Introduction
Colorectal cancer (CRC) is currently the third most
common cancer in males and the second in females worldwide, with an
estimated 93,090 novel cases and 49,700 mortalities in the USA in
2015 (1,2). The morbidity of CRC varies markedly
worldwide, mainly due to differences in lifestyle, environment and
genetics. Epidemiological studies have demonstrated that a number
of risk factors contribute to the initiation and progression of
CRC, including advanced age, hereditary components, obesity, excess
alcohol and red meat, smoking and a lack of physical exercise
(3–6).
Over the last decades, marked progress has been achieved in surgery
resection, radiotherapy and chemotherapy (7). However, the clinical outcome and
prognosis of remains poor for patients with CRC with metastasis or
recurrence (8,9). In total ~20% of patients with CRC were
identified to already bear distant metastases. Furthermore, >50%
patients eventually develop recurrent disease and metastasis
following surgery with curative intent (10,11). The
5-year overall survival rate for patients with CRC is ~90%, but
decreases to <5% in cases of CRC with metastases or recurrence
(12). Therefore, it is crucial to
understand the molecular mechanisms underlying CRC progression
which may provide a potential therapeutic target for the treatment
of CRC.
Previous studies have demonstrated that microRNAs
(miRNAs/miRs) are deregulated in a number of human cancers,
including CRC (13–15). miRNAs are a class of single-stranded
non-coding endogenous small RNA molecules of between 19 and 24
nucleotides in length (16). They
negatively regulate gene expression through binding to the 3′
untranslated regions (3′UTRs) of target mRNAs and inducing
degradation or translational repression of target gene mRNA
(16,17). miRNAs have been identified to be
involved in a wide range of physiological and pathological
processes, including cell proliferation, apoptosis, cell cycle,
metastasis, development and differentiation (18,19).
Furthermore, previous studies have demonstrated that miRNAs are
able to function as tumor suppressors or oncogenes depending on
their target mRNAs (20–22). Upregulated miRNAs function as
oncogenes by negatively regulating tumor suppressor genes. In
contrast, downregulated miRNAs act as tumor suppressors via
blockade of oncogenes in tumor progression (23,24).
Therefore, miRNAs have been investigated in carcinogenesis and
progression as a novel therapeutic choice for patients with
CRC.
In the present study, miR-184 was identified to be
downregulated and behave as a tumor suppressor in CRC by inhibiting
cancer cell growth, migration and invasion. Mechanistically, it was
demonstrated that miR-184 exerted its tumor suppressive roles by
directly targeting insulin-like growth factor 1 receptor (IGF-1R)
in CRC.
Materials and methods
Tissue specimens
The procedure of human tissue specimen collection
was approved by the Ethical Committee of the Affiliated Hospital of
Nanjing University of Traditional Chinese Medicine (Nanjing,
China). In addition, written informed consent was obtained from all
patients. In the present study, all experimental protocols were
performed according to the approved guidelines. In total, 32 pairs
of CRC tissues and matched normal adjacent tissues (NATs) were
obtained from patients (age range, 38–72 years; median age, 51; 19
males and 13 females) who had been diagnosed with CRC and underwent
surgery at the Affiliated Hospital of Nanjing University of
Traditional Chinese Medicine between June 2013 and February 2015.
CRC tissues and matched NATs were immediately frozen in liquid
nitrogen and stored at −80°C until use.
Cell culture
Human CRC cell lines (HCT116, HT29 and SW620) and
human normal colon epithelium cell line (FHC) were purchased from
the American Type Culture Collection (Manassas, VA, USA). The human
embryonic kidney HEK293T cell line was obtained from the Shanghai
Institute of Biochemistry and Cell Biology (Shanghai, China). All
cells were cultured in Dulbecco's modified Eagle's medium (HT29 and
FHC cells) or RPMI-1640 medium (HCT116 and SW620 cells) containing
10% fetal bovine serum (FBS), 100 mg/ml penicillin and 100 mg/ml
streptomycin (all from Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). All cells were incubated at 37°C in a humidified
incubator containing 5% CO2.
Transfection
miR-184 mimic, negative control (NC), IGF-1R small
interfering RNA (siRNA) and NC siRNA were purchased from Guangzhou
RiboBio Co., Ltd (Guangzhou, China). The miR-184 mimic sequence was
5′-GGCAUUCUGUAUACAUCGGAG-3′ and the NC sequence was
5′-UUCUCCGAACGUGUCACGUTT-3′. The IGF-1R siRNA sequence was
5′-CACCGCGGCTGGAAACTCTTCTACACGAATGTAGAAGAGTTTCCAGCCGC-3′ and the NC
siRNA sequence was
5′-CACCGCTCACCGGCTCCAGATTTATCGAAATAAATCTGGAGCCGGTGAGC-3′.
Luciferase reporter plasmids, PGL3-IGF-1R-3′UTR wild-type (WT) and
PGL3-IGF-1R-3′UTR mutant (Mut), were obtained from GenePharma
(Shanghai, China). Cell transfections were performed using
Lipofectamine™ 2000 (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted from tissues or cells using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. The expression levels of
miR-184 in CRC tissues and cell lines were determined using a
TaqMan miRNA assay (Applied Biosystems; Thermo Fisher Scientific,
Inc.). For determination of miRNA expression, cDNA was synthesized
from total RNA and amplified using a TaqMan MicroRNA Reverse
Transcription kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. The cycling
conditions for amplification were as follows: 40 cycles of
denaturation at 95°C for 15 sec; and annealing/extension at 60°C
for 60 sec. To quantify IGF-1R mRNA expression, cDNA was
synthesized using the PrimeScript RT-PCR kit (Takara Bio, Inc.,
Otsu, Japan). IGF-1R expression was assessed using a SYBR-Green PCR
kit (Takara Bio, Inc.) on an ABI 7500 Real-Time PCR Detection
system (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
thermocycling conditions for IGF-1R quantification were as follows:
95°C for 10 min; 40 cycles of 95°C for 15 sec and 60°C for 1 min.
