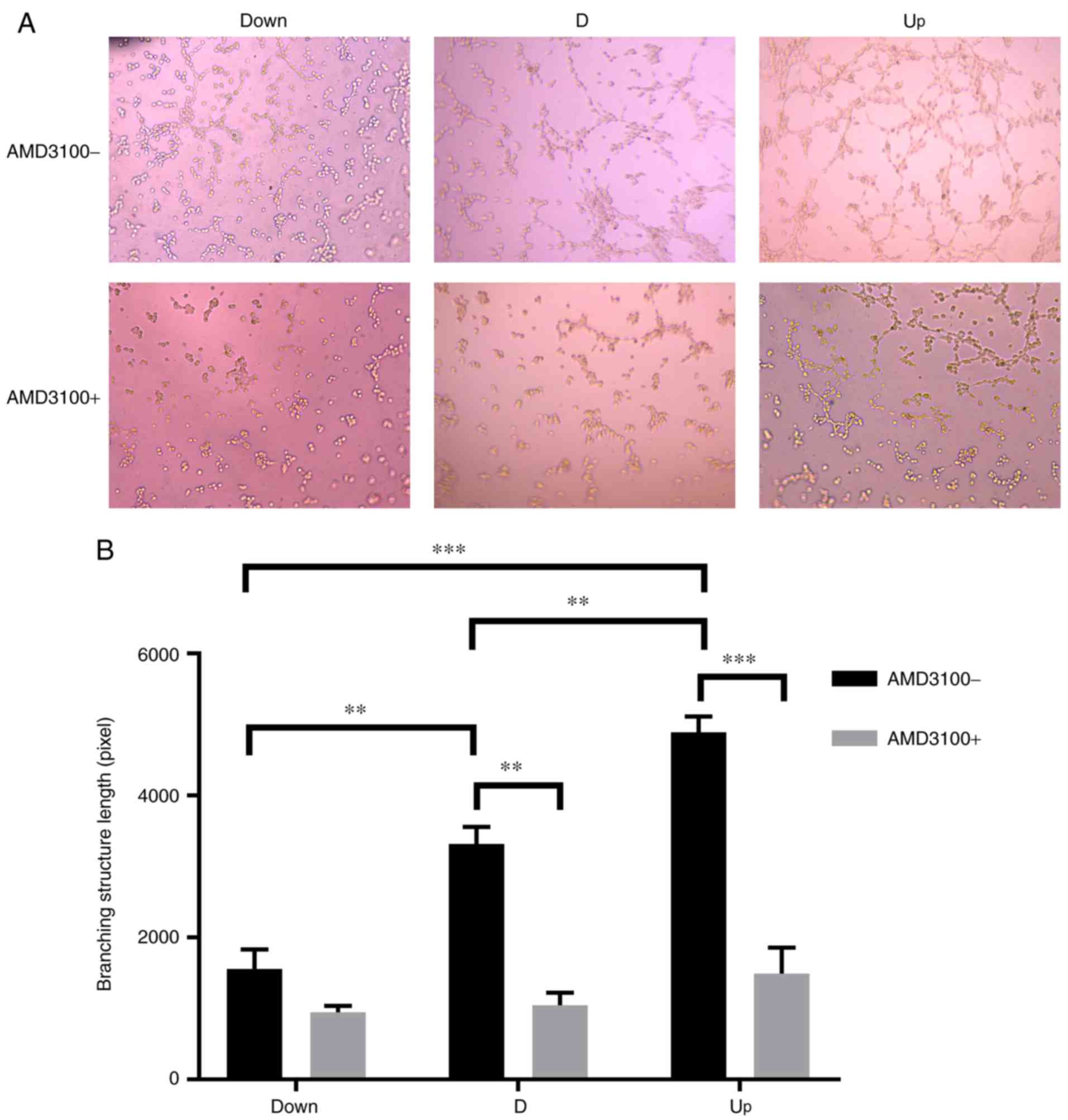 | Figure 6.Influence of nucleus pulposus cells on
the tube formation of vascular endothelial cells in the co-culture
system. (A) In the co-culture system, before adding AMD3100, the
branching structures were increased as stromal cell derived factor
1 increased in the Down, D and Up groups. After adding AMD3100, the
branching structures were decreased in all three groups.
