Introduction
Phytochemicals have been applied as a
multi-targeting approach to cancer medicine due to their potential
to improve the efficiency of chemotherapy protocols (1). Tumor cells produce high levels of
antioxidants that neutralize free radicals, thus creating a
negative balance of intracellular reactive oxygen species (ROS)
levels, which facilitates the survival of cancer cells.
Antioxidants help to alleviate the toxic effects of free
radical-producing drugs and preserve the health of normal tissues
patients with cancer (2). However, it
has been demonstrated that the up regulation of antioxidant systems
may provide the same protection to tumor cells against oxidative
damage, and subsequently, may stimulate tumor progression by
increasing the aggressiveness and chemoresistance of tumor cells
(2).
Nuclear factor erythroid 2-related factor 2 (Nrf2)
has been recognized as a member of the cap ‘n’ collar subfamily,
and regulates the intracellular antioxidant response through the
controlled activation of a series of genes, including phase-II
detoxifying enzymes, endogenous antioxidants and transporters that
shield cells from the harmful effects of carcinogens and
environmental toxins (3–5). Overexpression of Nrf2 and its downstream
target genes has been identified in numerous primary tumors, and
may protect cancer cells against the cytotoxic effects of
chemotherapeutic agents (6,7). Therefore, understanding the signaling
pathway of Nrf2 is essential in tumor biology, and the application
of Nrf2 inhibitors may be a useful method of treating tumors
(8–11).
Vitamin C (VC) is known as one of the most prominent
antioxidative components, which may exert chemopreventive effects
without perceptible toxic side effects (12). VC produces cytotoxic levels of
hydrogen peroxide and kills cancer cells at pharmacological
concentrations, as tumor cells are often catalase-deficient and
more susceptible to hydrogen peroxide than normal cells (12). It has been demonstrated that ascorbic
acid (a form of VC) protects normal cells against oxidative stress
in mice, suggesting that VC may be used as an adjuvant for cancer
treatment (13–15). Additionally, ascorbic acid inhibits
Nrf2 activation by interfering with Nrf2 nuclear localization and
it's binding to the antioxidant response element (ARE) sequence
(16).
One of the most abundant flavonoids in fruits and
vegetables is quercetin (Q), which has been shown to exert
anticancer actions, such as the blocking of tumor initiation
(17), in addition to exerting
anti-oxidative (18) and
anti-apoptotic activities in different cancer cell lines (19,20).
Notably, Q has the capacity to act either as an antioxidant or as a
pro-oxidant, depending on its concentration and the period of
exposure (21,22). It has been demonstrated that high
doses of Q decrease cell survival rates and diminish the levels and
activities of cellular antioxidants, thus enhancing antitumor
effects (23). By contrast, low doses
of Q augment the total antioxidant capacity of cancer cells and
counteract the cytotoxic effects of antineoplastic drugs in lung
cancer A549, colorectal cancer HCT116 and ovarian cancer cells
(22). Q rapidly stimulates Nrf2
phosphorylation and translocation to the cytosol (24). However, long-term treatment of the
cells with Q inhibits both these effects and transiently induces
the activation of p38 mitogen-activated protein kinases (24).
In the present study, it was hypothesized that the
combined effects of Q (a multiple signaling inhibitor) and VC (an
antioxidant agent with antineoplastic activity) could exert a
synergistic effect on ROS levels in cancer cells via inhibition of
Nrf2. The cytotoxicity of Q and VC in various cancer cells was
examined, and the effectiveness of the Nrf2 pathway was
investigated at the gene and protein levels.
Materials and methods
Reagents
RPMI-1640 medium and 10% fetal bovine serum (FBS)
were obtained from Invitrogen (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). Primary rabbit polyclonal anti-Nrf2 (sc-722) and
anti-β-actin antibodies (sc-47778) were purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). Q and VC were purchased from
Sigma-Aldrich (Merck Millipore, Darmstadt, Germany). The human
breast cancer cell lines MDA-MB 231, MDA-MB 468 and MCF-7, and the
human lung cancer cell line A549, were all obtained from the
National Cell Bank of Iran, Pasteur Institute of Iran (Tehran,
Iran).
Cell cytotoxicity study
Cancer cells were seeded at a density of
1×104 cells/well in a 96-well plate and cultured in
RPMI-1640 medium containing 10% FBS, 100 U/ml penicillin and 100
µg/ml streptomycin at 37°C in a humidified 5% CO2
atmosphere. Cells at passages 3–5 were used in subsequent
experiments after reaching 70% confluence. Cells were exposed to
varying concentrations of Q and VC (0.1–1,000 µM) for 24 h.
Subsequently, 100 µl 0.5 mg/ml MTT solution in PBS was added per
well, and the plate was incubated at 37°C for 3 h in the dark. The
absorbance was then measured at 570 nm in an ELISA reader (Mikura
Ltd., Horsham, UK).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Cancer cells were cultured at a density of
5×105 cells/well in a 6-well plate, and following
incubation for 18–24 h, cells were treated with 200 or 100 µM VC
for 24 h. Then, 50 or 75 µM Q, respectively, was added for 6 h.
Following isolation of RNA using BioZOL RNA extraction reagent
(BioFlux Corporation, Tokyo, Japan), the amount of RNA was
determined with a NanoDrop 1000 spectrophotometer (NanoDrop
Technologies; Thermo Fisher Scientific, Inc., Wilmington, DE, USA).
Total RNA was immediately reverse transcribed to generate
first-strand complementary DNA using an RT kit (Fermentas; Thermo
Fisher Scientific, Inc., Pittsburgh, PA, USA), according to the
manufacturer's protocol. Specific primers for Nrf2 were used to
detect Nrf2 expression: Forward, 5′-ACACGGTCCACAGCTCATC-3′ and
reverse, 5′-TGTCAATCAAATCCATGTCCTG-3′. The levels of β-actin and
ribosomal protein lateral stalk subunit P0 (RPLP0) were also
analyzed as reference genes. The primers used were as follows:
β-actin forward, 5′-AATCGTGCGTGACATTAAG-3′ and reverse,
5′-GAAGGAAGGCTGGAAGAG-3′; and RPLPO forward,
5′-GAAGGCTGTGGTGCTGATGG-3′ and reverse, 5′-CCGGATATGAGGCAGCAGTT-3′.
qPCR was performed using SYBR® Premix Ex Taq™ II (Tli
RNaseH Plus) (Takara Bio, Inc., Otsu, Japan) and analyzed using the
software provided in the StepOnePlus™ Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). qPCR amplification was
carried out for 25 cycles using the following protocol: 95°C for 10
min, 94°C for 30 sec, 55°C for 30 sec, 72°C for 30 sec and 72°C for
10 min. The Pfaffl method was used for the relative mRNA
quantification, as described previously (25,26).
Western blot analysis
To detect Nrf2 protein levels, cells at a density of
12×105 cells/T75 flask were cultured for 18–24 h.
Firstly, they were exposed to 200 or 100 µM VC for 24 h.
Subsequently, 50 or 75 µM Q (respectively) was added for 6 h. Cells
were then lysed at 4°C in a buffer containing 50 mM Tris, 20 mM
NaCl and 200 µl NP-40 in a final volume of 20 ml (pH 8.0). A total
of 10 µl 7X protease inhibitor cocktail (P8340; Sigma-Aldrich;
Merck Millipore) was mixed with 750 µl lysis buffer, and then 1X
lysis buffer was added to each flask. Cells were removed by a
scrapper and placed on a rotator for 30 min, followed by
centrifugation at 12,000 × g for 20 min at 4°C. The supernatant was
then collected, and protein concentrations were determined using
the Pierce BCA Protein Assay kit (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). To produce a cytosolic fraction, cells were
re-suspended at 4°C in 10 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (1.5 mM
MgCl2, 10 mM KCl, 0.5 mM dithiothreitol and 0.2 mM
phenylmethane sulfonyl fluoride; pH 7.9), placed on ice for 10 min
and vortexed for 10 sec. Samples were centrifuged at 10,000 × g for
2 min at 4°C, and the supernatant containing the cytosolic fraction
was stored at −80°C. Protein concentrations were measured using the
aforementioned Pierce BCA Protein Assay kit. Equal amounts of
protein (30 µg per sample) were separated by 12.5% SDS-PAGE and
transferred to a nitrocellulose membrane. Following blocking with
10% skimmed milk for 1 h, proteins were incubated with rabbit
polyclonal antibodies against Nrf2 (dilution, 1:700) and β-actin
(dilution, 1:5,000) at 4°C overnight. Upon washing three times, the
membranes were further incubated with horseradish
peroxidase-conjugated secondary antibodies (rabbit anti-mouse
immunoglobulin G; dilution, 1:10,000; ab97046; Abcam, Cambridge,
UK) for 2 h at room temperature. Finally, immunoreactive protein
bands were developed using enhanced chemiluminscence (123072;
Sigma-Aldrich; Merck Millipore). Normalization of western blot
analysis was ensured by using β-actin as a loading control. Western
blot quantification was performed using ImageJ software version
1.48 (https://imagej.nih.gov/ij/download.html).
