Introduction
Colorectal cancer (CRC) is one of the most common
types of cancer worldwide. An estimated 142,820 novel cases and
50,830 mortalities due to CRC occurred in 2013 alone (1). In spite of significant improvements in
current surgical techniques and chemoradiotherapy, the prognosis of
patients with advanced CRC remains poor, and the morbidity remains
high. Metastasis is responsible for <90% of cancer-associated
mortalities (2). Therefore, the
identification of a novel, specific and efficient molecular target
to prevent metastasis in CRC may be valuable in decreasing the
mortality rate of patients with this disease.
Forkhead box M1 (FoxM1), a member of the forkhead
family of transcription factors, is essential for cell cycle
progression and is an important cell-cycle regulator that controls
the transition from the G1 to the S phase (3–5). FoxM1
overexpression is observed in a variety of tumors and is associated
with a poor prognosis in patients with cancer (6–9).
Furthermore, the regulation of FoxM1 leads to the enhancement or
inhibition of the invasion of pancreatic cancer (10), prostate cancer (11), ovarian cancer (12) and non-small cell lung carcinoma
(NSCLC) (13) cells.
Epithelial-mesenchymal transition (EMT) is an
essential phenotypic event that occurs during embryonic
development, tissue remodeling and wound healing, and is a pivotal
event that affects cancer cell invasion (14–17). EMT
is hypothesized to be a critical mechanism of tumor metastasis
(18–20). Ueno et al (21) demonstrated that the histological
category of EMT in CRC was markedly prognostic, independent of
staging factors, and exhibited an increased prognostic
stratification power compared with the tumor and node stages.
The mechanisms underlying the FoxM1-induced
metastasis in cancer remain unclear. Kong et al (13) identified that FoxM1 may be used as a
prognostic indicator for patients with NSCLC and that FoxM1
promotes metastasis by inducing EMT in lung cancer cells. FoxM1
signaling serves an important role in tumor cell aggressiveness via
the acquisition of the EMT phenotype in gastric cancer cells
(22). The interaction between FoxM1
and EMT in CRC requires elucidation.
In the present study, the prognostic role of FoxM1
overexpression in CRC was determined, and the association between
FoxM1 expression and EMT in the metastasis of CRC was further
investigated.
Materials and methods
Patients and tissue samples
A total of 91 paraffin-embedded colon cancer
specimens were acquired from Zhejiang Provincial People's Hospital
(Hangzhou, China). The samples were collected between January 2011
and December 2013. All cases were classified according to the World
Health Organization Pathological Classification of Tumors, 2010
(23). Of the 91 individuals from
which specimens had been collected, 55 were male and 36 were
female. The patients' mean age was 54.4±10.8 years (range, 41–76
years). According to Dukes' classification, 16 patients were of
grade A, whereas 25 were of grade B, 37 were of grade C and 13 were
of grade D. None of the patients had received any radiotherapy or
chemotherapy prior to the operation. The specimens were fixed in
10% formalin at room temperature for 24 h, then embedded in
paraffin. The freshly resected tumor and adjacent normal mucosal
tissues from 30 cases were immediately stored at −70°C until use.
Paraffin-embedded blocks of colonic adenoma tissues were also
included in the present study (61 cases). The present study was
approved by the Ethics Committee of Zhejiang Provincial People's
Hospital.
Immunohistochemistry
Immunohistochemical analyses on the tissues were
performed using a Biotin-Streptavidin Horseradish Peroxidase
Detection system (OriGene Technologies, Inc., Beijing, China),
according to the manufacturer's protocol. Each 4 µm section was
deparaffinized in xylene, rehydrated in an alcohol gradient and
rinsed with PBS. Antigen retrieval was carried out in 0.01 M
citrate buffer (pH 6.0) for 3 min using the high-pressure retrieval
method. The sections were then incubated with 3%
H2O2 for 10 min, followed by 10% goat serum
(Thermo Fisher Scientific, Inc., Waltham, MA, USA) for 15 min, at
room temperature, in order to block endogenous peroxidase and
non-specific antigen binding.
Anti-FoxM1 rabbit monoclonal antibody (mAb) (cat.
no. sc-501; dilution, 1:100; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) and anti-E-cadherin mouse mAb (cat. no. UM800076;
dilution, 1:100; OriGene Technologies, Inc., Rockville, MD, USA)
were used as primary antibodies. The immunohistochemical
evaluations were independently performed by two pathologists
without knowledge of the clinical data. The immunoreactivity levels
in each case were estimated using a light microscope (BX40; Olympus
Corporation, Tokyo, Japan) by manually scoring the signal intensity
(0=negative, 1=weak, 2=moderate, 3=strong) and the proportion of
cells with positive staining (0, <5%; 1, 5–25%; 2, 26–50%; 3,
51–75%; 4, 76–100%). The intensity and proportion scores were then
multiplied to obtain a composite score: A score between 0 and 3 was
defined as negative and a score between 4 and 12 was defined as
positive.
Quantitative polymerase chain reaction
(qPCR)
Total RNA from tissues or cells was extracted using
TRIzol reagent (cat. no. 15596026; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. Complementary (c) DNA was
produced with reverse transcription using the PrimeScript First
Strand cDNA Synthesis kit (cat. no. DRR047A; Takara Biotechnology
Co., Ltd., Dalian, China) according to the manufacturer's protocol.
qPCR assays were performed using the MX3000P system (Stratagene;
Agilent Technologies, Inc., Santa Clara, CA, USA) and the SYBR
Premix ExTaq kit (cat. no. DRR042A; Takara Biotechnology Co.,
Ltd.). GAPDH was used as an internal control. The primers for qPCR
were as follows: FoxM1-Forward, GGA GCA GCG ACA GGT TAA GG;
FoxM1-Reverse, GTT GAT GGC GAA TTG TAT CAT GG; GAPDH-Forward, TGA
AGG TCG GAG TCA ACG G; GAPDH-Reverse, CTG GAA GAT GGT GAT GGG ATT.
