Introduction
Cell adhesion proteins form connections between
individual cells and mediate the interactions of cells with the
surrounding extracellular matrix (ECM) (1). In this way, they participate in various
cellular functions, including signal transduction, communication,
embryogenesis, inflammation and apoptosis.
The majority of cell adhesion proteins can be
grouped into one of five families: The immunoglobulin family, the
integrins, the cadherins, the selectins and the syndecans (1–3). Members
of the immunoglobulin family promote cell adhesion in a
calcium-independent manner. These proteins contain an extracellular
section with a variable number of immunoglobin (Ig)-like domains, a
relatively short intracellular domain and a single transmembrane
domain. For example, cell adhesion molecule-1 (CADM1; also known as
nectin-like protein 2) contains three extracellular Ig-like domains
and binds with its intracellular domain to the adapter protein
DAL1, which in turn anchors CADM1 to the actin cytoskeleton
(1–3).
Integrins are involved in the integration of the ECM with the
cytoskeleton. These proteins represent noncovalently linked
heterodimers composed of an α and a β subunit. The majority of
integrins acts as receptors for ECM proteins, including
fibronectin, vitronectin, collagens and laminin, and recognizes the
Arg-Gly-Asp sequence within target proteins (1–3). Cadherins
are calcium-dependent cell adhesion molecules. E-cadherin, for
example, contains an extracellular domain with 5 cadherin repeats,
an intracellular domain and a transmembrane domain. The
intracellular domain interacts with catenin and binds to the actin
cytoskeleton (1–3). Selectins mediate the interactions
between leukocytes and endothelial cells. They are composed of a
single transmembrane domain, a short intracellular domain and an
extracellular domain with a variable number of sushi motifs.
Typically, selectins assist with the homing of lymphocytes to the
lymph node (1–3). Finally, syndecans are proteoglycans
located on the cell membrane. They contain a variable number of
glycosaminoglycan chains attached to strategic serine residues of
the polypeptide chain. In addition to mediating cell-matrix and
cell-cell interactions, syndecans contribute to modulating the
activity of heparin binding growth factors (1–3).
Evidence from previous studies indicates that cell
adhesion proteins participate in tumor formation and metastasis
(2,3).
Indeed, the progression of cancer is often associated with the loss
of at least one cell adhesion protein. However, cell adhesion
proteins can also function as tumor suppressors when overexpressed
in tumor tissues (2,3). It appears that their forced expression
restores contact inhibition, a phenomenon observed in normal cells,
but not in the majority of malignant cells. E-cadherin, for
example, is often lost in tumors of epithelial origin and it has
been demonstrated that the disruption of cell adhesion mediated by
E-cadherin is associated with the development and progression of
cancer (4). The forced expression of
E-cadherin in tumor cell lines slows down cell proliferation and
reduces cell invasiveness. By contrast, inhibiting E-cadherin using
antibodies or antisense RNA restores the invasiveness of the cells
(4). Likewise, integrin α7 is
frequently mutated in human malignancies and such mutations are
associated with cancer recurrence (5). It has been demonstrated that the
increased expression of integrin α7 in leiomyosarcoma cells
resulted in the reduction of colony formation. Moreover, increasing
the expression of α7 integrin in mice with xenografted tumors
inhibited tumor growth (5). Likewise,
CADM1 has been implicated in cancer progression. Downregulation of
CADM1 synthesis has been observed in a variety of human tumors,
including breast cancer and esophageal squamous cell carcinoma
(6,7).
However, it has been demonstrated that restoring CADM1 expression
suppresses cell growth and slows down tumor invasion (7). It has therefore been concluded that the
majority of the cell adhesion proteins can function as tumor
suppressors (2,3).
A previous study by our group described a novel
transmembrane protein that resembles CADM1 (8,9). This
protein contains three extracellular Ig-like domains, a relatively
short intracellular domain and a single transmembrane domain. It
interacts with heparin and fibroblast growth factors (FGFs) in a
manner similar to the classical FGF receptors (FRFRs), FGFR1-FGFR4
(10,11). For this reason, the novel protein was
termed FGFR-like 1 protein (FGFRL1). However, FGFRL1 does not
possess the intracellular tyrosine kinase domain required for
signal transduction by transphosphorylation and consequently cannot
mediate FGF signaling by itself.
The function of the novel receptor is currently
unclear. Knockout mice with a targeted disruption of the FGFRL1
gene present a striking phenotype; they lack metanephric kidneys
(12) and die at birth due to a weak,
malformed diaphragm that cannot inflate the lungs after birth
(13,14). Cell culture experiments have
demonstrated that FGFRL1 can act as a typical cell adhesion protein
when coated on plastic dishes. FGFRL1 forms heterophilic
interactions with heparan sulfate proteoglycans, such as glypican,
at the cell surface of neighboring cells (15). A tetracycline-inducible system has
been utilized, in which the expression of FGFRL1 could be
controlled by the addition of the inducer doxycycline (16). It was observed that in the presence of
doxycycline, cells aggregated and formed huge clusters, whereas in
its absence, they tended to remain as individual clones (17).
In the present study, one of the
tetracycline-inducible cell clones was characterized in more
detail. The primary aim of the study was to determine whether
FGFRL1 functions as a tumor suppressor in a manner similar to other
cell adhesion proteins.
Materials and methods
Cell culture
The generation of tetracycline-inducible cell clones
has been described in detail in a recent publication (17). The stable clone K13ΔC was produced by
transfection of a cDNA for truncated FGFRL1 (corresponding to amino
acid residues 1–417) into HEK 293 Tet-On® cells
(Clontech Laboratories, Takara Bio Europe SAS,
Saint-Germain-en-Laye, France). The cells were cultivated in an
atmosphere of 5% CO2 in Dulbecco's modified Eagle's
medium (DMEM) supplemented with 10% fetal bovine serum, 100 U/ml
penicillin and 100 µg/ml streptomycin (all Sigma-Aldrich, Buchs,
Switzerland). To maintain the selective pressure for stable
transfection, 100 µg/ml Hygromycin B (InvivoGen, San Diego, CA,
USA) was used. Expression of FGFRL1 from the tetracycline
responsive promoter was induced by the addition of 1 µg/ml
doxycycline.