Determination of glutathione
peroxidase (GPx) activity
The procedures defined by Fecondo and Augusteyn
(27), which screen continuous
regeneration of reduced glutathione (GSH) from oxidized glutathione
(GSSG) in the presence of glutathione reductase (GR; Sigma-Aldrich;
Merck Millipore) and disodium (Na2) salt of reduced
nicotinamide adenine dinucleotide phosphate (NADPH; Sigma-Aldrich;
Merck Millipore) were used to determine the GPx activity, with
minor modifications. After culturing cells at a density of
12×105 in a T75 flask for 18–24 h, cancer cells were
treated with VC and Q as mentioned in the preceding paragraph. The
enzyme activity in the clear supernatant of tumor cell lysates was
expressed as µmol of NADPH oxidized/min/mg cell protein, using a
molar extinction coefficient of 6.22×106 M−1
cm−1 for NADPH. The GPx activity is defined as mU/mg of
cell protein.
Determination of GR activity
The activity of GR was assessed by the method
elucidated by Maiani et al (28), with minor modifications. Cancer cells
were cultured at a density of 12×105 cells/T75 flask,
and after 18–24 h of incubation, they were treated with 200 or 100
µM VC for 24 h. Then, 50 or 75 µM Q, respectively, was added for 6
h. The GR assay was performed in a cuvette in a total volume of 1
ml, containing 60 µM buffer, 5 mM EDTA (pH 8.0), 0.033 M GSSG, 2 mM
NADPH and sample. The decrease in absorbance, which represents the
oxidation of NADPH during the reduction of GSSG by the GR present
in the sample, was monitored spectrophotometrically at 340 nm for 3
min. Results were based on a molar extinction coefficient for NADPH
of 6.22×106 M−1 cm−1. The GR
activity was defined as mU/mg cell protein.
Determination of NADPH dehydrogenase
quinone 1 (NQO1) activity
Similarly, to the determination of GR activity,
following the seeding of cancer cells at a density of
12×105 cells/T75 flask and treatment with VC and Q,
cells were washed with FBS and resuspended in 2 ml 25 mM Tris-HCl
buffer (pH 7.4) and 250 mM sucrose (1:1). Then, cells were
sonicated on ice for 10 sec (twice) using a probe sonicator. The
resultant sonicate was centrifuged at 10,000 × g at 4°C for 30 sec
to remove large particles. The activity of NQO1 was determined
spectrophotometrically as the dicoumarol-inhibitable fraction of
the NADH-dependent reduction of dichloroindophenol (DCPIP). DCPIP
was used as an electron acceptor, as it loses color upon reduction.
Briefly, 100 ml extract was placed in an acid-cleaned quartz
cuvette containing 2.7 ml buffer [25 mM Tris-HCl (pH 7.4), 700
mg/ml bovine serum albumin (BSA) (A1933; Sigma-Aldrich; Merck
Millipore)], 100 ml NADH (6 mM) and 100 ml DCPIP (1.2 mM). The
cuvette was rapidly agitated, and the absorbance at 600 nm was
recorded over 2 min using a sample with no enzyme as a reference.
The assay was then repeated with a fresh sample containing 10 ml
dicoumarol inhibitor (10 mM in dimethyl sulfoxide). Dicoumarol
sensitive activity [rate of optical density (OD) change without
inhibitor/rate of OD change with inhibitor] was used to measure
NQO1 activity. The final activities were calibrated against protein
concentration and expressed as nM/min/mg protein. Protein
concentration was determined using the Pierce BCA Protein Assay kit
(Thermo Fisher Scientific, Inc.).
Heme oxygenase 1 (HO1) activity
assay
HO1 activity was measured in microsomal preparations
from cells. Similar to the previous test, following cell culture at
a density of 12×105 cells/T75 flask and treatment with
VC and Q for 30 h, cells were homogenized in 0.5 ml ice-cold 0.25 M
sucrose solution containing 50 mM potassium phosphate buffer (pH
7.4). Homogenates were centrifuged at 200 × g for 10 min. The
supernatants were then centrifuged at 10,000 × g for 20 min, and
further centrifuged at 30,000 × g for 60 min at 4°C. The resultant
pellet was resuspended in 50 mM potassium phosphate buffer (pH 7.4)
and the protein concentration was determined using the
aforementioned Pierce BCA Protein Assay kit. The extract
(containing 40 mM protein) was mixed with 20 µM hemin (H2250;
Sigma-Aldrich; Merck Millipore), 15 mM BSA, 1 mM NADPH, 0.1 M
potassium phosphate buffer (pH 7.4) and 1.5 unit purified
biliverdin reductase (B3687; Sigma-Aldrich; Merck Millipore). After
1 h of incubation at 37°C, the reaction was stopped with 0.6 ml
chloroform. Following the extraction of cells, bilirubin
concentrations in the chloroform cell extracts were determined by
spectrophotometry at an absorbance wavelength of 464–530 nm. HO1
activity was calculated as nM bilirubin/mg protein/min, assuming an
extinction coefficient of 40/(mmol/l)/cm in chloroform.
Determination of GSH
GSH assay using 5,5-dithio-bis-(2-nitrobenzoic acid)
(DTNB) was performed according to the Ellman's method (29). Standard curves were constructed from 1
mM GSH. Following the seeding of cancer cells at a density of
12×105 cells/T75 flask and treatment with VC and Q,
clear supernatant of cell lysate was analyzed for GSH levels. A
total of 2.3 ml potassium phosphate buffer (0.2 M, pH 7.6) was
added to 0.2 ml cell lysate supernatant, and then 0.5 ml DTNB
(0.001 M) was added to the solution. The absorbance was measured 5
min later at 412 nm.
Determination of intracellular
generation of ROS
2′-7′-Dichlorodihydrofluorescein diacetate (DCFH-DA)
fluorescent probes were used to measure the intracellular
generation of hydrogen peroxide (H2O2) and
superoxide anions (O2˙−), respectively (30). These probes are stable nonpolar
compounds that readily diffuse into cells. Once inside the cells,
the acetate groups of DCFH-DA are cleaved from the molecule by
intracellular esterases to yield DCFH, which is trapped within the
cells. Intracellular H2O2 or low-molecular
weight peroxides, oxidize DCFH to dichloride, which is a highly
fluorescent compound. Thus, the fluorescence intensity is
proportional to the quantity of peroxide produced by the cells.
Briefly, solid tumor cells were seeded at a density of
1×104 cells/well in a 96-well plate. Following cell
treatments with VC and Q, the media of each well were removed, and
100 µl 10 µM DCFH-DA was added to the plate, which was then
incubated for 30 min at 37°C in a humidified 5% CO2
atmosphere. Extracellular DCFH-DA was subsequently replaced with
200 µl PBS− (PBS without calcium or magnesium), and the
fluorescence intensity was determined with a fluorimeter, using 480
and 530 nm as the excitation and emission wavelengths,
respectively.
Statistical analysis
Data were collected and expressed as the mean ±
standard error of the mean from three independent experiments.
Statistical analysis was performed by applying one-way analysis of
variance and Tukey's test to compare the control and vehicle groups
against the treated groups. P<0.05 was considered to indicate a
statistically significant difference. The 50% inhibitory
concentration (IC50) values for VC and Q were calculated
using GraphPad Prism software version 5 (GraphPad Software, Inc.,
La Jolla, CA, USA).
