Introduction
Breast cancer is a common type of malignancy in the
world and a major cause of mortality in females between 30 and 59
years of age (1). Breast cancer is a
heterogeneous disease in terms of histology, pathology, and genetic
and molecular profiles (2). Despite
diagnostic and therapeutic advances, breast cancer patients still
often exhibit relapse or metastasis subsequent to therapy (3).
Tumors are morphologically heterogeneous, composed
of undifferentiated and differentiated cells (4). Cancer stem cells (CSCs) have been
identified as a subpopulation within the tumor possessing the
ability to self-renew and differentiate into non-tumorigenic cell
populations that constitute the bulk of the tumor (5). CSCs have been associated with tumor
initiation, therapy resistance and tumor recurrence. CSCs are a
major problem for cancer therapy, and the elimination of CSCs is
required for an effective treatment (6). The presence of CSC population in breast
cancer has been demonstrated in several studies (7,8). Breast
cancer stem cells (BCSCs) were first isolated by Al-Hajj et
al (9) in 2003 from the pleural
effusions of a patient. Specific cell surface markers and
biomarkers are used to identify and isolate BCSCs. The adhesion
molecule cluster of differentiation (CD) 44 is a multifunctional
cell surface transmembrane glycoprotein that serves a role in cell
adhesion, proliferation, differentiation, motility and migration
(10). In breast cancer,
CD44+/CD24− expression was demonstrated as
prospective phenotype to isolate BCSCs. Al-Hajj et al
(9) reported that breast cancer cells
exhibiting an increased expression of
CD44+/CD24− were able to form tumors when
injected into immunodeficient mice.
Cyclin-dependent kinases (CDKs) serve an essential
role in the control of the cell cycle, and are associated with
cytoskeletal dynamics, epigenetic regulation, controlling stem cell
self-renewal, regulating metabolism, cell migration, regulation of
transcription, DNA damage, and genomic and chromosomal instability
(11). The dysregulation of CDK
expression contributes to the loss of normal cell cycle control,
which leads to the formation and progression of cancer (12). Therefore, the inhibition of CDKs by
small-molecule CDK inhibitors may be an effective treatment of
cancer. The dysregulation of cyclin D and the CDK pathway in cancer
cells may inhibit senescence and promote cellular proliferation
(13). By using various different
mechanisms, malignant cells may increase cyclin D-dependent
activity. The cyclin D-CDK4/6-retinoblastoma pathway controls the
cell cycle restriction point, and is commonly dysregulated in
breast cancer, making it a possible target for anticancer therapy
(14).
Flavopiridol is a semisynthetic flavonoid that was
the first CDK inhibitor used in clinical trials (15). Flavopiridol exhibits an antitumor
effect against a variety of tumor types, including several solid
tumors, through cytostatic activity, and induces cell cycle arrest
and apoptosis (16). This flavonoid
is a promising anticancer drug that is undergoing phase I and II
clinical trials for chronic myeloid leukemia and pancreatic cancer
(17,18). Our previous study demonstrated that
flavopiridol induced growth inhibition and apoptosis in
CD133+/CD44+ prostate CSCs (19).
BCSCs have been proposed to be responsible for
numerous properties of breast cancer such as resistance, metastatic
properties and recurrence (20).
Conventional anticancer therapies may kill the majority of the
cancer cells, but CSCs are not affected by these therapies
(21). For a more effective treatment
of breast cancer, it may be necessary to target CSCs. Genome-wide
gene expression profiling based on microarray analysis is a
powerful tool to elucidate the possible mechanisms of cancer drugs.
The present study aimed to investigate the cytotoxic effects and
underlying mechanism of action of flavopiridol against human breast
CSCs.
Materials and methods
Cell culture conditions and
reagents
Human breast cancer MCF7 cells were obtained from
Interlab Cell Line Collection (Genova, Italy) and were grown in
monolayer cell culture in RPMI 1640 culture medium (Lonza Group AG,
Basel, Switzerland) containing 10% heat-inactivated fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 1%
penicillin and 1% streptomycin (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). The cells were cultured in 25-cm2
polystyrene flasks (Corning Life Sciences, Corning, NY, USA) and
incubated for 48 h at 37°C in a humidified atmosphere of 5%
CO2. Flavopiridol (Sigma-Aldrich; Merck KGaA) was
prepared as 10 mM stock solution in dimethyl sulfoxide (DMSO), and
the final volume of DMSO did not exceed 0.1% of the total
incubation volume and was not cytotoxic to the tumor cells at these
concentrations (data not shown).
Fluorescence-activated cell sorting
(FACS)
To sort the CSCs subpopulations in the human breast
cancer MCF7 cell line, the antibodies of expressed surface markers
CD44+/CD24−, anti-CD44 conjugated to
fluorescein isothiocyanate (10 µl/106 cell; FITC; cat.
no. 555478; BD Biosciences, Franklin Lakes, NJ, USA) and anti-CD24
conjugated to phycoerythrin (10 µl/106 cell; PE; clone
32D12; Miltenyi Biotec GmbH, Bergisch Gladbach, Germany) were used.