The reaction mixture consisted of 2 µl of cDNA template in a final
reaction volume of 20 µl; there were three replicates of each
sample. Initial denaturation at 95°C for 5 min was followed by 40
cycles of amplification at 95°C for 10 sec, 58°C for 20 sec and
72°C for 20 sec. A melting curve analysis was performed following
the PCR cycles. Data was analyzed with SDS software v2.3 (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Relative expression
levels were calculated using the 2−∆ΔCt method (24).
Cell culture
The colon cancer cell lines LoVo and SW480 were
obtained from the Cell Bank of the Shanghai Chinese Academy of
Sciences Institute (Shanghai, China). The cells were cultured in
RPMI-1640 medium (cat. no. 11875-093) containing 10% fetal bovine
serum (FBS; cat. no. 10099-141; Gibco; Thermo Fisher Scientific,
Inc.), 50 units/ml penicillin and 50 µg/ml streptomycin, and were
incubated at 37°C in a 5% CO2 incubator.
Transient transfection with FoxM1
small interfering RNA (siRNA)
In order to inhibit FoxM1 expression, FoxM1 siRNA
(cat. no. sc-37615; Santa Cruz Biotechnology, Inc.) was used. LoVo
and SW480 cells (2×105 cells/well) were seeded in 6-well
plates, and cells were transfected with FoxM1 siRNA using siRNA
Transfection Reagent (cat. no. sc-29528; Santa Cruz Biotechnology,
Inc.), according to the manufacturer's protocol. Simultaneously,
cells were transfected with scrambled siRNA (cat. no. sc-37007;
Santa Cruz Biotechnology, Inc.) as a negative control. Following a
24 h transfection period, the cells were collected for later
experiments.
Western blot analysis
For western blotting, cells were washed in PBS and
lysed in a protein lysis buffer (including 1% NP-40, 20 mmol/l
Tris-HCl (pH 8), 137 mmol/l NaCl, 10% glycerol and 2 mmol/l EDTA)
by pipetting up and down to re-suspend. The lysates were gently
rocked at 2–8°C for 30 min and centrifuged at 14,000 × g for 5 min,
and the supernatant was transferred to a clean test tube. Sample
protein concentrations were quantitated using the BCA method and
then the extractions were aliquoted and stored at −80°C.
Total protein (40 µg) was used for 10% SDS-PAGE. The
proteins were electrotransferred onto polyvinylidene fluoride
membranes using the semi-dry transfer method. The membranes were
blocked with 5% BSA for 2 h at room temperature and then incubated
overnight with primary antibodies at 4°C. The primary antibodies
used were the previously described anti-FoxM1 rabbit and
anti-E-cadherin monoclonal antibodies, as well as an
anti-N-cadherin (D4R1H) XP® rabbit monoclonal antibody
(cat. no. 13116; dilution, 1:3,000; Cell Signaling Technology,
Inc., Danvers, MA, USA). Following incubation with a HRP labeled
secondary antibody (cat no. HA1001-100, 1:5,000, HuaAn
Biotechnology Co., Ltd., Hangzhou, China) for 2 h, the membranes
were treated with an electrochemiluminescence reagent (cat. no.
36208ES60, Yishen Biotech Co., Ltd., Shanghai, China) and the
chemiluminescence signal was detected using the Chemiluminescence
Gel Imaging System (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Three replicates were performed.
In vitro cell migration and invasion
assays
In the migration assays, 1×105 colon
cancer cells (LoVo or SW480) transfected with FoxM1 or negative
control siRNA were plated (in triplicate) in the upper chambers of
Transwell plates (EMD Millipore, Billerica, MA, USA) with a 8-µm
pore size membrane with 200 µl FBS-free RPMI 1640 medium. A
Matrigel-precoated membrane (cat. no. ECM554; EMD Millipore) was
used to assess cell invasion. The degree of tumor cell migration
and invasion was evaluated according to a previous protocol
(25). After 24 h or 48 h of
incubation, cells remaining on the top layers of the inserts were
removed with a cotton swab. The cells on the lower surface of the
membrane were fixed and stained with hematoxylin and eosin
staining. The cell numbers in five random fields (with
magnification, ×200) were manually counted, and the mean value was
calculated.
MTT assay
The effect of FoxM1 expression on the proliferation
of colon cancer cells was evaluated using an MTT assay. LoVo or
SW480 colon cancer cells that were transfected with FoxM1 or
negative control siRNA were plated (in triplicate) in 96-well
plates at 1×103 cells/well. The cells were incubated at
37°C in a 5% CO2 incubator for between 1 and 7 days.
Each day, 10 µl MTT (5 mg/ml) (Sigma; Merck KGaA, Darmstadt,
Germany) was added to each well and the cells were incubated for
another 4 h prior to determination of cell viability. The medium
was removed, and 150 µl dimethylsulfoxide was added to dissolve the
formazan product. The optical density at 570 nm (for measurement)
and 630 nm (for background) was measured using a microplate
reader.
Wound healing assay
siRNA-transfected LoVo or SW480 cells or negative
control siRNA-transfected cells were seeded in Matrigel-coated (50
µg/ml) 35-mm diameter culture dishes. When the cultured cells
reached 80% confluence, a sterilized pipette tip was used to mark a
line of the same width on the bottom of each dish. Dishes were
re-incubated at 37°C, and images were captured 0 or 48 h after
wounding. At 48 h after scratching, the medium was aspirated and
the cells were rinsed with PBS and fixed in 100% ethanol for 10 min
at room temperature. A wound-healing image of each group was then
photographed with a phase-contrast IX71 microscope (Olympus
Corporation).
