Introduction
Structural abnormalities with breakpoints in the
euchromatic section of a Y chromosome may lead to its instability
and loss in subsequent cell cycles, with formation of a mosaic 45,X
cell line. As a result, numerous morphologic forms of the aberrant
Y chromosome occur in the mosaic karyotype. Dicentrics are the most
common structural abnormalities of the Y chromosome (1,2).
They are mitotically unstable and may consequently be lost during
development, resulting in the majority of patients with dicentric Y
chromosome having a mosaic karyotype, mostly with a 45,X cell line
(3). Clinical features of such
patients may range from Turner-syndrome-like females to infertile
males, depending on the structure of the dicentric Y chromosome,
the sites of breakage and types of mosaicism.
The identification of Y chromosome-derived material
in cases with dysgenetic gonads is associated with an increased
risk of developing gonadoblastoma. The presence of a Y chromosome,
or part of it, is associated with a 25–70% risk of gonadoblastoma,
the highest risk being associated with mixed gonadal dysgenesis and
increasing age (4). In patients
with the 45,X/46,XY karyotype, the risk appears lowest in
phenotypic males, intermediate (0.5%) in patients with ambiguous
external genitalia and highest (22%) in phenotypic females
(1).
The Y chromosome contains crucial loci for normal
male sexual development. The sex-determining region of the Y
chromosome (SRY), a transcription factor on the short arm of the Y
chromosome, is the critical switch leading to testis development.
Mutations in SRY result in XY individuals developing as females;
patients with a 45,X karyotype with insertion of SRY into an
autosome have a male phenotype (5,6).
Genes in the euchromatic region of the long arm of the Y chromosome
(Yq11) are required for normal spermatogenesis, and Y chromosome
microdeletions are an important genetic cause of primary
testiculopathies. Deletion mapping was directed at the
identification of genes related to spermatogenesis, and defined
three regions as the azoospermia factors (AZFa, AZFb and AZFc)
mapped to Yq11 (7). The main
candidate gene in AZFc is the DAZ (deleted in azoospermia) cluster,
a set of genes transcribed in the testis and expressed exclusively
in germ cells, apparently encoding an RNA binding protein (8).
In this study, we present a detailed
molecular-cytogenetic characterization of a patient with phenotypic
features of the Klinefelter syndrome with mosaicism involving an
isodicentric Y chromosome.
Patient and methods
Patient history
A 17-year-old male was referred for cytogenetic
analysis in October 2008 due to a small penis with small testes
(the right and left testicular volume of 4 and 4.5 ml,
respectively). Informed consent was obtained from the patient and
ethics approval for the study was provided by the Bio-safety and
Bioethics Member Committee/S.A.E.C. The patient's weight was 69 kg,
with a height of 170 cm, and he did not show any dysmorphic signs
or symptoms. The mother was a 37-year-old healthy female without
any signs or symptoms of virilization or drug abuse during
pregnancy. The mother's and the father's height were 167 and 184
cm, respectively. No family history of sterility or abortion was
reported. Ultrasound of the pelvis area revealed no uterus and
ovaries. Laboratory examination showed FSH levels of 2.1 mIU/ml, LH
levels of 1.6 mIU/ml and a testosterone concentration of 0.89
ng/dl. Volume semen analysis was 2.5 ml.
Cytogenetic and molecular cytogenetics
analysis
Cytogenetic analysis using GTG-banding was performed
according to standard procedures (9). A total of 100 metaphases analyzed
from stimulated peripheral blood cultures were analyzed. Karyotypes
were described according to the International System for Human
Cytogenetic Nomenclature (10).