Adhesion experiments
Recombinant FGFRL1 protein was isolated from the
conditioned media of HEK 293 cells that had been stably transfected
with a cDNA for human FGFRL1 (corresponding to amino acid residues
1–357), as previously described (18). The recombinant protein was purified by
chromatography on a column of Heparin Sepharose® 6 Fast
Flow (GE Healthcare Bio-Sciences, Pittsburgh, PA, USA) as
previously described (18). FGFRL1
protein or bovine serum albumin (BSA; Sigma-Aldrich), which served
as a control, was diluted to 20 µg/ml in phosphate-buffered saline
(PBS) and droplets of the solution (15 µl) were spotted onto 35-mm
petri dishes (non-tissue culture; catalog no. 82.1135; Sarstedt
Co., Nümbrecht, Germany). Following incubation in a humidified
chamber at 4°C for 16 h, the solution was carefully aspirated and
all residual sites of the petri dish were blocked with 1% BSA in
PBS. Cells were seeded onto pre-coated petri dishes in serum-free
medium (2×106 cells/plate) and incubated for 1 h at
37°C. Non-adherent cells were carefully removed by washing with
PBS. Adherent cells were fixed with 4% paraformaldehyde
(Sigma-Aldrich) and inspected under a microscope (Nikon Eclipse
E800; Nikon AG, Zurich, Switzerland).
In a further experiment, the cells were cultivated
in complete growth medium (as aforementioned) on uncoated petri
dishes (non-tissue culture) for 1–3 days, and cell adhesion and
cell-cell clustering were documented with a Zeiss Axiovert 10
microscope (Carl Zeiss AG, Oberkochen, Germany).
Immunocytochemistry
Cells grown on coverslips were washed with ice-cold
PBS, fixed with 4% paraformaldehyde and permeabilized with 0.2%
Triton X-100 (Sigma-Aldrich) in PBS. Non-specific sites were
blocked with 3% BSA in PBS. The fixed cells were incubated for 3 h
at room temperature with a humanized monoclonal antibody (1 µg/ml)
against FGFRL1 that had been prepared by our group in a previous
study (19). Following three steps of
washing with PBS, bound antibodies were detected with Cy2-labeled
secondary antibodies (catalog no. 109-225-097; dilution 1:200;
Jackson ImmunoResearch Laboratories, West Grove, PA, USA). The
nuclei of the cells were stained with 1 µg/ml
4′,6-diamidino-2-phenylindole (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). To detect filamentous actin,
fixed cells were treated in a similar way with
tetramethylrhodamine-labeled phalloidin (Sigma-Aldrich).
Electron microscopy
Cells grown on cover slips were fixed with 2.5%
glutaraldehyde in 30 mM potassium phosphate buffer. The fixed cells
were dehydrated in ethanol, critical point dried and sputter-coated
with gold, as previously described (20). Finally, the specimens were inspected
with a Philips XL 30 FEG scanning electron microscope operated at
10 kV (Philips, Amsterdam, The Netherlands).
Nude mice xenograft experiments
To investigate tumorigenicity, immunodeficient
CD-1® NU/NU-Foxn1 nude mice (Charles River Wiga GmbH,
Sulzfeld, Germany) were utilized. Approximately 1×107
K13ΔC cells in PBS were subcutaneously injected at two ventral
sites into the animals. Half of the mice (4 mice per group,
randomly selected) were treated with 100 µg/ml doxycycline, which
was directly added to the drinking water of the animals; the other
half received regular water. After 5 weeks, the mice were
sacrificed and images of the tumors were captured. All animal
experiments had been approved by the Ethics committee of the County
of Bern.
Statistical analysis
The significance of the results from the xenograft
experiments was analyzed with the exact Fisher test utilizing an
online calculation tool (http://www.quantitativeskills.com). P≤0.05 was
considered to indicate a statistically significant difference.
Results
FGFRL1 overexpression in cell
culture
All the following experiments were performed with
clone K13ΔC from the Tet-On-FGFRL1ΔC cell line (17). This clone had been prepared with cDNA
for FGFRL1, which covered the extracellular domain and the
transmembrane helix of the protein, but lacked the intracellular
domain. Hence, all effects observed with this clone may be
attributed to the extracellular and the transmembrane domains of
FGFRL1.