The MCF7 cells were seeded and grown to 80% confluence. The cells
were detached using a non-enzymatic cell dissociation solution
(Sigma-Aldrich; Merck KGaA) and resuspended in RPMI 1640 culture
medium. A total of ~5×104 cells were incubated with
anti-CD44-fluorescein isothiocyanate (FITC; clone G44 26; BD
Biosciences, Franklin Lakes, NJ, USA) and anti-CD24-phycoerythrin
(PE; clone 32D12; Miltenyi Biotec GmbH, Bergisch Gladbach, Germany)
in FACS stain buffer (cat. no. 554657; BD Pharmingen, Franklin
Lakes, NJ, USA) for 15 min at 4°C. After 15 min, the cells were
washed with the above FACS wash buffer and resuspended in FACS
stain buffer (cat. no 554657, BD Pharmingen, Franklin Lakes, NJ,
USA) to a density of 107 cells/ml. The cells were sorted
into a CD44+/CD24− population (sorted cells)
using a FACSAria flow cytometer (BD Biosciences).
Analysis of cell viability
The viability of the cells following treatment was
determined using the Muse® Count & Viability kit
(Muse Cell Analyzer; EMD Millipore, Billerica, MA, USA) according
to the protocol of the manufacturer. The cells were seeded in
triplicate in 6-well plates at a density of 1×104
cells/well. Subsequent to a 24-h incubation, the cells were exposed
to 500, 750 and 1,000 nM flavopiridol. The plates were then
incubated at 37°C in a 5% CO2 incubator for 24, 48 and
72 h. Subsequent to incubation, all cells were collected and
diluted with PBS. In total, 50 µl of the cell suspension was then
added to 450 µl Muse® Count & Viability reagent
(dilution, 10X), incubated for 5 min at room temperature and
analyzed using the Muse Cell Analyzer. Data were presented as
proportional viability (%) by comparing the treated group with the
untreated cells.
RNA isolation and microarray
analysis
The BCSCs were treated with a dose of flavopiridol
equivalent to its half maximal inhibitory concentration
(IC50). Total RNA was extracted from the treated and
untreated cells using the RNeasy Mini kit (Qiagen, Inc., Valencia,
CA, USA) according to the protocol of the manufacturer.
Biotin-labeled RNA samples for hybridization on Illumina Human
HT-12 v4 Expression BeadChip (Illumina, Inc., San Diego, CA, USA)
were prepared according to the recommended sample labeling
procedure of Illumina, Inc. A total of 250 ng total RNA was used
for cDNA synthesis, followed by an amplification/labeling step to
synthesize biotin-labeled cRNA. The quality of the cRNA was