Fluorescence in situ hybridization (FISH) was
carried out on the metaphases using a commercially available probes
whole painting chromosome (WCP), LSI SRY, subtelomeric for Xp/Yp
and Xq/Yq, centromeric for X and Y (DXZ1; DYZ3) (Abbott
Molecular/Vysis, Des Plaines, IL, USA) and Y chromosome-specific
multicolor banding (MCB). The procedure was performed as described
in a previous study (11). A total
of 50–100 metaphase spreads were analyzed. The results were
evaluated on a fluorescence microscope (AxioImager.Z1 mot, Zeiss)
equipped with appropriate filter sets to discriminate between a
maximum of five fluorochromes and the counterstain DAPI (4′,
6′-diamino-2-phenylindole). Image capturing and processing were
carried out using an Isis imaging system (MetaSystems, Altlussheim,
Germany) for the evaluation of MCB.
Molecular analysis
Genomic DNA was extracted from peripheral blood
using Genequality AZF MX kit (AB Analitica, Italy) according to the
manufacturer's instructions. To screen for Y-specific regions, we
tested for the presence of 26 Y-DNA loci via a sequence-tagged site
(STS) and genes. The STS primers tested were: sY81, sY82, sY84,
sY86, DBY, USP9Y (AZFa); sY95; sY117, sY121, sY124, sY125, sY127,