miR-184 expression was normalized to that of U6 RNA, and GADPH was
used as an internal control for IGF-1R expression. The primers were
designed as follows: miR-184 forward, 5′-GCATGCCTAAATGTTGACAGCC-3′
and reverse, 5′-GTGCAGGGTCCGAGGT-3′; U6 forward,
5′-CTCGCTTCGGCAGCACATATACT-3′ and reverse,
5′-ACGCTTCACGAATTTGCGTGTC-3′; IGF-1R forward,
5′-GGAGGCTGAATACCGCAAAGTC-3′ and reverse,
5′-AAAGACGAAGTTGGAGGCGCT-3′; and GAPDH forward,
5′-ATCTGGAGTTTACCGCTGG-3′ and reverse, 5′-TACCGATGTCTGGTAGACGAT-3′.
Relative expression levels were calculated using the
2−ΔΔCq method (25).
Cell proliferation assay
Cell proliferation was determined using the Cell
Counting kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). Cells were seeded into 96-well plates at a
density of 3,000 cells/well in 150 µl culture medium. Following
incubation at 37°C overnight, cells underwent transfection.
Following various incubation times (24, 48, 72 or 96 h), cell
proliferation assay was conducted. A 10 µl volume of CCK-8 assay
solution was added to each well, and the 96-well plates were
incubated at 37°C for a further 2 h. The absorbance at 450 nm was
determined using an automatic multiwell spectrophotometer (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). All experiments were
performed in triplicate.
Transwell assay
To evaluate cell migration and invasion abilities,
Transwell chambers (Costar; Corning Incorporated, Corning, NY, USA)
with 8 µm diameter pores were used. For analysis of cell invasive
ability, Matrigel (BD Biosciences, San Jose, CA, USA) was added
into the Transwell apparatus. A total of 5×104
transfected cells in 200 µl serum-free culture medium was placed in
the upper chamber and 500 µl culture medium containing 20% FBS was
added to the lower chamber as a chemoattractant. Following
incubation at 37°C for 24 h, Transwell chambers were stained with
0.5% crystal violet and washed with PBS three times. Non-migrating
and non-invading cells were carefully wiped from the upper chambers
with cotton wool. Values for cell migration and invasion were
obtained by counting 5 fields per membrane using a light
microscope.
Bioinformatic analysis
Bioinformatic analysis was performed to predict the
potential targets of miR-184 using TargetScan (http://www.targetscan.org/) and PicTar (http://pictar.mdcberlin.de/).
Western blot analysis
Transfected cells were harvested and lysed in
radioimmunoprecipitation buffer (Thermo Fisher Scientific, Inc.).
Total protein concentration was quantified using a bicinchoninic
acid protein assay kit (Thermo Fisher Scientific, Inc.). Equal
amounts of protein were separated using SDS-PAGE (10%). Proteins
were transferred onto polyvinylidene fluoride membranes (EMD
Millipore, Billerica, MA, USA), followed by incubation at room
temperature with 5% non-fat dry milk in TBST for 1 h. The membranes
were incubated with primary antibodies at 4°C overnight, including
mouse anti-human monoclonal IGF-1R (1:500 dilution; cat. no.
sc-81464; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) and
GADPH (1:500 dilution; cat. no. sc-59540; Santa Cruz Biotechnology,
Inc.). On the second day, the membranes were incubated with a
corresponding secondary antibody conjugated to horseradish
peroxidase (1:1,000 dilution; cat. no. sc-2005; Santa Cruz
Biotechnology, Inc.). Each blot was visualized using enhanced
chemiluminescence (Pierce; Thermo Fisher Scientific, Inc.), and
densitometry analysis was performed using ImageJ 1.49 (National
Institutes of Health, Bethesda, MD, USA).
Luciferase reporter assay
HEK293T cells were seeded in a 24-well plates. Cells
were co-transfected with miR-184 mimic or NC, and co-transfection
with PGL3-IGF-1R-3′UTR WT or PGL3-IGF-1R-3′UTR Mut, using
Lipofectamine 2000. At 48 h after transfection, luciferase activity
was determined using the Dual-Luciferase Reporter assay system
(Promega Corporation, Madison, WI, USA), and normalized to the
activity of Renilla luciferase. Each assay was performed
three times.
Statistical analysis
All results are expressed as the mean ± standard
deviation. Results were compared using SPSS software (version 17.0;
SPSS Inc., Chicago, IL, USA). P<0.05 (two-tailed) was considered
to indicate a statistically significant difference.