controlled using the Agilent 2100 Bioanalyzer (Agilent
Technologies, Inc., Santa Clara, CA, USA). Hybridization was
performed at 58°C in GEX-HCB buffer (Illumina, Inc.) at a
concentration of 150 ng cRNA/µl. The BeadChips were subsequently
washed, blocked and conjugated with cyanine 3-streptavidin (Thermo
Fisher Scientific, Inc.). The microarrays were scanned in the iScan
System (Illumina, Inc.). The obtained amplification data
(fold-changes in the quantification cycle values of all the genes)
were processed in Agilent GeneSpring Data Analysing Software
(Agilent Technologies, Inc.) and >2 fold-change was used for
filtering criteria.
Statistical analysis
The statistical software package SPSS version 20.0
for Windows (IBM Corp., Armonk, NY USA) was used for all
statistical analysis. All experiments were performed independently
three times. Statistical analysis was tested by one-way analysis of
variance, followed by Tukey's or Dunnett's post hoc tests. All data
are presented as mean ± standard deviation from 3 independent
experiments. P<0.05 was considered to indicate a statistically
significant difference.
Results
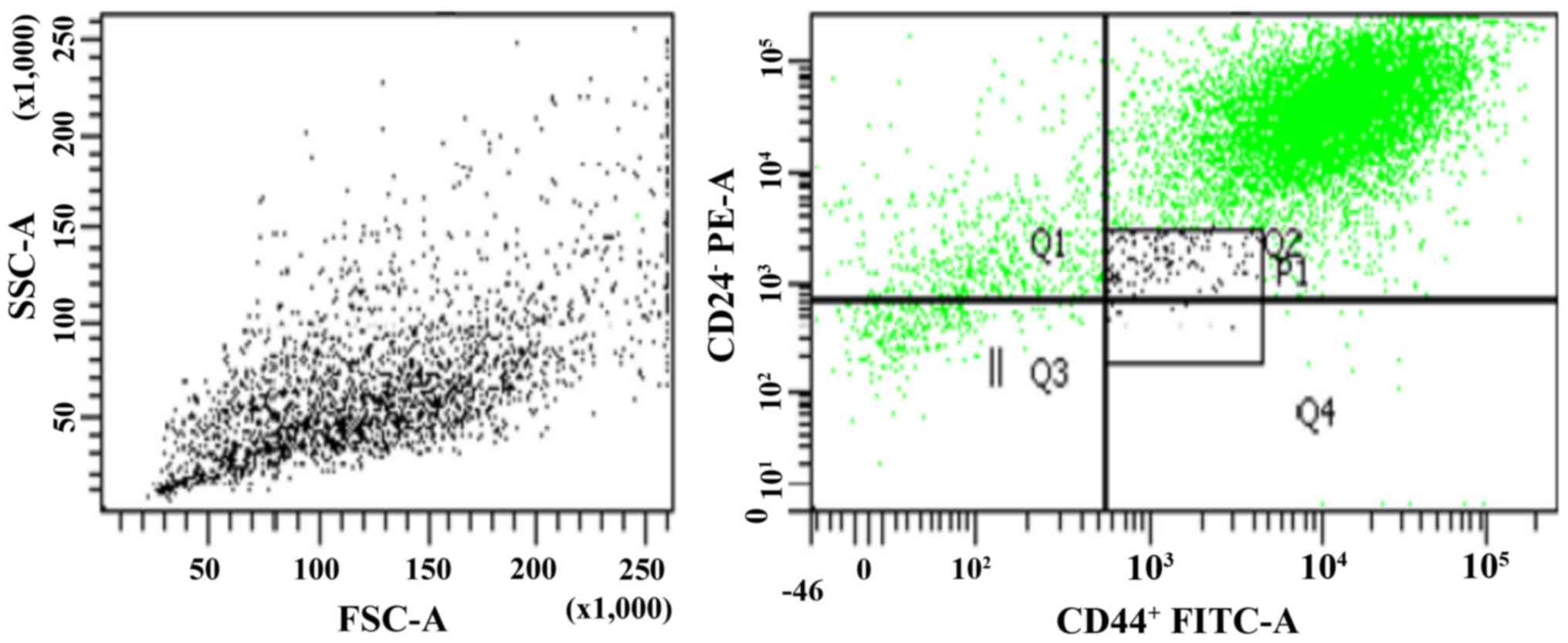
Sorting breast cancer MCF7 cells and
purity of the CD44+/CD24− sorted
subpopulations
Human breast cancer MCF7 cells were separated with
FACS, yielding a CD44+/CD24− population
(Fig. 1). The present study obtained
MCF7 CSC and non-CSC subpopulations. According to the results, the
mean percentage of MCF7 CSCs and non-CSCs were 1.6 and 98.4%,
respectively. The purity of the CSCs samples was tested with
anti-CD44 and anti-CD24 antibodies. The sorting rate analysis and
purity of the cells were evaluated sequentially, and the rate was
96.7±5.4% for the sorted cells. To confirm the flow cytometry
analyses, the cells were re-evaluated following sorting, and the
analyses were repeated subsequent to one passage. The results
revealed that the cell purity following sorting was >90%.
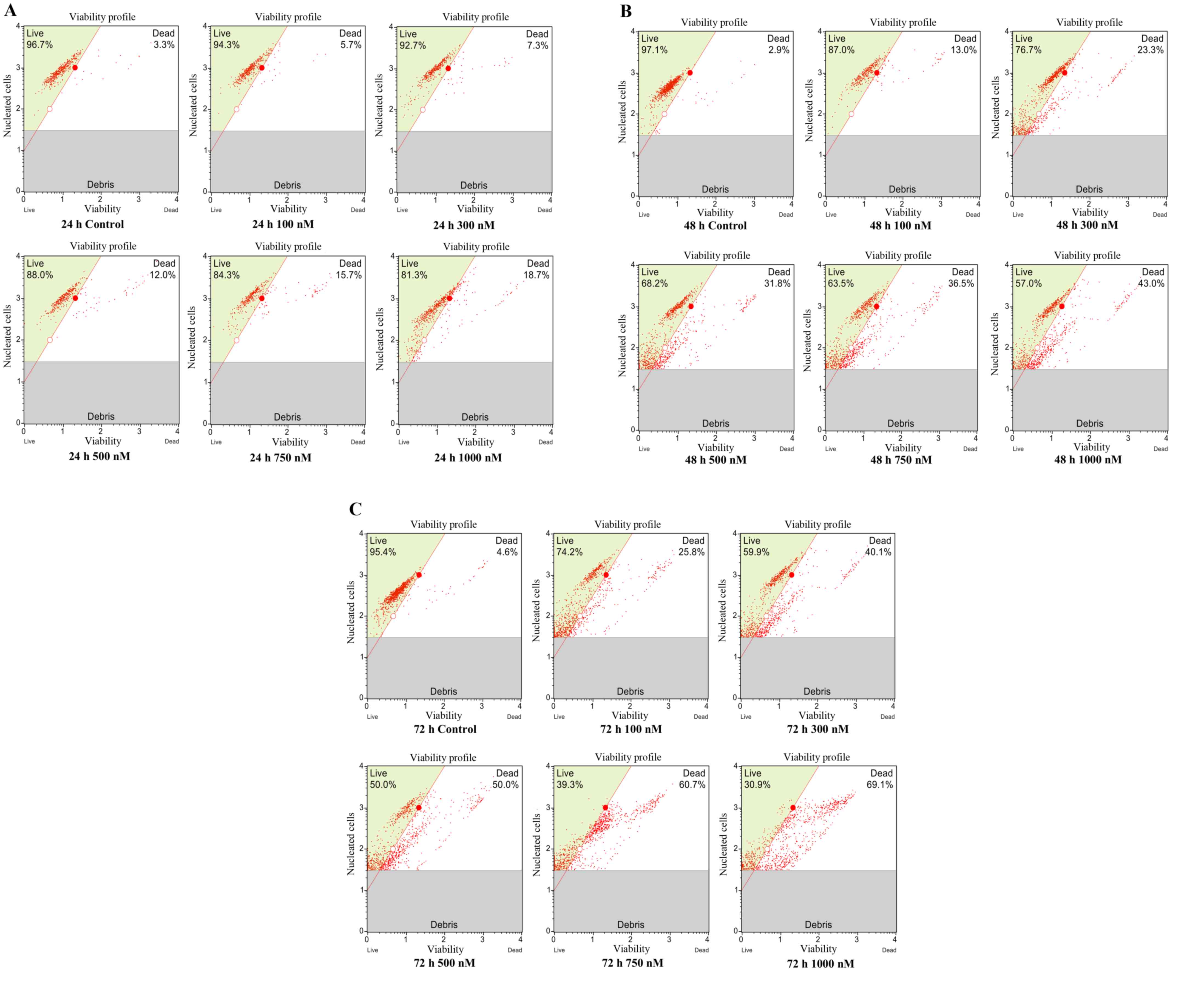
Increasing cytotoxicity of
CD44+/CD24− BCSCs with flavopiridol
Cytotoxicity assays were performed to determine the
therapeutic effect of flavopiridol. MCF7 CSCs were exposed to
100–1,000 nM flavopiridol for 24, 48 and 72 h, and the percentage
of viable cells in the samples was determined by a cell viability
assay. Flavopiridol reduced the cell viability of CSCs in a time-
and concentration-dependent manner (Fig.