Magnification, ×200. (B) Statistical results, showing that there
was a branching structure length difference between each group
before adding AMD3100. After adding AMD3100, the branching
structure length in the D and Up groups decreased significantly.
There was no significant difference in the Down group before or
after adding AMD3100. **P<0.01, ***P<0.001. Down,
downregulation; D, degeneration; Up, upregulation; AMD3100-,
without adding AMD3100 into the co-culture system; AMD3100+, adding
AMD3100 into the co-culture system. |
Discussion
Chemokines are a class of polypeptides consisting of
~100 amino acids that act as cytokines leading to chemotaxis, and
SDF1 is an important example of these proteins. Many types of cells
express SDF1, such as cardiomyocytes, endothelial cells, smooth
muscle cells, macrophages and progenitor cells (23,24),
and it is involved in an array of physiological and pathological
activities in vivo. In the field of orthopedics, a previous
report demonstrated that the SDF1/CXCR4 axis participated in the
regulation of changes in matrix components and cartilage
degeneration during osteoarthritis (25). Intervertebral discs also belong to
the motor system, the same as articular cartilage, enduring high
stress and the lack of a blood supply. Degenerative disc disease
and osteoarthritis are both degenerative diseases of the motor
system. Therefore, it was speculated that the SDF1/CXCR4 axis may
participate in the disc degeneration process.
Zhang et al (26) reported that SDF1 and CXCR4
expression were upregulated in degenerated discs, as detected by
immunohistochemistry. Further research from Liu et al
(27) revealed that SDF1/CXCR4 was
not only increased in degenerated intervertebral discs, but also
resulted in disc degeneration by inducing apoptosis in NPCs. SDF1
is a secretory protein and is secreted into the extracellular
matrix after its production by cells. Previous reports (26,27)
used immunohistochemical methods to illustrate the expression of
SDF1, and showed that there are large areas of positive staining in
the extracellular matrix of degenerated discs, but whether SDF1 is
expressed by NPCs was not clear. In the present experiments, the
NPCs of degenerated intervertebral discs were identified as the
source of SDF1 using RT-qPCR, western blotting and
immunofluorescence analysis. It was therefore concluded that the
degenerated NPCs can secret SDF1 to play a physiological and
pathological role.
In this study, the expression of SDF1 in NPCs was
upregulated and downregulated by virus transfection, and the
expression was successfully confirmed at the mRNA and protein
level. Then, using a conditioned medium produced by NPCs or by
direct co-culture, the interaction between NPCs and VECs was
observed. The results showed that the proliferation, migration and
tube formation of VECs were enhanced with the increase in SDF1
expression. The intervertebral disc is an organ that lacks blood
vessels, and only the one-third of the outer layer of the annulus
has blood vessels; this is a result of the balance between
anti-vascular ingrowth factors and vascular ingrowth factors. Under
normal conditions, the aggrecan component contained in the nucleus
pulposus has an inhibitory effect on vascular ingrowth (28). In the case of discogenic LBP, it
can be seen that the new blood vessels grow from the annulus
fibrosus into the nucleus pulposus, which is accompanied by the
occurrence of nucleus pulposus fibrosis (29). This indicates that when
intervertebral disc degeneration occurs, the aggrecan component in
the nucleus pulposus is reduced, which works as a factor of
anti-vascular ingrowth. To the contrary, the factors that promote
vascular ingrowth are increased. The comprehensive result is that
neovascularization occurs into the degenerative intervertebral
disc, even deep into the nucleus pulposus, causing painful
intervertebral disc degeneration. As a factor of angiogenesis, SDF1
has been reported in many physiological and pathological processes.
Cai et al (30) reported
that SDF1, which is enhanced during hyperglycemia, can promote
choroidal vascularization. Yu et al (31) reported that SDF1, which was highly
expressed in renal cells, promoted the neovascularization of
tumors. Virgintino et al (32) reported that SDF1 was involved in
the regulation of angiogenesis during human brain development. The
present results showed that with the increase in SDF1 expression in
NPCs, the proliferation and chemotactic migration ability of VECs
were enhanced. The ability of tube-like structure formation was
also enhanced, which is involved in the synthesis of a series of
proteins, collagen and proteases in VECs (33). These results suggested that SDF1
also plays an important role in the process of vascularization in
degenerated intervertebral discs. Furthermore, the degree of
vascularization in the intervertebral disc is positively correlated
with the degree of degeneration (10).
