Introduction
Breast cancer is clinically divided into four
categories, based on the following receptor profiles: Luminal A
[estrogen receptor (ER)+, progesterone receptor
(PgR)+, human epidermal growth factor receptor 2
(HER2)−], luminal B (ER+, PgR+,
HER2+), basal like (ER−, PgR−,
HER2−) and HER2-like (ER−, PgR−,
HER2+). Luminal A/B tumors correspond to
hormone-responsive breast cancer phenotypes (1) and anti-hormonal therapy, including
anti-estrogens and aromatase inhibitors, represents the gold
standard of treatment for this type of cancer (2). However, resistance to these agents may
ultimately occur (3). Resistance to
standard chemotherapy and to anti-hormone therapy is the main cause
of treatment failure in patients with solid tumors.
Chemoresistance has a multifactorial origin.
Molecular changes in different signaling pathways, including
apoptosis and cell cycle regulation, modifications of the cellular
phenotype, metabolic alterations of chemotherapeutics and
dysregulation of inhibitors of apoptosis proteins (IAPs) are all
events which may be involved in chemoresistance and in the
increased malignancy of several types of tumor (4). As the steroid hormone signaling pathway
activates phosphoinositide 3-kinase (PI3k)/protein kinase B (Akt)
and that mammalian target of rapamycin (mTOR) is a downstream
mediator of Akt, certain drugs acting as mTOR inhibitors have been
developed and tested in order to overcome resistance to
anti-hormonal therapy (5).
Sirolimus (SRL) was the first mTOR inhibitor
molecule to be developed. It was obtained from the bacterium
Streptomyces hygroscopicus and has been approved for renal
transplantation (6). Everolimus
(RAD001) is derived from SRL and contains a 2-hydroxy-ethyl chain
that makes the drug more hydrophilic than SRL, increasing its oral
bioavailability by ~10–16% (7). The
mechanism underlying the action of SRL and RAD001 is the inhibition
of mTOR complex 1 and the regulation of factors involved in several
cellular functions, including protein synthesis, regulation of
angiogenesis, lipid biosynthesis, mitochondrial biogenesis and
function, cell cycle and autophagy (8). RAD001 has received approval from the US
Food and Drug Administration for the treatment of hormone
receptor-positive advanced breast cancer in combination with
exemestane in post-menopausal patients with non-steroidal aromatase
inhibitor-refractory disease (9).
Unfortunately, conflicting data have been reported concerning the
responsiveness of breast cancer cells to mTOR inhibitors. For
example, a previous study has demonstrated that not all
hormone-sensitive mammary tumor cell lines have good reactivity to
RAD001 (10).
Among the multiple mechanisms involved in
chemoresistance is escape from apoptosis, which is often determined
by the increased expression of IAPs. Our group previously
demonstrated that the upregulation of one of these IAPs, survivin,
is involved in the resistance of human breast cancer cells to
taxanes and also to K858, an inhibitor of kinesins (11,12). Given
that survivin and estrogens are involved in the PI3k/Akt/mTOR
transduction pathway, it is possible to hypothesize that survivin
may be involved in the establishment of RAD001 resistance in
certain hormone-responsive breast cancers. For this purpose, the
effects of RAD001 in two human breast cancer cell lines, BT474
(luminal B) and MCF7 (luminal A) were analyzed, and the former was
demonstrated to be responsive while the latter was resistant to
RAD001. Following this, the potential involvement of survivin in
the establishment of this resistance to RAD001 was examined.
Materials and methods
Cell culture and treatments
Two human breast cancer cell lines were utilized in
the present study: MCF7 (Luminal A;
ER+/PgR+/HER2−) and BT474 (Luminal
B; ER+/PgR+/HER2+), both obtained
from the American Type Culture Collection (Manassas, VA, USA). The
cell lines were grown at 37°C and 5% CO2 in Dulbecco's
modified Eagle's medium supplemented with 10% fetal bovine serum, 2
mM glutamine and 50 U/ml penicillin-streptomycin (all
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). RAD001 (a gift from
Novartis International AG, Basel, Switzerland) was solubilized in
DMSO to form a 100 mM stock solution that was utilized at final
concentration of 100 nM. YM155 (Selleck Chemicals LLC, Houston, TX,
USA) was solubilized in DMSO to form a 10 mM stock solution and
used at a final concentration of 2.5 nM. Cells used as negative
controls were treated with equivalent quantities of DMSO rather
than RAD001, but were otherwise treated identically.
Cytotoxicity assay
To determine cytotoxicity, a sulforhodamine B
colorimetric assay was performed. Cells (1.5×104) were
plated on a 96-well plate, grown for 24 h and then treated with 100
nM RAD001 for 24, 48 and 72 h at 37°C. Cells were then fixed with
50 % trichloroacetic acid for 1 h at 4°C and stained for 30 min at
room temperature with 0.4% sulforhodamine B in 1% acetic acid.
Excess dye was removed by washing four times with 1% acetic acid.
Protein-bound dye was dissolved in 10 mM TRIS (pH 10), and optical
density was determined at 510 nm using a microplate reader.