Statistical analysis
All statistical analyses were performed using SPSS
13.0 statistical software (SPSS Inc., Chicago, IL, USA). The data
was presented as mean ± standard deviation (n=3). Fisher's exact
test and the χ2 test were used to evaluate the
association between the clinicopathological variables and the
expression of FoxM1 and E-cadherin. Spearman's rank correlation
coefficients were used to quantify the association between the
expression of FoxM1 and E-cadherin. P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression of FoxM1 is increased and
expression of E-cadherin is decreased in CRC tissues
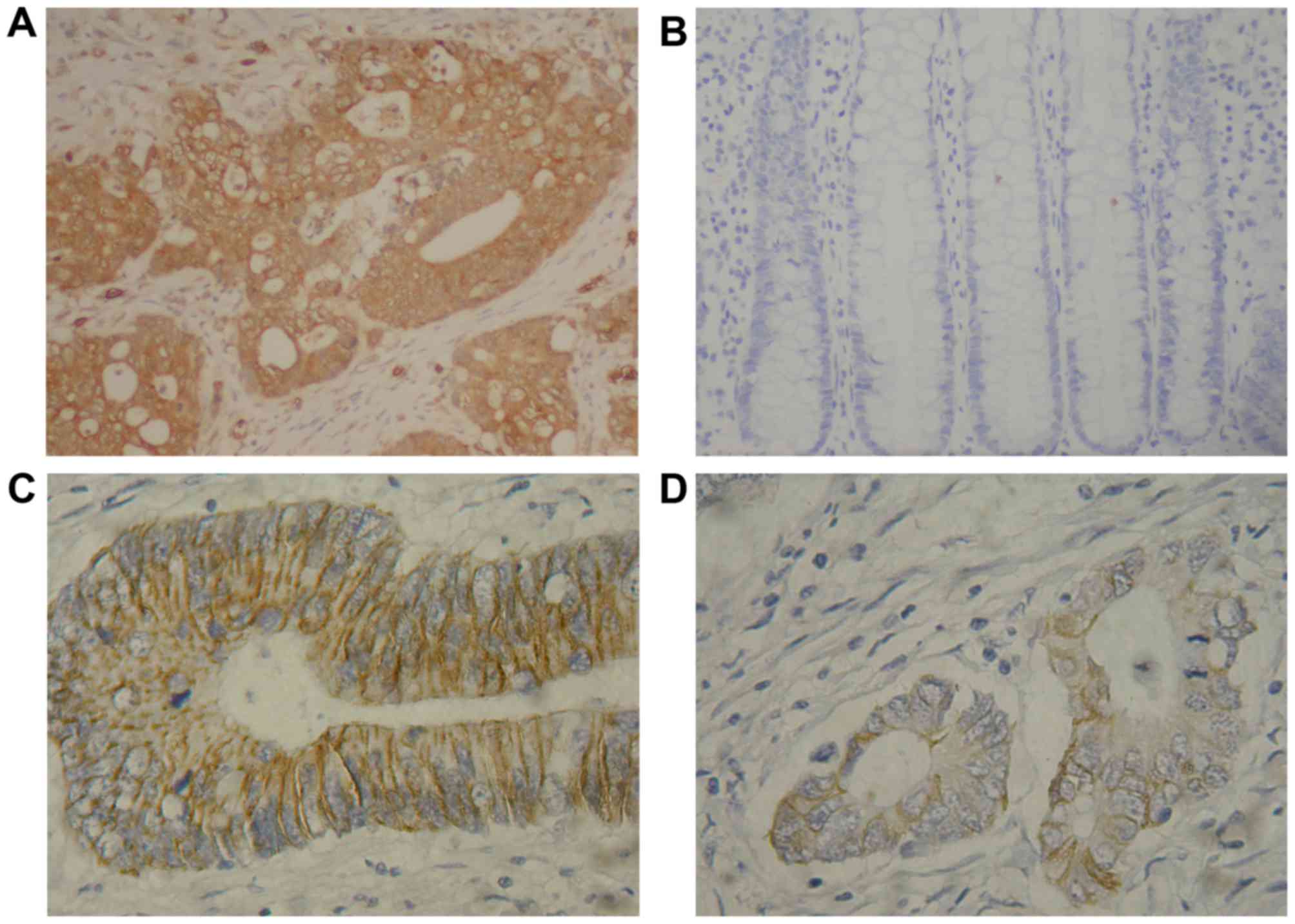
Immunostaining of FoxM1 protein was primarily
located in the cytoplasm of the tumor cells, and that of E-cadherin
protein was primarily located in the cell membrane (Fig. 1). As included in Table I, the rates of positive FoxM1 and
E-cadherin expression were 76.9% (70/91) and 34.1% (31/91) in the
CRC tissues, and 19.7% (12/61) and 82.0% (50/61) in the colorectal
adenoma tissues, respectively. The expression of FoxM1 was
increased significantly in the CRC tissues compared with the
colorectal adenoma (P<0.01) and normal colonic (P<0.01)
tissues. In contrast, E-cadherin expression was decreased
significantly in the CRC tissues compared with the colorectal
adenoma (P<0.01) and normal colonic (P<0.01) tissues. No
significant differences in the FoxM1 or E-cadherin expression
levels between the colorectal adenoma and normal colonic tissues
were identified.
 | Table I.The rate of positive FoxM1 and
E-cadherin expression in colonic tissues. |
Table I.
The rate of positive FoxM1 and
E-cadherin expression in colonic tissues.
|
|
| FoxM1 expression, n
(%) | E-cadherin
expression, n (%) |
|---|
|
|
|
|
|
|---|
| Tissue type | Total, n | Negative | Positive | Negative | Positive |
|---|
| Colorectal
cancer | 91 | 21 (23.1) | 70 (76.9) | 60 (65.9) | 31 (34.1) |
| Colorectal
adenoma | 61 | 49 (80.3) | 12 (19.7) | 11 (18.0) | 50 (82.0) |
| Normal colon | 32 | 31 (96.9) | 1 (3.1) | 4 (12.5) | 28 (87.5) |
The association of FoxM1 and E-cadherin expression
with the clinicopathological factors of the patients with cancer is
presented in Table II. The
expression levels of FoxM1 and E-cadherin were identified to be
significantly associated with advanced tumor-node-metastasis (TNM)
stage, poor tissue differentiation and lymph node metastasis.
| Table II.Association between FoxM1 and
E-cadherin expression and the clinicopathological factors of
patients with colorectal cancer. |
Table II.