sY134, sY142, sY143 (AZFb); sY147, sY149, sY158, sY239, sY242,
sY254 (DAZ), sY255 (DAZ), sY283 (AZFc); sY145, sY153 (AZFd).
Moreover, sY160 (heterochromatin region) and human zinc-finger
protein-encoding genes (ZFX/ZFY) located on the X and Y chromosomes
were selected. The ZFX/ZFY and SRY (i.e., STS sY14) acted as
internal control primers. Following an initial denaturation step at
94°C for 5 min, cycle parameters were: 40 cycles at 94°C for 1 min,
60°C for 1 min, and 72°C for 1 min. The program was followed by the
final extension step at 72°C for 5 min. The reaction products were
electrophoresed on an ethidium bromide-stained 2.5% agarose-TAE-gel
and observed under UV-light.
PCR amplification of SRY and sequencing
analysis
According to the procedure of Al-Achkar et al
(12), mutation screening was
carried out using direct DNA sequence analysis. The whole coding
sequence of the SRY gene was amplified by PCR using the primers
previously described (13).
Results
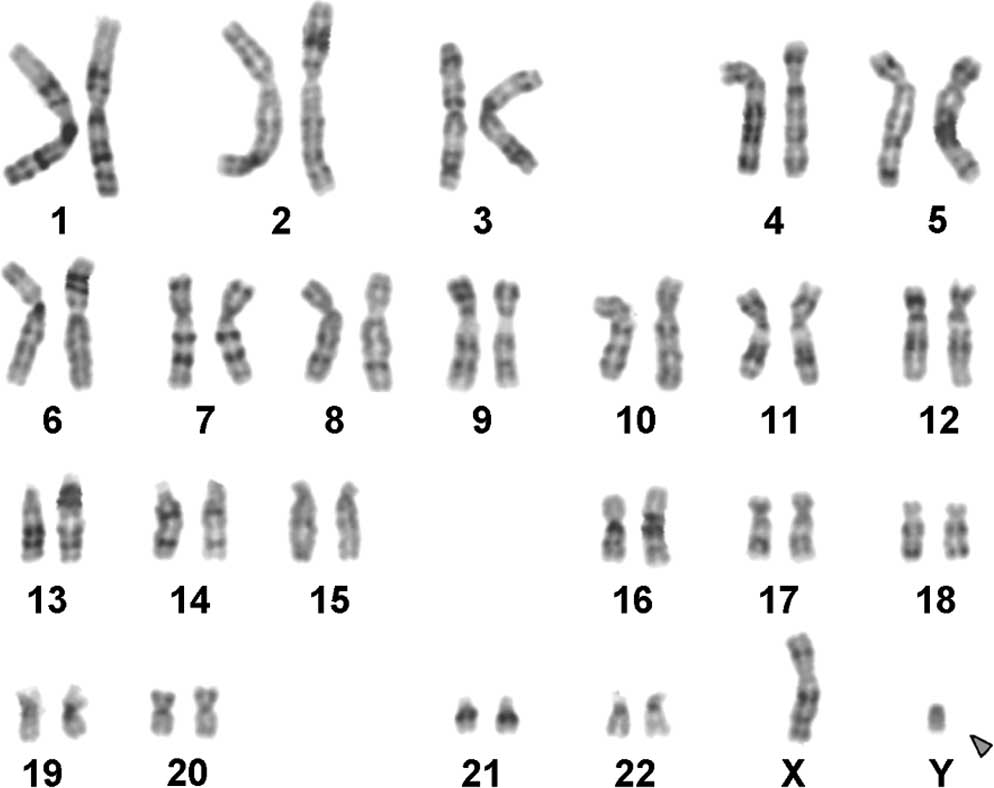
The karyotype determined by GTG-banding was
identified as mos 45,X/46,X,+mar/47,XX,+mar (Fig. 1). FISH using a Y
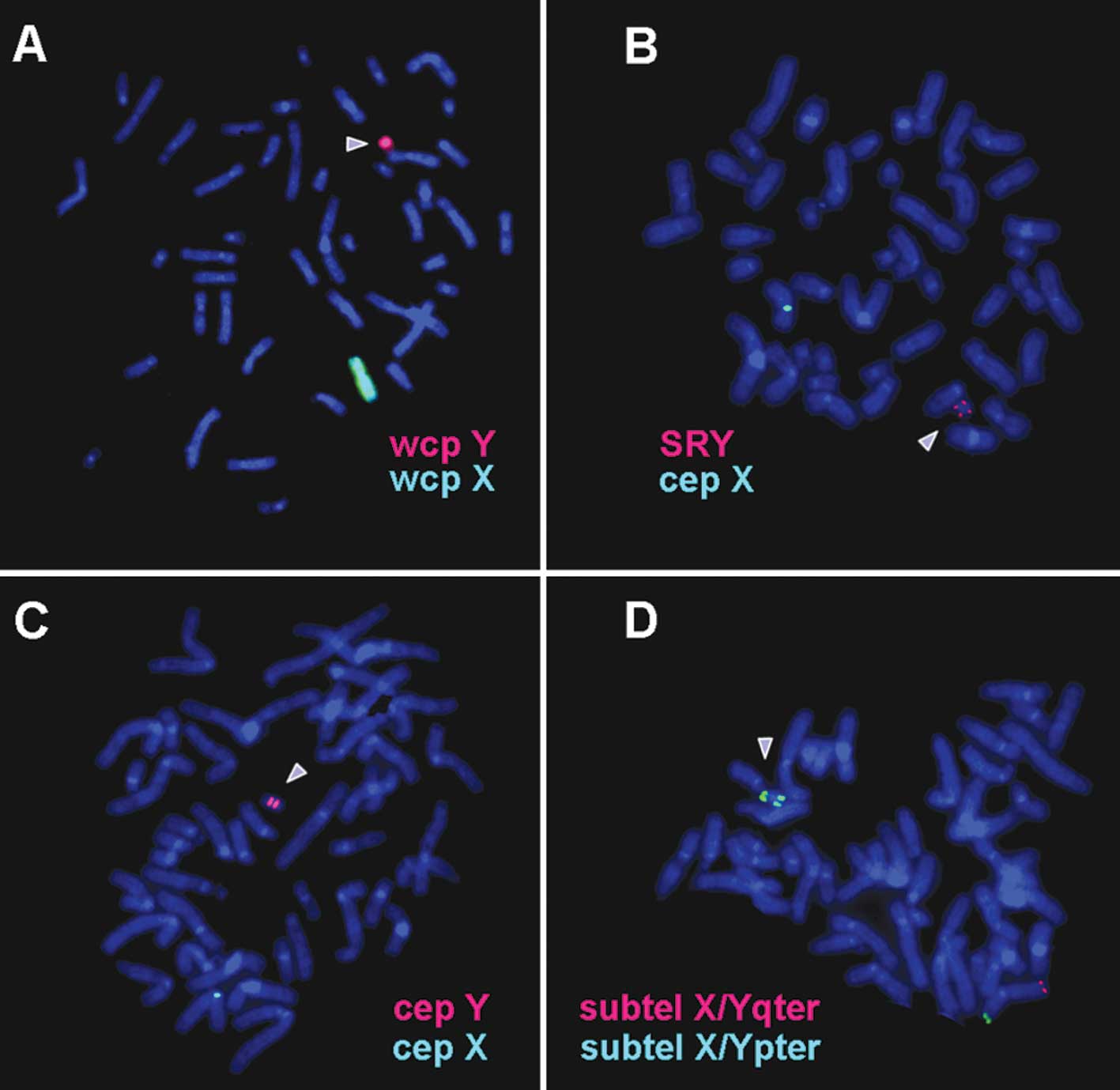
chromosome-specific whole chromosome painting probe stained the
entire derivative chromosome (Fig.