Western blotting
Cells (2×106) were treated with 100 nM
RAD001 for 48 h at 37°C, and then with and without 2.5 nM YM155 1 h
prior to RAD001 treatment. Control cells were treated with
equivalent quantities of DMSO. Cells then were lysed by incubating
in lysis buffer (1% Triton, 0.1 % sodium dodecyl sulfate-SDS, 150
mM NaCl, 50 mM Tris HCl pH 7.4, 2 mM EDTA) plus a protease
inhibitor cocktail tablet (Roche Applied Science, Penzburg,
Germany) for 30 min at 4°C. Lysates were then centrifuged at 16,000
× g for 15 min at 4°C and the supernatant was collected. The
protein concentration was evaluated using the Bio-Rad Protein
Concentration assay (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). Protein lysate samples (50–100 µg) were separated by
molecular weight on 10, 12 or 14% SDS-PAGE and then transferred
onto nitrocellulose membranes. Membranes were blocked for 1 h at
room temperature in 5% non-fat dry milk and then incubated with
primary antibodies overnight at 4°C, washed in Tris-buffered saline
with 0.1% Tween-20 and then incubated with horseradish
peroxidase-conjugated anti-mouse IgG (cat. no. A4416) or
anti-rabbit IgG (cat. no. A0545) (both 1:5,000; Sigma-Aldrich, St.
Louis, MO, USA) for 1 h at room temperature. The filters were then
developed using enhanced chemiluminescence (Super Signal West Pico
Chemiluminescent Substrate; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) using Kodak X-Omat films (Kodak, Rochester, NY,
USA). The primary antibodies used were as follows: Rabbit
anti-survivin (1:1,000; cat. no. NB500-201; Novus Biologicals,
Littleton, CO, USA), mouse anti-poly (ADP-ribose) polymerase 1
(PARP1; 1:500; cat. no. SC-8007; Santa Cruz Biotechnology, Dallas,
TX, USA), which is able to detect both cleaved and uncleaved PARP1,
mouse anti-caspase-8 (1:500; cat. no. 9746; Cell Signaling
Technology, Danvers, MA, USA) which is able to detect both cleaved
and uncleaved caspase-8, mouse anti-caspase-9 (1:500; cat. no.
9508; Cell Signaling Technology), mouse anti-B cell lymphoma 2
(Bcl2; 1:250; cat. no. Sc7382; BD Biosciences, Franklin Lakes, NJ,
USA), rabbit anti-BCL2 associated X, apoptosis regulator (Bax;
1:250; cat. no. Sc493; BD Biosciences), rabbit anti-LC3 (1:500;
cat. no. ABC232; Sigma-Aldrich, St. Louis, MO, USA), which is
capable of recognizing the doublet of LC3 composed of the two
single bands of LC3I and LC3II; rabbit anti-beclin-1 (1:500; cat.
no. 62557; Abcam, Cambridge, UK), mouse anti-β-actin (1:750; cat.
no. A5060; Sigma Aldrich, St. Louis, MO, USA).
Experiments were performed in triplicate and each
band from the blots was quantified using ImageJ v.1.48 software
(National Institutes of Health, Bethesda, MD, USA) and the mean
value was calculated and expressed as densitometric units (DU).
Reverse transcription polymerase chain
reaction (RT-PCR) assay
A total of 2×106 cells were treated with
100 nM RAD001 for 48 h at 37°C and then total RNA was isolated
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Moloney murine leukemia
virus reverse transcriptase (New England BioLabs, Inc., Ipswich,
MA, USA) was used to reverse-transcribe 1 µg total RNA into cDNA at
42°C. cDNA (5 µg) was then subjected to PCR in a buffer (New
England BioLabs, Inc.) containing 25 pmol upstream and downstream
primers and 1.25 U Platinum Taq polymerase and dNTP (both
EuroClone, Pero, Italy). The number of amplified products,
expressed in arbitrary optical density units, were normalized to
GADPH expression, which was used as the housekeeping gene. The
amplification reaction was performed in a Piko-Thermal Cycler
(Finnzymes Instruments; Thermo Fisher Scientific, Inc.). The
resulting PCR products were separated on 2% agarose gel and
visualized using Gel-Red (Biotium, Inc., Fremont, CA, USA). The
sequences of the human gene-specific primers and the sizes of the
amplified products were as follows: GAPDH forward,
5′-AGATGTTCCAATATGATTCC-3′ and reverse,
5′-TGGACTCCACGACGTACTCAG-3′; 161 bp (Sigma-Aldrich, St. Louis, MO,
USA); survivin forward, 5′-CAGATTTGAATCGCGGGACCC-3′ and reverse,
5′-CCAAGTCTGGCTCGTTCTCAG-3′; 206 bp (Primm, Milano, Italy). The PCR
program was as follows: 94°C for 5 min, followed by 30 cycles at
94°C for 30 sec, 60°C for 30 sec and 72°C for 30 sec, and a final
step of 72°C for 5 min.
Cell apoptosis assay
Cells (2×106) were treated with 100 nM
RAD001 or DMSO for 48 h at 37°C. Detached and adherent cells were
harvested using trypsin-EDTA and washed with cold PBS. The cells
were double stained with APC-Annexin V-allophycocyanie (cat. no.
550474, BD Biosciences, Franklin Lakes, NJ, USA) and
7-amino-actinomycin (7-AAD) (cat. no. 559925; BD Biosciences) in a
calcium binding buffer (cat. no. 556454; BD Biosciences) according
to the manufacturer's protocol and analyzed using the fluorescence
activated cell sorting (FACS) cytofluorimeter FACSCalibur (BD
Biosciences) and the CellQuest Pro software version 5.1 (BD
Biosciences).