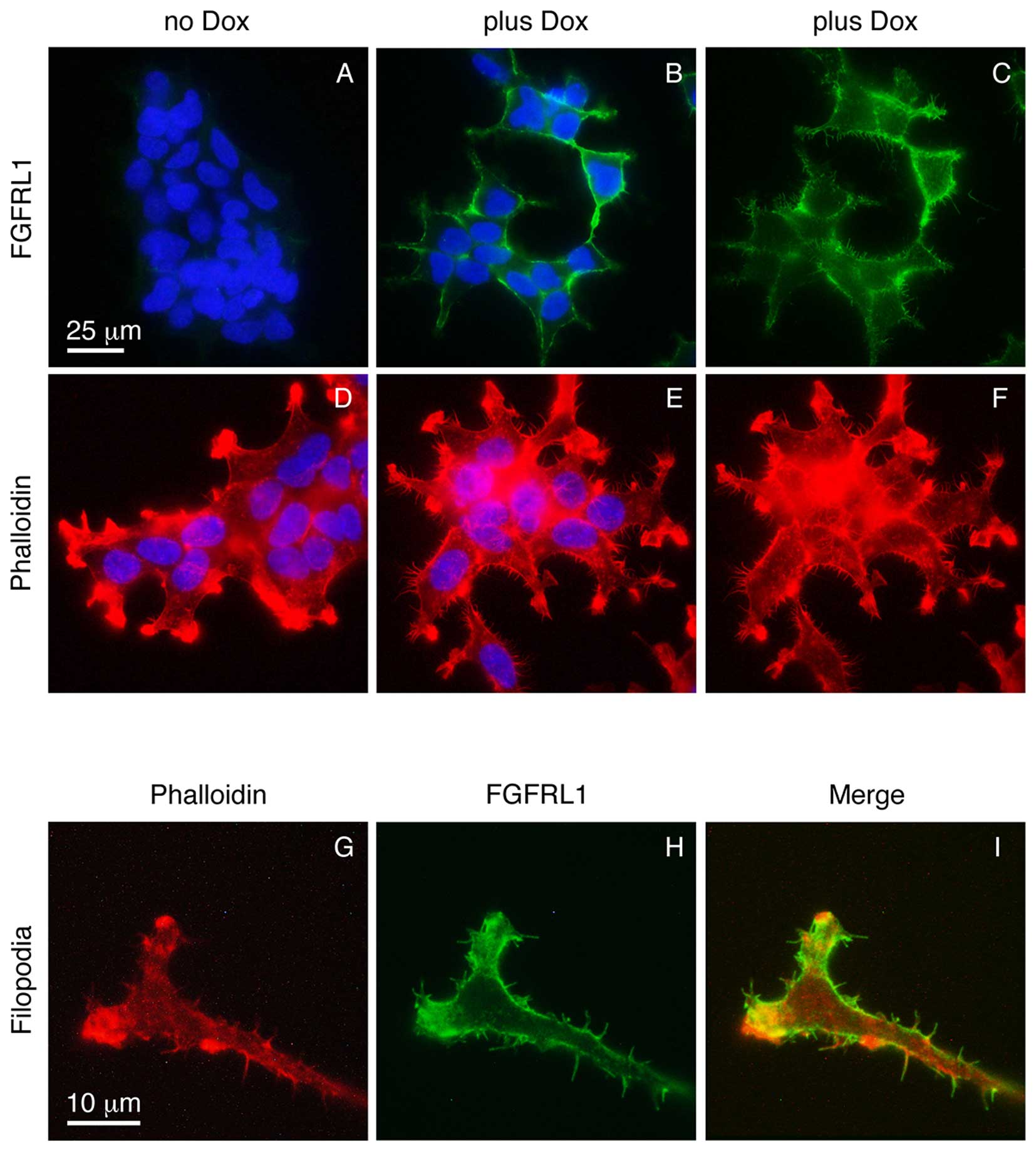
In the absence of doxycycline, the K13ΔC clone did
not express any detectable FGFRL1ΔC protein, as verified with our
monoclonal antibodies (Fig. 1A).
Following induction with doxycycline, a strong signal for FGFRL1
was observed at the cell membrane (Fig.
1B). Thus, K13ΔC cells tightly controlled the expression of
FGFRL1ΔC depending on the presence or absence of doxycycline. When
the focus of the microscope lens was changed to the plane of the
glass slide (rather than the site of strongest fluorescence), some
fluorescent signal was observed at numerous protrusions that
emerged from the plasma membrane of the cells in a spike-like
fashion (Fig. 1C). To investigate
whether these microspikes represented normal filopodia, the cells
were stained with fluorescently labeled phalloidin, which is known
to interact with filamentous actin. Again, a number of spikes was
observed that protruded in a perpendicular manner from the plasma
membrane (Fig. 1D-F). At higher
magnification (objective lens, 100X), the FGFRL1ΔC signal (green)
partially co-localized with the phalloidin signal (red) (Fig. 1G-I). Additionally, the spikes were
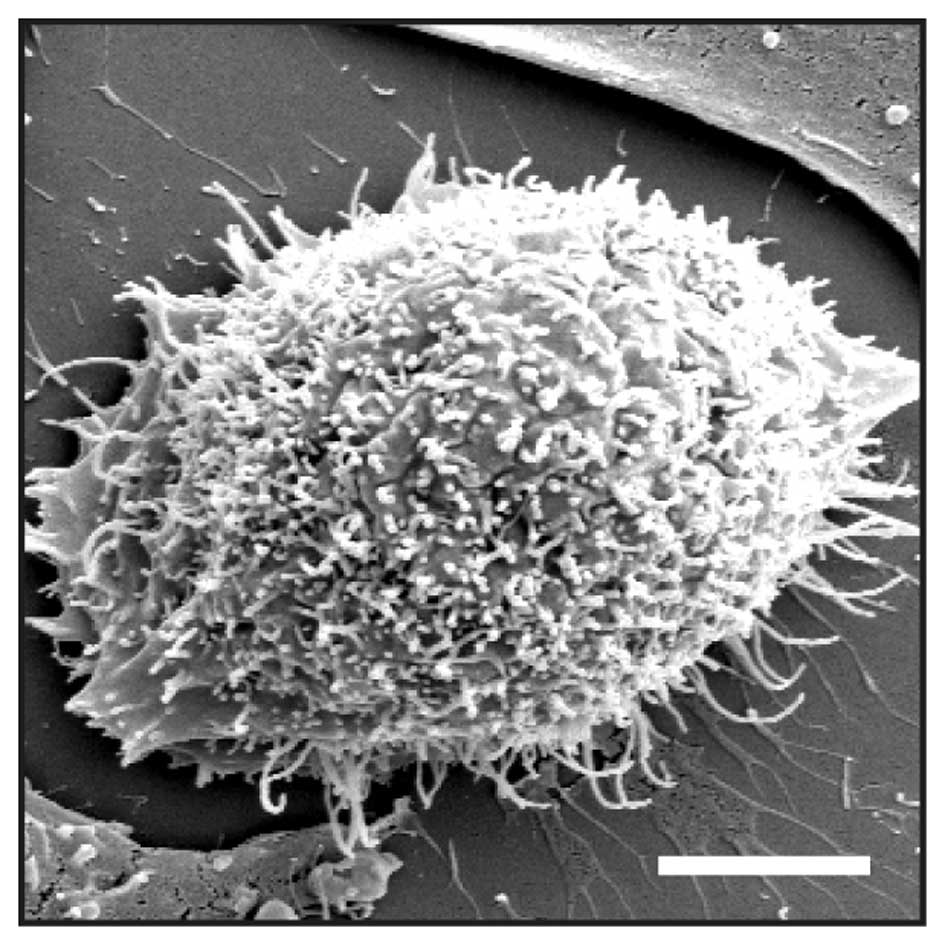
visualized under a scanning electron microscope (Fig. 2). Here, numerous protrusions were
detected that had an average diameter of 180±28 nm (n=6). It is
therefore likely that the spikes represent regular filopodia.