2A-C). According to the data, there were no significant
decreases in cell viability at the low doses (100 and 300 nM) of
flavopiridol treatment for 24 h compared with that of the untreated
cells (P=0.642). After 48 h of treatment, flavopiridol
significantly reduced the cell viability of BCSCs at 500, 750 and
1,000 nM compared with that of the untreated cells (P=0.000). After
72 h of treatment with flavopiridol, the IC50 was
calculated as 500 nM.
Microarray analysis for the
identification of differentially expressed genes in MCF7
CD44+/CD24− cells treated with
flavopiridol
To analyze the molecular mechanisms underlying the
anticancer effect of flavopiridol in BCSCs, the MCF7
CD44+/CD24− cells were treated with 500 nM
flavopiridol for 72 h. To identify flavopiridol-regulated genes and
determine the possible mechanism underlying the differential role
of flavopiridol on the growth of MCF7
CD44+/CD24− cells, global gene expression
profiling was undertaken following treatment with flavopiridol
using the Illumina Human HT-12 v4 Expression BeadChip. According to
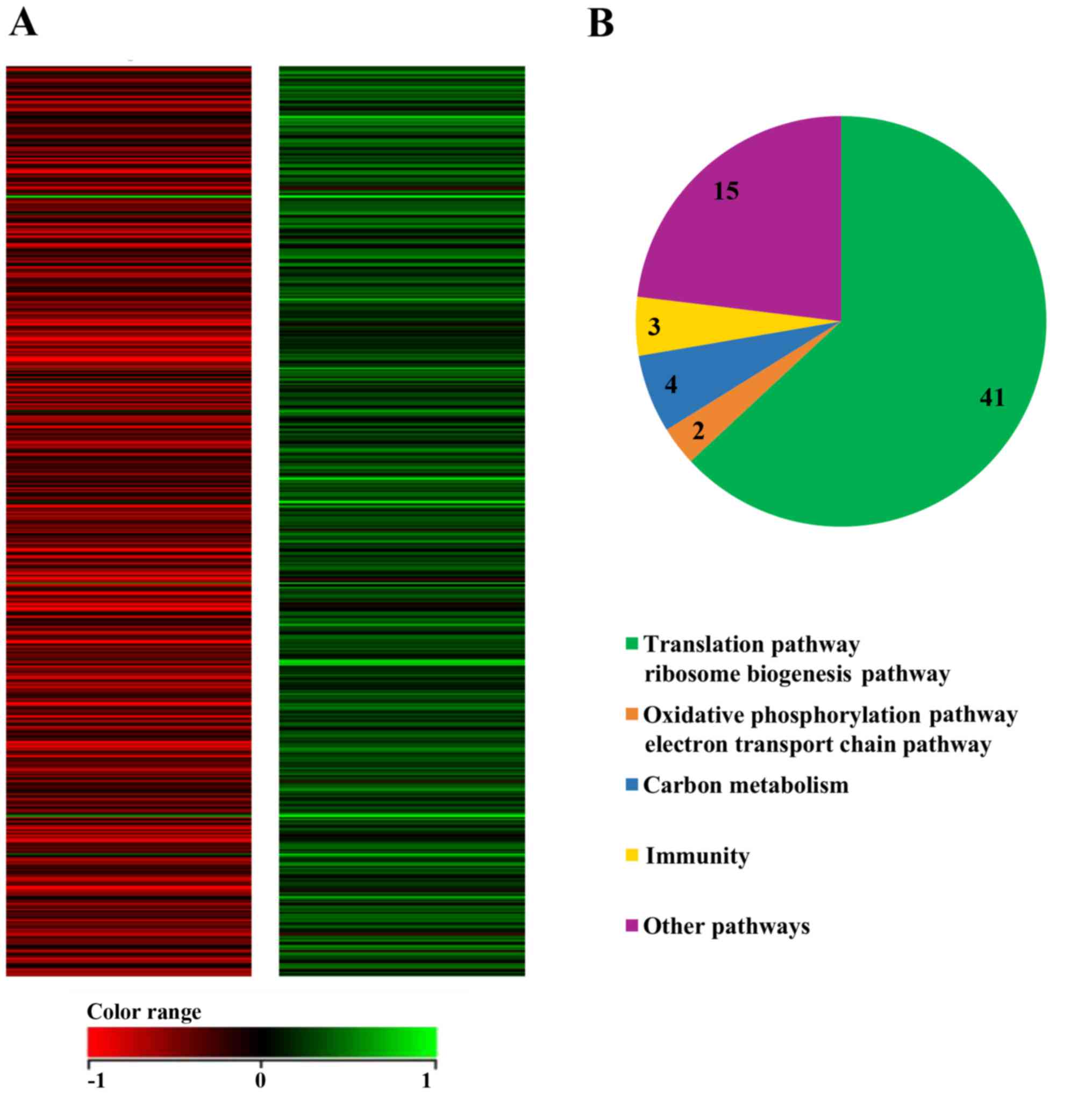
the results of microarray analysis, 65 genes were identified as
significantly affected subsequent to treatment with flavopiridol,
since the expression of 57 genes decreased and the expression of 8
genes increased compared with that in untreated cells at 72 h
(Table I).