Results
Evaluating the cytotoxic effects of VC
and Q against solid tumor cell lines
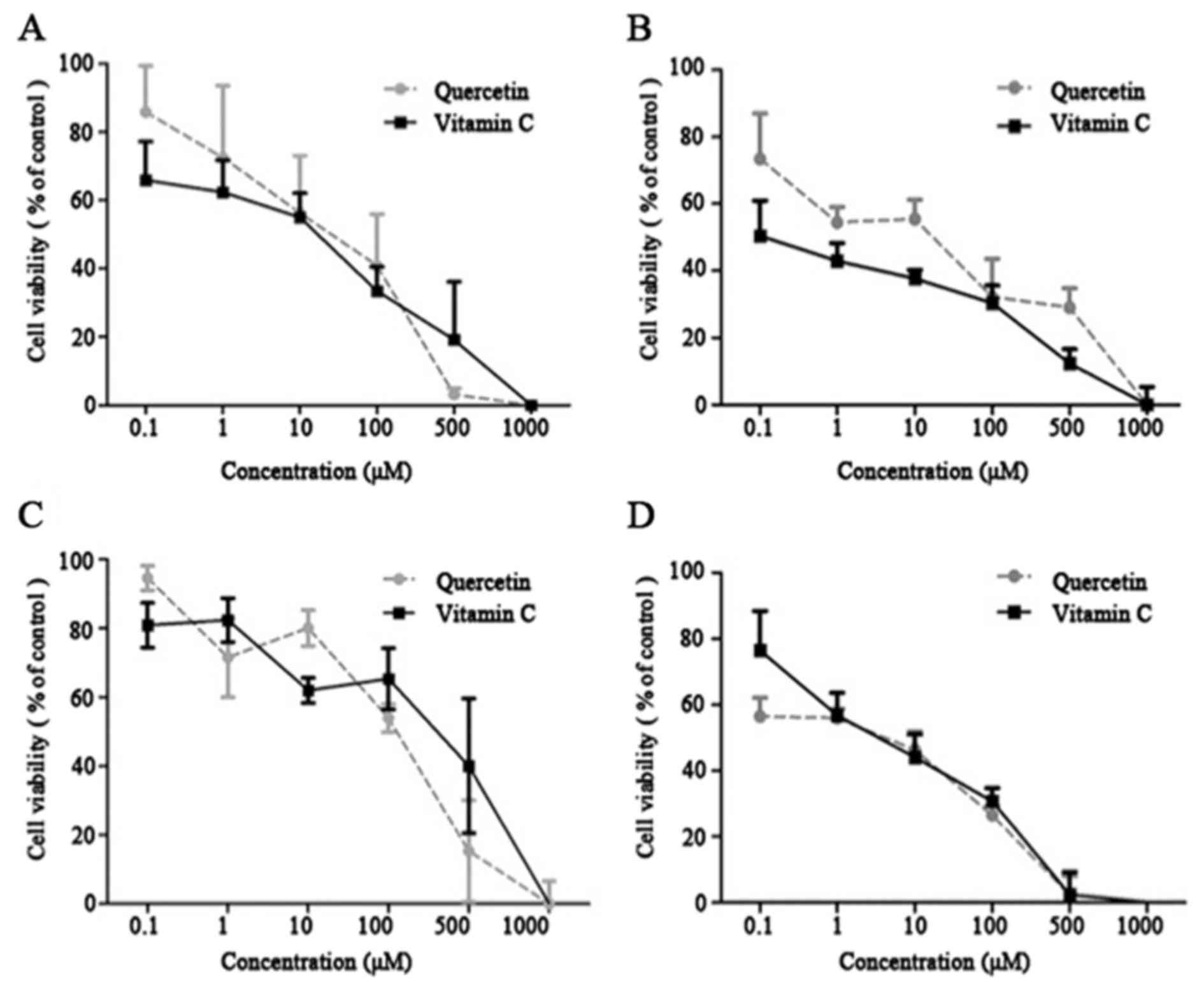
VC exhibited cytotoxicity against the cancer cell
lines, with an IC50 of 271.6–480.1 µM. Q displayed
comparable cytotoxic profiles against the tumor cell lines, with an
IC50 of 155.1–232.9 µM (Table
I). VC and Q exhibited a growth-inhibitory effect in a
dose-dependent manner. Dose-response values were measured (Fig. 1A and D). The results indicated a
concentration-dependent decrease in cell viability following
treatment with VC and Q. VC and Q significantly decreased cell
viability of tumor cells at concentrations >100 µM (Fig. 1) (P=0.045). Based on this observation,
subtoxic concentrations of Q (50 and 75 µM) and VC (100 and 200 µM)
were selected to investigate the effect of Q and VC on Nrf2
signaling.
 | Table I.IC50 values of vitamin C
and quercetin in different tumor cell lines. Data are expressed as
the mean ± standard deviation of three independent experiments
(n=3). |
Table I.
IC50 values of vitamin C
and quercetin in different tumor cell lines. Data are expressed as
the mean ± standard deviation of three independent experiments
(n=3).
| A549 | MCF-7 | MDA-MB 468 | MDA-MB 231 |
IC50 |
|---|
| 232.90±17.75 | 155.10±33.80 | 183.20±22.50 | 196.70±40.90 | Quercetin (µM) |
| 480.10±25.05 | 271.60±31.40 | 365.90±24.95 | 382.10±8.69 | Vitamin C (µM) |
Roles of VC and Q on Nrf2
expression
In a preliminary study, MDA-MB 231 cells were seeded
and cultured for 24 h prior to treatment with various
concentrations of VC (50, 100 and 200 µM) or Q (25, 50 and 75 µM),
individually or in combination. With sequential treatment, the most
significant decrease in Nrf2 mRNA expression was observed following
the incubation of cells with 200 µM VC in combination with 50 µM Q,
or 100 µM VC with 75 µM Q (data not shown). Therefore, the same
combinations of VC and Q were applied for subsequent treatments of
the other cancer cell lines. The results indicated that incubating
breast cancer cells with VC and Q induced a marked decrease in the
mRNA and protein expression of Nrf2 in a dose-dependent manner
(Figs. 2 and 3). Despite an aggressive genotype, MDA-MB
231 cells exhibited a greater reduction in the mRNA and nuclear
fraction of Nrf2 than other cells, suggesting that MDA-MB 231 cells
with higher Nrf2 levels are more sensitive to the suppressive
effect of VC and Q than cells with lower levels of Nrf2 (P=0.024
for all cell lines). Treatment with VC and Q decreased nuclear Nrf2
levels (Fig. 2B) and the
nuclear/cytosolic Nrf2 ratio (Fig.
3C) in all cell lines. The nuclear/cytosolic Nrf2 ratio
decreased by 1.7-fold in MDA-MB 231 cells, 2-fold in MDA-MB 468
cells, 1.4-fold in MCF-7 cells and 1.2-fold in A549 cells following
treatment of tumor cells with 200 µM VC and 50 µM Q. This ratio was
much lower in cells treated with 100 µM VC and 75 µM Q: 3.4-fold in
MDA-MB 231 cells, 6-fold in MDA-MB 468 cells, 3.1-fold in MCF-7
cells and 1.2-fold in A549 cells (P=0.027 for breast cancer cell
lines and P=0.505 for A549 cells). These results suggest that 100
and 200 µM VC, as well as 50 and 75 µM Q, have a prominent effect
on the modulation of Nrf2 expression in tumor cells.