Notably, a much larger number of filopodia was observed in the
presence of doxycycline compared with the number observed in its
absence (Fig. 1D-F). Thus, forced
expression of FGFRL1ΔC appears to stimulate the formation of
filopodia.
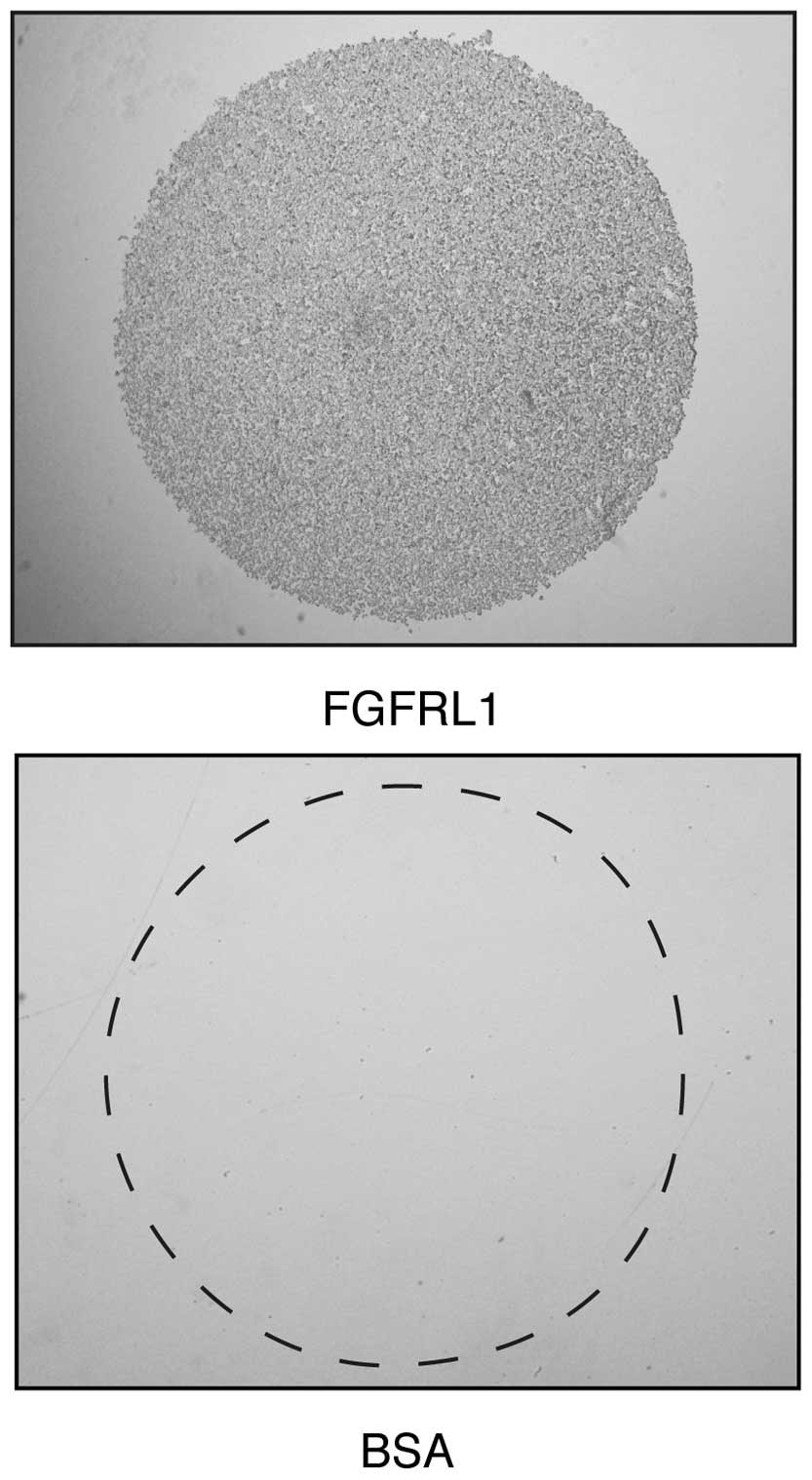
Next, it was investigated whether K13ΔC cells would
bind to purified FGFRL1 protein, as has previously been documented
in Chinese hamster ovary (CHO) cells (15). The surface of a plastic plate
(non-tissue culture) was coated with droplets of recombinant FGFRL1
solution comprising the Ig1-Ig3 domains, or with droplets of BSA
that served as a control. Within 1 h, the K13ΔC cells attached to
the recombinant FGFRL1 protein, however, they did not attach to the
control BSA (Fig. 3). It is likely
that this interaction was accomplished by the binding of FGFRL1 to
cell surface heparan sulfate proteoglycans, since the binding could
be blocked with soluble heparin (data not shown), as previously
reported in CHO cells (15).
