Introduction
Osteosarcoma is the most common type of primary
malignant bone tumor in children and young adults (1). Since the development of neoadjuvant
chemotherapy, the postoperative five-year survival rate of patients
with non-metastatic osteosarcoma has improved from <20% to
60–70% (2); however, resistance to
chemotherapy is common, and numerous chemotherapeutic drugs have
serious side effects. In recent years an increasing number of
studies have focused on identifying safe and effective antitumor
drugs (3,4).
Curcumol, the dried root of Rhizoma Curcumae
zedoariae, has been used for the treatment of antiviral‚
anti-inflammatory and for hepatoprotection (5–7) and its
derivatives (8–17), which have been reported to have
anticancer activities in various types of cancer such as leukemia,
glioma, lung, liver and colorectal cancer. Furthermore, Zhao et
al (18) reported that β-element
curcumol derivatives potentiated the effect of taxanes on p53
mutant H23 cells and p53 null H358 cells. Lin et al
(19), Liu et al (20) and Liu et al (21) reported that β-Elemene, the curcumol
derivatives, could induce protective autophagy from apoptosis in
hapatoma cell, non-small-cell lung cancer and gastric cells
respectively. However, Zhou et al (22) demonstrated that β-Elemene increased
autophagic apoptosis and drug sensitivity in SPC-A-1/DDP cells by
inducing beclin-1 expression. Besides, Mu et al (23) reported that β-Elemene enhanced the
efficacy of gefitinib on glioblastoma multiforme cells through the
inhibition of the EGFR signaling pathway.
Programmed cell death has been subdivided into two
categories (24): Apoptosis (type I)
and autophagic cell death (type II). There is an intricate
crosstalk between autophagy and apoptosis (25). In certain instances, autophagy may
inhibit apoptosis and promote cell survival, whereas in other
instances, autophagy may enhance apoptosis (26,27). To
the best of our knowledge, no previous studies have investigated
the effects of curcumol in the MG-63 osteosarcoma cell line, and
autophagy has not been documented. The aim of the present study was
to explore a possible association between autophagy and apoptosis
in MG-63 human osteosarcoma cells exposed to curcumol and the
potential underlying mechanism.
Materials and methods
Reagents
MG-63 osteosarcoma cells were purchased from the
Shanghai Institute of Cell Biology, Chinese Academy of Sciences
(Shanghai, China). Curcumol (purity, >98%) was obtained from
National Institutes for Food and Drug Control (Beijing, China) and
dissolved in dimethyl sulfoxide (DMSO) as a stock solution and
stored at 20°C. Curcumol was then diluted with Dulbecco's modified
Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) to the desired working concentration prior to
each experiment. SP600125 was purchased from Gibco (Thermo Fisher
Scientific, Inc.). The broad- spectrum caspase inhibitor
(z-VAD-fmk) was obtained from EMD Millipore (Billerica, MA, USA).
Fetal bovine serum was purchased from Hangzhou Sijiqing Biological
Engineering Materials Co., Ltd. (Hangzhou, China). Chloroquine (CQ)
and MTT were purchased from Sigma-Aldrich (Merck Millipore,
Darmstadt, Germany). Lipofectamine 2000 transfection reagent was
obtained from Invitrogen (Thermo Fisher Scientific, Inc.). Rabbit
monoclonal anti-caspase-3 and anti-light chain 3 (LC3) antibodies
were purchased from Cell Signaling Technology, Inc. (Danvers, MA,
USA).
Cell culture and viability assay
MG-63 cells were maintained in DMEM supplemented
with 10% fetal bovine serum, 100 U/ml penicillin and 100 µg/ml
streptomycin at 37°C in a 5% CO2 incubator. The cells in
mid-log phase were used in the experiments. Cell viability was
determined using an MTT assay. The MG-63 cells were seeded in
96-well flat bottom microtiter microplates (1×104
cells/well), and then treated with 15, 30, 60 and 120 mg/l curcumol
at room temperature for 0, 12, 24 and 48 h, respectively. The
control group and zero adjustment well were also set up. The
absorbance value per well at 570 nm was read using an automatic
multiwell spectrophotometer (Power Wave X; BioTek Instruments Inc.,
Winooski, VT, USA). All the MTT assays were performed in
triplicate. The inhibitory rate for the proliferation of MG-63
cells was determined according to the formula: (1 - experimental
absorbance value/control absorbance value) × 100%. Half maximal
inhibitory concentration (IC50) values were then
evaluated using SPSS software version 16.0 (SPSS, Inc., Chicago,
IL, USA).
Detection of apoptosis
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) double staining assay was performed to
detect the apoptotic ratio of MG-63 cells. The cells were cultured
with 15, 30, 60 and 120 mg/l curcumol for 48 h, trypsinized and
then washed twice with ice-cold PBS. The cells were then reacted
with FITC-conjugated Annexin V and PI for 15 min at room
temperature in the dark, followed by cytometric analysis (EPICS XL;
Beckman Coulter, Inc., Brea, CA, USA) within 30 min of staining.
Each group was repeatedly evaluated three times and each sample
included 1×104 cells.
Hoechst 33258 staining
MG-63 cells at the logarithmic growth phase were
seeded in 96-well plates with a cell density of
1×104/ml. Following fixation with 3.7% paraformaldelyde
for 30 min at room temperature, cells were washed with PBS and
stained with 10 mg/l Hoechst 33258 at 37°C for 15 min. The confocal
fluorescence microscope (DM2500; Leica Microsystems GmbH, Wetzlar,
Germany) equipped with an ultraviolet filter was used to observed
MG-63 cells. The images were recorded and processed on a computer
with a digital camera attached to the microscope. Normal nuclei
stained blue and apoptotic nuclei were identified as condensed or
fragmented nuclei stained bright blue.