2A). Findings of dual color FISH experiments (Fig. 2B-D) showed that the chromosome was
an isodicentric mainly derived from the short arm of the Y
chromosome. Together with the MCB-findings (Fig. 3) the karyotype was obtained was:
45,X[4]/46,X,idic(Y)(q11.21)[95]/47,XX,+idic(Y)(q11.21)[1].
The presence or absence of 26 Y-specific loci was
analyzed molecularly. Of the 26 loci, 12 loci were absent in the
present case: sY95, sY117, sY125, sY127, sY134, sY145, sY147,
sY149, sY158, sY254, sY255 and sY283 (Fig. 4). Thus, the breakpoint of the
derivative Y chromosome was determined to be between marker sY84
(13.4 MB) and sY95 (15.1 MB). According to cytogenetics, the
breakpoint was in band Yq11.21, located between 12.5 and 14.3 MB.
Therefore, the breakpoint is likely to be between 13.4 and 14.3 MB
(hg18).
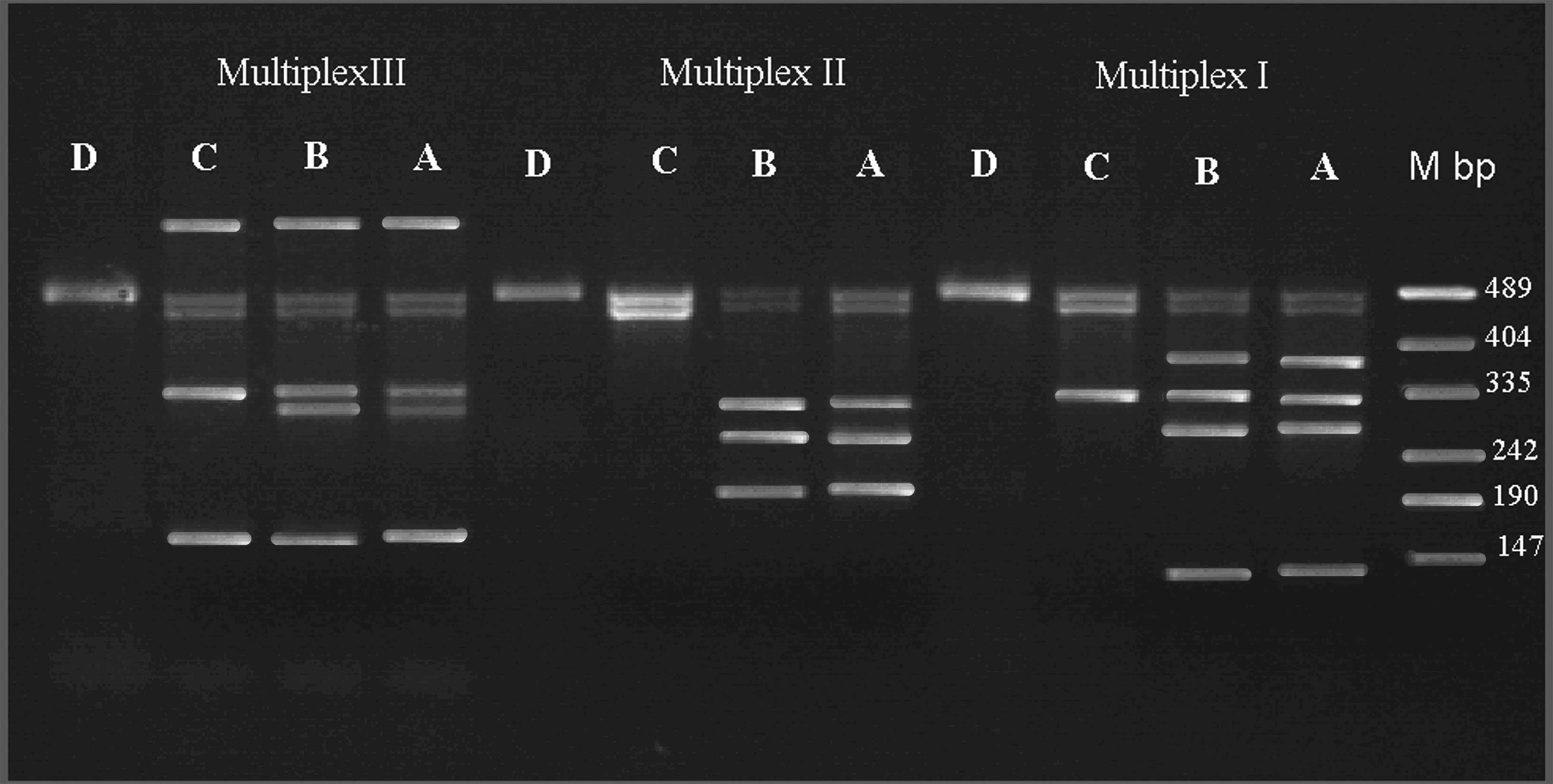 | Figure 4Multiplex PCR for the presence or
absence of 26 Y-specific loci were analyzed in the patient with a Y
chromosome deletion. Multiplex I: Zinc-finger protein-encoding
genes (ZFX/ZFY) (495 bp), sex-determining region of Y chromosome
(SRY) (472 bp), sY254 (380 bp), sY86 (320 bp), sY127 (274 bp) and
sY255 (120 bp); Multiplex II: ZFX/ZFY (495 bp), SRY (472 bp), sY95
(303 bp), sY117 (262 bp) and sY125 (200 bp); and Multiplex III: DBY
(689 bp), ZFX/ZFY (495 bp), SRY (47 bp), sY84 (326 bp), sY134 (301
bp) and DFFRY (155 bp). This patient had seven (sY95, sY117, sY125,
sY127, sY134, sY254 and sY255) deletions. Each reaction was