Association between FoxM1 and
E-cadherin expression and the clinicopathological factors of
patients with colorectal cancer.
|
|
| FoxM1
expression | E-cadherin
expression |
|---|
|
|
|
|
|
|---|
| Variable | n | Negative | Positive | P-value | Negative | Positive | P-value |
|---|
| Sex |
|
|
| 0.19 |
|
| 0.15 |
|
Male | 55 | 12 | 43 |
| 35 | 20 |
|
|
Female | 36 | 9 | 27 |
| 25 | 11 |
|
| Age, years |
|
|
| 0.19 |
|
| 0.18 |
|
<60 | 41 | 10 | 31 |
| 27 | 14 |
|
|
≥60 | 50 | 11 | 39 |
| 33 | 17 |
|
| Tumor
localization |
|
|
| 0.13 |
|
| 0.15 |
|
Rectum | 40 | 11 | 29 |
| 25 | 15 |
|
|
Colon | 51 | 10 | 41 |
| 35 | 16 |
|
| Tumor
differentiation |
|
|
| 0.02 |
|
| 0.04a |
|
Poor | 25 | 4 | 21 |
| 20 | 5 |
|
|
Moderate | 44 | 7 | 37 |
| 30 | 14 |
|
|
Well | 22 | 10 | 12 |
| 10 | 12 |
|
| Lymph node
metastasis |
|
|
| 0.01 |
|
| <0.01 |
|
Present | 48 | 6 | 42 |
| 38 | 10 |
|
|