Statistical analysis and graphic
programs
All results were analyzed using one-way analysis of
variance and the significance was evaluated using the Tukey honest
significant difference post hoc test. All figures were created
using Adobe Photoshop CS5 (Adobe Systems, Inc., San Jose, CA, USA),
all graphs were produced and statistical analyses conducted using
Graph Pad Prism 5.0 (GraphPad Software, Inc., La Jolla, CA,
USA).
Results
The effect of RAD001 on cell viability was assessed,
and the concentration of 100 nM RAD001 was selected based on the
results of a previous study performed on a large panel of breast
cancer cell lines (10). MCF7 and
BT474 cells were treated for 24, 48 and 72 h with 100 nM RAD001. No
significant effect was evident on cell viability in either cell
line following 24 h of treatment, while a significant decrease in
cell viability was evidenced following treatment for 48 h in BT474
cells, but not in MCF7 cells. The described effects on cell
viability were similar when cells were treated for 72 h, without
any significant variation compared with 48 h treatment (Fig. 1A). Based on these results, BT474 cells
were indicated as sensitive and MCF7 cells indicated as resistant
to RAD001. According to these data, RAD001 was used for the
following experiments at the concentration of 100 nM for 48 h.
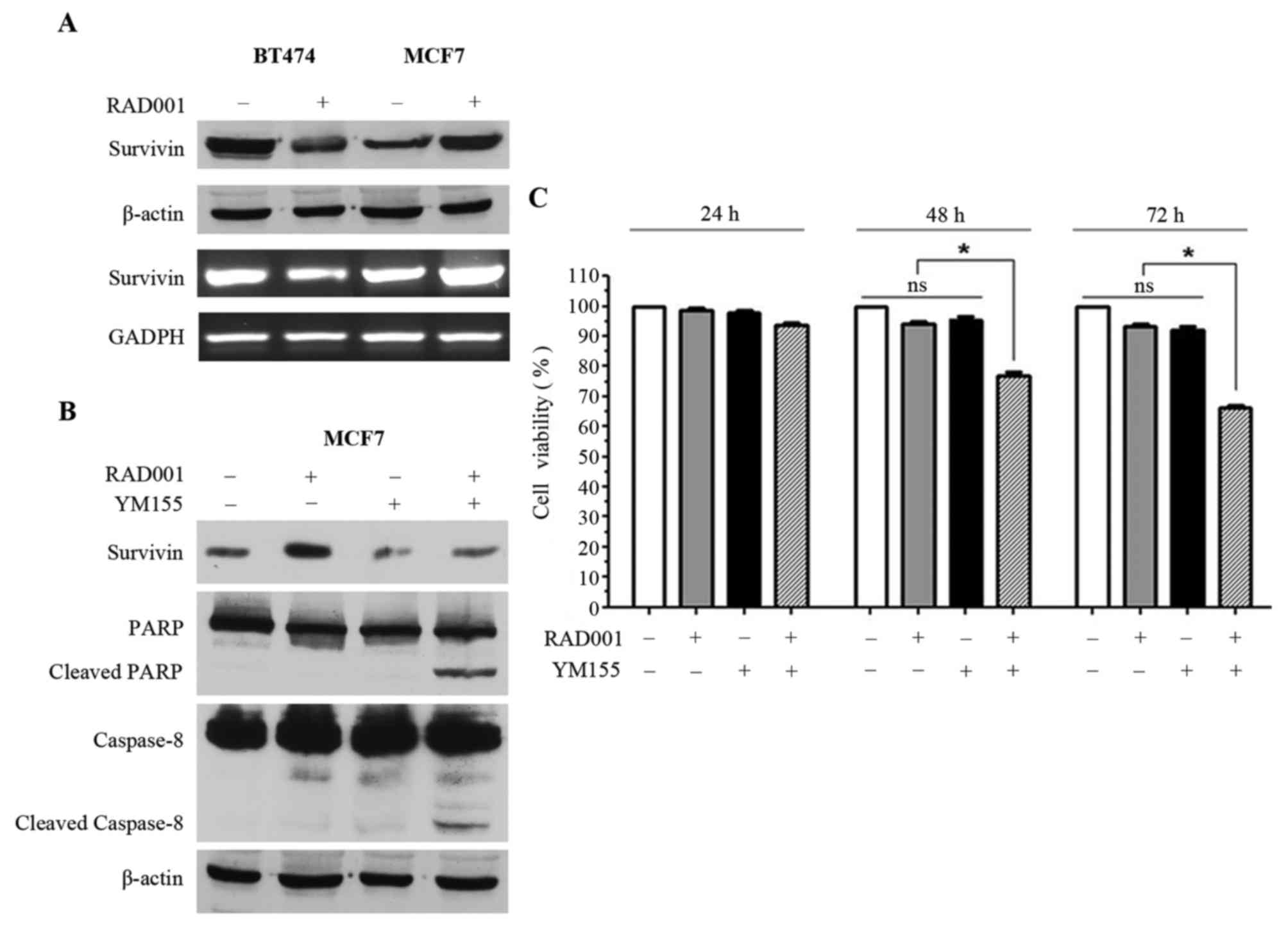
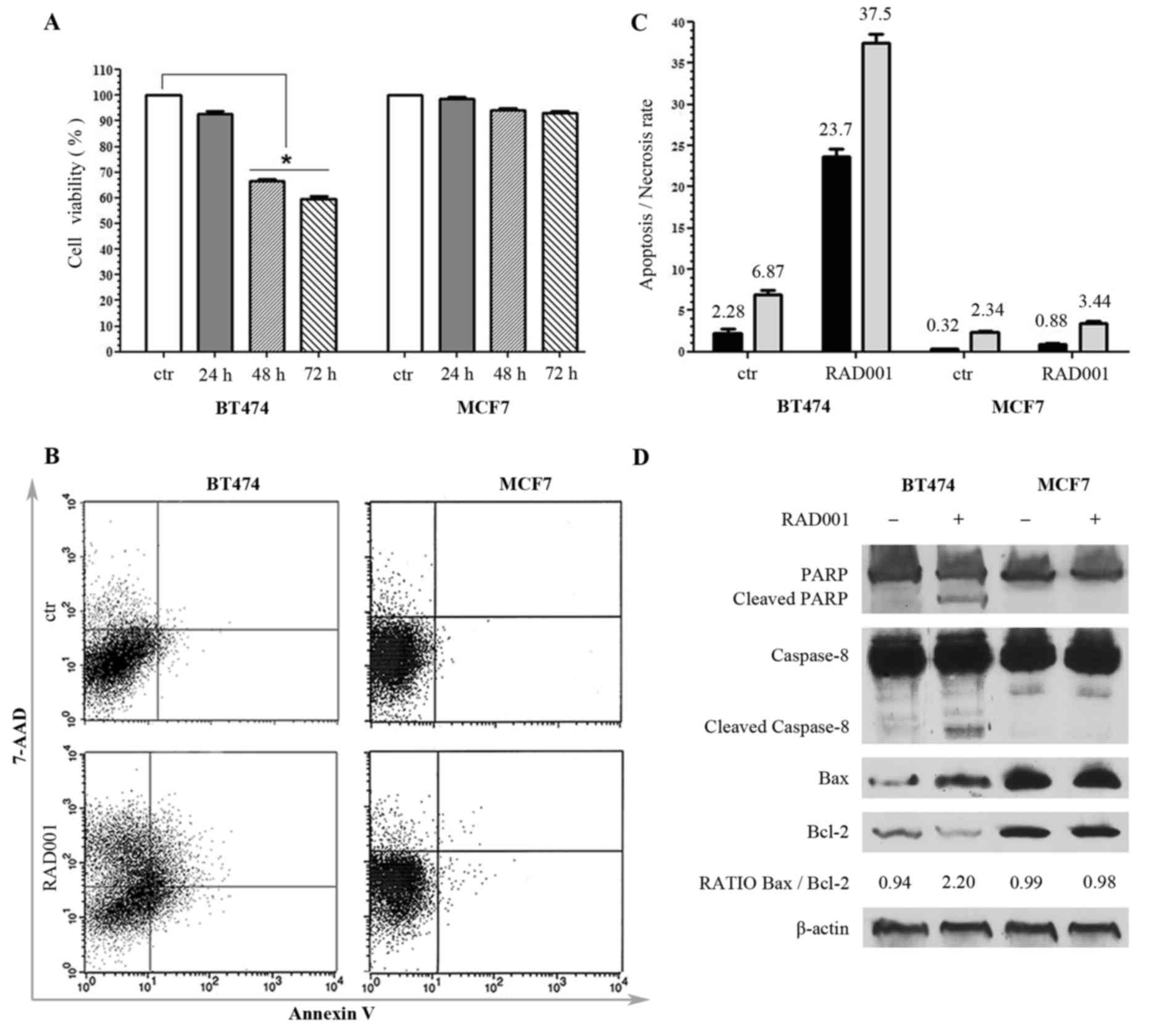 | Figure 1.Responsiveness of breast cancer cell
lines to RAD001 (A) Viability of BT474 and MCF7 cells following