compared with a fertile man serving as the control (A and B); C,
DNA sample of patient; D, sample from a woman as a negative
control; and M, DNA molecular weight marker. Additionally the loci
(sY145, sY147, sY149, sY158 and sY283) were deleted (data not
shown). |
To identify a possible mutation, we analyzed the
SRY-specific PCR fragment by DNA sequencing using a normal male as
a control. No mutation was exhibited by the patient (data not
shown).
Discussion
The most common abnormality in the human Y
chromosome is a dicentric derivative present as part of a mosaic
karyotype including a 45,X cell line (3). A number of mechanisms of dicentric
isochromosome formation have been described and the most frequent
mechanism of this alteration is the sister-chromatid breakage and
reunion (14–17). Isodicentric Y chromosomes appear to
be formed by a single break in one of the Y chromatids, followed by
a fusion of the broken ends of sister chromatids and the loss of
the acentric fragment during gametogenesis prior to the formation
of spermatids. Such rearrangements are generally unstable and an
additional 45,X cell line is frequently present (18). Patients with dicentric
isochromosome Y are mosaics without a 46,XY cell line, indicating
that gametogenic or early post-zygotic origins are the most
frequent (1), and a 45,X cell line
is usually found as part of this mosaicism at the same time
(1,3). In the present study, the dicentric
isochromosome Y was also mosaic but without a normal cell line
(1,3).