Absent | 43 | 15 | 28 |
| 22 | 21 |
|
| TNM stage |
|
|
| 0.01 |
|
| 0.01 |
|
I+II | 41 | 15 | 26 |
| 21 | 20 |
|
|
III+IV | 50 | 6 | 44 |
| 39 | 11 |
|
As presented in Table
III, an increase in FoxM1 expression was identified to be
associated with a decrease in E-cadherin expression (r=−0.60,
P<0.01).
| Table III.Spearman's correlation analysis
between FoxM1 and E-cadherin expression in colorectal cancer
patients. |
Table III.
Spearman's correlation analysis
between FoxM1 and E-cadherin expression in colorectal cancer
patients.
|
| E-cadherin
expression |
|
|---|
|
|
|
|
|---|
| FoxM1
expression | Negative | Positive | P-value |
|---|
| Negative | 3 | 18 |
|
| Positive | 57 | 13 | <0.01 |
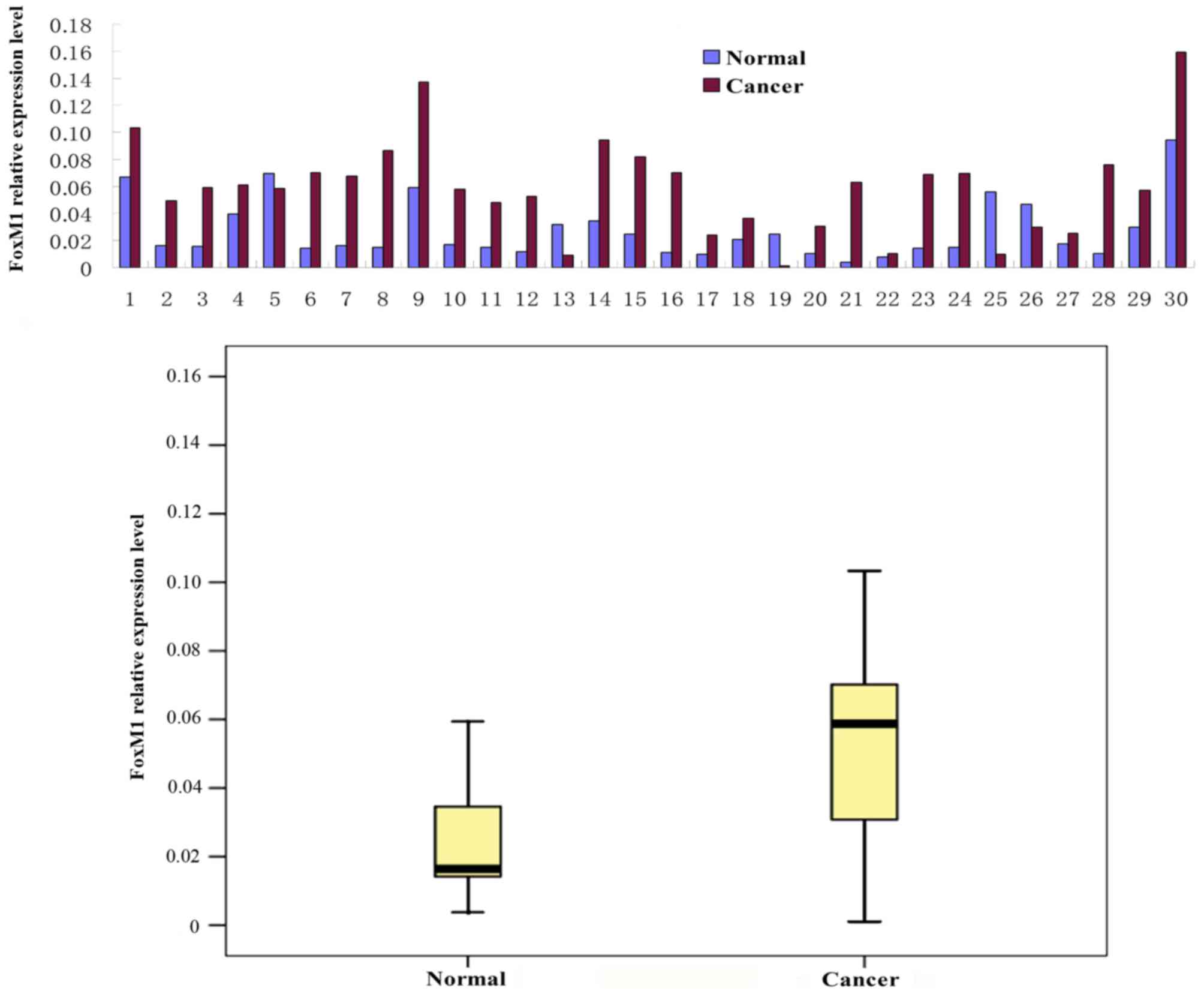
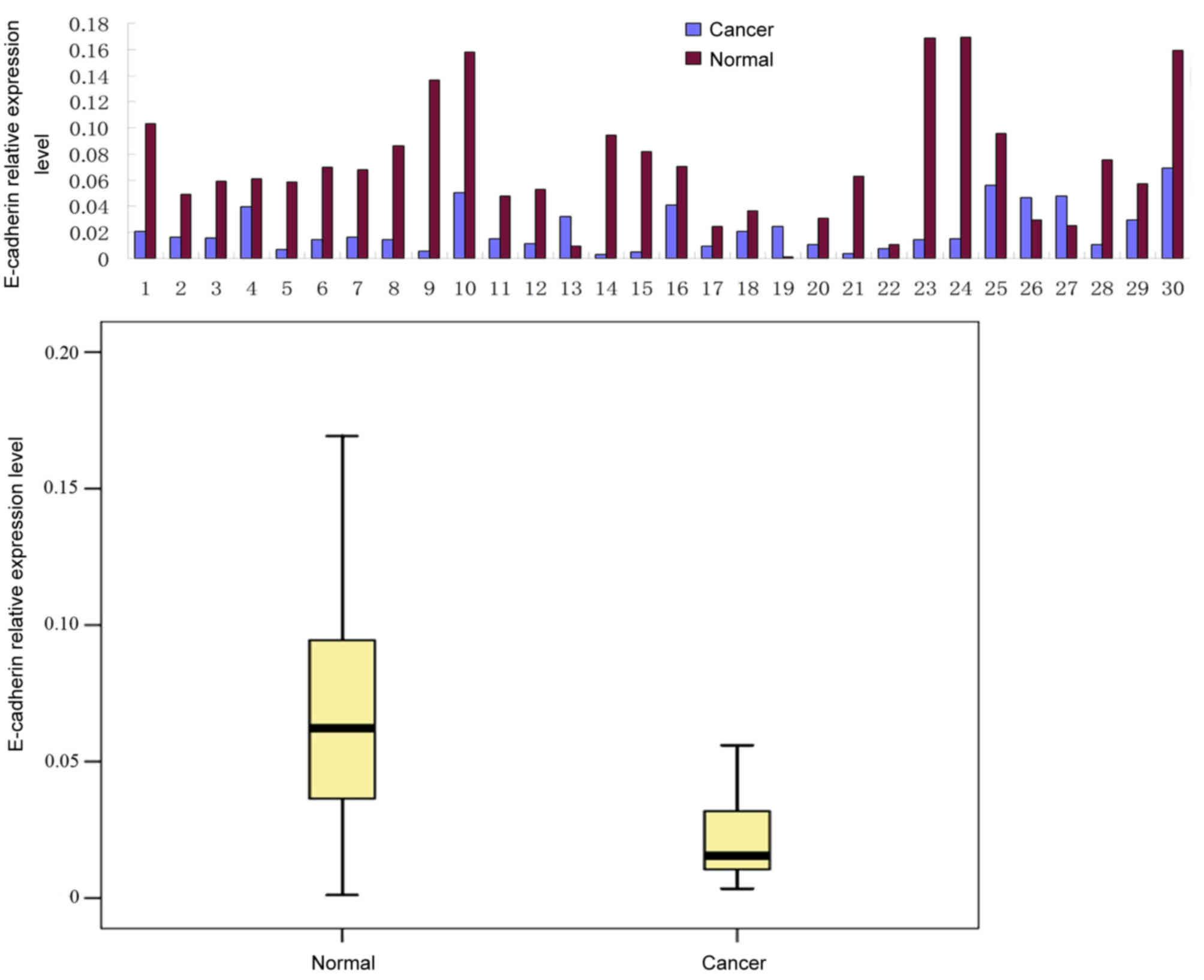
FoxM1 and E-cadherin mRNA expression levels were
determined using a qPCR assay in 30 fresh CRC tissue samples. The