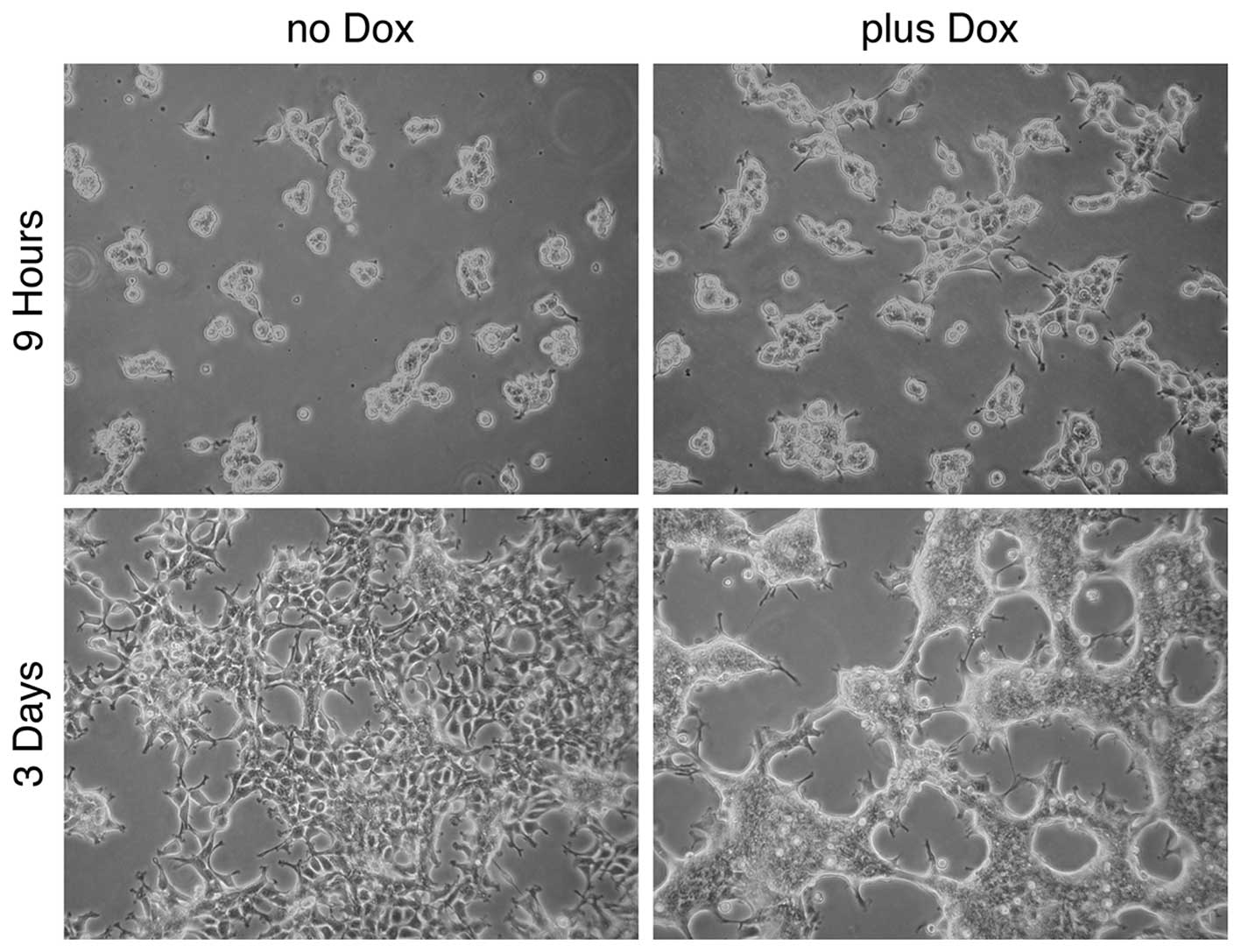
In the following experiment, the effect of
doxycycline (and therefore the effect of FGFRL1ΔC) on cell-cell
adhesion and cell clustering was recapitulated (Fig. 4). K13ΔC cells formed large patches of
10–30 cells following 9 h of incubation on bacterial plates in the
presence of doxycycline. In the absence of the inducer, the cells
also started to form clusters, however, these clusters were much
smaller. After 3 days in culture, the K13ΔC cells had merged to
form a continuous network of cells. Notably, the borders between
the individual cells were no longer distinguishable in the presence
of doxycycline, whereas they were clearly detectable in its
absence. Thus, the presence of FGFRL1ΔC promotes tight, intimate
interactions between cells. Taken together, the three experiments
suggested that FGFRL1 represents a typical cell adhesion
protein.