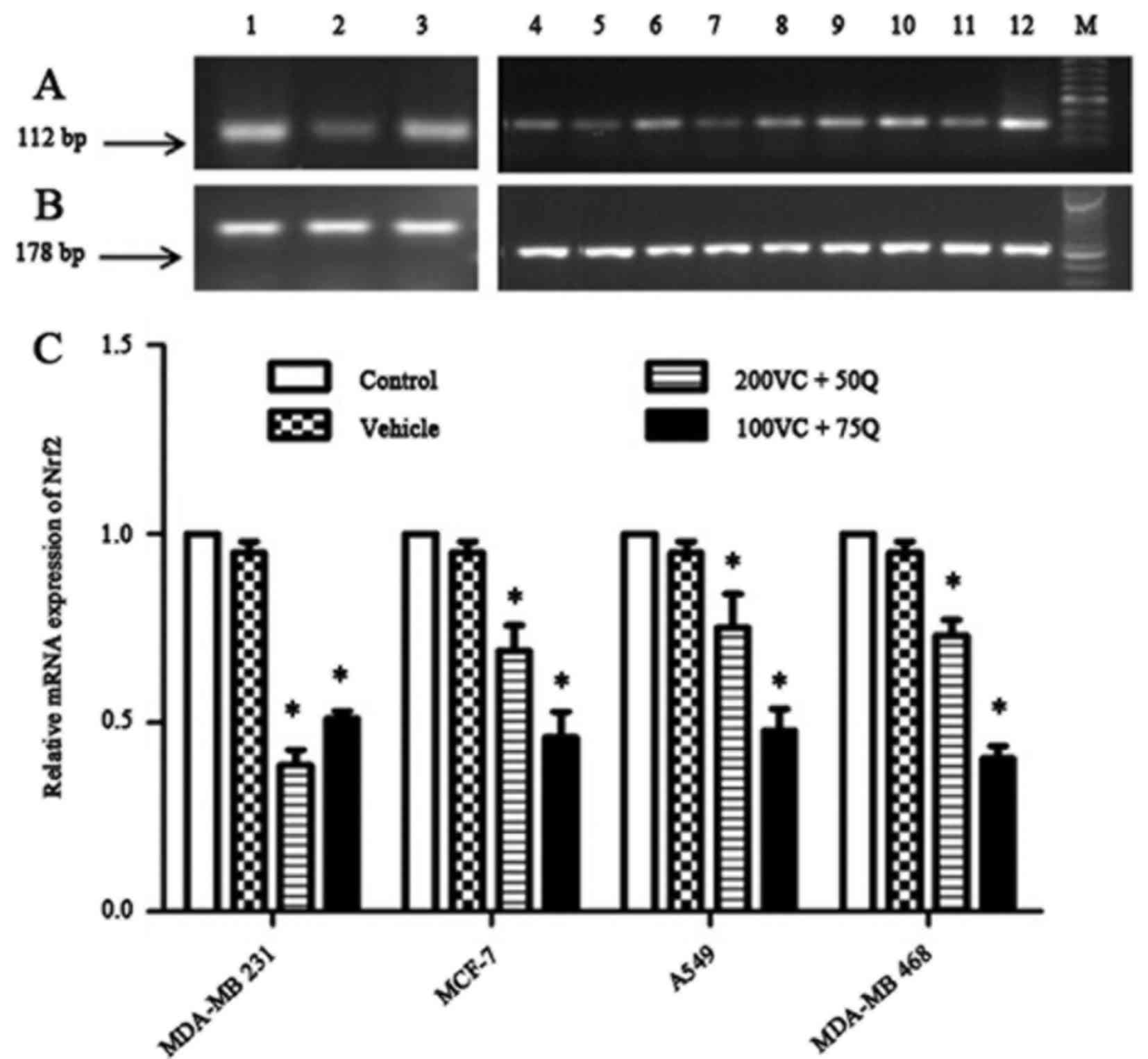 | Figure 2.Dose-dependent effects of VC and Q on
Nrf2 mRNA expression following 30 h of sequential treatment on
tumor cell lines. (A) Bands represent the results from RT-qPCR for
Nrf2: Lanes 1, 2 and 3 correspond to different treatments in MDA-MB
231 cells, which consist of 200 µM VC and 50 µM Q, 100 µM VC and 75
µM Q, and control, respectively; lanes 4, 5 and 6 correspond to
MCF-7 subjected to the same treatment described in lanes 1–3; lanes
7, 8 and 9 correspond to A549 cells subjected to the same treatment
described in lanes 1–3; and lanes 10, 11 and 12 correspond to
MDA-MB 468 cells subjected to the same treatment described in lanes
1–3. (B) Bands represent the results from RT-qPCR results for
β-actin. (C) Results of RT-qPCR with the same concentrations of VC
and Q as the ones mentioned above in solid tumor cell lines.
Percentage data of Nrf2 mRNA expression in various cell lines
following the aforementioned treatments relative to the controls.
Data are presented as the mean ± standard error of the mean, n=6.
P=0.024 for all cell lines; *P<0.05 compared with the control
group and vehicle. RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; VC, vitamin C; Q, quercetin; Nrf2,
nuclear factor erythroid 2-related factor 2; mRNA, messenger RNA;
M, marker. |
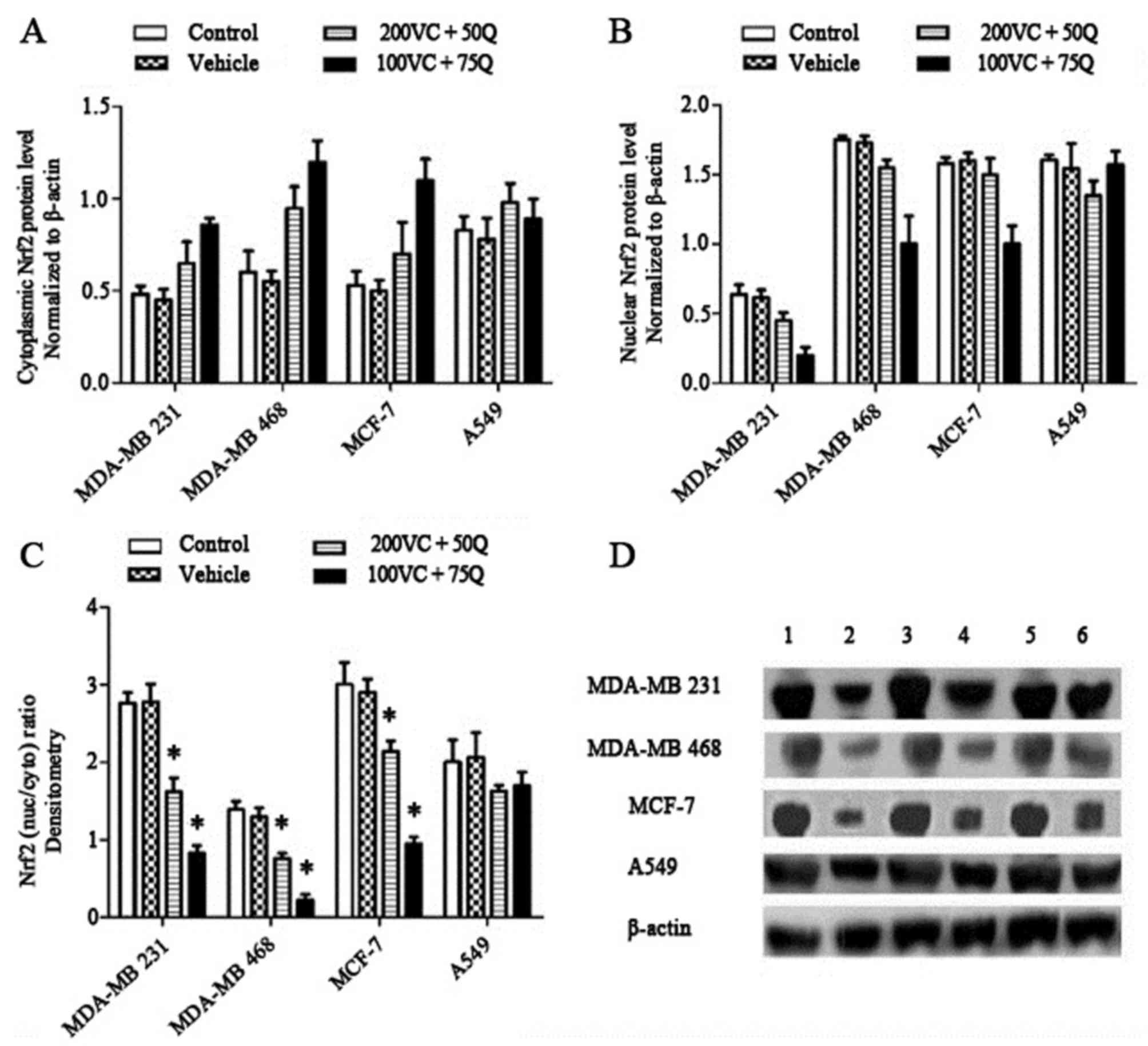 | Figure 3.Dose-dependent effects of VC and Q on
the levels of Nrf2 protein following 30 h of sequential treatment
of tumor cell lines. Nrf2 levels were measured in the whole cell
lysate and cytoplasmic fraction, and Nrf2 levels in nuclear
fractions were obtained by subtracting the cytoplasmic fraction
levels from the whole cell lysate levels. (A) Percentage values of
cytosolic levels of Nrf2 relative to the controls. (B) Percentage
values of nuclear levels of Nrf2 relative to the controls. (C)
Nuclear/cytosolic Nrf2 ratio of bands densitometric quantification.