relative level of FoxM1 mRNA was significantly increased (Fig. 2) and the expression of E-cadherin mRNA
was significantly decreased (Fig. 3)
in the CRC tissues compared with the levels in the adjacent
wild-type mucosal tissues (P<0.01).
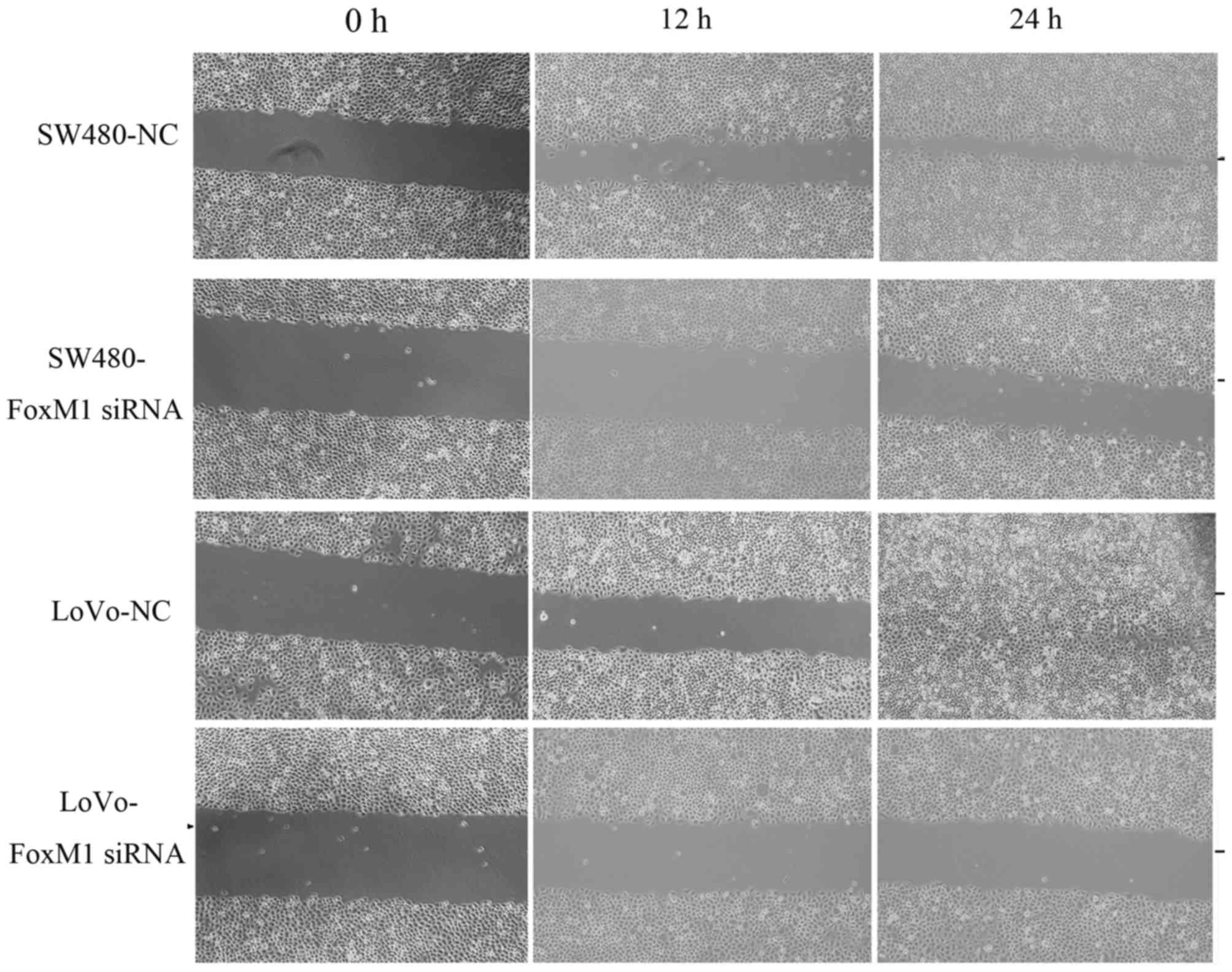
FoxM1 knockdown decreases wound
healing, in vitro migration and invasion, and proliferation of CRC
cells
Downregulation of FoxM1 using siRNA inhibited cell
migration using an in vitro wound healing assay. At 24 h
after wounding, the healing ability of FoxM1 siRNA-transfected LoVo
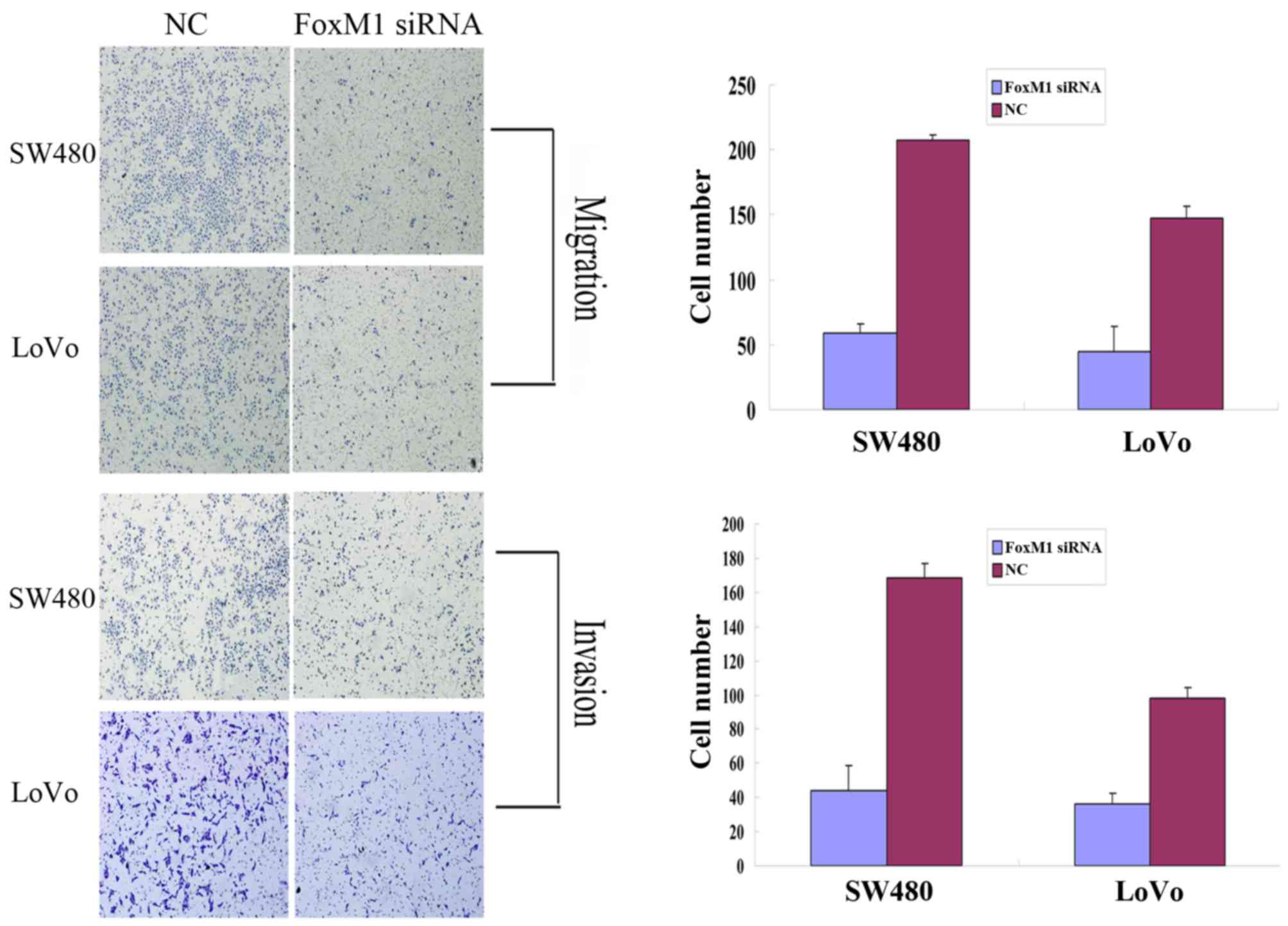
or SW480 cells was significantly suppressed (Fig. 4). Cell migration and invasion assays
were performed using a Transwell system. Compared with the control
cells, siRNA-transfected LoVo or SW480 cells exhibited a
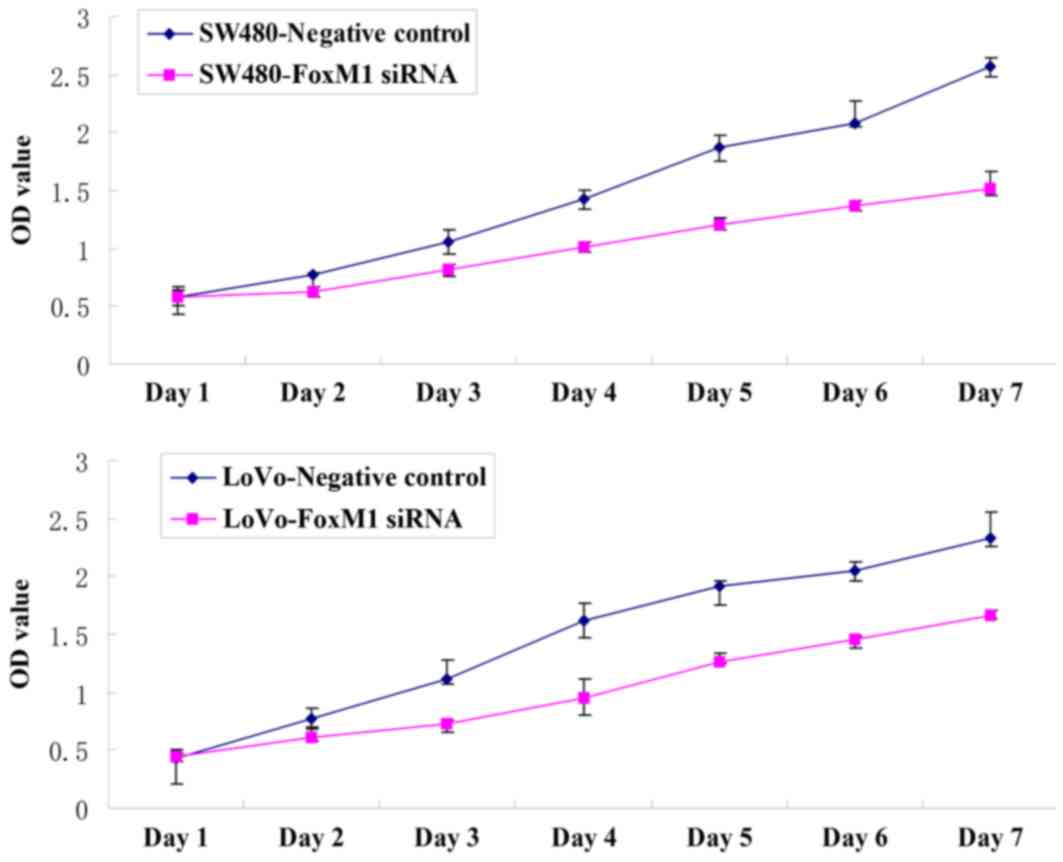