RAD001 treatment, expressed as the mean ± standard deviation. (B)
Annexin V-allophycocyanin and 7-AAD expression as evaluated by
FACS. Quadrant location for the representative images: Lower left
indicates living cells, lower and upper right indicate apoptotic
cells and the upper left indicates necrotic cells. (C) Each bar
represents the FACS-reported value of apoptotic (black) and
necrotic (grey) cells, expressed as mean ± standard deviation of
three different experiments. (D) Western blotting analysis for
PARP1, caspase-8, Bax, Bcl2 and β-actin. The densitometry
quantitation is indicated for Bax/Bcl2 ratio. RAD001, everolimus;
7-AAD, 7-amino-actinomycin; FACS, fluorescence activated cell
sorting; PARP1, poly(ADP-ribose) polymerase 1; Bax, BCL2 associated
X, apoptosis regulator; Bcl2, B cell lymphoma 2. |
In order to detect whether the decrease of cell
viability determined by RAD001 was due to necrosis or to apoptosis,
untreated cells and cells treated with 100 nM RAD001 for 48 h were
double-stained with APC-conjugated Annexin V and with 7-AAD, and
then analyzed by FACS. There was no significant difference in MCF7
cells following RAD001 treatment, and the cells remained
practically negative for Annexin V and 7-AAD, confirming the
resistance to RAD001. BT474 cells responded to RAD001 with a
significant increase of Annexin V staining and a lower increase of
7-AAD, indicative of a higher rate of apoptosis (a 10-fold increase
compared with the control) as compared with the rate of necrosis (a
5-fold increase compared to the control) (Fig. 1B and C).