Green fluorescent protein (GFP)-LC3
dot assay
Cells were cultured in 6-well plates at 37°C and
transfected with GFP-LC3 at 15–25°C using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. Subsequently, the cells were treated with
63.5 mg/l curcumol with or without CQ for 48 h. For observation,
the cells were fixed with 4% formaldehyde for 15 min, and then
washed twice in cold PBS. The induction of autophagy was quantified
by evaluating the percentage of cells in each group that contained
LC3 aggregates by observation under a Leica DM2500 confocal
laser-scanning microscope.
Western blot analysis protein samples were separated
electrophoretically by SDS-PAGE (12%) and transferred onto a
polyvinylidene difluoride membrane. The membrane was incubated with
5% non-fat dry milk overnight at room temperature. The membrane was
incubated with specific primary antibodies (dilution, 1:1,000)
against cleaved caspase-3, LC3I, LC3II, t-JNK and p-JNK. IgG goat
anti-rabbit and goat anti-mouse secondary antibodies (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). The bound antibodies were
detected by the enhanced chemiluminescence method and densitometric
analysis.
Statistical analysis
Data are presented as the mean ± standard deviation.
The statistical analysis was performed using analysis of variance
followed by Dunnett's t-test. P<0.05 was considered to indicate
a statistically significant difference.
Results
Curcumol inhibits MG-63 cell
proliferation in a dose- and time-dependent manner
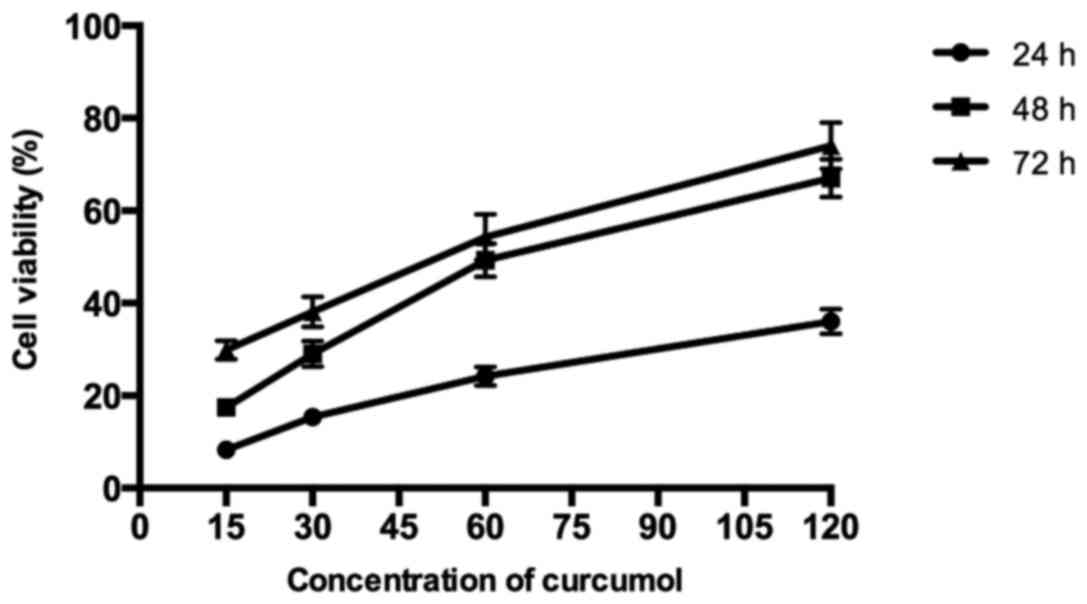
As shown in Fig. 1,
the results demonstrated that curcumol inhibited the proliferation
of MG-63 cells in a dose- and time-dependent manner following
treatment with various concentrations of curcumol (15, 30, 60 and
120 mg/l) for 24, 48 and 72 h, respectively, compared with the
control group. According to the MTT assays, the IC50
values of MG-63 cells following curcumol treatment for 48 h was
63.5 mg/l. As the length of the curcumol treatment increased and
the concentration of the dose administered increased, the
proliferation inhibitory effect of curcumol on MG-63 cells
demonstrated greater significance (P<0.05).