substantial decrease in migratory and invasive capacity (Fig. 5). An MTT assay demonstrated that the
proliferative capability of the FoxM1 siRNA-transfected LoVo or
SW480 cells was significantly decreased compared with the negative
control siRNA-transfected cells (Fig.
6).
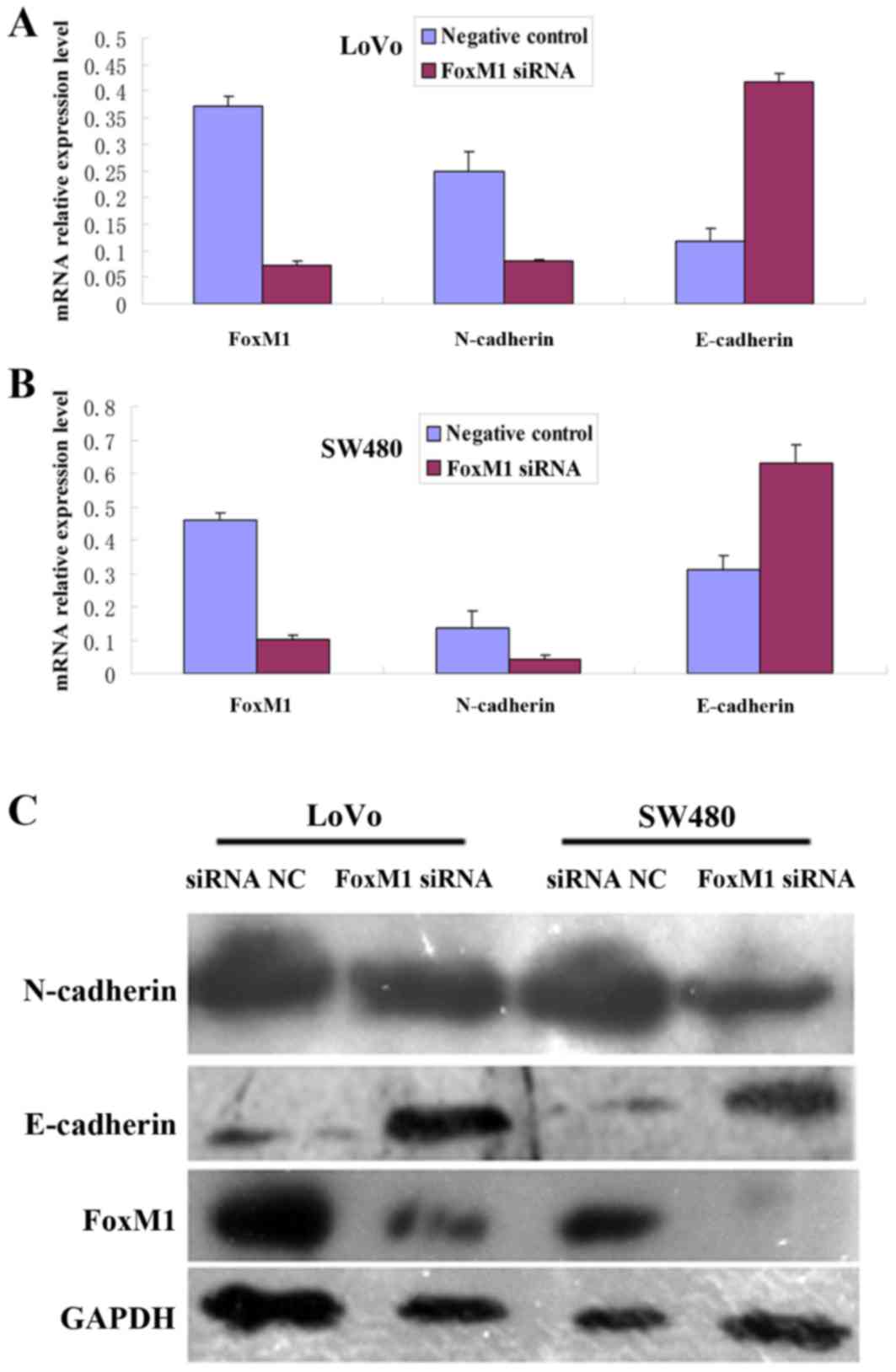
Downregulation of FoxM1 expression
alters the expression of EMT-related proteins in CRC cells
To investigate the effect of FoxM1 knockdown on the
EMT phenotype in CRC cells, FoxM1, E-cadherin and N-cadherin
protein and mRNA expression levels were determined using Western
blot and qPCR analyses. Following transfection of cells with FoxM1
siRNA, the expression levels of FoxM1 and N-cadherin protein and
mRNA were significantly decreased, whereas the expression levels of
E-cadherin protein and mRNA were increased in the LoVo or SW480
cells, compared with the negative control siRNA-transfected cells
(Fig. 7).