Normalization of western blotting results was ensured by β-actin.
(D) Bands of representative experiments: Lane 1, whole cell lysate
without treatment; lane 2, cytoplasmic lysate without treatment;
lane 3, whole cell lysate with 200 µM VC and 50 µM Q; lane 4,
cytoplasmic lysate with 200 µm VC and 50 µM Q; lane 5, whole cell
lysate with 100 µM VC and 75 µM Q; lane 6, cytoplasmic lysate with
100 µM VC and 75 µM Q (mean ± standard error of the mean, n=3).
P=0.027 for breast cancer cell lines and P=0.505 for A549 cells;
*P<0.05 compared with the control group and vehicle. Nrf2,
nuclear factor erythroid 2-related factor 2; VC, vitamin C; Q,
quercetin; nuc/cyto, nuclear/cytoplasmic Nrf2 ratio. |
Effects of sequential treatment with
VC and Q on xenobiotic metabolizing enzymes and thiol content
To investigate how treatment with VC and Q affects
Nrf2-regulated genes, the levels of xenobiotic metabolizing enzymes
and thiol content in solid tumor cells were determined. There were
no significant changes in GPx or GR activities in MDA-MB 231, MCF7
or A549 cells following exposure of cells to sequential treatment
of VC and Q (Fig. 4A and B). However,
both these parameters were significantly decreased in MDA-MB-468
cells (P=0.027), indicating a significant reduction in the level of
antioxidant enzymes. In the MDA-MB 231 and MCF-7 cell lines, HO1
was significantly suppressed following treatment with VC and Q
(Fig. 4C). Additionally, NQO1
activity was significantly decreased in all treated cells (Fig. 4D). The most prominent changes were
observed in MDA-MB 231 cells, suggesting that this cell line, which
has higher levels of Nrf2 expression, is more sensitive to the
suppressive effects of VC and Q than the other cell lines
evaluated.
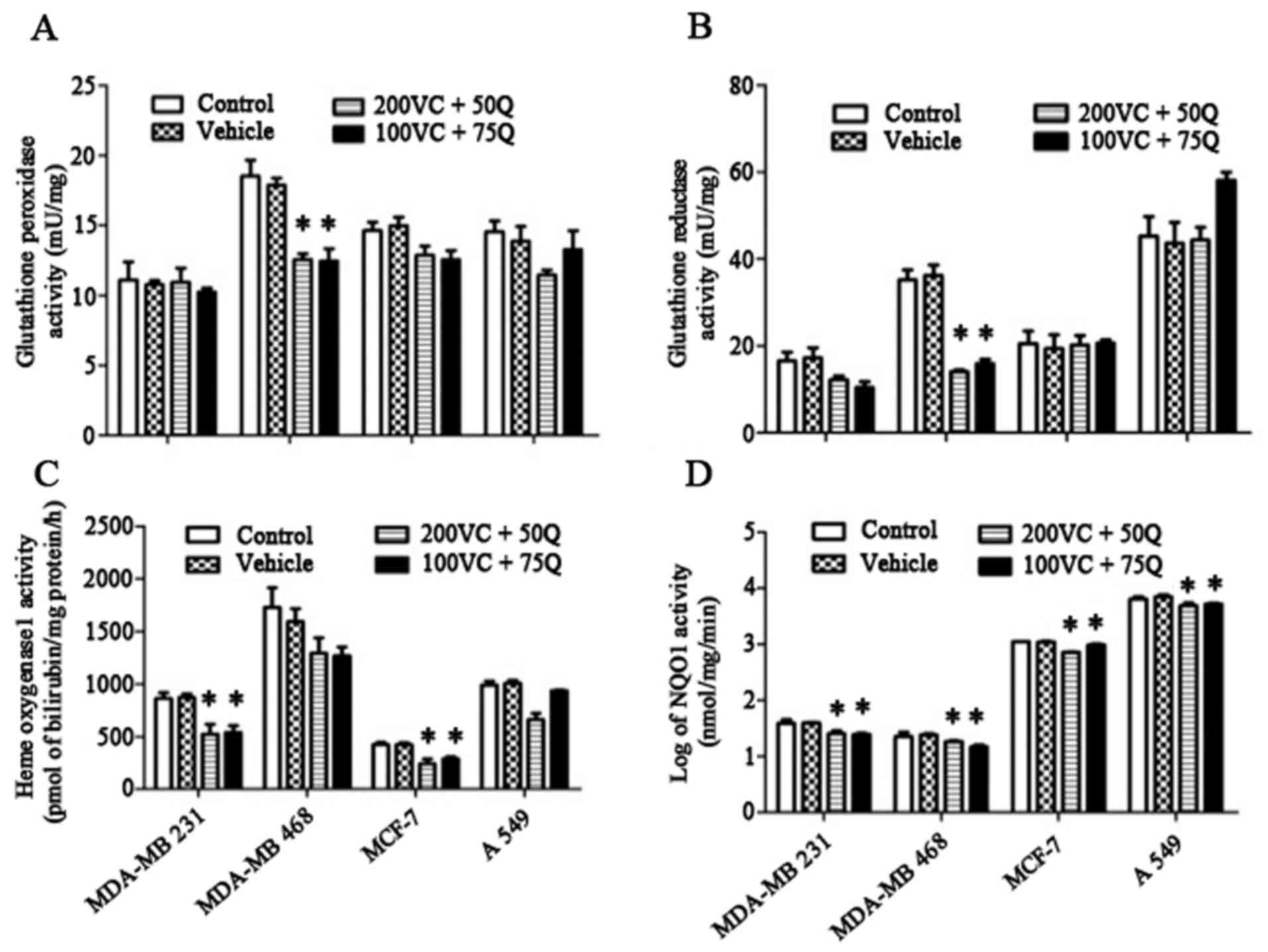 | Figure 4.Effects of VC and Q treatment on the
level of (A) GPx, (B) GR (C) HO1 and (D) NQO1 activities. Tumor
cells were incubated with 200 µM VC and 50 µM Q, or with 100 µM VC
and 75 µM Q for 30 h. Values are means of three different samples
per condition. Data are represented as the mean ± standard error of
the mean, n=3. P=0.05 for MDA-MB 231 and A549 cells, and P=0.027
for MDA-MB 468 and MCF-7 cells for NQO1 activity; P=0.027 for HO1
activity in MDA-MB 231 and MCF-7 cells; P=0.027 for GPx and GR
activity in MDA-MB 468 cells; *P<0.05 compared with the control
group and vehicle. VC, vitamin C; Q, quercetin; GPx, glutathione
peroxidase; GR, glutathione reductase; HO1, heme oxygenase 1; NQO1,
nicotinamide adenine dinucleotide phosphate dehydrogenase quinone
1. |
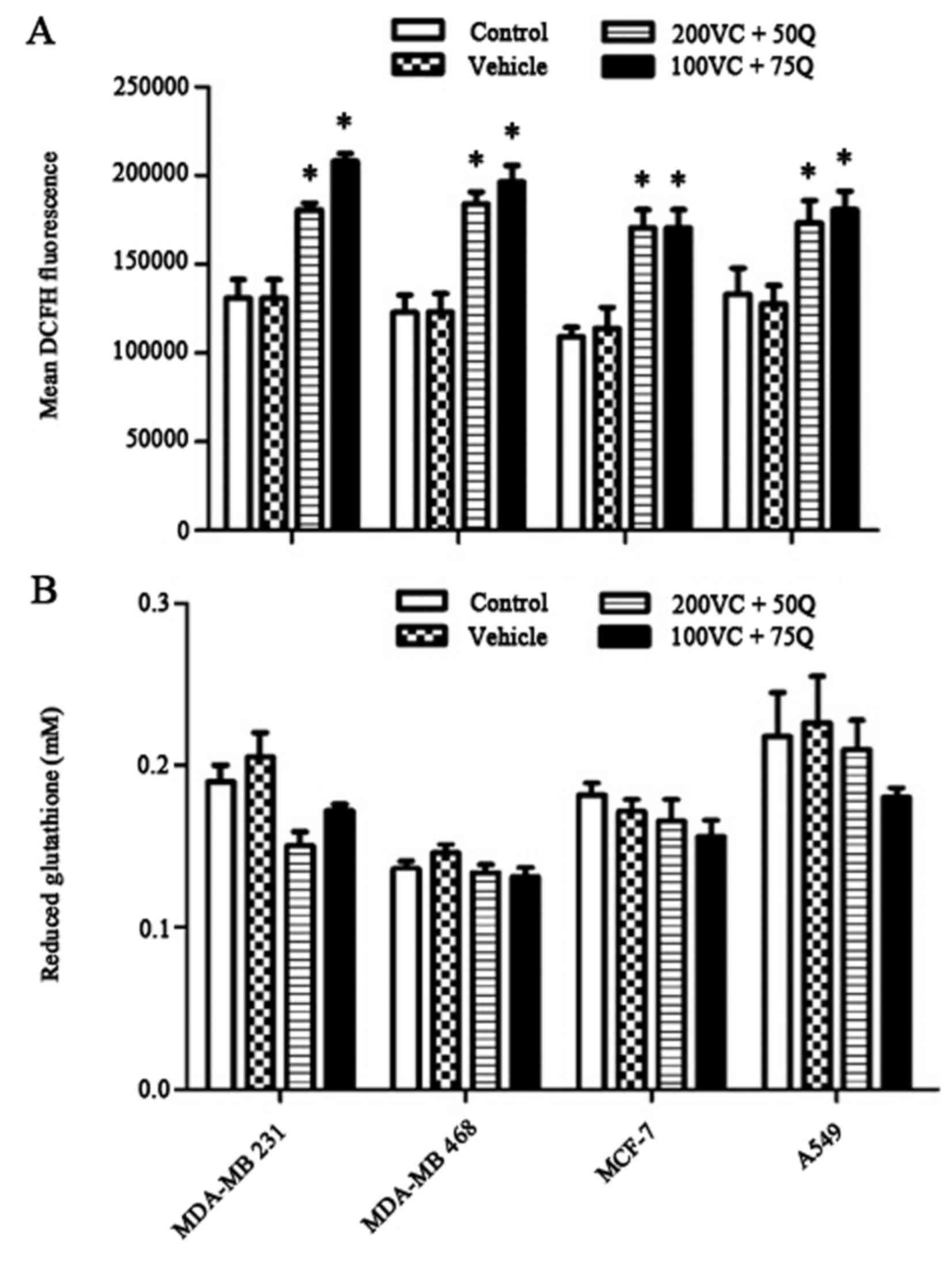
Inhibitory effect of sequential
treatment with VC and Q on intracellular ROS levels
The baseline DCF florescence measurement indicated
that 30 h of sequential treatment with VC and Q significantly
decreased endogenous ROS levels in a dose-dependent manner
(Fig. 5A). By contrast, sequential
treatment of cells with VC and Q did not modulate cellular thiol
levels in tumor cells (Fig. 5B).
Discussion
Identifying novel and potential molecular targets
for cancer therapy is the goal of current studies that aim to
decrease the side effects of chemotherapy agents and overcome
chemo-resistance. Nrf2 and downstream target genes serve a pivotal
role in cellular redox homeostasis, elimination of ROS and
xenobiotic metabolism (5). Persistent
Nrf2-mediated antioxidant responses promote malignant progression,
development of acquired apoptotic resistance and chemo-resistance
in cancer cells (30,31). Therefore, the identification of
stable, safe and potent Nrf2 inhibitors to decrease the antioxidant
response and reduce drug metabolism in tumor cells is urgently
required. In a pilot study in our laboratory (Recombinant Protein
Laboratory, School of Medicine, Shiraz University of Medical
Sciences), it was demonstrated that treatment of MDA-MB 231 cells
with 25 µM Q increased the expression of Nrf2, while 50 and 75 µM Q
decreased the mRNA levels of Nrf2. In addition, suppression of Nrf2
mRNA was detected when the cells were treated with 50–400 µM VC
(data not shown).
It has been reported that treatment of HepG2 cells
with 50 µM Q inhibited Nrf2 activation by decreasing the nuclear
translocation of Nrf2 and the nuclear content of phosphorylated