Although in the literature patients with mosaic
karyotypes 45,X/46,X,idic(Y)(q11.21) have frequently been described
(http://www.med.uni-jena.de/fish/sSMC/sturner.html),
the present patient is to the best of our knowledge the first case
with mos 45,X/46,X,idic(Y)(q11.21)/47,XX,+idic(Y)(q11.21) and a
Klinefelter syndrome phenotype.
The Klinefelter syndrome is most common, occurring
in 1:500 to 1:1000 live births (19). The phenotype of patients is
variable such as tall stature, hypogonadism, gynecomastia, small
testes, decreased facial hair, small penis, infertility and low
testosterone (20). The classic
form of Klinefelter syndrome is the polyploidy (extra chromosome)
condition of 47,XXY, which characterizes 80% of Klinefelter cases.
Another 15% of the cases are mosaic types (46,XY/47,XXY,
46,XY/48,XXYY, 45,X0/46,XY/47,XXY and 46XX/47,XXY). The remaining
5% are XX forms with the SRY gene (testes determining factor)
translocated to an X chromosome, Poly X + Y forms, or combined
mosaic and poly X + Y forms (21).
In the present study, the 47,XX,+idic(Y)(q11.21) condition was
present in only 1% of the peripheral blood cells.
The presence of a Y chromosome is associated with a
25–70% risk, the highest risk being associated with mixed gonadal
dysgenesis and increasing age. The development of gonadoblastoma is
thought to be less common in patients with idic(Y)(q11.2) (22).
In conclusion, we present a rare case of a clinical
presentation of Klinefelter syndrome with only 1% of the peripheral
blood cells having the causative two X-chromosomes. We suggest that
the patient has more cells with 47,XX,+idic(Y)(q11.21) karyotype in
other tissues. This case may be useful to explain similar clinical
Klinefelter syndrome cases without typical karyotypic findings in
peripheral blood. The breakpoint of the derivative Y chromosome was
also narrowed down to 0.9 MB.
Acknowledgements
The authors thank Professor I. Othman, General
Director of Atomic Energy Commission of SYRIA (AECS) and Dr N.
Mirali, Head of the Department of Molecular Biology and
Biotechnology for their support. This study was supported by the
Syrian Atomic Energy Commission and the Else
Kröner-Fresenius-Stiftung.
References
|
1
|
Hsu LY: Phenotype/karyotype correlations
of Y chromosome aneuploidy with emphasis on structural aberrations
in postnatally diagnosed cases. Am J Med Genet. 53:108–140. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yoshida A, Nakahori Y, Kuroki Y, Motoyama
M, Araki Y, Miura K and Shirai MM: Dicentric Y chromosome in an
azoospermic male. Mol Hum Reprod. 3:709–712. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tuck-Muller CM, Chen H, Martinez JE, Shen
CC, Li S, Kusyk C, Batista DA, Bhatnagar YM, Dowling E and
Wertelecki W: Isodicentric Y chromosome: cytogenetic, molecular and
clinical studies and review of the literature. Hum Genet.
96:119–129. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Krasna IH, Lee ML, Smilow P, Sciorra L and
Eierman L: Risk of malignancy in bilateral streak gonads: the role
of the Y chromosome. J Pediatr Surg. 27:1376–1380. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sinclair AH, Berta P, Palmer MS, Hawkins
JR, Griffiths BL, Smith MJ, Foster JW, Frischauf AM, Lovell-Badge R
and Goodfellow PN: A gene from the human sex-determining region
encodes a protein with homology to a conserved DNA-binding motif.