Curcumol triggers caspase-dependent
apoptosis in MG-63 cells
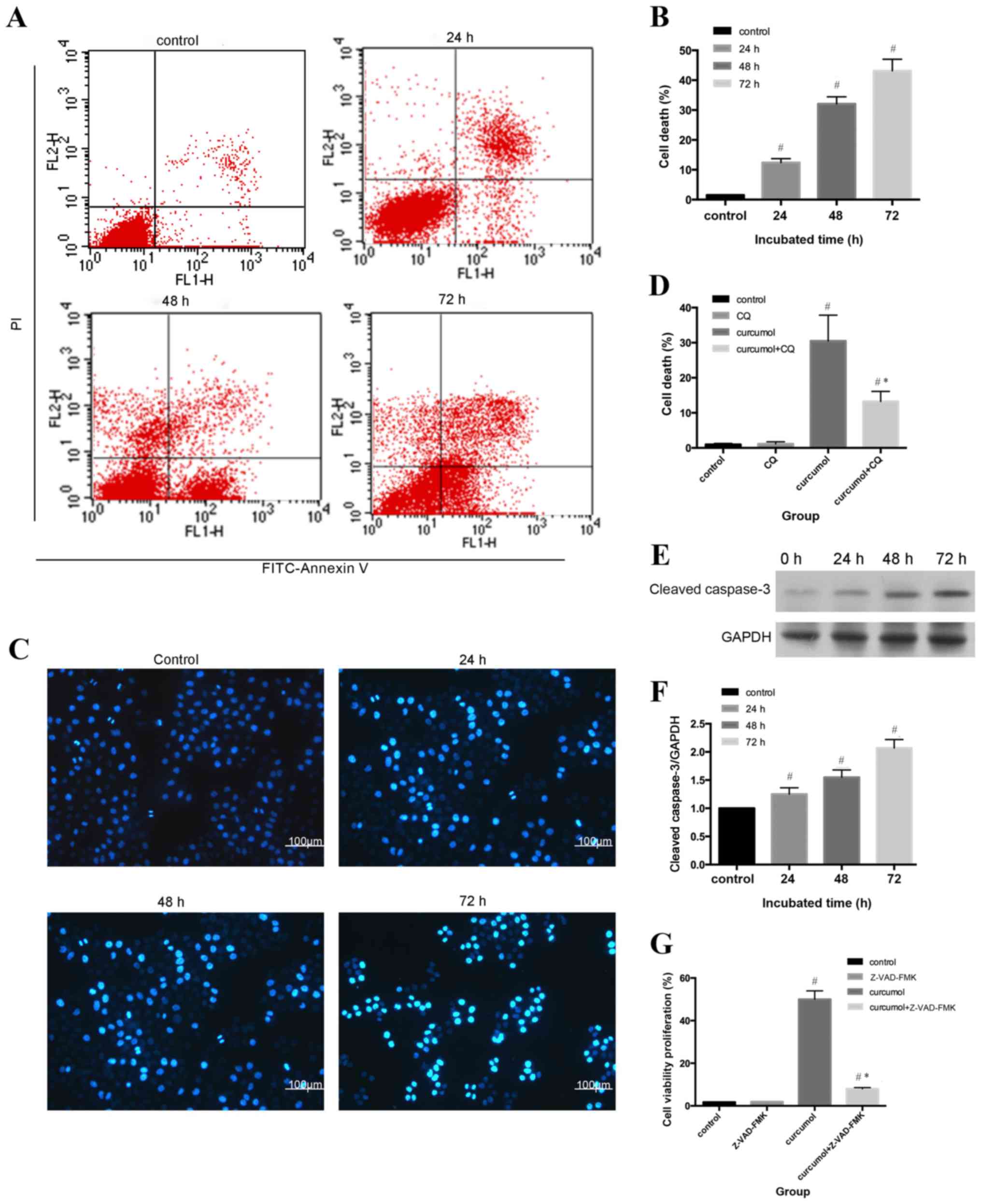
Flow cytometric analysis and Hoechst 33258 staining
were used to evaluate the apoptotic rates of MG-63 cells. As shown
in Fig. 2A and B, compared with the
control group (1.53±0.21%), the apoptosis ratio shifted as the
treatment durations increased (P<0.05). The morphological
alterations of apoptotic nuclei were revealed by Hoechst 33258
staining; the control group demonstrated uniformly stained nuclei,
whereas apoptotic nuclei were condensed or fragmented with light
blue or white fluorescence (Fig. 2C and
D). An increasing number of apoptotic cells with brighter
fluorescence were observed as the treatment durations
lengthened.
In order to further assess the rate of apoptosis
induced by curcumol, western blot analysis was used to determine
the expression levels of cleaved caspase-3. Apoptosis can be
activated by extrinsic stimuli by cell surface death receptors and
intrinsic stimuli by the mitochondrial signaling pathway (28); however, caspase-3 is the core effector
of these two methods and the activation of caspase-3 ultimately
induces irreversible cell death (29,30). As
shown in Fig. 2E and F, cleaved
caspase-3 expression gradually increased as the incubation periods
increased. In order to determine whether cell death was
caspase-dependent, the present study further evaluated the effect
of the pan caspase inhibitor z-VAD-fmk on curcumol-induced cell
viability inhibition. As presented in Fig. 2G, z-VAD-fmk induced the reduction of
cell viability, decrease from 48.8 to 8.99% (Fig. 2G). Taken together, the results
indicated that curcumol induced MG-63 cell death by a
caspase-dependent pathway.