Xenograft tumor model
The majority of cell adhesion proteins can function
as tumor suppressors (2,3); therefore, the effects of FGFRL1 on tumor
formation were investigated. K13ΔC cells were injected
subcutaneously into immunocompromised nude mice at two ventral
sites. Half of the mice received doxycycline in their drinking
water in order to induce the expression of FGFRL1ΔC. The other half
served as controls and received regular drinking water. Within 5
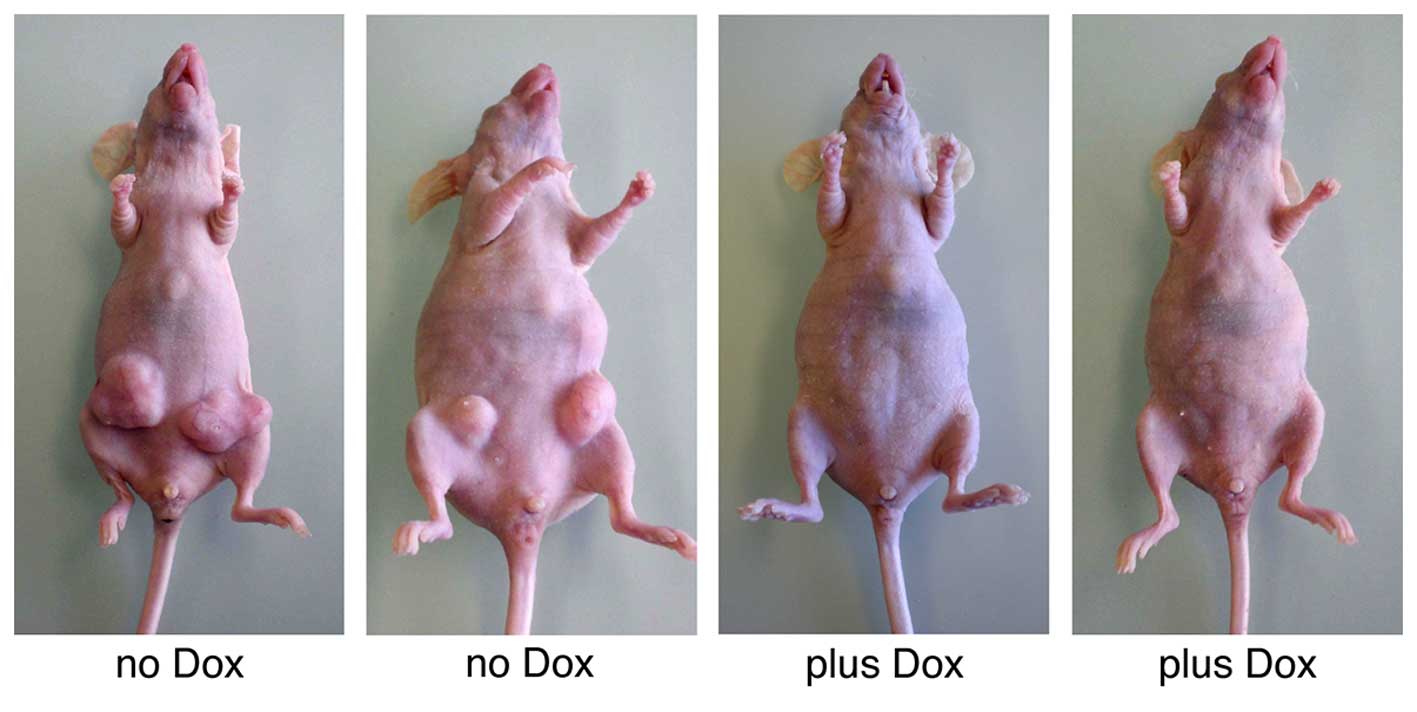
weeks, the control animals had developed large tumors (diameter ≥12
mm) at five out of the eight injection sites (Fig. 5). In sharp contrast, none of the
doxycycline-treated mice had produced any tumors (P=0.0257). A
control experiment confirmed that doxycycline itself had no effect
on tumor growth (data not shown). Thus, FGFRL1 may function as a
typical tumor suppressor that effectively inhibits the outgrowth of
xenografted tumors in vivo.
Discussion
The current study, taken together with results from
the literature, demonstrates that the novel receptor FGFRL1
represents a regular cell-cell adhesion protein. This conclusion is
based on the following facts: i) The domain structure of FGFRL1,
which has a single transmembrane domain, three extracellular
Ig-like repeats and a short intracellular domain (21), resembles the structure of other cell
adhesion proteins from the Ig superfamily, namely the nectins and
nectin-like molecules (22,23). ii) Recombinant polypeptides
corresponding to the extracellular domain of FGFRL1 promote cell
adhesion in vitro. If a mutation is introduced into the
polypeptide chain, the activity is completely lost (15). iii) When overexpressed in different
cell lines, FGFRL1 protein accumulates at intersections where two
cells touch each other (15).
Overexpression in HEK 293 Tet-On cells leads to the aggregation of
the cells and to the formation of large clusters (17). iv) Finally, the results of the current
study indicate that FGFRL1, like other typical cell adhesion
proteins, acts as a tumor suppressor in a xenograft tumor model. It
was determined that the forced expression of FGFRL1 in HEK 293
Tet-On cells completely inhibited the outgrowth of tumors in
immunocompromised mice.
During the xenograft experiments, it was noted that
the cells were not extremely tumorigenic. A large number of HEK 293
Tet-On-FGFRL1ΔC cells had to be injected into the mice to initiate
any tumor growth. Other studies have also noted that HEK 293 cells
exhibit particularly low tumorigenicity. Shen et al
(24) demonstrated that HEK 293 cell
tumorigenicity increased with increasing passage number, and
finally reached 100% when the passage number was >65; however,
the original isolates of the cells did not form tumors at all in
nude mice. In the present study, an outgrowth of tumors was
detected in the absence of doxycycline in 5 cases (63%). Following
induction of FGFRL1 synthesis, tumor growth was observed in no
cases (0%). Doxycycline itself had no effect on tumor growth, as
demonstrated in a control experiment and as previously published in
the literature (25). Thus, it is
FGFRL1 that can act as a tumor suppressor.