Results
miR-184 is downregulated in CRC
tissues and cell lines
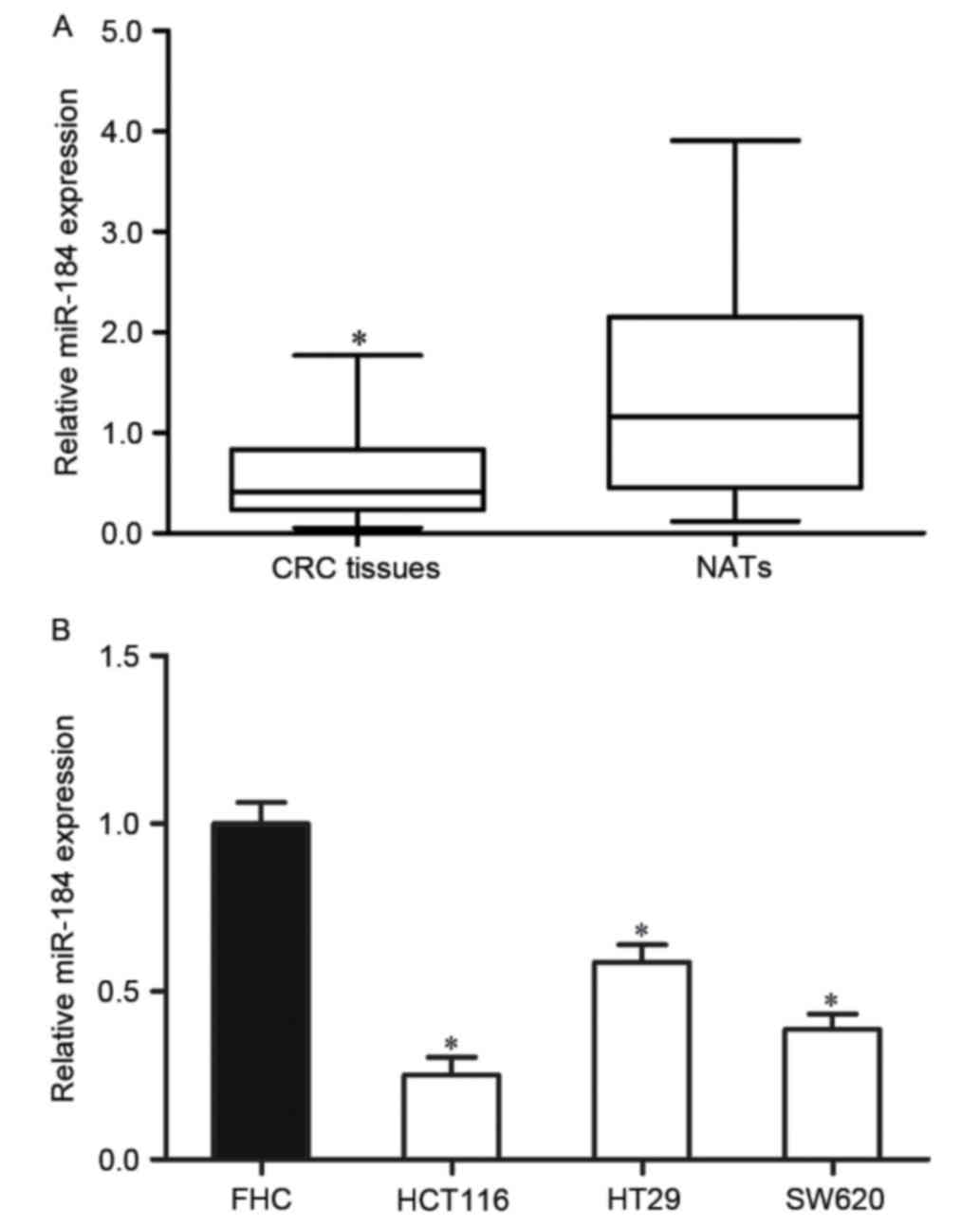
To investigate the roles of miR-184 in CRC, the
miR-184 expression level was examined in CRC tissues and matched
NATs using RT-qPCR. The results indicated that miR-184 was
significantly downregulated in CRC tissues compared with matched
NATs (P<0.05; Fig. 1A).
In addition, the expression level of miR-184 in
human CRC cell lines was also determined. The results indicated
that the miR-184 expression level was significantly decreased in
the three CRC cell lines compared with that of the human normal
colon epithelium cell line FHC (P<0.05; Fig. 1B). These results indicated that
miR-184 may serve an important role in CRC.
miR-184 inhibits CRC cell
proliferation, migration and invasion
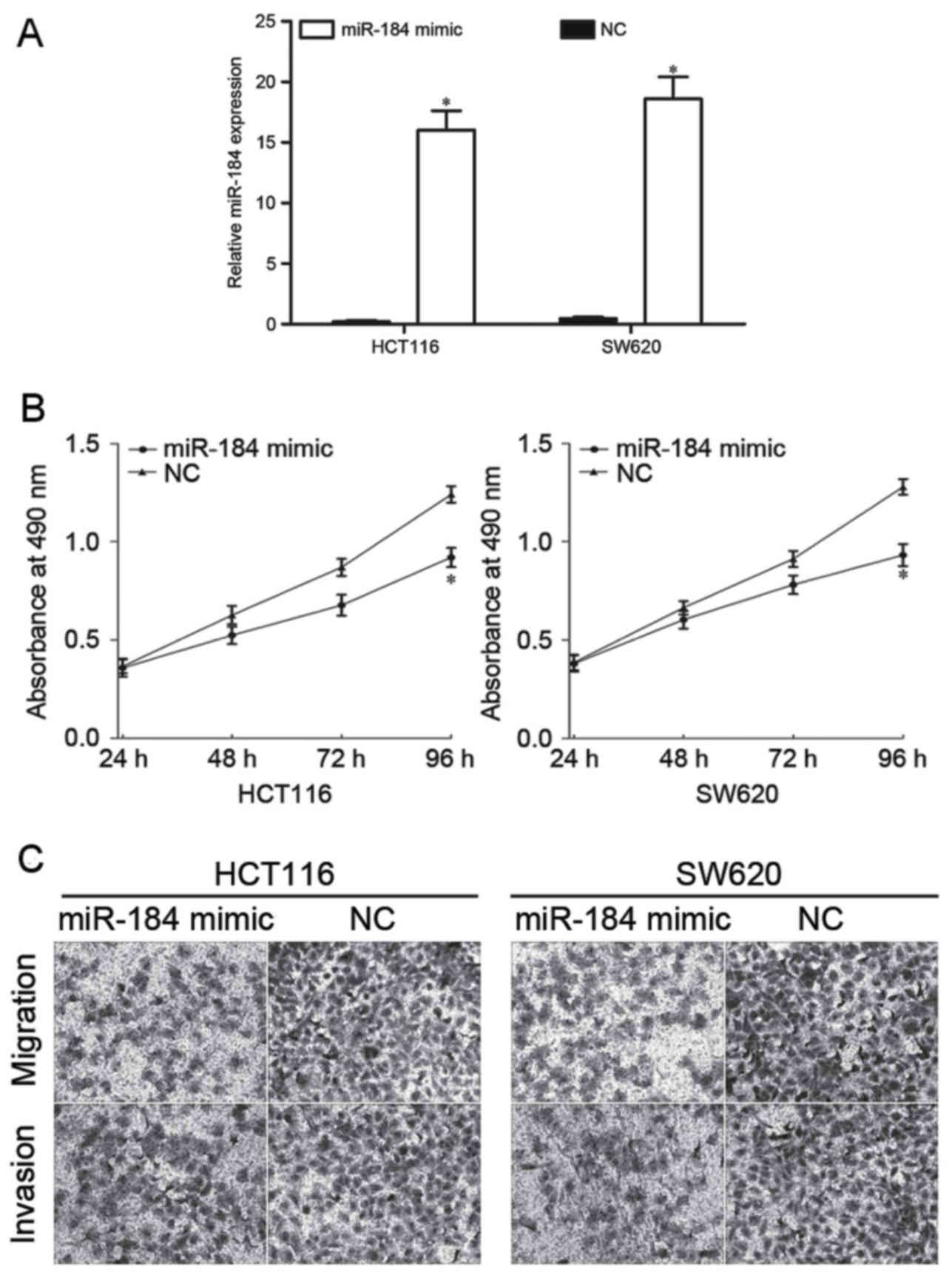
To investigate the effect of miR-184 in CRC
proliferation, migration and invasion, miR-184 mimic or NC was
transfected into HCT116 and SW620 cells. Following transfection,
RT-qPCR was performed to detect miR-184 expression. RT-qPCR
analysis demonstrated that miR-184 was significantly upregulated in
HCT116 and SW620 cells (Fig. 2A). The
effect of overexpression of miR-184 on cell proliferation using an
MTT assay was subsequently evaluated. The results indicated that