Curcumol-induced autophagy in MG-63
cells
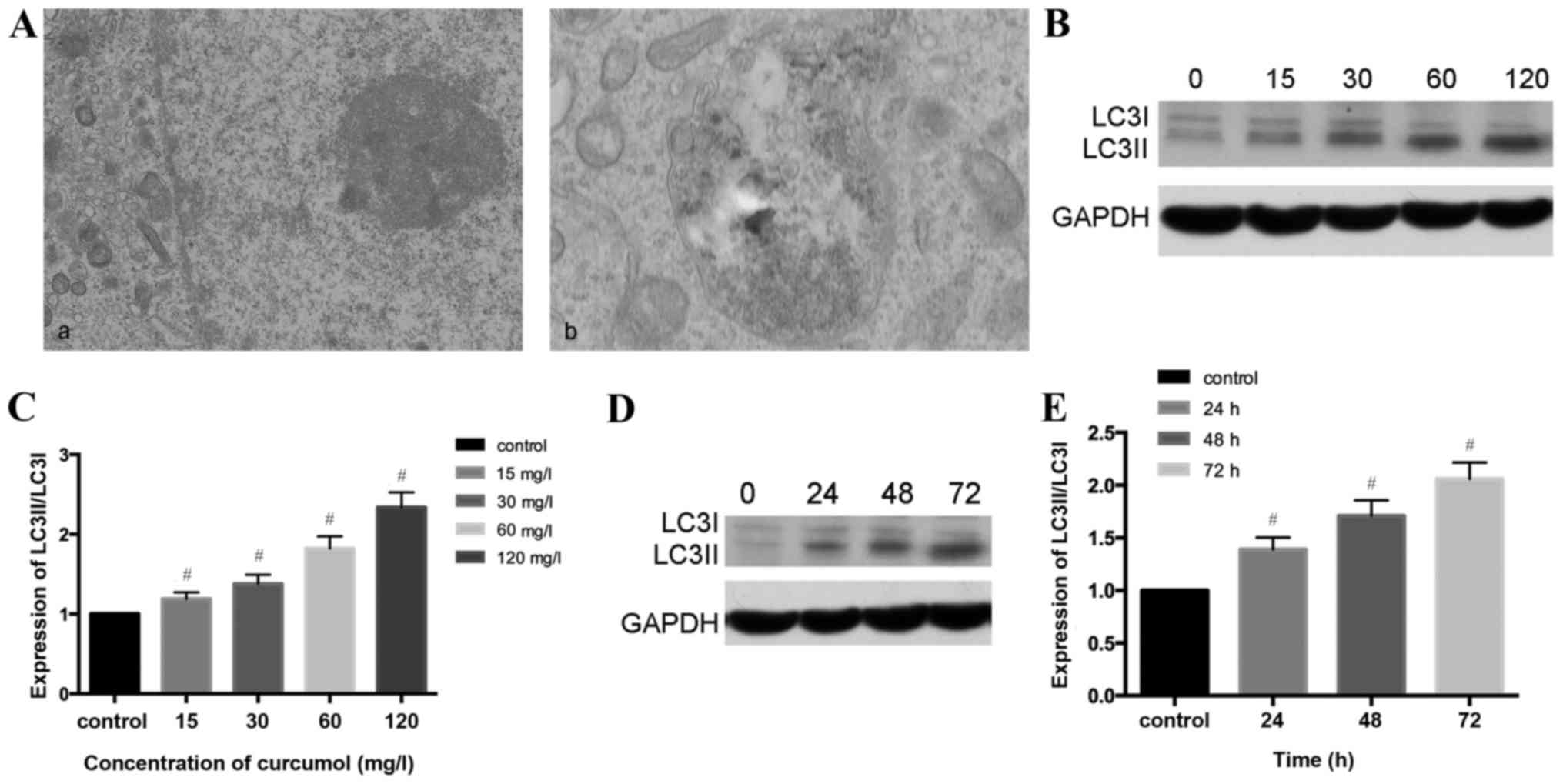
As presented in Fig.
3A, the present study observed that the ultrastructural
characteristics of MG-63 cells treated with curcumol demonstrated a
classical image of autophagic vacuoles sequestrating cytoplasm and
organelles by transmission electron microscopy.
Microtubule-associated protein 1, LC3, similar to yeast
autophagy-related protein 8, generally serve as autophagic markers
that exist as two forms, LC3-I and LC3-II (31). When autophagy is activated, LC3I
transforms into LC3II and LC3II/LC3I is upregulated (32). In order to verify this previous
finding, the present study investigated whether curcumol increased
the LC3II/LC3II level. The results of western blotting demonstrated
that the ratio of LC3II/LC3I shifted in a dose- and time- manner
(Fig. 3B-E).
Autophagy is a dynamic process and could be
accumulated in the condition of inhibition at the final stage, and
therefore autophagic flux should be synchronously monitored. CQ has
been used as an autophagy inhibitor due to destruction of the
acidic environment and subsequent blocking of autophagosome in
combination with lysosomes (31,32). In
the present study autophagy was increased by curcumol treatment and
CQ was used in the following experiment as an autophagy
inhibitor.
GFP-LC3 plasmid transient transfection and altering
the LC3II/LC3I ratios were performed in order to monitor the
efficiency of autophagosomes and to reflect the autophagic flux. As
presented in Fig. 3F and G, compared
with the control group (1.89±0.58%), the CQ treatment alone group
demonstrated no statistical significance, the curcumol group
demonstrated increased percentages of MG-63 cells positive for
LC3II/LC3I (23.12±1.75%) and the combined curcumol and CQ group was
(39.17±2.33%; P<0.05). Western blotting results (Fig. 3H and I) revealed that compared with
the control and CQ treatment groups, the ratio of LC3II/LC3I in the
curcumol treatment group shifted; pre-treatment with CQ followed by
incubation in 63.5 mg/l curcumol increased the ratio and higher
expression levels of LC3I/LC3II were observed (P<0.05). In
conclusion, curcumol treatment induced LC3-II accumulation and
autophagic flux in MG-63 cells.