 | Table I.Changes in the expression of
upregulated and downregulated genes following treatment with
flavopiridol. |
Table I.
Changes in the expression of
upregulated and downregulated genes following treatment with
flavopiridol.
| A, Translation
pathway and ribosome biogenesis pathway |
|---|
|
|---|
| Probe ID | Symbol |
Fold-changea | Regulation | Definition |
|---|
| 4920193 | RPL27A | −2.7026234 | Down | RPL27a |
| 6060356 | RPL13A | −2.1516730 | Down | RPL13a |
| 3360228 | RPS20 | −2.0229893 | Down | RPS20 |
| 5290082 | RPLP1 | −2.1037197 | Down | RiP, large, P1 |
| 1710369 | RPL3 | −2.5006313 | Down | RPL3, transcript
variant 2 |
| 620754 | RPS5 | −2.3889322 | Down | RPS5 |
| 7040095 | RPL17 | −2.0128388 | Down | RPL17, transcript
variant 2 |
| 990273 | RPL37A | −2.2212677 | Down | RP L37a |
| 3060477 | RPL8 | −2.4217634 | Down | RPL8, transcript
variant 2 |
| 3610241 | RPL19 | −3.2778310 | Down | RPL19 |
| 3800332 | RPS25 | −2.3831854 | Down | RPS25 |
| 5260682 | RPS14 | −2.5092149 | Down | RPS14, transcript
variant 2 |
| 5220037 | RPS2 | −3.9913297 | Down | RPS2 |
| 6960181 | RPS12 | −2.9727620 | Down | RPS12 |
| 7510482 | RPS4X | −2.1651378 | Down | RPS4, X-linked |
| 20021 | RPS15 | −2.4493800 | Down | RPS15 |
| 5560349 | RPS11 | −2.6253710 | Down | RRPS11 |
| 5890730 | RPS26L | −2.8872151 | Down | Predicted: Homo
sapiens 40S RPS26-like |
| 510195 | RPL27 | −2.4229383 | Down | RPL27 |
| 840647 | RPL36 | −2.7411752 | Down | RPL36, transcript
variant 1 |
| 4250445 | RPL4 | −2.0928760 | Down | RPL4 |
| 6250097 | RPS9 | −2.3448272 | Down | RPS9 |
| 1410537 | RPSA | −2.1424713 | Down | RPSA, transcript
variant 1 |
| 6270546 | RPS6 | −2.3560820 | Down | RPS6 |
| 6590377 | RPS26 | −2.1171474 | Down | RPS26 |
| 4250445 | RPL4 | −2.0012200 | Down | RPL4 |
| 3610309 | LOC653881 | −2.6014566 | Down | Predicted: Similar
to RPL3 |
| 2490450 | LOC91561 | −2.2661705 | Down | Predicted: Similar
to RPS2, transcript variant 3 |
| 3440670 | LOC402251 | −2.3879724 | Down | Predicted: Similar
to eukaryotic translation elongation factor 1 α 2 |
| 4060446 | LOC649150 | −3.2092447 | Down | Predicted: Similar
to eukaryotic translation elongation factor 1 α 2 |
| 1440398 | LOC644511 | −2.2366867 | Down | Predicted: Similar
to RPL13a, transcript variant 1 |
| 1570491 | LOC648000 | −2.3058624 | Down | Predicted: Similar
to 60S RPL7, transcript variant 1 |
| 2320494 | LOC653314 | −3.1144562 | Down | Homo sapiens
similar to RPL19 |
| 6280021 | LOC441876 | −2.8330393 | Down | Predicted: Similar
to 40S RPS16, |
| 870593 | LOC285053 | −2.3860030 | Down | Predicted: Similar
to RPL18a, transcript variant 1 |
| 5720747 | LOC441775 | −2.6068625 | Down | Predicted: Similar
to 60S RPL18 |
| 2190546 | LOC388654 | −2.2629400 | Down | Predicted: Similar
to laminin receptor 1 (RPSA) |
| 5720747 | LOC441775 | −2.6068625 | Down | Predicted: Similar
to 60S RPL18 |
| 6330373 | EEF1B2 | −2.3026142 | Down | Eukaryotic
translation elongation factor 1 β 2, transcript variant 1 |
| 3850121 | EEF1A1 | −3.1777650 | Down | Eukaryotic
translation elongation factor 1 α 1 |
|
| B, Oxidative
phosphorylation and electron transport chain pathway |
|
| 3850110 | COX6A1 | −2.1455740 | Down | Cytochrome c
oxidase subunit Vıa polypeptide 1 |
| 4490259 | COX8A | −2.9969997 | Down | Cytochrome c
oxidase subunit 8A (ubiquitous) |
|
| C, Carbon
metabolism |
|
| 2760358 | NME1-2 | −2.3299380 | Down | NME1-NME2
readthrough |
| 1940360 | TPI1 | −2.8388138 | Down | Triosephosphate