The apoptosis induced by 48 h treatment with 100 nM
RAD001 was further investigated by evaluating the expression of
PARP, which is a key mediator involved in DNA repair and apoptosis
that is activated in stress conditions by caspase-dependent
cleavage in the late phase of apoptosis: Treatment with RAD001
resulted in PARP cleavage in BT474 cells but not in MCF7 cells, as
evidenced by an uncleaved PARP1 band of 116 kDa present in every
sample and a cleaved PARP1 band of 89 kDa appearing only in
BT474-treated cells (Fig. 1D). The
cleavage of caspase-8 and caspase-9 was also analyzed, and RAD001
treatment was revealed not to induce any cleavage of caspase-9 in
either cell lines (data not shown). However, RAD001 treatment
induced the cleavage of caspase-8 exclusively in BT474 cells,
visualized as a band migrating at 43 kDa, while only the uncleaved
band of caspase-9 of 57 kDa was visible in MCF7 cells (Fig. 1D). These data confirmed MCF7
resistance to RAD001-induced apoptosis, and demonstrated that the
activation of apoptosis in BT474 cells followed the extrinsic
pathway. The state of Bax, which induces apoptosis, and Bcl2, which
inhibits apoptosis, was then investigated. These proteins are
members of the Bcl2 family, which controls mitochondria
permeability and cytochrome C release, and an increase of Bax/Bcl2
ratio is often indicative of apoptosis. The 21 kDa Bax band
increased and 26 kDa Bcl-2 band decreased in BT474 cells treated
with 100 nM RAD001 for 48 h, but not in MCF7 cells, resulting in an
increase of the Bax/Bcl2 ratio in treated BT474 cells, with 0.94
densitometric units (DU) in the control compared with 2.20 DU in
the treated cells, while it was not modified in MCF7 (Fig. 1D). These data confirmed the
sensitivity of BT474 cells to RAD001 and the induction of
apoptosis, and at the same time confirmed the lack of
responsiveness of MCF7 cells to RAD001.
In order to investigate the mechanisms underlying
the resistance of MCF7 to RAD001, the effect of 48 h 100 nM RAD001
treatment on survivin was analyzed. The 16 kDa survivin band and
survivin mRNA were downregulated in BT474 cells, while survivin
protein and mRNA were upregulated in MCF7 cells (Fig. 2A). Notably, RAD001 affected survivin
in opposite ways in the two cell lines, which may justify the
sensitivity of BT474 cells and, at the same time, the resistance of
MCF7 cells. Thus, it was speculated that forced negative control of
the upregulation of survivin in MCF7 may restore the sensitivity of
these cells to RAD001. YM155, an inhibitor of survivin, was
selected, however, since YM155 has also been described as an
inducer of apoptosis (13–15), we different concentrations of YM155
were tested on MCF7 cells in order to identify the concentration
capable of inhibiting survivin expression without inducing
immediate apoptosis. The concentration of 2.5 nM was selected for
YM155 and used for the following experiments. Consequently, MCF7
cells were treated with 2.5 nM YM155 1 h prior to treatment with
100 nM RAD001 for 48 h, and upregulation of survivin, which was
visible in the cells treated with RAD001 only, was reversed by
treatment with YM155 (Fig. 2B). At
the same time, treatment of MCF7 cells with RAD001 and YM155
induced apoptosis, as supported by the presence of PARP1 and
caspase-8 cleavage (Fig. 2B).
Treatment with YM155 alone was also performed as control and a
slight downregulation of survivin was observed, but PARP1 and
caspase-8 cleavage were not induced (Fig.
2B). Treatment of MCF7 cells with RAD001 and YM155 did not
induce cleavage of caspase-9 (data not shown), confirming that only
the extrinsic pathway and not the intrinsic pathway was activated,
as demonstrated in Fig. 1D.
MCF7 viability was investigated using sulforhodamine
B staining following 24, 48 and 72 h of treatment with 100 nM
RAD001 and 2.5 nM YM155, and a significant decrease of cell
viability was observed when MCF7 cells were treated for 48 and 72 h
with RAD001 together with YM155 (Fig.
2C). This effect was not evident following treatment for 24 h,
and there was no significant effect on cell viability when the
cells were treated alone with RAD001 or with YM155 (Fig. 2C). Taken together, these results
demonstrated that YM155 was able to reverse the upregulation of
survivin induced by RAD001, and at the same time was able to
restore sensitivity to RAD001 with the induction of apoptosis in
MCF7 cells.