Nrf2 (24). Furthermore, changes in
the redox state caused by antioxidants such as VC inhibited
Nrf2-mediated gene expression and overcame resistance to imatinib
(16). Therefore, the doses of VC and
Q used in the current study were pharmacological and selected
according to their low levels of toxicity, as well as their
efficacy at inhibiting Nrf2 expression. The results of the present
study demonstrated that treatment with Q, a tumor-active
phytochemical, and VC, an antioxidant agent with antineoplastic
activity, resulted in a significant decrease in Nrf2 expression and
induced oxidative stress in cancer cells. Sequential treatment of
solid tumor cells with VC and Q reduced the mRNA and protein levels
of Nrf2. Suppression of Nrf2 protein expression was notable, and
the overall response indicated that the aforementioned treatment
decreased Nrf2 mRNA and protein expression, as well as reducing the
stability of Nrf2 protein, leading to the suppression of Nrf2.
In various types of cancer cells, Q inhibits cell
growth and induces apoptosis; however, it also induces the
expression of antioxidant proteins involved in the elimination of
ROS, thus protecting cells against oxidative damage (32,33).
Flavonoids, including VC and Q, have emerged as an effective
adjuvant in cancer therapy, due to acting as free radical
scavengers and immune system modulators, in addition to exhibiting
antioxidant properties (34). It has
been demonstrated that treatment of MSTO-211H lung cancer cells
with 20 µM Q stimulated an increase in the levels of cellular Nrf2,
upregulation of Nrf2 mRNA and protein and an increase in the
affinity of Nrf2 for binding to ARE-driven reporter sequences,
consequently boosting the expression of downstream genes compared
with untreated cells (35). However,
at doses of Q ≥60 µM, the levels of Nrf2 protein were not affected
(35). It has been suggested that Q
may act as a ‘double-edged sword’ due to its unique properties,
since it behaves as an antioxidant and/or pro-oxidant depending on
its concentration and the duration of exposure (35). Treatment of HepG2 cells with 50 µM Q
induced activation of p38 following 4 h of treatment. By contrast,
following18 h of incubation, the level of p38 expression detected
was similar to that of the control cells. Nrf2 expression was
inhibited at both incubation times, and Q (50 µM) induced a
time-dependent activation of p38, in parallel with a transient
stimulation of Nrf2, provoking its inhibition later (24).
VC has been employed as an adjuvant for the
treatment of cancer patients, as it acts as pro-oxidant by
generating ascorbate radicals and hydrogen peroxide against the
growth of tumor cells but not against that of normal cells
(36). It has been indicated that VC
significantly inhibits tumor growth in Lewis lung carcinoma
(LLC)-bearing mice at low and high doses (37). Addition of 0.125 mM ascorbic acid to
KCL22/SR cells markedly reduced their peroxide levels and inhibited
the formation of the Nrf2/DNA complex in KCL22/SR cells, without
any changes in the level of Nrf2 protein in the total cell lysate,
suggesting that ascorbic acid represses the translocation of Nrf2
into the nucleus (16). Since
oxidative stress stimulated the translocation of Nrf2 into the
nucleus, a shift in intracellular redox balance towards a reduced
condition may hinder the movement of Nrf2 in KCL22/SR cells. The
results of the current study demonstrated that HO1 activity was
reduced in MDA-MB 231 and MCF-7 cells, and that the activity of
NQO1 diminished significantly in all tumor cell lines following
treatment with VC and Q. To maintain homeostasis during oxidative
stress, cells enhance the GSH concentration and upregulate
glutathione-related enzymes to prevent potential oxidative insults
and suppress oxidative-stress induced injuries (38,39). NQO1
and HO-1 are two major downstream targets of Nrf2, and serve a
pivotal role in the maintenance of cellular redox homeostasis, thus
preventing the transformation of normal cells to precancerous or
malignant ones by counteracting ROS-mediated carcinogenesis
(40). However, it was demonstrated
that NQO1, in parallel with Nrf2 overexpression, aberrantly
elevated the levels of HO1 in different types of cancer (40,41).