Inhibition of curcumol-mediated
autophagy attenuates apoptosis
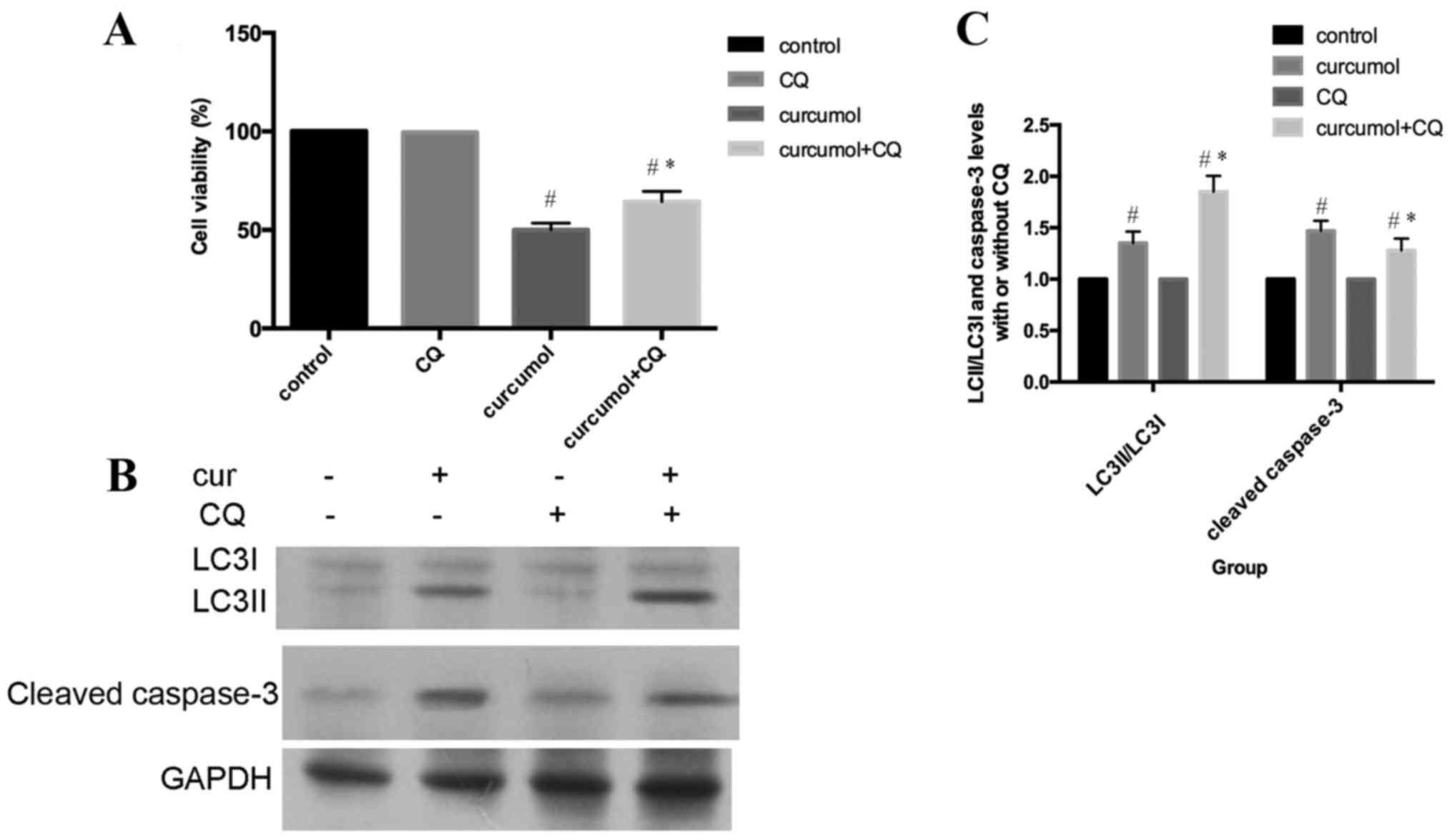
As presented in Fig.
4A, MG-63 cells were incubated with 63.5 mg/l curcumol for 48
h, with or without pre-treatment with CQ for 1 h, and subsequently
cell viability was evaluated by MTT assay. Compared with the
curcumol treatment group, the cell viability was increased in cells
that had also been pre-treated with CQ for 1 h. In addition,
pretreatment with CQ increased LC3-II/LC3I accumulation and
decreased the cleaved caspase-3 expression level (Fig. 4B and C). In conclusion, these results
clearly demonstrated that autophagic inhibition by CQ treatment
attenuated apoptosis in MG-63 cells. Autophagic cell death induced
by curcumol treatment contributed to apoptosis of MG-63 cells.