the proliferation of CRC cells was markedly inhibited following
transfection with miR-184 mimic compared with cells transfected
with NC (Fig. 2B). Furthermore,
Transwell assays were used to assess the effect of upregulated
miR-184 in CRC cell migration and invasion. The results identified
that overexpression of miR-184 significantly decreased the
migratory and invasive abilities of HCT116 and SW620 cells
(Fig. 2C). These results suggested
that miR-184 inhibited the proliferation, migration and invasion of
CRC cells in vitro.
miR-184 negatively regulates IGF-1R
expression
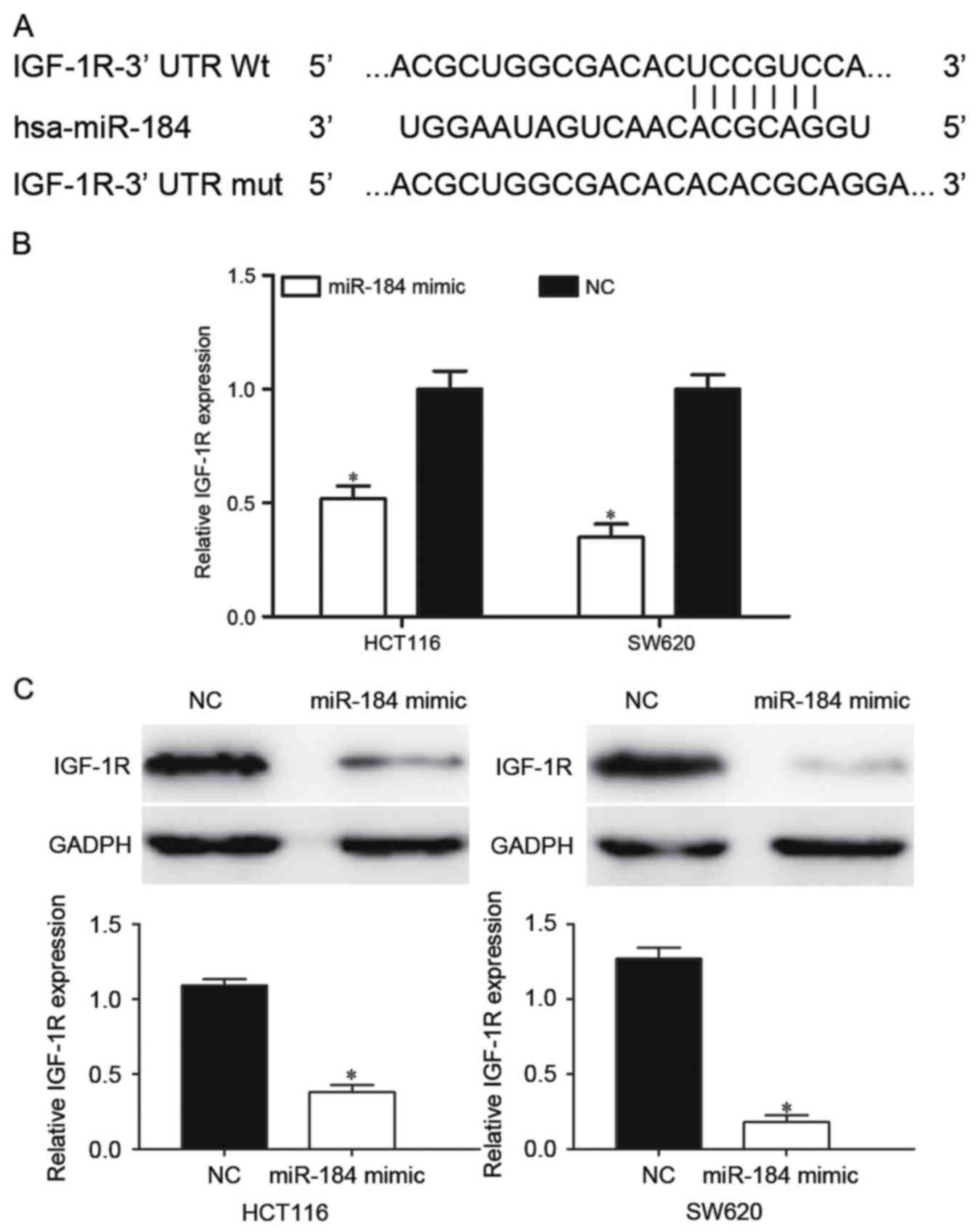
Bioinformatic databases were used to predicted
potential targets of miR-184. Among these predicted targets, IGF-1R
was a clear candidate since it was predicted by TargetScan and
PicTar. As presented in Fig. 3A,
IGF-1R contained a miR-184 seed match of the IGF-1R 3′-UTR. To
investigate the effect of miR-184 in the regulation of IGF-1R
expression, RT-qPCR and western blot analysis were performed. It
was identified that the IGF-1R mRNA was significantly downregulated
in miR-184 mimic-transfected HCT116 and SW620 cells (P<0.05;
Fig. 3B). Western blot analysis
results indicated that miR-184 significantly decreased IGF-1R
protein expression in HCT116 and SW620 cells compared with the NC
groups (P<0.05; Fig. 3C).