CXCR4 is a specific receptor for SDF1 and is
expressed in a variety of cells, including VECs. SDF1 and CXCR4
constitute a signaling axis and participate in the pathological
process of various diseases, such as HIV infection, tumor
metastasis, pathological angiogenesis, myocardial ischemia and
pulmonary fibrosis (34). CXCR4
not only promotes angiogenesis in normal tissues, such as the
cornea and retina, but also promotes angiogenesis under
pathological conditions such as tumors. In this experiment, the
interaction of each group of NPCs (or the CM of NPCs) and VECs
exhibited properties of facilitating angiogenesis. However, after
pre-incubation of the VECs with AMD3100, which is a small molecule
inhibitor of CXCR4, the effects of each group of NPCs (or the CM of
NPCs), not only on the endothelial cell proliferation and
chemotactic migration but also on the tube-like structure
formation, were blocked and declined to varying degrees. These
results suggested that SDF1 passes signals through CXCR4 on the
endothelial cell surface, and the SDF1/CXCR4 axis plays an
important role in promoting the angiogenesis of VECs induced by
NPCs. At the same time, according to a previous study (35), it can be speculated that in
degenerated intervertebral discs, because of the large amount of
inflammatory substances such as IL-1β (36), the expression of CXCR4 in VECs will
be further upregulated, and the response to SDF1 will be more
intense, showing stronger chemotaxis. The macroscopic manifestation
is that in degenerated intervertebral discs accompanied by a large
number of inflammatory mediators, more neovascularization will lead
to ingrowth into the nucleus pulposus.
The present results also showed that in the SDF1
downregulation group (Down), there was a further decrease in the
cell viability, migration ability and tube formation ability before
and after incubation with AMD3100, but there were no significant
differences. It was considered that the decreased expression of
SDF1 greatly affected the angiogenesis of VECs. Therefore, further
inhibition of CXCR4 did not further inhibit angiogenesis. On the
other hand, these results also indicated that the effect of SDF1 on
angiogenesis may be as important as that of CXCR4.
The shortcoming of these experiments is that the
NPCs and VECs were in a monolayer culture with a sufficient oxygen
supply. Although the P1 generation of NPCs were chosen for this
experiment to keep the cell phenotype as close to the original
generation as possible, there were still differences in regard to
the high pressure, low oxygen and acidic environment observed in
vivo. Therefore, the addition of continuous pressure (or a
regularly changing pressure to simulate the change of pressure
in vivo) under hypoxic culture conditions may be closer to
the in vivo situation, and will provide more in-depth and
objective data in exploring the angiogenesis of degenerated
intervertebral discs.
The present study upregulated and downregulated the
expression of SDF1 in degenerated NPCs by virus transfection and
identified the main source of SDF1 in nucleus pulposus tissue. At
the same time, the NPCs (or CM of NPCs) with different SDF1
expression levels were co-cultured with VECs, and the results
suggested that the SDF1/CXCR4 axis plays a role in new blood vessel
ingrowth into degenerated intervertebral discs. As the expression
of SDF1/CXCR4 increases, the effect on angiogenesis is also
increased. This will help to further understand the process of
intervertebral disc degeneration, and provide new ideas for
exploring the pathogenesis and treatment of disc degeneration.
Acknowledgements
The authors thank the Department of Orthopedics at
The First Affiliated Hospital of Chongqing Medical University for
their collaboration and support. The authors express special
gratitude to the Chongqing Key Laboratory of Ophthalmology for
providing the experimental platform.
Funding
The present study was supported by the Guizhou
Science and Technology Department Cooperation Project (2015) (grant
no. 2152).
Availability of data and materials
The datasets used and/or analyzed during the current
study are included in this published article.
Authors' contributions
HZ participated in the study design, data
collection, statistical analysis, manuscript preparation and
literature search. BH participated in the study design, data
collection, statistical analysis and collection of funds.