isomerase 1 |
| 6590253 | ALDOA | −2.1273860 | Down | ALDOA |
| 6520128 | GPX4 | −2.1577030 | Down | Glutathione
peroxidase |
|
| D, Mammary gland
development pathway |
|
| 5860138 | RIPK4 | −2.1919790 | Down |
Receptor-interacting serine-threonine
kinase 4 |
|
| E, G
protein-mediated signaling pathway via Gα12/Gα13 family |
|
| 2850402 | PFN1 | −2.4898353 | Down | Profilin 1 |
|
| F, Tumor
necrosis factor-mediated signaling pathway |
|
| 670673 | BCL2L1 | −2.0002713 | Down | BCL2-like 1,
nuclear gene encoding mitochondrial |
|
| G, Signaling
pathway pertinent to immunity |
|
| 1980594 | FTHL8 | −3.2119188 | Down | Ferritin, heavy
polypeptide-like 8 |
| 2970431 | FTHL7 | −4.2565985 | Down |
FTHL7 |
|
| H, Toll-like
receptor signaling pathway |
|
| 3840154 | SPP1 | −3.0712519 | Down | SPP1, transcript
variant 1 |
|
| I, Signaling by
TGF-β receptor complex |
|
| 1430239 | UBC | −2.8056865 | Down | UBC |
|
| J, Regulatory
and cell adhesion signaling pathways |
|
| 5570132 | ACTB | −3.4210854 | Down | Actin, β |
|
| K, NRF2
pathway |
|
| 4920767 | FTL | −3.1391878 | Down | Ferritin, light
polypeptide |
|
| L,
Folate-alcohol and cancer pathway |
|
| 6510754 | ALDH1A1 | −2.9203625 | Down | Aldehyde
dehydrogenase 1 familyer A1 |
|
| M, Cell adhesion
signaling pathway |
|
| 610437 | CD24 | 2.9155455 | Up | CD24 molecule |
|
| N,
Calcium/calcium-mediated signaling pathway |
|
| 7100711 | CALM2 | 2.7985630 | Up | Calmodulin 2
(phosphorylase kinase, delta) |
|
| O, Amino acid
metabolism |
|
| 450161 | FAHD1 | 2.1592160 | Up | Fumarylacetoacetate
hydrolase domain containing 1 |
|
| P, Protein
modification pathway |
|
| Probe
ID | Symbol |
Fold-changea |
Regulation |
Definition |
|
| 4590110 | SEPT9 | 2.2080840 | Up | Septin 9 |
|
| Q, Cell
cycle |
|
| 870491 | BUB3 | 2.1889267 | Up | BUB3 budding
uninhibited by benzimidazoles 3 |
|
| R, Regulation of
actin cytoskeleton |
|
| 2760292 | PPP1CC | 2.6673288 | Up | Protein phosphatase
1, catalytic subunit |
|
| S,
Vasopressin-regulated water reabsorption |
|
| 4230520 | DNCL1 | 2.0563870 | Up | Dynein,
cytoplasmic, light polypeptide 1 |
|
| T, Transport
pathway |
|
| 1740136 | SLC38A2 | 2.1263490 | Up | Solute carrier
family 38, member 2 |
To investigate the mechanism involved in the
flavopiridol-induced antiproliferative effect on MCF7
CD44+/CD24− CSCs, pathway analysis was
performed using the WikiPathways database (www.wikipathways.org). Specifically, these pathways
are involved in the translation pathway, ribosome biogenesis,
oxidative phosphorylation, the electron transport chain pathway,
carbon metabolism, mammary gland development, protein modification
and the cell cycle (Fig. 3A and
B).
Discussion
BCSCs have been identified as subpopulations of
cells within breast tumors that possess tumor-initiating potential
in addition to the ability to self-renew and differentiate into a
diverse range of progeny cells that make up the tumor (22) These cells are resistant to traditional
therapies against cancer, including chemotherapy and radiation
therapy (5). Although treatments
associated with cancer therapy kill the majority of tumor cells,
CSCs are not killed (23). Therefore,
a more effective strategy for the treatment of breast cancer may
target CSCs. The present study investigated the effect and
underlying mechanism of flavopiridol on BCSCs with respect to
antitumor properties. The results demonstrated that flavopiridol
dose-dependently induced the growth inhibition of BCSCs.