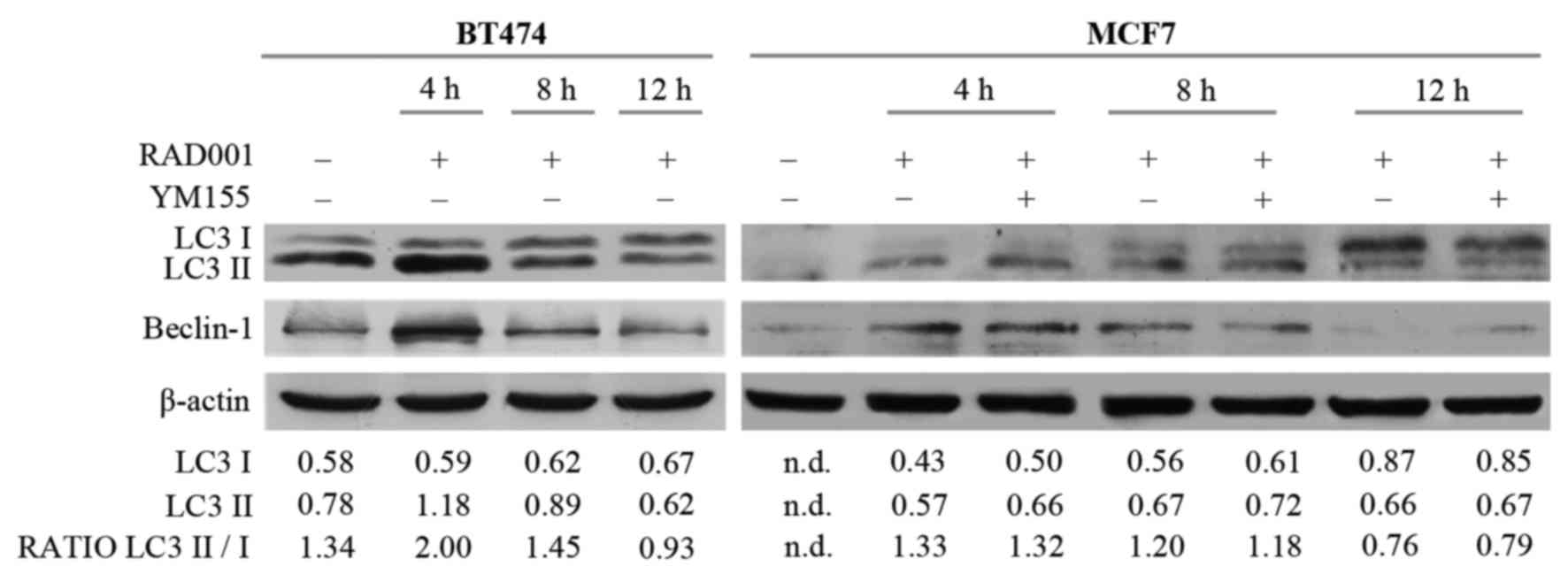
The potential involvement of autophagy was then
investigated, since RAD001 and mTOR inhibitors are described as
autophagy activators (8,16), and also because the induction of
apoptosis required a longer duration of RAD001 treatment. Autophagy
and the autophagosome are controlled by specific interactions of
several protein complexes: A protein complex including beclin-1 is
required for phagophore nucleation, while another complex including
the cytosolic and the lipidated form of LC3 protein (LC3I/LC3II) is
necessary for elongation and closure of the autophagosome (17). Therefore, the effect of RAD001 on the
expression of LC3I, LC3II and beclin-1 was evaluated. There was an
increase of 14 kDa LC3II and of 52 kDa beclin-1 in BT474 cells
treated with RAD001 for 4 h, but LC3II and beclin-1 expression
decreased so that it was similar to the untreated condition
following treatment with 100 nM RAD001 for 8 and 12 h (Fig. 3). A high ratio between the LC3II 16
kDa band and LC3I 14 kDa band is indicative of autophagy, and an
increase of this ratio was observed following treatment for 4 h
(Fig. 3), confirming that RAD001
induced autophagy in BT474 cells, but only as an early event. At
the same time, RAD001 induced autophagy in MCF7 cells, as evidenced
by an increase of LC3II and beclin-1 expression following 4 and 8 h
of treatment with 100 nM RAD001, while the beclin-1 and LC3II/LC3I
ratio were slightly reduced following treatment for 12 h (Fig. 3). The addition of YM155 to RAD001
treatment in MCF7 failed to modify the described effects on LC3 and
beclin-1. Longer treatment of BT474 and MCF7 cells with 100 nM
RAD001 also failed to modify the expression of LC3I–II and beclin-1
(data not shown). These results confirmed that autophagy was
induced by RAD001 as an early event, and that it stopped following
longer treatment durations.
Discussion
RAD001 is an inhibitor of m-TOR derived from SRL
(7) that is utilized for the
treatment of hormone receptor-positive advanced breast cancer in
combination with exemestane (9).
Unfortunately, a previous study has demonstrated that not all
hormone-sensitive mammary tumors respond well to RAD001 in
vitro (10). Consequently, two
cell lines that responded differently to RAD001 were selected: One
sensitive (BT474) and the other resistant (MCF7). Our group has
previously demonstrated that the upregulation of survivin, a
specific IAP component, contributes to establishing resistance to
taxanes and kinesin inhibitors in breast cancer cells (11,12).
Survivin is a downstream target of the PI3k/Akt/mTOR pathway that
is involved in drug resistance (18).