Previous studies have reported that alterations in
the synthesis of FGFRL1 occur in tumor cells (26–31). The
screening of 241 different human tumor samples with a
cancer-profiling array suggested that major changes in the relative
expression of FGFRL1 occur in ovarian tumors (26). In several samples, a significant
decrease in FGFRL1 expression was observed in the tumor tissue
relative to the matched control tissue. However, in one ovarian
tumor sample there was a 25-fold increase (26). Furthermore, the overexpression of
FGFRL1 in certain ovarian tumor samples was confirmed in a study
aiming to identify novel tumor-specific marker genes (27). An association of FGFRL1 expression
with tumor growth and metastasis was also suggested by a study of
patients suffering from head and neck tumors (28,29). In
this case, FGFRL1 overexpression appeared to correlate with tumor
growth. Another study noted a significant decrease of FGFRL1
protein expression in bladder tumors and bladder cancer cell lines
(30). This decrease was explained by
heterozygous deletions at the chromosomal region 4p16.3, which
includes the locus of the FGFRL1 gene. Finally, mutations affecting
the reading frame of FGFRL1 have been observed relatively
frequently in colorectal cancer cell lines (31). The majority of these mutations
occurred in the intracellular domain of the protein.
The molecular mechanism governing how FGFRL1 may
inhibit tumor growth in a xenograft model is complex. With other
cell adhesion proteins it has been concluded that forced expression
partially restores contact inhibition of the tumor cells. In the
case of CADM1, it was demonstrated that homophilic interactions of
CADM1 at the surface of two adjacent cells activated the
phosphatidylinositol 3-kinase (PI3K) pathway and led to the
reorganization of the actin cytoskeleton (32). The intracellular domain of CADM1
formed a complex with membrane-associated guanylate kinase
homologues, including MPP3 and Dlg, thus linking CADM1 and PI3K. In
the case of FGFRL1, the effect may be exerted by heterophilic
interactions of the extracellular domain of FGFRL1 with another
transmembrane protein, since the protein expressed by K13ΔC cells
lacks the intracellular domain of FGFRL1. The extracellular domain
has the ability to interact with target proteins of neighboring
cells containing heparan sulfate chains, including syndecans and
glypicans (15). Two target proteins,
glypican-4 and glypican-6, which specifically interact with FGFRL1,
have previously been identified by our group using tandem LC mass
spectrometry (16). Therefore, it is
hypothesized that FGFRL1 inhibited tumor growth in the current
study in the following manner: HEK 293 Tet-On cells lost their
normal contact inhibition and grew in an unrestricted way, as they
were subcloned multiple times during the generation of the HEK 293
Tet-On-FGFRL1ΔC clones. When injected into nude mice, these cells
gave rise to large tumors due to a defect in contact inhibition.
Enforcing the expression of FGFRL1ΔC in the injected cells by
adding doxycycline induced the adhesion of the cells with each
other and restored contact inhibition, thereby inhibiting tumor
growth.
It remains to be demonstrated whether the
identification of FGFRL1 as a tumor suppressor may be exploited to
improve the diagnosis and therapy of cancer patients. FGFRL1 may
serve as a tumor marker to identify aggressive tumors that have
lost FGFRL1 expression (27,31). Furthermore, there may be the
possibility of enhancing FGFRL1 synthesis in the tumor tissue.
Levels of FGFRL1 mRNA are regulated by microRNA-120 (28,33), which
specifically interacts with the 3′ end of the FGFRL1 mRNA and leads
to its degradation. Therefore, if it were possible to downregulate
microRNA-120 expression in the tumor tissue, the endogenous levels
of FGFRL1 would increase and contact inhibition of the cells in the
tumor tissue would be restored, thus suppressing tumor growth.
Future studies are necessary to determine whether this could be
developed as a novel therapeutic strategy for patients with
cancer.
In conclusion, the current study taken together with
results from the literature, demonstrates that FGFRL1 is a
cell-cell adhesion protein that acts as a tumor suppressor similar
to numerous other cell adhesion proteins.
Acknowledgements
The current study was supported by grants from the
Swiss National Science Foundation (Bern, Switzerland; no.
31003A-143350), the Novartis Foundation for Medical-Biological
Research (Basel, Switzerland; no. 15A006) and the Swiss Foundation
for Research on muscular diseases (Cortaillod, Switzerland; no.
2015–12-18).
Glossary
Abbreviations
Abbreviations:
|
FGF
|
fibroblast growth factor
|
|
FGFR
|
fibroblast growth factor receptor
|
|
FGFRL1
|
FGFR-like protein 1
|
References
|
1
|
Kreis T and Vale R: Guidebook to the
Extracellular MatrixAnchor and Adhesion Proteins. 2nd. Oxford
University Press; 1999
|
|
2
|
Okegawa T, Li Y, Pong RC and Hsieh JT:
Cell adhesion proteins as tumor suppressors. J Urol. 167:1836–1843.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Moh MC and Shen S: The roles of cell
adhesion molecules in tumor suppression and cell migration: A new
paradox. Cell Adh Migr. 3:334–336. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pećina-Slaus N: Tumor suppressor gene
E-cadherin and its role in normal and malignant cells. Cancer Cell
Int. 3:172003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ren B, Yu YP, Tseng GC, Wu C, Chen K, Rao
UN, Nelson J, Michalopoulos GK and Luo JH: Analysis of integrin
alpha7 mutations in prostate cancer, liver cancer, glioblastoma
multiforme, and leiomyosarcoma. J Natl Cancer Inst. 99:868–880.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Heller G, Geradts J, Ziegler B, Newsham I,
Filipits M, Markis-Ritzinger EM, Kandioler D, Berger W, Stiglbauer
W, Depisch D, et al: Downregulation of TSLC1 and DAL-1 expression
occurs frequently in breast cancer. Breast Cancer Res Treat.
103:283–291. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ito T, Shimada Y, Hashimoto Y, Kaganoi J,
Kan T, Watanabe G, Murakami Y and Imamura M: Involvement of TSLC1
in progression of esophageal squamous cell carcinoma. Cancer Res.