Ethics approval and consent to
participate
Informed consent was obtained from all donors, and
the experimental protocol was approved by the Ethics Committee of
Chongqing Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Andersson GB: Epidemiological features of
chronic low-back pain. Lancet. 354:581–585. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Martin BI, Turner JA, Mirza SK, Lee MJ,
Comstock BA and Deyo RA: Trends in health care expenditures,
utilization, and health status among US adults with spine problems,
1997–2006. Spine (Phila Pa 1976). 34:2077–2084. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dieleman JL, Baral R, Birger M, Bui AL,
Bulchis A, Chapin A, Hamavid H, Horst C, Johnson EK, Joseph J, et
al: US spending on personal health care and public health,
1996–2013. JAMA. 316:2627–2646. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
O'Halloran DM and Pandit AS:
Tissue-engineering approach to regenerating the intervertebral
disc. Tissue Eng. 13:1927–1954. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Urban JP, Smith S and Fairbank JC:
Nutrition of the intervertebral disc. Spine (Phila Pa 1976).
29:2700–2709. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Matveeva N, Zivadinovik J, Zdravkovska M,
Jovevska S and Bojadzieva B: Histological composition of lumbar
disc herniations related to the type of herniation and to the age.
Bratisl Lek Listy. 113:712–717. 2012.PubMed/NCBI
|
|
7
|
Ahmed M, Bjurholm A, Kreicbergs A and
Schuftzberg A: Neuropeptide Y, tyrosine hydroxylase and vasoactive
intestinal polypeptide-immunoreactive nerve fibers in the vertebral
bodies, discs, dura mater and spinal ligaments of the rat lumbar
spine. Spine (Phila Pa 1976). 18:268–273. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stefanakis M, Al-Abbasi M, Harding I,
Pollintine P, Dolan P, Tarlton J and Adams MA: Annulus fissures are
mechanically and chemically conducive to the ingrowth of nerves and
blood vessels. Spine (Phila Pa 1976). 37:1883–1891. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Freemont AJ, Watkins A, Le Maitre C, Baird
P, Jeziorska M, Knight MT, Ross ER, O'Brien JP and Hoyland JA:
Nerve growth factor expression and innervation of the painful
intervertebral disc. J Pathol. 197:286–292. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rätsep T, Minajeva A and Asser T:
Relationship between neovascularization and degenerative changes in
herniated lumbar intervertebral discs. Eur Spine J. 22:2474–2480.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nagasawa T, Kikutani H and Kishimoto T:
Molecular cloning and structure of a pre-B-cell growth-stimulating
factor. Proc Natl Acad Sci USA. 91:2305–2309. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Loetscher M, Geiser T, O'Reilly T, Zwahlen
R, Baggiolini M and Moser B: Cloning of a human seven-transmembrane
domain receptor, LESTR, that is highly expressed in leukocytes. J
Biol Chem. 269:232–237. 1994.PubMed/NCBI
|
|
13
|
Zou YR, Kottmann AH, Kuroda M, Taniuchi I
and Littman DR: Function of the chemokine receptor CXCR4 in
haematopoiesis and in cerebellar development. Nature. 393:595–599.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moser B and Loetscher P: Lymphocyte
traffic control by chemokines. Nat Immunol. 2:123–128. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Katsumoto K and Kume S: The role of
CXCL12-CXCR4 signaling pathway in pancreatic development.
Theranostics. 3:11–17. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sainz J and Sata M: CXCR4, a key modulator
of vascular progenitor cells. Arterioscler Thromb Vasc Biol.
27:263–265. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sasaki T, Fukazawa R, Ogawa S, Kanno S,
Nitta T, Ochi M and Shimizu K: Stromal cell-derived factor-1alpha
improves infarcted heart function through angiogenesis in mice.
Pediatr Int. 49:966–971. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shyu WC, Lin SZ, Yen PS, Su CY, Chen DC,
Wang HJ and Li H: Stromal cell-derived factor-1 alpha promotes
neuroprotection, angiogenesis, and mobilization/homing of bone
marrow-derived cells in stroke rats. J Pharmacol Exp Ther.
324:834–849. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pfirrmann CW, Metzdorf A, Zanetti M,
Hodler J and Boos N: Magnetic resonance classification of lumbar
intervertebral disc degeneration. Spine (Phila Pa 1976).