IGF-1R is a direct target of
miR-184
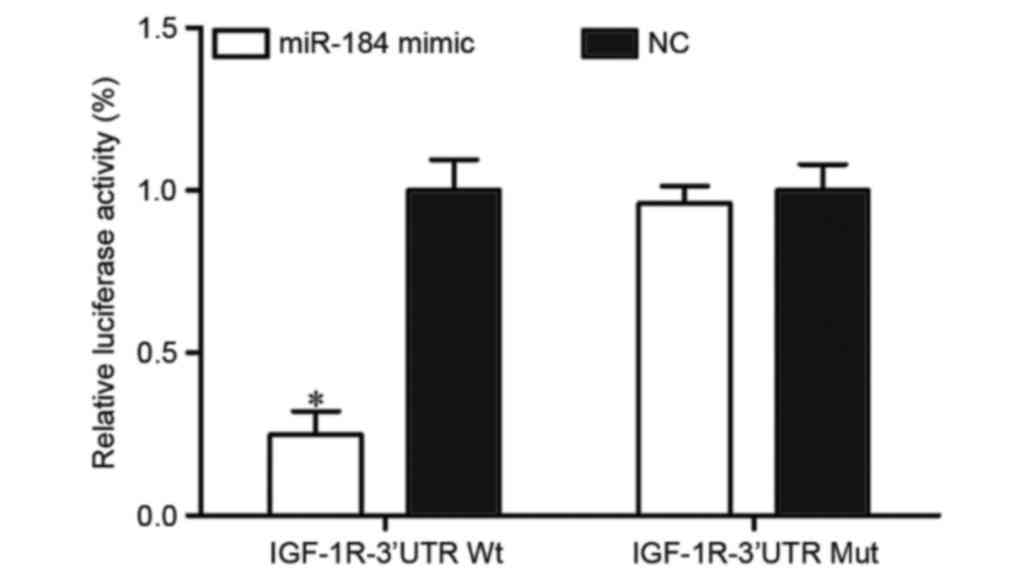
To verify whether IGF-1R was a direct target of
miR-184 in CRC, PGL3-IGF-1R-3′UTR WT and PGL3-IGF-1R-3′UTR Mut
along with miR-184 mimic or NC were transfected into HEK293T cells.
Following transfection for 48 h, luciferase reporter assays were
performed. The results identified that miR-184 significantly
inhibited the luciferase activity of PGL3-IGF-1R-3′UTR WT
(P<0.05; Fig. 4). However, the
activity of PGL3-IGF-1R-3′UTR Mut was not affected (Fig. 4), which suggested that IGF-1R was a
direct target of miR-184.
IGF-1R is involved in miR-184-mediated
proliferation, migration and invasion in CRC cells
To explore further whether IGF-1R acts as a
downstream effector in miR-184-mediated suppressive effects in CRC
cells, IGF-1R siRNA or NC siRNA was transfected into HCT116 and
SW620 cells. Following transfection, western blot analysis was
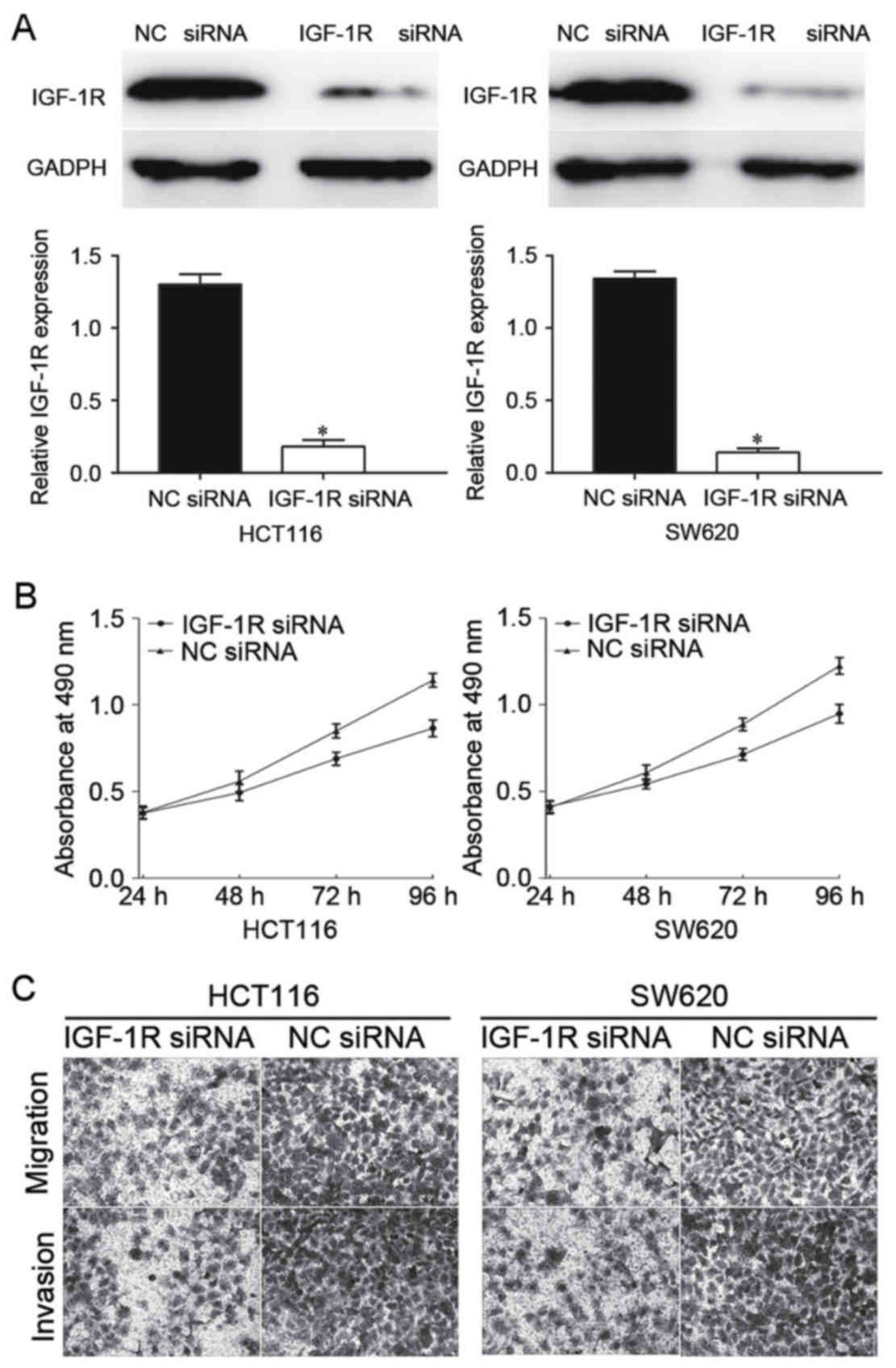
performed to determine the expression of IGF-1R protein. As
presented in Fig. 5A, IGF-1R was
significantly downregulated in IGF-1R siRNA-transfected HCT116 and
SW620 cells compared with NC siRNA-transfected cells
(P<0.05).
Subsequently, MTT assays and Transwell assays were
performed. The results identified that knockdown of IGF-1R
inhibited CRC cell proliferation, migration and invasion (Fig. 5B and C), indicating that IGF-1R was
involved in the miR-184-mediated proliferation, migration and
invasion of CRC cells.
Discussion
Previous studies have demonstrated that the
expression of miR-184 differs between normal tissues and tumors. It
has been identified to be upregulated in hepatocellular carcinoma
(26) and squamous cell carcinoma of
the tongue (27). However,
downregulation of miR-184 has also been identified in various types
of human cancer, including renal cell carcinoma (28), nasopharyngeal carcinoma (29), neuroblastoma (30), non-small cell lung cancer (31), glioma (32,33) and
breast cancer (32). The
contradictory results of these studies indicated that the
expression levels of miR-184 in cancer exhibit tissue-specificity.
In the present study, it was identified that miR-184 was
downregulated in CRC tissues and cell lines. The results suggested
that miR-184 may serve tumor suppressive functions in the
carcinogenesis and progression of CRC.
miR-184 was demonstrated to be a tumor suppressor in
cancer. Lin et al (31)
demonstrated that miR-184 was downregulated in non-small cell lung
cancer. Furthermore, a low expression level of miR-184 may be a
predictor of poor prognosis in patients with non-small cell lung
cancer. Upregulation of miR-184 decreased cell proliferation and