Underlying mechanism of
curcumol-induced autophagy
An increasing number of studies have indicated that
the c-Jun N-terminal kinase (JNK) signaling pathway and the
activation of mitogen-activated protein kinase (MAPK) family
members may serve an important role in various forms of autophagy
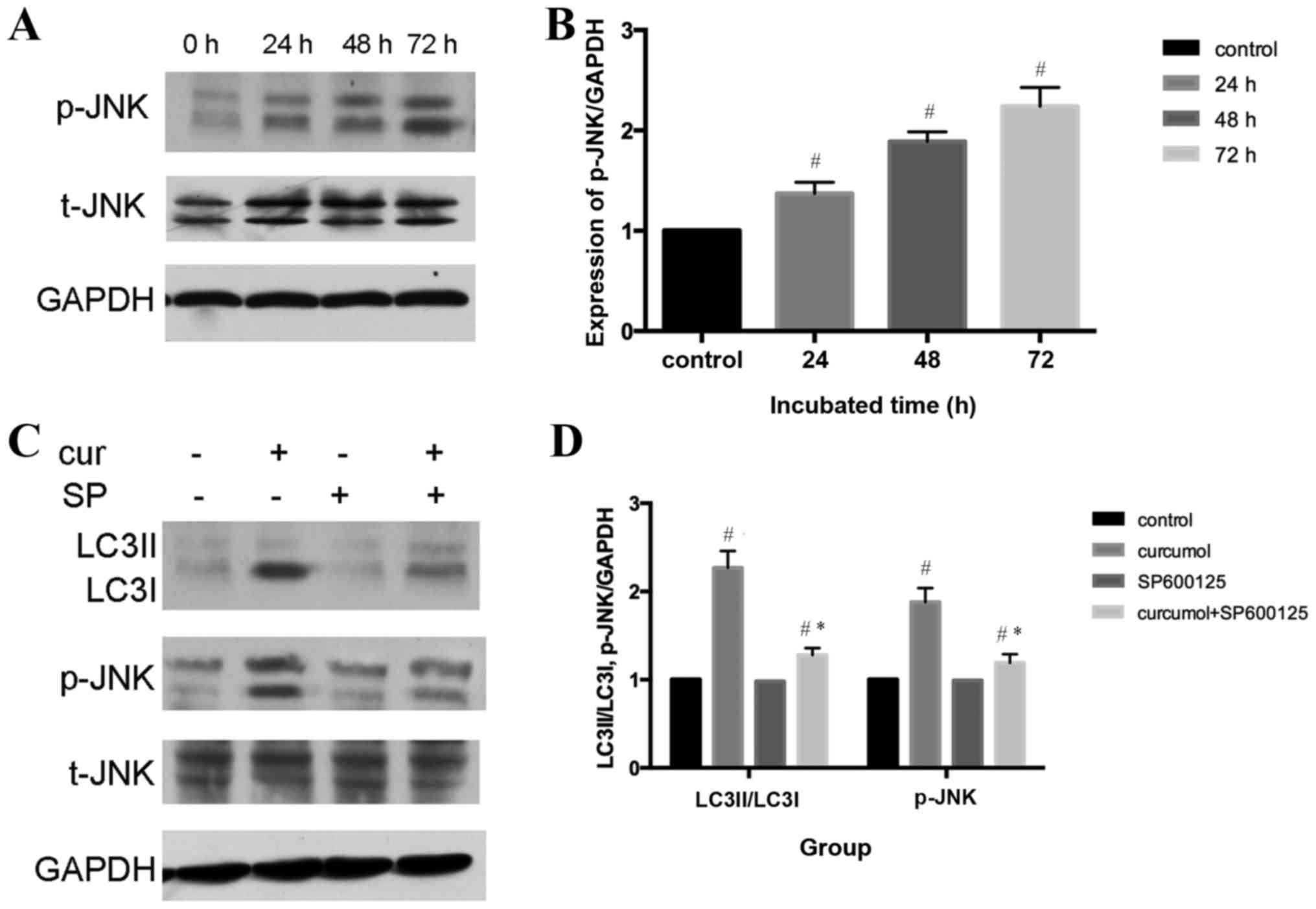
(33,34). The present study demonstrated that
curcumol induced the phosphorylation of JNK in a time-dependent
manner (P<0.05), whereas total-JNK content demonstrated no
change, indicating that curcumol activated the JNK signaling
pathway in MG-63 cells (Fig. 5A and
B). To further confirm the role of the JNK signaling pathway in
curcumol-induced autophagy, a selective inhibitor of JNK, SP600125,
was administered. Furthermore, pretreatment with SP600125
significantly attenuated the expression levels of LC3II/LC3I and
p-JNK compared with the curcumol group (Fig. 5C and D).
Discussion
The results of the present study have demonstrated
that curcumol inhibited cell proliferation of MG-63 cells in a
concentration- and time-dependent manner. Consistent with previous
studies (8–16), flow cytometric analysis and Hoechst
33258 staining revealed that curcumol could trigger apoptosis.
Zhang et al (10) and Chen
et al (16) demonstrated that
curcumol induced apoptosis in lung cancer cells via the
caspase-independent mitochondrial pathway and suppression of B-cell
lymphoma 2. However, the present study revealed that cleaved
caspase-3 accumulated during curcumol treatment in MG-63 cells.
Pre-incubation with the pancaspase inhibitor Z-VAD-FMK attenuated
apoptosis induced by curcumol, which was observed by performing an
MTT assay. Taken together, these results indicated that curcumol
induced caspase-dependent apoptosis.
Autophagy is a highly conserved process in
eukaryotes in which the cytoplasm, including excess or aberrant
organelles, is sequestered into double-membrane vesicles and
delivered to the degradative organelle, the lysosome/vacuole, for
breakdown (35), leading to the
eventual recycling of the resulting macromolecules. While under
pathological conditions, autophagy contributes to the turnover of
long-lived proteins and elimination of damaged or aged organelles
to maintain cell homeostasis (36).
However, extensive autophagy or inappropriate activation of
autophagy results in autophagic cell death (type II programmed cell
death) (37,38).
Previous studies have investigated
curcumol-triggered apoptosis (8–16). To the
best of our knowledge, the present study provided evidence for the
first time that curcumol may trigger autophagy in MG-63 cells: The
classical autophagic double membrane image of MG-63 cells was
observed using a transmission electron microscope; western blot
analysis revealed that the level of LC3II/LC3I shifted in a dose-
and time- manner; furthermore, compared with the curcumol treatment
group, GFP-LC3 plasmids and LC3II/LC3I were markedly upregulated in
the curcumol and CQ treatment group. All these results suggested
that autophagic flux was triggered. Taken together, these results
demonstrated that curcumol may activate autophagy.