To isolate populations of BCSCs within tumors, the
phenotypic definition of a CSC must first be established. CSCs have
been identified using cell surface markers in the majority of
cancer types. The present study isolated BCSCs based on the
CD44+/CD24− phenotype from the breast cancer
MCF7 cell line. Al-Hajj et al (9) revealed that breast cancer tumorigenic
cells exhibit a CD44+/CD24−/low phenotype.
Several studies have used the
CD44+/CD24−and/or the aldehyde dehydrogenase
(ALDH)+ phenotype for BCSC isolation (9,24).
Ginestier et al (25) isolated
stem-like cells from primary breast xenografts using
CD44+/CD24− and ALDH activity, revealing that
these cells displayed the greatest tumor-initiating capacity,
generating tumors in non-obese diabetic/severe combined
immunodeficiency mice from as little as 20 cells.
Cyclins and CDK inhibitors are involved in cell
morphogenesis, adhesion, migration, DNA repair, transcription,
cytoskeleton dynamics and cell motility. Flavopiridol is the first
CDK inhibitor that exhibits an antitumor effect against a variety
of tumor types in several solid tumors (26) The results of the present study
revealed that flavopiridol reduced the level of cell viability of
BCSCs in a dose- and time-dependent manner, and that flavopiridol
appears to possess multiple targets within tumor cells. The number
of publications involving the effect of flavopiridol on CSC is
quite limited. Soner et al (19) demonstrated that flavopiridol induced
growth inhibition and apoptosis by the upregulation of p53 and
caspases 3 and 8 in CD133+/CD44+ prostrate
CSCs.
The translation and ribosome biogenesis pathways
serve important roles in numerous cellular processes and are more
active in cancer cells compared with those in normal cells. The
inhibition of translation and ribosome biogenesis have been
reported to be associated with alterations in the cell cycle and
the regulation of cell growth (27).
The present study demonstrated that flavopiridol induced the
downregulation of translation and ribosome biogenesis genes in
CSCs. According to previous studies, flavopiridol induced
G1/S-phase cell cycle arrest (28,29). The
mechanism of flavopiridol on the cell cycle may be associated with
ribosome biogenesis. Cancer cells have been suggested to exhibit a
higher rate of ribosome biogenesis compared with that in normal
cells. Changes of proto-oncogenes and tumor-suppressor genes
activate the mechanisms that stimulate cell growth and
proliferation, and initiate certain pathways that enhance ribosome
biogenesis (30,31). Derenzini et al (32) demonstrated that the inhibition of
ribosomal RNA synthesis caused an accelerated or delayed G1/S-phase
progression in rat hepatoma cells.
The present study demonstrated the effects of
flavopiridol on BCSCs, suggesting that flavopiridol induced growth
inhibition in CD44+/CD24− BCSCs and inhibited
the translation and ribosome biogenesis pathways. Flavopiridol is
one of the most promising chemotherapy drug candidates for the
treatment of cancer. However, the data on the effects of
flavopiridol on cancer remain limited. An increased understanding
of the mechanisms responsible for the effects of the drug is
required to improve novel therapeutic strategies for breast cancer.
Combination drug therapies targeting CSCs may be an effective
method to prevent relapse and resistance in cancer therapies.
References
|
1
|
DeSantis C, Ma J, Bryan L and Jemal A:
Breast cancer statistics, 2013. CA Cancer J Clin. 64:52–62. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stingl J and Caldas C: Molecular
heterogeneity of breast carcinomas and the cancer stem cell
hypothesis. Nat Rev Cancer. 7:791–799. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weigelt B, Peterse JL and van 't Veer LJ:
Breast cancer metastasis: Markers and models. Nat Rev Cancer.
5:591–602. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu S and Wicha MS: Targeting breast
cancer stem cells. J Clin Oncol. 28:4006–4012. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Frank NY, Schatton T and Frank MH: The
therapeutic promise of the cancer stem cell concept. J Clin Invest.
120:41–50. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bhat-Nakshatri P, Goswami CP, Badve S,
Sledge GW Jr and Nakshatri H: Identification of FDA-approved drugs
targeting breast cancer stem cells along with biomarkers of
sensitivity. Sci Rep. 3:25302013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mukherjee S, Mazumdar M, Chakraborty S,
Manna A, Saha S, Khan P, Bhattacharjee P, Guha D, Adhikary A,
Mukhjerjee S and Das T: Curcumin inhibits breast cancer stem cell
migration by amplifying the E-cadherin/β-catenin negative feedback
loop. Stem Cell Res Ther. 5:1162014. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA. 100:pp.
3983–3988. 2003; View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Williams K, Motiani K, Giridhar PV and
Kasper S: CD44 integrates signaling in normal stem cell, cancer
stem cell and (pre)metastatic niches. Exp Biol Med (Maywood).
238:324–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Besson A, Dowdy SF and Roberts JM: CDK
inhibitors: Cell cycle regulators and beyond. Dev Cell. 14:159–169.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Casimiro MC, Crosariol M, Loro E, Li Z and
Pestell RG: Cyclins and cell cycle control in cancer and disease.