invasion by targeting cell division cycle 25A and c-Myc (31). In renal cell carcinoma, overexpression
of miR-184 suppressed cell proliferation and migration, and
enhanced cell apoptosis in vitro (28). Cheng et al (33) demonstrated that miR-184-targeted tumor
necrosis factor α-induced protein 2 decreased glioma cell
proliferation and migration in vitro, and the xenografted
tumor size in vivo (33).
Taken together, the results of these studies suggested a possible
role for miR-184 in regulating cancer carcinogenesis and
progression.
However, miR-184 has also been verified as an
oncogene in certain types of cancer. For example, in squamous cell
carcinoma of the tongue, miR-184 increased cell proliferation and
inhibited apoptosis (27). In
addition, surgical removal of the primary squamous cell carcinoma
of tongue markedly suppressed miR-184 expression in plasma
(34). In hepatocellular carcinoma,
overexpression of miR-184 significantly increased cell
proliferation, tumorigenicity and cell cycle progression by
directly targeting sex-determining region Y box 7 and inositol
polyphosphate phosphatase-like 1 (26,35). These
results appear contradictory in that miR-184 was demonstrated to be
an oncogene in certain types of cancer and a tumor suppressor in
others. This contradiction may be explained by the ‘imperfect
complementarity’ of the interactions between miRNAs and target
genes (36).
In the present study, the functions of miR-184 in
CRC cell proliferation, migration and invasion were analyzed by
transfecting miR-184 mimic and NC into HCT116 and SW620 CRC cells.
It was identified that miR-184 significantly inhibited the
proliferation, migration and invasion of CRC cells. These results
provided novel insight into the functions of miR-184 in the
initiation and development of CRC. Subsequently, the potential
underlying molecular mechanism involved in miR-184-mediated
suppressive functions of CRC cell proliferation, migration and
invasion were sought. In the present study, an important molecular
link between miR-184 and IGF-1R was identified in CRC. First,
bioinformatic analysis predicted that IGF-1R contained a miR-184
seed match at position 701–708 of the IGF-1R 3′-UTR. Secondly,
RT-qPCR and western blot analysis demonstrated that miR-184
decreased IGF-1R expression at the mRNA and protein level. Thirdly,
a luciferase reporter assay demonstrated that miR-84 directly
targeted IGF-1R 3′-UTR. Finally, IGF-1R siRNA also decreased CRC
cell proliferation, migration and invasion. These results indicated
that miR-184 targeted IGF-1R to inhibit CRC cell proliferation and
metastasis in vitro. Identification of the targets of
miR-184 is essential for understanding its role in CRC, and
developing novel targeted therapies for CRC.