Minaei et al demonstrated the effectiveness of nano-Qin
decreasing the levels of NQO1 and multidrug resistance-associated
protein 1without altering Nrf2 expression (41). Ren et al reported that Q
decreased the half-life of Nrf2 by means of ubiquitination systems,
which led to a reduction in the gene expression levels. It was also
demonstrated that treatment of human keratinocytes with 50 µM VC
had no effect on NQO1 or HO1 activities (42). Furthermore, it was identified that
administering injections of VC to tumor cells increased the
carbonyl levels in the liver, but reduced the GSH/GSSG ratio in the
liver and kidney (43). Therefore, it
was suggested that high-dose VC has a bifunctional role: i)
Pro-oxidant activity against tumor growth; and ii) antioxidant
activity against oxidative stress and nephrotoxicity induced by
cisplatin in LLC-bearing mice (37).
It was demonstrated that treatment of human keratinocytes with 50
µM VC did not affect NQO1 or HO1 activity (43). Additionally, it has been demonstrated
that treatment of HepG2 cells with Q for 4 h enhanced the activity
of GPx and GR, and increased GSH levels as well as GCS expression,
therefore suggesting that Q can mediate the expression of
GSH-related enzymes (44). This may
be associated with the fact that Nrf2 activation is a master
regulator upstream the GPx and GR genes. Similarly, following acute
stress, GSH levels may be temporarily suppressed and subsequently
recovered, due to an increase in GCS activity and mRNA levels. GSH
levels therefore may be a signal of cellular self-protection
against a sub-lethal toxic insult. High levels of Q, which induce
toxicity, overcome the defense mechanisms of the cell, such as the
Nrf2 response (45). By contrast, the
results of the current study suggest that unchanged levels of GPx,
GR and GSH may imply impairment in the machinery involved in the
gene transcription and mRNA synthesis of antioxidant enzymes. It
was demonstrated that ROS levels significantly increased in tumor
cells treated with VC and Q. The capability of Q to reduce the
levels of accumulated intracellular ROS indicated that the
protective effects of flavonoids are not only limited to their
antioxidant properties, but they can also act as ROS scavengers in
the extracellular medium (46).
Treatment with 1 or 5 mM ascorbate increased the levels of
intracellular ROS in ReN cells but not in mesothelium cells.
Additionally, it was demonstrated that malignant mesothelioma cells
enhanced superoxide production and induced overexpression of the
superoxide-producing NADPH oxidase 4 (47). This discrepancy between data collected
in the current study and previous studies may be due to different
doses of agents used, duration of treatments and the sequential
treatments that were employed.
The results of the present study indicate that
targeting Nrf2 may be a promising strategy to induce oxidative
stress, which in turn represents a potential efficient method of
sensitizing tumor cells. Furthermore, Nrf2 may be a determining
factor for inhibition in chemotherapy protocols. The sequential
treatment of solid tumor cells with VC and Q reduced the expression
of Nrf2 at the mRNA and protein levels. Therefore, further studies
to establish the efficacy and safety of antioxidant adjuvants in
vivo and in humans are required to establish evidence-based
guidelines on their use in cancer therapy, in order to obtain
optimal therapeutic outcomes in patients with cancer.
Acknowledgements
The present study was performed both as a part of a
PhD student thesis by Miss Fatemeh Ramezani (Department of
Biochemistry, School of Medicine, Shiraz University of Medical
Sciences, Shiraz, Iran) and an MSc student thesis by Mrs. Fatemeh
Keshavarzi (Department of Biochemistry, Recombinant Protein
Laboratory, School of Medicine, Shiraz University of Medical
Sciences), who received grants (grant nos. 92–6659 and 94–7441,
respectively) from the office of Vice Chancellor for Research and
the Committee for Advanced Biomedical Sciences, Shiraz University
of Medical Sciences.
References
|
1
|
Ramos S: Effects of dietary flavonoids on
apoptotic pathways related to cancer chemoprevention. J Nutr
Biochem. 18:427–442. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
No JH, Kim YB and Song YS: Targeting nrf2
signaling to combat chemoresistance. J Cancer Prev. 19:111–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wu T, Harder BG, Wong PK, Lang JE and
Zhang DD: Oxidative stress, mammospheres and Nrf2-new implication
for breast cancer therapy? Mol Carcinog. 54:1494–1502. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jaramillo MC and Zhang DD: The emerging
role of the Nrf2-Keap1 signaling pathway in cancer. Genes Dev.
27:2179–2191. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kensler TW, Wakabayashi N and Biswal S:
Cell survival responses to environmental stresses via the
Keap1-Nrf2-ARE pathway. Annu Rev Pharmacol Toxicol. 47:89–116.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Foygel K, Sekar TV and Paulmurugan R:
Monitoring the antioxidant mediated chemosensitization and
ARE-signaling in triple negative breast cancer therapy. PloS One.
10:e01419132015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang XJ, Sun Z, Villeneuve NF, Zhang S,
Zhao F, Li Y, Chen W, Yi X, Zheng W, Wondrak GT, et al: Nrf2
enhances resistance of cancer cells to chemotherapeutic drugs, the
dark side of Nrf2. Carcinogenesis. 29:1235–1243. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Amadori D, Frassineti G, Zoli W, Milandri
C, Serra P, Tienghi A, Ravaioli A, Gentile A and Salzano E:
Doxorubicin and paclitaxel (sequential combination) in the
treatment of advanced breast cancer. Oncology (Williston Park).
11:(4 Suppl 3) 30–33. 1997.PubMed/NCBI
|
|
9
|
Danesi R, Conte PF and Del Tacca M:
Pharmacokinetic optimisation of treatment schedules for
anthracyclines and paclitaxel in patients with cancer. Clin
Pharmacokinet. 37:195–211. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ibrahim T, Fabbri M, Frassineti GL, Zoli
W, Monti M, Ricotti L and Amadori D: Doxorubicin, paclitaxel and
gemcitabine: A Phase I study of a new sequential treatment in stage
III B-IV breast cancer. J Chemother. 15:488–494. 2003.PubMed/NCBI
|
|
11
|
Chou TC: Theoretical basis, experimental
design, and computerized simulation of synergism and antagonism in
drug combination studies. Pharmacol Rev. 58:621–681. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee KW, Lee HJ, Surh YJ and Lee CY:
Vitamin C and cancer chemoprevention: Reappraisal. Am J Clin Nutr.
78:1074–1078. 2003.PubMed/NCBI
|
|
13
|
Zielinski CC: Gemcitabine, anthracycline,
and taxane combinations for advanced breast cancer. Oncology
(Williston Park). 17:(12 Suppl 14) 36–40. 2003.PubMed/NCBI
|
|
14
|
Chao MW, Lai MJ, Liou JP, Chang YL, Wang
JC, Pan SL and Teng CM: The synergic effect of vincristine and
vorinostat in leukemia in vitro and in vivo. J Hematol Oncol.