Therefore, the present study aimed to investigate whether survivin
was involved in the described resistance of hormone-sensitive
mammary tumors to RAD001. RAD001 was demonstrated to modify
survivin expression in opposite ways in the two cell lines,
characterized by downregulation in the sensitive cells and
upregulation in the resistant cells. These data suggest a potential
association between increased survivin expression and resistance to
RAD001. In order to further investigate this association, YM155, an
inhibitor of the promoter of survivin, was used to inhibit survivin
expression. When upregulation of survivin was blocked with YM155,
the sensitiveness to RAD001 was restored in MCF7 cells, as
demonstrated by the increased apoptosis rate induced when MCF7
cells were treated with YM155 in combination with RAD001. These
data strengthen the hypothesis that survivin is a mediator of drug
resistance. This is supported by data suggesting that the increase
of this protein in cancer tissues is an unfavorable prognostic
marker and is associated with increased risk of recurrence
(19). The overexpression of survivin
has been proposed as a predictive factor in determining the
response to chemotherapy (20).
Autophagy has been described as an alternative cell
death pathway that is induced by rapamycin inhibitors when
apoptosis is defective (21).
Notably, short durations of RAD001 treatment were able to induce
autophagy in the two cell lines, while 48 h of treatment were
required for the induction of apoptosis. The dysregulation of
autophagy serves a dual effect in cancer development, since its
chronic inhibition promotes cancer but, at the same time, increased
induction of autophagy is a mechanism of tumor cell survival in
advanced conditions of acidosis and hypoxia (22). In particular, increased autophagy has
been correlated with the development of resistance to chemotherapy
in breast cancer (22,23). Autophagy is a multistep process that
generates double-membrane vesicles called autophagosomes (24) which engulf the cytosolic form of LC3
(LC3-I) which is then conjugated to phosphatidylethanolamine to
form LC3-II (18), so LC3-I is
localized in the cytosol while LC3-II is present in the
autophagosomes. The ratio between LC3-II and LC3-I, which measures
the rate of autophagy, was significantly increased following
treatment with RAD001 for 4 h. In parallel, beclin-1, which is also
involved in the autophagy of breast cancer cells (25), demonstrated increased expression
following treatment with RAD001 for 4 h. Therefore, a short period
of RAD001 treatment induced autophagy in these cell lines, and
autophagy was progressively depressed from 8–48 h, and at 48 h
RAD001 activates apoptosis. These data suggest that autophagy and
apoptosis are two sequential and independent effects induced by
RAD001 in breast cancer cells. However, the inhibition of survivin
induced by YM155 failed to modify the rate of autophagy induced by
RAD001, which may be because the RAD001-induced control of survivin
expression lasts longer than RAD001-induced autophagy.
Consequently, it is possible to state that survivin controls
apoptosis but does not interfere with the autophagy induced by
RAD001.
In conclusion, survivin may be a potential target
for the inhibition of the survival and proliferation of cancer
cells, although its activation may involve several collateral
pathways, switched on in case of interruption of the principal
route of transduction. This is in accordance with different
clinical trials conducted with YM155 in combination with other
anti-cancer agents, as described in a previous review (26). A phase II study in HER2−
metastatic breast cancer performed with YM155 plus docetaxel
reported no significant differences compared with docetaxel alone
(27). However, the authors have
observed that, unless the pre-clinical data provided a good
rationale for utilizing YM155, their study presented various
limitations, including the lack of pharmacokinetic interaction
analysis between the two drugs. At the same time, a phase II study
on aggressive B-cell lymphoma treated with YM155 plus rituximab
reported encouraging antitumor activity and a durable response
(28).
Taken together with the results of previous studies,
the results of the present study suggested that YM155 may be a
valuable treatment for cancer. Furthermore, these data justify
further research into drugs effective at targeting the survivin
pathway, since the ability to negatively regulate survivin on
multiple fronts may help to control the evasion of cancer cells
from therapy-induced apoptosis.
References
|
1
|
Cancer Genome Atlas Network: Comprehensive
molecular portraits of human breast tumors. Nature. 490:61–70.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hurviz SA and Pietras RJ: Rational
management of endocrine resistance in breast cancer: A
comprehensive review of estrogen receptor biology, treatment
options, and future directions. Cancer. 113:2385–2397. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sobral AF, Amaral C, Correia-da-Silva G
and Teixeria N: Unravelling exemestane: From biology to clinical
prospects. J Steroid Biochem Mol Biol. 163:1–11. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hunter AM, LaCasse EC and Korneluk RG: The
inhibitors of apoptosis (IAPs) as cancer targets. Apoptosis.
12:1543–1568. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ghayad SE, Bieche I, Vendrell JA, Keime C,
Lidereau R, Dumontet C and Cohen PA: mTOR inhibition reverses
acquired endocrine therapy resistance of breast cancer cells at the
cell proliferation and gene-expression levels. Cancer Sci.
99:1992–2003. 2008.PubMed/NCBI
|
|
6
|
Kajiwara M and Masuda S: Role of mTOR
inhibitors in kidney disease. Int J Mol Sci. 17:E9752016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kirchner GI, Meier-Wiedenbach I and Manns
MP: Clinical pharmacokinetics of everolimus. Clin Pharmacokinet.