The association between autophagy and apoptosis is
complicated. According to various cell types and the effects of
stimuli, autophagy is involved in the promotion or inhibition of
cancer cell death (36). In the
present study, the MTT analysis demonstrated that following
pretreatment with autophagy inhibitor CQ, the cell viability of the
curcumol and CQ treatment group was upregulated. The western blot
analysis also indicated the level of cleaved caspase-3 decreased
and LC3II/LC3I expression increased following CQ pre-incubation for
1 h. Therefore, autophagy curcumol triggered MG-63 cell death.
The JNK signaling pathway, one of the MAPK signal
transduction pathways, serves a pivotal role in regulatory
mechanisms in eukaryotic cells (39).
Once activated by upstream kinases, JNK mediates various
physiological processes including, inflammation, stress, cell
growth, cell development, differentiation and cell death (40). An increasing amount of evidence
suggests that the JNK signaling pathway is an important regulator
of autophagy under various conditions (41). Thus, the present study investigated
whether curcumol-induced autophagy involves this pathway. As shown
in Fig. 5A and B, it was demonstrated
that curcumol activated the JNK signaling pathway, as p-JNK protein
expression level was upregulated in a time-dependent manner. In
order to determine whether the JNK signaling pathway participated
in curcumol-induced autophagy, selective inhibitors of JNK were
administered. As shown in Fig. 5C and
D, pretreatment with JNK inhibitor SP600125 inhibited the
phosphorylation of the JNK signaling pathway and LC3-II
accumulation.
In conclusion, the present findings suggest that
curcumol is a potent antitumor agent and exerts its antineoplastic
action by inducing cell apoptosis and autophagic cell death via
activation of the JNK signaling pathway. These results may be
useful for further development of the clinical application of this
compound in treating osteosarcoma.
References
|
1
|
Anderer U, Nöhren H, Koch I, Harms D and
Dietel M: Organization of the pediatric tumor cell bank of the
society of pediatric oncology and hematology (GPOH). Klin Padiatr.
210:1–9. 1998.(In German). View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferrari S and Palmerini E: Adjuvant and
neoadjuvant combination chemotherapy for osteog enic sarcoma. Curr
Opin Oncol. 19:341–346. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Whelan J, Seddon B and Perisoglou M:
Management of osteosarcoma. Curr Treat Options Oncol. 7:444–455.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Durfee RA, Mohammed M and Luu HH: Review
of osteosarcoma and current management. Rheumatol Ther. 3:221–243.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen LX, Zhang H, Zhao Q, Yin SY, Zhang Z,
Li TX and Qiu F: Microbial transformation of curcumol by
Aspergillus niger. Nat Prod Commun. 8:149–152. 2013.PubMed/NCBI
|
|
6
|
Chen X, Zong C, Gao Y, Cai R, Fang L, Lu
J, Liu F and Qi Y: Curcumol exhibits anti-inflammatory properties
by interfering with the JNK mediated AP-1 pathway in
lipopolysaccharide-activated RAW264.7 cells. Eur J Pharmacol.
723:339–345. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li J, Mao C and Li L, Ji D, Yin F, Lang Y,
Lu T, Xiao Y and Li L: Pharmacokinetics and liver distribution
study of unbound curdione and curcumol in rats by microdialysis
coupled with rapid resolution liquid chromatography (RRLC) and
tandem mass spectrometry. J Pharm Biomed Anal. 95:146–150. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zou L, Liu W and Yu L: Beta-elemene
induces apoptosis of K562 leukemia cells. Zhonghua Zhong Liu Za
Zhi. 23:196–198. 2001.(In Chinese). PubMed/NCBI
|
|
9
|
Zhou HY, Hou JS and Wang Y: Dose-and
time-dependence of elemene in the induction of apoptosis in two
glioma cell lines. Zhonghua Zhong Liu Za Zhi. 28:270–271. 2006.(In
Chinese). PubMed/NCBI
|
|
10
|
Zhang W, Wang Z and Chen T: Curcumol
induces apoptosis via caspases-independent mitochondrial pathway in
human lung adenocarcinoma ASTC-a-1 cells. Med Oncol. 28:307–314.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu HY, Pen AB, Liao AJ and Shi W:
Preliminary research on the mechanism of apoptosis hepatic stellate
cells induced by zedoary turmeric oil. Zhonghua Gan Zang Bing Za
Zhi. 17:790–791. 2009.(In Chinese). PubMed/NCBI
|
|
12
|
Shi H, Tan B, Ji G, Lu L4, Cao A, Shi S
and Xie J: Zedoary oil (Ezhu You) inhibits proliferation of AGS
cells. Chin Med. 8:132013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang J, Huang F, Bai Z, Chi B, Wu J and
Chen X: Curcumol inhibits growth and induces apoptosis of
colorectal cancer LoVo cell line via IGF-1R and p38 MAPK Pathway.
Int J Mol Sci. 16:19851–19867. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang J, Chen X and Zeng JH: Effect of
curcumol on proliferation and apoptosis of nasopharyngeal carcinoma
cellline CNE-2. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 27:790–792.
2011.(In Chinese). PubMed/NCBI
|
|
15
|
Xu LC, Bian KM, Liu ZM and Wang G: The
inhibitory effect of the curcumol on women cancer cells and
synthesis of RNA. Tumor. 25:570–572. 2005.
|
|
16
|
Chen G, Wang Y, Li M, Xu T, Wang X, Hong B
and Niu Y: Curcumol induces HSC-T6 cell death through suppression
of Bcl-2: Involvement of PI3K and NF-κB pathways. Eur J Pharm Sci.