Nature. 346:240–244. 1990. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yenamandra A, Deangelo P, Aviv H, Suslak L
and Desposito F: Interstitial insertion of Y-specific DNA sequences
including SRY into chromosome 4 in a 45,X male child. Am J Med
Genet. 72:125–128. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yen PH: A long-range restriction map of
deletion interval 6 of the human Y chromosome: a region frequently
deleted in azoospermic males. Genomics. 54:5–12. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Reijo R, Lee TY, Salo P, Alagappan R,
Brown LG, Rosenberg M, Rozen S, Jaffe T, Straus D, Hovatta O, et
al: Diverse spermatogenic defects in humans caused by Y chromosome
deletions encompassing a novel RNA-binding protein gene. Nat Genet.
10:383–393. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Al-Achkar W, Wafa A, Moassass F and Liehr
T: Partial trisomy 9p22 to 9p24.2 in combination with partial
monosomy 9pter in a Syrian girl: a case report. Mol Cytogenet.
3:182010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shaffer L, Slovak M and Cambell L: ISCN
(2009): An International System for Human Cytogenetic Nomenclature.
S. Karger; Basel: 2009
|
|
11
|
Liehr T, Heller A, Starke H, Rubtsov N,
Trifonov V, Mrasek K, Weise A, Kuechler A and Claussen U:
Microdissection based high resolution multicolor banding for all 24
human chromosomes. Int J Mol Med. 9:335–339. 2002.PubMed/NCBI
|
|
12
|
Al-Achkar W, Moassass F, Al-Halabi B and
Al-Ablog A: Mutations of the Connexin 26 gene in families with
non-syndromic hearing loss. Mol Med Report. 4:331–335. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang X, Wang XR, Liu MG, Wang Q and Liu
JY: Genetic analysis of a family with 46, XY ‘female’ associated
with infertility. Acta Genetica Sinica. 33:19–25. 2006.
|
|
14
|
Therman E, Sarto GE and Patau K:
Apparently isodicentric but functionally monocentric X chromosome
in man. Am J Hum Genet. 26:83–92. 1974.PubMed/NCBI
|
|
15
|
Wolff DJ, Miller AP, Van Dyke DL, Schwartz
S and Willard HF: Molecular definition of breakpoints associated
with human Xq isochromosomes: implications for mechanisms of
formation. Am J Hum Genet. 58:154–160. 1996.PubMed/NCBI
|
|
16
|
Robinson DO, Dalton P, Jacobs PA, Mosse K,
Power MM, Skuse DH and Crolla JA: A molecular and FISH analysis of
structurally abnormal Y chromosomes in patients with Turner
syndrome. J Med Genet. 36:279–284. 1999.PubMed/NCBI
|
|
17
|
Codina-Pascual M, Oliver-Bonet M, Navarro
J, Starke H, Liehr T, Gutiérrez-Mateo C, Sánchez-García JF, Arango
O, Egozcue J and Benet J: FISH characterization of a dicentric Yq
(p11.32) isochromosome in an azoospermic male. Am J Med Genet A.
127A:302–306. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Quilter CR, Nathwani N, Conway GS,
Stanhope R, Ralph D, Bahadur G, Serhal P, Taylor K and Delhanty JD:
A comparative study between infertile males and patients with
Turner syndrome to determine the influence of sex chromosome
mosaicism and the breakpoints of structurally abnormal Y
chromosomes on phenotypic sex. J Med Genet. 39:e802002. View Article : Google Scholar
|
|
19
|
Hamberton JL, Canning N, Ray M and Smith
S: A cytogenetic survey of 14,069 newborn infants. I Incidence of
chromosome abnormalities. Clin Genet. 8:223–243. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Smyth CM and Bremner WJ: Klinefelter
syndrome. Arch Intern Med. 158:1309–1314. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Paulsen CA, Gordon DL, Carpenter RW, Gandy
HM and Drucker WD: Klinefelter's syndrome and its variants: a
hormonal and chromosomal study. Recent Prog Horm Res. 24:321–363.
1968.
|
|
22
|
Lukusa T, Fryns JP and van den Berghe H:
Gonadoblastoma and Y-chromosome fluorescence. Clin Genet.
29:311–316. 1986. View Article : Google Scholar : PubMed/NCBI
|