43:83–95. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Granata S, Dalla Gassa A, Carraro A,
Brunelli M, Stallone G, Lupo A and Zaza G: Sirolimus and Everolimus
pathway: Reviewing candidate genes influencing their intracellular
effects. Int J Mol Sci. 17:E7352016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Baselga J, Campone M, Piccart M, Burris HA
III, Rugo HS, Sahmoud T, Noguchi S, Gnat M, Pritchard KI, Lebrun F,
et al: Everolimus in postmenopausal hormone-receptor-positive
advanced breast cancer. N Engl J Med. 366:520–529. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hurvitz SA, Kalous O, Conklin D, Desai AJ,
Dering J, Anderson L, O'Brien NA, Kolarova T, Finn RS, Linnartz R,
et al: In vitro activity of the mTOR inhibitor everolimus, in a
large panel of breast cancer cell lines and analysis for predictor
response. Breast Cancer Res Treat. 149:669–680. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
De Iuliis F, Salerno G, Giuffrida A,
Milana B, Taglieri L, Rubinacci G, Giantulli S, Terella F,
Silvestri I and Scarpa S: Breast cancer cells respond differently
to docetaxel depending on their phenotype and on survivin
upregulation. Tumor Biol. 37:2603–2611. 2016. View Article : Google Scholar
|
|
12
|
De Iuliis F, Taglieri L, Salerno G,
Giuffrida A, Milana B, Giantulli S, Carradori S, Silvestri I and
Scarpa S: The kinesin Eg5 inhibitor k858 induces apoptosis but also
survivin-related chemoresistance in breast cancer cells. Invest New
Drugs. 34:399–406. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang C, Cao X, Gei Y, Wang Y, Liu G,
Cheng G and Liu Q: Silencing of survivin by YM155 induces apoptosis
and growth arrest in hepatocellular carcinoma cells. Oncol Lett.
10:1627–1631. 2015.PubMed/NCBI
|
|
14
|
Zhang S, Liu B, Fan Z, Wang D, Liu Y, Li
J, Wang N, Liu Y and Zhang B: Targeted inhibition of survivin with
YM155 promotes apoptosis of hypoxic human pulmonary arterial smooth
muscle cells via the upregulation of voltage-dependent
K+ channels. Mol Med Rep. 13:3415–3422. 2016.PubMed/NCBI
|
|
15
|
Li WL, Lee MR and Cho MY: The small
molecule survivin inhibitor YM155 may be an effective treatment
modality for colon cancer through increasing apoptosis. Biochem
Biophys Res Commun. 471:309–314. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Saran U, Foti M and Dufour JF: Cellular
and molecular effects of the mTOR inhibitor everolimus. Clin Sci
(Lond). 129:895–914. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tanida I: Autophagy basics. Microbiol
Immunol. 55:1–11. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ou DL, Lee BS, Lin LI, Liou JY, Liao SC,
Hsu C and Cheng AL: Vertical blockade of the IGFR-PI3 K/Akt/mTOR
pathway for treatment of hepatocellular carcinoma: The role of
survivin. Mol Cancer. 13:22014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pavlidou A, Kroupis C and Dimas K:
Association of survivin splice variants with prognosis and
treatment of breast cancer. World J Clin Oncol. 5:883–894. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Song J, Su H, Zhou YY and Guo LL:
Prognostic value of survivin expression in breast cancer patients:
A meta analysis. Tumor Biol. 34:2053–2062. 2013. View Article : Google Scholar
|
|
21
|
Kim KW, Mutter RW, Cao C, Albert JM,
Freeman M, Hallahan DE and Lu B: Autophagy for cancer therapy
through inhibition of pro-apoptotic proteins and mammalian target
of rapamycin signaling. J Biol Chem. 281:36883–36890. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zarzynska JM: The importance of autophagy
regulation in breast cancer development and treatment. Biomed Res
Int. 2014:7103452014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zambrano J and Yeh ES: Autophagy and
apoptotic crosstalk: Mechanism of therapeutic resistance in
HER2-positive breast cancer. Breast Cancer (Auckl). 10:13–23.
2016.PubMed/NCBI
|
|
24
|
Hjelmeland A and Zhang J: Metabolic,
autophagic, and mitophagic activities in cancer initiation and
progression. Biomed J. 39:98–106. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jung YY, Lee YK and Koo JS: The potential
of Beclin 1 as a therapeutic target for the treatment of breast
cancer. Expert Opin Ther Targets. 20:167–178. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rauch A, Henning D, Schäfer C, Wirth M,
Marx C, Heinzel T, Schneider G and Kramer OH: Survivin and YM155:
How faithful is the liaison? Biochim Biophys Acta. 1845:202–220.
2014.PubMed/NCBI
|
|
27
|
Clemens MR, Gladkov OA, Gartner E,
Vladimirov V, Crown J, Steinberg J, Jie F and Keating A: Phase II,
multicenter, open-label, randomized study of YM155 plus docetaxel
as first-line treatment in patients with HER2-negative metastatic
breast cancer. Breast Cancer Res Treat. 149:171–179. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Papadopoulos KP, Lopez-Jimenez J, Smith
SE, Steinberg J, Keating A, Sasse C, Jie F and Thyss A: A
multicenter phase II study of sepantronium bromide (YM155) puls
rituximab in patients with relapsed aggressive B-cell non-Hodgkin
lymphoma. Leuk Lymphoma. 57:1848–1855. 2016. View Article : Google Scholar : PubMed/NCBI
|