65:21–28. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li X, Wang G, Zhao J, Ding H, Cunningham
C, Chen F, Flynn DC, Reed E and Li QQ: Antiproliferative effect of
beta-elemene in chemoresistant ovarian carcinoma cells is mediated
through arrest of the cell cycle at the G2-M phase. Cell Mol Life
Sci. 62:894–904. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao J, Li QQ, Zou B, Wang G, Li X, Kim
JE, Cuff CF, Huang L, Reed E and Gardner K: In vitro combination
characterization of the new anticancer plant drug β-elemene with
taxanes against human lung carcinoma. Int J Oncol. 31:241–252.
2007.PubMed/NCBI
|
|
19
|
Lin Y, Wang K, Hu C, Lin L, Qin S and Cai
X: Elemene injection induced autophagy protects human hepatoma
cancer cells from starvation and undergoing apoptosis. Evid Based
Complement Alternat Med. 2014:6375282014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu J, Hu XJ, Jin B, Qu XJ, Hou KZ and Liu
YP: β-Elemene induces apoptosis as well as protective autophagy in
human non-small-cell lung cancer A549 cells. J Pharm Pharmacol.
64:146–153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu J, Zhang Y, Qu J, Xu L, Hou K, Zhang
J, Qu X and Liu Y: β-Elemene-induced autophagy protects human
gastric cancer cells from undergoing apoptosis. BMC Cancer.
11:1832011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou K, Wang L, Cheng R, Liu X, Mao S and
Yan Y: Elemene increases autophagic apoptosis and drug sensitivity
in human cisplatin (DDP)-resistant lung cancer cell line
SPC-A-1/DDP By inducing beclin-1 expression. Oncol Res. May
23–2017.(Epub ahead of print). View Article : Google Scholar
|
|
23
|
Mu L, Wang T, Chen Y, Tang X, Yuan Y and
Zhao Y: β-Elemene enhances the efficacy of gefitinib on
glioblastoma multiforme cells through the inhibition of the EGFR
signaling pathway. Int J Oncol. 49:1427–1436. 2016.PubMed/NCBI
|
|
24
|
Raff MC, Barres BA, Burne JF, Coles HS,
Ishizaki Y and Jacobson MD: Programmed cell death and the control
of cell survival. Philos Trans R Soc Lond B Biol Sci. 345:265–268.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bursch W, Ellinger A, Gerner C, Fröhwein U
and Schulte-Hermann R: Programmed cell death (PCD) Apoptosis,
autophagic PCD, or others? Ann N Y Acad Sci. 926:1–12. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lockshin RA and Zakeri Z: Apoptosis,
autophagy, and more. Int J Biochem Cell Biol. 236:2405–2419. 2004.
View Article : Google Scholar
|
|
27
|
Ogier-Denis E and Codogno P: Autophagy: A
barrier or an adaptive response to cancer. Biochim Biophys Acta.
1603:113–128. 2003.PubMed/NCBI
|
|
28
|
Nicholson DW: Caspase structure,
proteolytic substrates, and function during apoptotic cell death.
Cell Death Differ. 6:1028–1042. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Thornberry NA: Caspases: Key mediators of
apoptosis. Chem Biol. 5:R97–R103. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xiong S, Mu T, Wang G and Jiang X:
Mitochondria-mediated apoptosis in mammals. Protein Cell.
5:737–749. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Poole B and Ohkuma S: Effect of weak bases
on the intralysosomal pH in mouse peritoneal macrophages. J Cell
Biol. 90:665–669. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nilsson JR: Does chloroquine, an
antimalarial drug, affect autophagy in Tetrahymena pyriformis? J
Protozool. 39:9–16. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou YY, Li Y, Jiang WQ and Zhou LF:
MAPK/JNK signaling: A potential autophagy regulation pathway.
Biosci Rep. 35:pii: e001992015.
|
|
34
|
Byun JY, Yoon CH, An S, Park IC, Kang CM,
Kim MJ and Lee SJ: The Rac1/MKK7/JNK pathway signals upregulation
of Atg5 and subsequent autophagic cell death in response to
oncogenic Ras. Carcinogenesis. 30:1880–1888. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Morgan-Bathke M, Lin HH, Ann DK and
Limesand KH: The role of autophagy in salivary gland homeostasis
and stress responses. J Dent Res. 94:1035–1040. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Apel A, Zentgraf H, Büchler MW and Herr I:
Autophagy-A double-edged sword in oncology. Int J Cancer.
125:991–995. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rami A and Kögel D: Apoptosis meets
autophagy-like cell death in the ischemic penumbra: Two sides of
the same coin? Autophagy. 4:422–426. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Su Z, Yang Z, Xu Y, Chen Y and Yu Q:
Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol
Cancer. 14:482015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Barr RK and Bogoyevitch MA: The c-Jun
N-terminal protein kinase family of mitogen-activated protein
kinases (JNK MAPKs). Int J Biochem Cell Biol. 33:1047–1063. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wei Y, Pattingre S, Sinha S, Bassik M and
Levine B: JNK1-mediated phosphorylation of Bcl-2 regulates
starvation-induced autophagy. Mol Cell. 30:678–688. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sui X, Kong N, Ye L, Han W, Zhou J, Zhang
Q, He C and Pan H: p38 and JNK MAPK pathways control the balance of
apoptosis and autophagy in response to chemotherapeutic agents.
Cancer Lett. 344:174–179. 2014. View Article : Google Scholar : PubMed/NCBI
|