Genes Cancer. 3:649–657. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
O'Sullivan CC: Overcoming endocrine
resistance in hormone-receptor positive advanced breast cancer-the
emerging role of CDK4/6 inhibitors. Int J cancer Clin Res. 2:pii:
0292015. View Article : Google Scholar
|
|
14
|
Spring L, Bardia A and Modi S: Targeting
the cyclin D-cyclin-dependent kinase (CDK) 4/6-retinoblastoma
pathway with selective CDK 4/6 inhibitors in hormone
receptor-positive breast cancer: Rationale, current status, and
future directions. Discov Med. 21:65–74. 2016.PubMed/NCBI
|
|
15
|
DiPippo AJ, Patel NK and Barnett CM:
Cyclin-dependent kinase inhibitors for the treatment of breast
cancer: Past, present, and future. Pharmacotherapy. 36:652–667.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shapiro GI: Preclinical and clinical
development of the cyclin-dependent kinase inhibitor flavopiridol.
Clin Cancer Res. 10:4270s–4275s. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin TS, Ruppert AS, Johnson AJ, Fischer B,
Heerema NA, Andritsos LA, Blum KA, Flynn JM, Jones JA, Hu W, et al:
Phase II study of flavopiridol in relapsed chronic lymphocytic
leukemia demonstrating high response rates in genetically high-risk
disease. J Clin Oncol. 27:6012–6018. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Carvajal RD, Tse A, Shah MA, Lefkowitz RA,
Gonen M, Gilman-Rosen L, Kortmansky J, Kelsen DP, Schwartz GK and
O'Reilly EM: A phase II study of flavopiridol (alvocidib) in
combination with docetaxel in refractory, metastatic pancreatic
cancer. Pancreatology. 9:404–409. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Soner BC, Aktug H, Acikgoz E, Duzagac F,
Guven U, Ayla S, Cal C and Oktem G: Induced growth inhibition, cell
cycle arrest and apoptosis in CD133+/CD44+ prostate cancer stem
cells by flavopiridol. Int J Mol Med. 34:1249–1256. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Velasco-Velázquez MA, Homsi N, De La
Fuente M and Pestell RG: Breast cancer stem cells. Int J Biochem
Cell Biol. 44:573–577. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Skvortsova I, Debbage P, Kumar V and
Skvortsov S: Radiation resistance: Cancer stem cells (CSCs) and
their enigmatic pro-survival signaling. Semin Cancer Biol.
35:39–44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lawson JC, Blatch GL and Edkins AL: Cancer
stem cells in breast cancer and metastasis. Breast Cancer Res
Treat. 118:241–254. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dean M, Fojo T and Bates S: Tumour stem
cells and drug resistance. Nat Rev Cancer. 5:275–284. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Charafe-Jauffret E, Ginestier C, Iovino F,
Tarpin C, Diebel M, Esterni B, Houvenaeghel G, Extra JM, Bertucci
F, Jacquemier J, et al: Aldehyde dehydrogenase 1-positive cancer
stem cells mediate metastasis and poor clinical outcome in
inflammatory breast cancer. Clin Cancer Res. 16:45–55. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ginestier C, Hur MH, Charafe-Jauffret E,
Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG,
Liu S, et al: ALDH1 is a marker of normal and malignant human
mammary stem cells and a predictor of poor clinical outcome. Cell
Stem Cell. 1:555–567. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Senderowicz AM: Flavopiridol: The first
cyclin-dependent kinase inhibitor in human clinical trials. Invest
New Drugs. 17:313–320. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brighenti E, Treré D and Derenzini M:
Targeted cancer therapy with ribosome biogenesis inhibitors: A real
possibility? Oncotarget. 6:38617–38627. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Newcomb EW, Tamasdan C, Entzminger Y,
Alonso J, Friedlander D, Crisan D, Miller DC and Zagzag D:
Flavopiridol induces mitochondrial-mediated apoptosis in murine
glioma GL261 cells via release of cytochrome c and apoptosis
inducing factor. Cell Cycle. 2:243–250. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
van 't Veer LJ, Dai H, van de Vijver MJ,
He YD, Hart AA, Mao M, Peterse HL, Van der Kooy K, Marton MJ,
Witteveen AT, et al: Gene expression profiling predicts clinical
outcome of breast cancer. Nature. 415:530–536. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sherr CJ: The pezcoller lecture: Cancer
cell cycles revisited. Cancer Res. 60:3689–3695. 2000.PubMed/NCBI
|
|
31
|
Kusnadi EP, Hannan KM, Hicks RJ, Hannan
RD, Pearson RB and Kang J: Regulation of rDNA transcription in
response to growth factors, nutrients and energy. Gene. 556:27–34.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Derenzini M, Montanaro L, Chillà A, Tosti
E, Vici M, Barbieri S, Govoni M, Mazzini G and Treré D: Key role of
the achievement of an appropriate ribosomal RNA complement for G1-S
phase transition in H4-II-E-C3 rat hepatoma cells. J Cell Physiol.
202:483–491. 2005. View Article : Google Scholar : PubMed/NCBI
|