IGF-1R, a transmembrane tyrosine kinase receptor of
the insulin receptor family, contains two extracellular α subunits
with the ligand-binding site and two transmembrane β subunits with
intracellular tyrosine kinase activity (37). Overexpression of IGF-1R has been
demonstrated to serve important functions in CRC, hepatocellular
carcinoma, osteosarcoma, non-small cell lung cancer and prostate
cancer (38–41). IGF-1R was identified to be involve in
a range of biological processes, including cell proliferation,
apoptosis, cell cycle regulation and metastasis (42–44).
Furthermore, IGF-1R expression level has been
identified to be associated with an aggressive phenotype, tumor
progression, drug resistance and poor outcome in several types of
tumor, including colorectal, ovary, prostate, endometrial, gastric,
bladder, sarcoma, glioblastoma, leukemia, myeloma, gastrointestinal
stromal and breast (45). This
indicated that inhibition of IGF-1R expression may serve as a novel
therapeutic method for the treatment of cancer. A number of agents
targeting IGF-1R have been developed or are in development, and
certain agents are in clinical trials for cancer treatment
(46). In the present study, miR-184
was identified for the first time, to the best of our knowledge, to
negatively regulate IGF-1R expression and therefore function as a
tumor suppressor in CRC. It may be investigated as a targeted
therapy for the treatment of CRC.
The results of the present study indicate that
miR-184 is downregulated in CRC. Furthermore, miR-184 inhibited CRC
cell proliferation, migration and invasion. Mechanistically, it was
verified that miR-184 directly targeted IGF-1R. miR-184 may
therefore be a therapeutic target in the treatment of CRC.
References
|
1
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Andrews L: Dietary flavonoids for the
prevention of colorectal cancer. Clin J Oncol Nurs. 17:671–672.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Altobelli E, Lattanzi A, Paduano R,
Varassi G and di Orio F: Colorectal cancer prevention in Europe:
Burden of disease and status of screening programs. Prev Med.
62:132–141. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sugarbaker PH: Colorectal cancer:
Prevention and management of metastatic disease. Biomed Res Int.
2014:7828902014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chan DS, Lau R, Aune D, Vieira R,
Greenwood DC, Kampman E and Norat T: Red and processed meat and
colorectal cancer incidence: Meta-analysis of prospective studies.
PLoS One. 6:e204562011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu M, Xu Y, Ge M, Gui Z and Yan F:
Regulation of UHRF1 by microRNA-9 modulates colorectal cancer cell
proliferation and apoptosis. Cancer Sci. 106:833–839. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gupta GP and Massague J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Spano D, Heck C, De Antonellis P,
Christofori G and Zollo M: Molecular networks that regulate cancer
metastasis. Semin Cancer Biol. 22:234–249. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiong Y, Zhang YY, Wu YY, Wang XD, Wan LH,
Li L and Zhou LM: Correlation of over-expressions of miR-21 and
Notch-1 in human colorectal cancer with clinical stages. Life Sci.
106:19–24. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Manfredi S, Bouvier AM, Lepage C, Hatem C,
Dancourt V and Faivre J: Incidence and patterns of recurrence after
resection for cure of colonic cancer in a well defined population.
Br J Surg. 93:1115–1122. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Van Cutsem E, Cervantes A, Nordlinger B
and Arnold D; ESMO Guidelines Working Group, : Metastatic
colorectal cancer: ESMO Clinical Practice Guidelines for diagnosis,
treatment and follow-up. Ann Oncol. 25 Suppl 3:iii1–9. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yuan W, Sui C, Liu Q, Tang W, An H and Ma
J: Up-regulation of microRNA-145 associates with lymph node
metastasis in colorectal cancer. PLoS One. 9:e1020172014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu L, Chen L, Xu Y, Li R and Du X:
microRNA-195 promotes apoptosis and suppresses tumorigenicity of
human colorectal cancer cells. Biochem Biophys Res Commun.
400:236–240. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen DL, Wang ZQ, Zeng ZL, Wu WJ, Zhang
DS, Luo HY, Wang F, Qiu MZ, Wang DS, Ren C, et al: Identification
of microRNA-214 as a negative regulator of colorectal cancer liver
metastasis by way of regulation of fibroblast growth factor
receptor 1 expression. Hepatology. 60:598–609. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
O'Hara SP, Mott JL, Splinter PL, Gores GJ
and LaRusso NF: MicroRNAs: key modulators of posttranscriptional
gene expression. Gastroenterology. 136:17–25. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Croce CM and Calin GA: miRNAs, cancer and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Martens-Uzunova ES, Olvedy M and Jenster
G: Beyond microRNA-novel RNAs derived from small non-coding RNA and
their implication in cancer. Cancer Lett. 340:201–211. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu X, Zhang X, Dhakal IB, Beggs M,
Kadlubar S and Luo D: Induction of cell proliferation and survival
genes by estradiol-repressed microRNAs in breast cancer cells. BMC
Cancer. 12:292012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yanokura M, Banno K, Kobayashi Y, Kisu I,
Ueki A, Ono A, Masuda K, Nomura H, Hirasawa A, Susumu N and Aoki D:
MicroRNA and endometrial cancer: Roles of small RNAs in human
tumors and clinical applications (Review). Oncol Lett. 1:935–940.