63:6320–6326. 2003.PubMed/NCBI
|
|
8
|
Wiedemann M and Trueb B: Characterization
of a novel protein (FGFRL1) from human cartilage related to FGF
receptors. Genomics. 69:275–279. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Trueb B: Biology of FGFRL1, the fifth
fibroblast growth factor receptor. Cell Mol Life Sci. 68:951–964.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Trueb B, Zhuang L, Taeschler S and
Wiedemann M: Characterization of FGFRL1, a novel fibroblast growth
factor (FGF) receptor preferentially expressed in skeletal tissues.
J Biol Chem. 278:33857–33865. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ornitz DM and Itoh N: The fibroblast
growth factor signaling pathway. Wiley Interdiscip Rev Dev Biol.
4:215–266. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gerber SD, Steinberg F, Beyeler M,
Villiger PM and Trueb B: The murine Fgfrl1 receptor is essential
for the development of the metanephric kidney. Dev Biol.
335:106–119. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Baertschi S, Zhuang L and Trueb B: Mice
with a targeted disruption of the Fgfrl1 gene die at birth due to
alterations in the diaphragm. FEBS J. 274:6241–6253. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Amann R, Wyder S, Slavotinek AM and Trueb
B: The FgfrL1 receptor is required for development of slow muscle
fibers. Dev Biol. 394:228–241. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rieckmann T, Kotevic I and Trueb B: The
cell surface receptor FGFRL1 forms constitutive dimers that promote
cell adhesion. Exp Cell Res. 314:1071–1081. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Steinberg F, Gerber SD, Rieckmann T and
Trueb B: Rapid fusion and syncytium formation of heterologous cells
upon expression of the FGFRL1 receptor. J Biol Chem.
285:37704–37715. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang X, Steinberg F, Zhuang L, Bessey R
and Trueb B: Receptor FGFRL1 does not promote cell proliferation
but induces cell adhesion. Int J Mol Med. 38:30–38. 2016.PubMed/NCBI
|
|
18
|
Zhuang L, Gerber SD, Kuchen S, Villiger PM
and Trueb B: Deletion of exon 8 from the EXT1 gene causes multiple
osteochondromas (MO) in a family with three affected members.
Springerplus. 5:712016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rieckmann T, Zhuang L, Flück CE and Trueb
B: Characterization of the first FGFRL1 mutation identified in a
craniosynostosis patient. Biochim Biophys Acta. 1792:112–121. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Henning A, Schneider M, Bur M, Blank F,
Gehr P and Lehr CM: Embryonic chicken trachea as a new in vitro
model for the investigation of mucociliary particle clearance in
the airways. AAPS PharmSciTech. 9:521–527. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhuang L, Falquet L and Trueb B:
Genome-wide comparison of FGFRL1 with structurally related surface
receptors. Exp Ther Med. 1:161–168. 2010.PubMed/NCBI
|
|
22
|
Mandai K, Rikitake Y, Mori M and Takai Y:
Nectins and nectin-like molecules in development and disease. Curr
Top Dev Biol. 112:197–231. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Samanta D and Almo SC: Nectin family of
cell-adhesion molecules: Structural and molecular aspects of
function and specificity. Cell Mol Life Sci. 72:645–658. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shen C, Gu M, Song C, Miao L, Hu L, Liang
D and Zheng C: The tumorigenicity diversification in human
embryonic kidney 293 cell line cultured in vitro. Biologicals.
36:263–268. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tanaka T and Rabbitts TH: Interfering with
RAS-effector protein interactions prevent RAS-dependent tumour
initiation and causes stop-start control of cancer growth.
Oncogene. 29:6064–6070. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schild C and Trueb: Aberrant expression of
FGFRL1, a novel FGF receptor, in ovarian tumors. Int J Mol Med.
16:1169–1173. 2005.PubMed/NCBI
|
|
27
|
Barrett CL, DeBoever C, Jepsen K, Saenz
CC, Carson DA and Frazer KA: Systematic transcriptome analysis
reveals tumor-specific isoforms for ovarian cancer diagnosis and
therapy. Proc Natl Acad Sci USA. 112:E3050–E3057. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tsuchiya S, Fujiwara T, Sato F, Shimada Y,
Tanaka E, Sakai Y, Shimizu K and Tsujimoto G: MicroRNA-210
regulates cancer cell proliferation through targeting fibroblast
growth factor receptor-like 1 (FGFRL1). J Biol Chem. 286:420–428.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shimada Y, Okumura T, Nagata T, Hashimoto
I, Sawada S, Yoshida T, Fukuoka J, Shimizu K and Tsukada K:
Expression analysis of fibroblast growth factor receptor-like 1
(FGFRL1) in esophageal squamous cell carcinoma. Esophagus.
11:48–53. 2014. View Article : Google Scholar
|
|
30
|
di Martino E, Taylor CF, Roulson JA and
Knowles MA: An integrated genomic, transcriptional and protein
investigation of FGFRL1 as a putative 4p16.3 deletion target in
bladder cancer. Genes Chromosomes Cancer. 52:860–871. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Donnard E, Asprino PF, Correa BR, Bettoni
F, Koyama FC, Navarro FC, Perez RO, Mariadason J, Sieber OM,
Strausberg RL, et al: Mutational analysis of genes coding for cell
surface proteins in colorectal cancer cell lines reveal novel
altered pathways, druggable mutations and mutated epitopes for
targeted therapy. Oncotarget. 5:9199–9213. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Murakami S, Sakurai-Yageta M, Maruyama T
and Murakami Y: Trans-homophilic interaction of CADM1 activates
PI3K by forming a complex with MAGuK-family proteins MPP3 and Dlg.
PLoS One. 9:e1100622014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Huang X, Ding L, Bennewith KL, Tong RT,
Welford SM, Ang KK, Story M, Le QT and Giaccia AJ:
Hypoxia-inducible mir-210 regulates normoxic gene expression
involved in tumor initiation. Mol Cell. 35:856–867. 2009.
View Article : Google Scholar : PubMed/NCBI
|