26:1873–1878. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kwon WK, Moon HJ, Kwon TH, Park YK and Kim
JH: Influence of rabbit notochordal cells on symptomatic
intervertebral disc degeneration: Anti-angiogenic capacity on human
endothelial cell proliferation under hypoxia. Osteoarthritis
Cartilage. 25:1738–1746. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vernon-Roberts B, Moore RJ and Fraser RD:
The natural history of age-related disc degeneration: The pathology
and sequelae of tears. Spine (Phila Pa 1976). 32:2797–2804. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jaerve A, Schira J and Müller HW: Concise
review: The potential of stromal cell-derived factor 1 and its
receptors to promote stem cell functions in spinal cord repair.
Stem Cells Transl Med. 1:732–739. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dong F, Harvey J, Finan A, Weber K,
Agarwal U and Penn MS: Myocardial CXCR4 expression is required for
mesenchymal stem cell mediated repair following acute myocardial
infarction. Circulation. 126:314–324. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu W, Shi J, Zhang J, Lv Z, Guo F, Huang
H, Zhu W and Chen A: CXCL12/CXCR4 axis regulates aggrecanase
activation and cartilage degradation in a post-traumatic
osteoarthritis rat model. Int J Mol Sci. 17(pii): E15222016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang H, Zhang L, Chen L, Li W, Li F and
Chen Q: Stromal cell-derived factor-1 and its receptor CXCR4 are
upregulated expression in degenerated intervertebral discs. Int J
Med Sci. 11:240–245. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu Z, Ma C, Shen J, Wang D, Hao J and Hu
Z: SDF-1/CXCR4 axis induces apoptosis of human degenerative nucleus
pulposus cells via the NF-κB pathway. Mol Med Rep. 14:783–789.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Johnson WE, Caterson B, Eisenstein SM and
Roberts S: Human intervertebral disc aggrecan inhibits endothelial
cell adhesion and cell migration in vitro. Spine (Phila Pa 1976).
30:1139–1147. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peng B, Chen J, Kuang Z, Li D, Pang X and
Zhang X: Expression and role of connective tissue growth factor in
painful disc fibrosis and degeneration. Spine (Phila Pa 1976).
34:E178–E182. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cai Y, Li X, Wang YS, Shi YY, Ye Z, Yang
GD, Dou GR, Hou HY, Yang N, Cao XR and Lu ZF: Hyperglycemia
promotes vasculogenesis in choroidal neovascularization in diabetic
mice by stimulating VEGF and SDF-1 expression in retinal pigment
epithelial cells. Exp Eye Res. 123:87–96. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yu P, Ge YZ, Zhao Y, Wu JP, Wu R, Zhou LH
and Jia RP: Identification and significance of mobilized
endothelial progenitor cells in tumor neovascularization of renal
cell carcinoma. Tumour Biol. 35:9331–9341. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Virgintino D, Errede M, Rizzi M, Girolamo
F, Strippoli M, Wälchli T, Robertson D, Frei K and Roncali L: The
CXCL12/CXCR4/CXCR7 ligand-receptor system regulates
neuro-glio-vascular interactions and vessel growth during human
brain development. J Inherit Metab Dis. 36:455–566. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Arnaoutova I and Kleinman HK: In vitro
angiogenesis: Endothelial cell tube formation on gelled basement
membrane extract. Nat Protoc. 5:628–635. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Debnath B, Xu SL, Grande F, Garofalo A and
Neamati N: Small molecule inhibitors of CXCR4. Theranostics.
3:47–75. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gupta SK, Lysko PG, Pillarisetti K,
Ohlstein E and Stadel JM: Chemokine receptors in human endothelial
cells. Functional expression of CXCR4 and its transcriptional
regulation by inflammatory cytokines. J Biol Chem. 273:4282–4287.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang W, Yu XH, Wang C, He WS, Zhang SJ,
Yan YG, Zhang J, Xiang YX and Wang WJ: Interleukin-1β in
intervertebral disk degeneration. Clin Chim Acta. 450:262–272.
2015. View Article : Google Scholar : PubMed/NCBI
|