2010.PubMed/NCBI
|
|
23
|
Ventura A and Jacks T: MicroRNAs and
cancer: Short RNAs go a long way. Cell. 136:586–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang T, Thakur A, Chen T, Yang L, Lei G,
Liang Y, Zhang S, Ren H and Chen M: MicroRNA-15a induces cell
apoptosis and inhibits metastasis by targeting BCL2L2 in non-small
cell lung cancer. Tumour Biol. 36:4357–4365. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gao B, Gao K, Li L, Huang Z and Lin L:
miR-184 functions as an oncogenic regulator in hepatocellular
carcinoma (HCC). Biomed Pharmacother. 68:143–148. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wong TS, Liu XB, Wong BY, Ng RW, Yuen AP
and Wei WI: Mature miR-184 as potential oncogenic microRNA of
squamous cell carcinoma of tongue. Clin Cancer Res. 14:2588–2592.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Su Z, Chen D, Li Y, Zhang E, Yu Z, Chen T,
Jiang Z, Ni L, Yang S, Gui Y, et al: microRNA-184 functions as
tumor suppressor in renal cell carcinoma. Exp Ther Med. 9:961–966.
2015.PubMed/NCBI
|
|
29
|
Zhen Y, Liu Z, Yang H, Yu X, Wu Q, Hua S,
Long X, Jiang Q, Song Y, Cheng C, et al: Tumor suppressor PDCD4
modulates miR-184-mediated direct suppression of C-MYC and BCL2
blocking cell growth and survival in nasopharyngeal carcinoma. Cell
Death Dis. 4:e8722013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tivnan A, Foley NH, Tracey L, Davidoff AM
and Stallings RL: MicroRNA-184-mediated inhibition of tumour growth
in an orthotopic murine model of neuroblastoma. Anticancer Res.
30:4391–4395. 2010.PubMed/NCBI
|
|
31
|
Lin TC, Lin PL, Cheng YW, Wu TC, Chou MC,
Chen CY and Lee H: MicroRNA-184 deregulated by the microRNA-21
promotes tumor malignancy and poor outcomes in non-small cell lung
cancer via targeting CDC25A and c-Myc. Ann Surg Oncol. 22 Suppl
3:S1532–S1539. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Feng R and Dong L: Inhibitory effect of
miR-184 on the potential of proliferation and invasion in human
glioma and breast cancer cells in vitro. Int J Clin Exp Pathol.
8:9376–9382. 2015.PubMed/NCBI
|
|
33
|
Cheng Z, Wang HZ, Li X, Wu Z, Han Y, Li Y,
Chen G, Xie X, Huang Y, Du Z, et al: MicroRNA-184 inhibits cell
proliferation and invasion and specifically targets TNFAIP2 in
Glioma. J Exp Clin Cancer Res. 34:272015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wong TS, Ho WK, Chan JY, Ng RW and Wei WI:
Mature miR-184 and squamous cell carcinoma of the tongue.
Scientific World Journal. 9:130–132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu GG, Li WH, He WG, Jiang N, Zhang GX,
Chen W, Yang HF, Liu QL, Huang YN, Zhang L, et al: Mir-184
post-transcriptionally regulates SOX7 expression and promotes cell
proliferation in human hepatocellular carcinoma. PLoS One.
9:e887962014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yu Z, Ni L, Chen D, Zhang Q, Su Z, Wang Y,
Yu W, Wu X, Ye J, Yang S, et al: Identification of miR-7 as an
oncogene in renal cell carcinoma. J Mol Histol. 44:669–677. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hu Q, Gong JP, Li J, Zhong SL, Chen WX,
Zhang JY, Ma TF, Ji H, Lv MM, Zhao JH and Tang JH: Down-regulation
of miRNA-452 is associated with adriamycin-resistance in breast
cancer cells. Asian Pac J Cancer Prev. 15:5137–5142. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang YH, Wang ZX, Qiu Y, Xiong J, Chen YX,
Miao DS and De W: Lentivirus-mediated RNAi knockdown of
insulin-like growth factor-1 receptor inhibits growth, reduces
invasion and enhances radiosensitivity in human osteosarcoma cells.
Mol Cell Biochem. 327:257–266. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang YH, Han XD, Qiu Y, Xiong J, Yu Y,
Wang B, Zhu ZZ, Qian BP, Chen YX, Wang SF, et al: Increased
expression of insulin-like growth factor-1 receptor is correlated
with tumor metastasis and prognosis in patients with osteosarcoma.
J Surg Oncol. 105:235–243. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Scharf JG and Braulke T: The role of the
IGF axis in hepatocarcinogenesis. Horm Metab Res. 35:685–693. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shiratsuchi I, Akagi Y, Kawahara A,
Kinugasa T, Romeo K, Yoshida T, Ryu Y, Gotanda Y, Kage M and
Shirouzu K: Expression of IGF-1 and IGF-1R and their relation to
clinicopathological factors in colorectal cancer. Anticancer Res.
31:2541–2545. 2011.PubMed/NCBI
|
|
42
|
Werner H and LeRoith D: The role of the
insulin-like growth factor system in human cancer. Adv Cancer Res.
68:183–223. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pollak M: The insulin and insulin-like
growth factor receptor family in neoplasia: An update. Nat Rev
Cancer. 12:159–169. 2012.PubMed/NCBI
|
|
44
|
King H, Aleksic T, Haluska P and Macaulay
VM: Can we unlock the potential of IGF-1R inhibition in cancer
therapy? Cancer Treat Rev. 40:1096–1105. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hewish M, Chau I and Cunningham D:
Insulin-like growth factor 1 receptor targeted therapeutics: Novel
compounds and novel treatment strategies for cancer medicine.
Recent Pat Anticancer Drug Discov. 4:54–72. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wu J and Zhu AX: Targeting insulin-like
growth factor axis in hepatocellular carcinoma. J Hematol Oncol.
4:302011. View Article : Google Scholar : PubMed/NCBI
|