8:822015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Weiss RB, Woolf SH, Demakos E, Holland JF,
Berry DA, Falkson G, Cirrincione CT, Robbins A, Bothun S, Henderson
IC, et al: Natural history of more than 20 years of node-positive
primary breast carcinoma treated with cyclophosphamide,
methotrexate, and fluorouracil-based adjuvant chemotherapy: A study
by the cancer and leukemia group B. J Clin Oncol. 21:1825–1835.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tarumoto T, Nagai T, Ohmine K, Miyoshi T,
Nakamura M, Kondo T, Mitsugi K, Nakano S, Muroi K, Komatsu N and
Ozawa K: Ascorbic acid restores sensitivity to imatinib via
suppression of Nrf2-dependent gene expression in the
imatinib-resistant cell line. Exp Hematol. 32:375–381. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Khanduja KL, Gandhi RK, Pathania V and
Syal N: Prevention of N-nitrosodiethylamine-induced lung
tumorigenesis by ellagic acid and quercetin in mice. Food Chem
Toxicol. 37:313–318. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saw CL, Guo Y, Yang AY, Paredes-Gonzalez
X, Ramirez C, Pung D and Kong AN: The berry constituents quercetin,
kaempferol, and pterostilbene synergistically attenuate reactive
oxygen species: Involvement of the Nrf2-ARE signaling pathway. Food
Chem Toxicol. 72:303–311. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Borska S, Chmielewska M, Wysocka T,
Drag-Zalesinska M, Zabel M and Dziegiel P: In vitro effect of
quercetin on human gastric carcinoma: Targeting cancer cells death
and MDR. Food Chem Toxicol. 50:3375–3383. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Granado-Serrano AB, Martín MA, Bravo L,
Goya L and Ramos S: Quercetin induces apoptosis via caspase
activation, regulation of Bcl-2, and inhibition of PI-3-kinase/Akt
and ERK pathways in a human hepatoma cell line (HepG2). J Nutr.
136:2715–2721. 2006.PubMed/NCBI
|
|
21
|
Li N, Sun C, Zhou B, Xing H, Ma D, Chen G
and Weng D: Low concentration of quercetin antagonizes the
cytotoxic effects of anti-neoplastic drugs in ovarian cancer. PloS
One. 9:e1003142014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Robaszkiewicz A, Balcerczyk A and Bartosz
G: Antioxidative and prooxidative effects of quercetin on A549
cells. Cell Biol Int. 31:1245–1250. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Samuel T, Fadlalla K, Mosley L, Katkoori
V, Turner T and Manne U: Dual-mode interaction between quercetin
and DNA-damaging drugs in cancer cells. Anticancer Res. 32:61–71.
2012.PubMed/NCBI
|
|
24
|
Granado-Serrano AB, Martín MA, Bravo L,
Goya L and Ramos S: Quercetin modulates Nrf2 and
glutathione-related defenses in HepG2 cells: Involvement of p38.
Chem Biol Interact. 195:154–164. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pfaffl MW: Quantification strategies in
real-time PCR. A-Z of quantitative PCR. 3:87–112. 2004.
|
|
26
|
Sabzichi M, Samadi N, Mohammadian J,
Hamishehkar H, Akbarzadeh M and Molavi O: Sustained release of
melatonin: A novel approach in elevating efficacy of tamoxifen in
breast cancer treatment. Colloids Surf B Biointerfaces. 145:64–71.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fecondo JV and Augusteyn RC: Superoxide
dismutase, catalase and glutathione peroxidase in the human
cataractous lens. Exp Eye Res. 36:15–23. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Maiani G, Mobarhan S, Nicastro A, Virgili
F, Scaccini C and Ferro-Luzzi A: Determination of glutathione
reductase activity in erythrocytes and whole blood as an indicator
of riboflavin nutrition. Acta Vitaminol Enzymol. 5:171–178.
1982.(In Italian).
|
|
29
|
Ellman GL: Tissue sulfhydryl groups. Arch
Biochem Biophys. 82:70–77. 1959. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Furfaro A, Traverso N, Domenicotti C,
Piras S, Moretta L, Marinari UM, Pronzato MA and Nitti M: The
Nrf2/HO-1 axis in cancer cell growth and chemoresistance. Oxid Med
Cell Longev. 2016:19581742016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
van der Wijst MG, Brown R and Rots MG:
Nrf2, the master redox switch: The Achilles' heel of ovarian
cancer? Biochim Biophys Acta. 1846:494–509. 2014.PubMed/NCBI
|
|
32
|
Liu KC, Yen CY, Wu RS, Yang JS, Lu HF, Lu
KW, Lo C, Chen HY, Tang NY, Wu CC and Chung JG: The roles of
endoplasmic reticulum stress and mitochondrial apoptotic signaling
pathway in quercetin-mediated cell death of human prostate cancer
PC-3 cells. Environ Toxicol. 29:428–439. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee WJ, Hsiao M, Chang JL, Yang SF, Tseng
TH, Cheng CW, Chow JM, Lin KH, Lin YW, Liu CC, et al: Quercetin
induces mitochondrial-derived apoptosis via reactive oxygen
species-mediated ERK activation in HL-60 leukemia cells and
xenograft. Arch Toxicol. 89:1103–1117. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bournival J, Francoeur MA, Renaud J and
Martinoli MG: Quercetin and sesamin protect neuronal PC12 cells
from high-glucose-induced oxidation, nitrosative stress, and
apoptosis. Rejuvenation Res. 15:322–333. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bouayed J and Bohn T: Exogenous
antioxidants-double-edged swords in cellular redox state: Health
beneficial effects at physiologic doses versus deleterious effects
at high doses. Oxid Med Cell Longev. 3:228–237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Carr AC, Vissers MC and Cook JS: The
effect of intravenous vitamin C on cancer-and chemotherapy-related
fatigue and quality of life. Front Oncol. 4:2832014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen MF, Yang CM, Su CM and Hu ML: Vitamin
C protects against cisplatin-induced nephrotoxicity and damage
without reducing its effectiveness in C57BL/6 mice xenografted with
Lewis lung carcinoma. Nutr Cancer. 66:1085–1091. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang X, Ye XL Liu R, Chen HL, Bai H, Liang
X, Zhang XD, Wang Z, Li WL and Hai CX: Antioxidant activities of
oleanolic acid in vitro: Possible role of Nrf2 and MAP kinases.
Chem Biol Interact. 184:328–337. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ding M, Zhao J, Bowman L, Lu Y and Shi X:
Inhibition of AP-1 and MAPK signaling and activation of Nrf2/ARE
pathway by quercitrin. Int J Oncol. 36:59–67. 2010.PubMed/NCBI
|
|
40
|
Chun KS, Kundu J, Kundu JK and Surh YJ:
Targeting Nrf2-Keap1 signaling for chemoprevention of skin
carcinogenesis with bioactive phytochemicals. Toxicol Lett.
229:73–84. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Minaei A, Sabzichi M, Ramezani F,
Hamishehkar H and Samadi N: Co-delivery with nano-quercetin
enhances doxorubicin-mediated cytotoxicity against MCF-7 cells. Mol
Biol Rep. 43:99–105. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ren D, Villeneuve NF, Jiang T, Wu T, Lau
A, Toppin HA and Zhang DD: Brusatol enhances the efficacy of
chemotherapy by inhibiting the Nrf2-mediated defense mechanism.
Proc Natl Acad Sci USA. 108:1433–1438. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wagner AE, Ernst I, Iori R, Desel C and
Rimbach G: Sulforaphane but not ascorbigen, indole-3-carbinole and
ascorbic acid activates the transcription factor Nrf2 and induces
phase-2 and antioxidant enzymes in human keratinocytes in culture.
Exp Dermatol. 19:137–144. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Scharf G, Prustomersky S, Knasmuller S,
Schulte-Hermann R and Huber WW: Enhancement of glutathione and
g-glutamylcysteine synthetase, the rate limiting enzyme of
glutathione synthesis, by chemoprotective plant-derived food and
beverage components in the human hepatoma cell line HepG2. Nutr
Cancer. 45:74–83. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Goldring CE, Kitteringham NR, Elsby R,
Randle LE, Clement YN, Williams DP, McMahon M, Hayes JD, Itoh K,
Yamamoto M and Park BK: Activation of hepatic Nrf2 in vivo by
acetaminophen in CD-1 mice. Hepatology. 39:1267–1276. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hanneken A, Lin FF, Johnson J and Maher P:
Flavonoids protect human retinal pigment epithelial cells from
oxidative-stress-induced death. Invest Ophthalmol Vis Sci.
47:3164–3177. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Parrow NL, Leshin JA and Levine M:
Parenteral ascorbate as a cancer therapeutic: A reassessment based
on pharmacokinetics. Antioxid Redox Signal. 19:2141–2156. 2013.
View Article : Google Scholar : PubMed/NCBI
|