Discussion
FoxM1 is overexpressed in a variety of tumors,
including lung cancer, bladder cancer, clear cell renal cell
carcinoma and early stage cervical cancer (7,9,12,26).
Furthermore, FoxM1 expression is significantly associated with a
poor prognosis in many types of cancers, including gastric cancer,
pancreatic cancer and lung carcinoma (13,22,27). In
addition, the results of the present study indicated that FoxM1
expression in tumor specimens is positively associated with
advanced TNM stage, poor tissue differentiation and lymph node
metastasis. siRNA-mediated FoxM1 inhibition decreased the
proliferation and substantially decreased the migratory and
invasive capacity of CRC cells. These results suggest that FoxM1
serves an important role in the invasion and metastasis of CRC.
Recently, Yang et al (28)
reported that FoxM1 overexpression was significantly associated
with regional lymph node metastasis and tumor recurrence.
Therefore, FoxM1 has the potential to be a novel therapeutic target
in the treatment of CRC.
Metastasis involves a series of complex steps that
include decreased adhesion, increased motility, cell attachment,
matrix dissolution and migration (29). EMT is a key process in tumor
metastasis and is associated with decreased E-cadherin levels,
increased N-cadherin levels and altered expression of several
cytoskeletal proteins (30,31). The decreased expression of E-cadherin
promotes the invasion of tumor cells and is a defining
characteristic of EMT (32).
A previous meta-analysis indicated that
downregulated E-cadherin expression appeared to be associated with
worse prognoses in Asian patients with CRC (33). In the analyses of the present study,
the expression levels of E-cadherin protein and mRNA in the CRC
tissues were significantly decreased compared with those in the
adjacent wild-type mucosal tissues. Decreased E-cadherin expression
was significantly associated with advanced TNM stage, poor tissue
differentiation and lymph node metastasis in patients with CRC.
These results are consistent with previous studies; for instance,
Yun et al (34) recently
reported that the loss of E-cadherin expression in stage III CRC is
associated with a less favorable long-term prognosis in Korean
patients. However, other EMT markers (including fibronectin and
vimentin) were not identified to be associated with long-term
survival outcomes. An improved understanding of the molecular
pathways underlying EMT may reveal novel targets for the treatment
of CRC by preventing this aspect of invasion.
The present study has demonstrated that an increased
level of FoxM1 expression was associated with a decreased level of
E-cadherin expression and metastasis in patients with CRC, which
preliminarily indicates that the association between FoxM1 and
metastasis in CRC patients is associated with EMT. Previous studies
have demonstrated that increased FoxM1 expression induces the
occurrence of an EMT in pancreatic, lung and hepatocellular cancer
cells (13,35,36). In
the present study, siRNA-mediated FoxM1 inhibition decreased
N-cadherin protein and mRNA expression and increased E-cadherin
protein and mRNA expression levels in LoVo or SW480
siRNA-transfected CRC cells, which had markedly decreased migratory
and invasive capacities. These results suggest that FoxM1
expression is associated with metastasis by regulating EMT in
colorectal cells. The underlying molecular mechanisms and the
question of whether FoxM1 directly or indirectly affects the
expression of key EMT factors require further investigation.
The results of the present study suggest that FoxM1
expression is associated with metastasis in CRC. In the cell
culture experiments, RNA interference-mediated FoxM1 knockdown
significantly inhibited the proliferation, migration and invasion
of CRC cells, and induced EMT of the CRC cells. The results of the
present study raise the possibility of FoxM1 being an attractive
target for CRC therapy.
Acknowledgements
The present study was supported by the Zhejiang
Provincial Science Foundation of Medicine and Health (grant no.
2011KYA016).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Laoukili J, Kooistra MR, Brás A, Kauw J,
Kerkhoven RM, Morrison A, Clevers H and Medema RH: FoxM1 is
required for execution of the mitotic programme and chromosome
stability. Nat Cell Biol. 7:126–136. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sullivan C, Liu Y, Shen J, Curtis A,
Newman C, Hock JM and Li X: Novel interactions between FOXM1 and
CDC25A regulate the cell cycle. PLoS One. 7:e512772012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nakamura S, Hirano I, Okinaka K, Takemura
T, Yokota D, Ono T, Shigeno K, Shibata K, Fujisawa S and Ohnishi K:
The FOXM1 transcriptional factor promotes the proliferation of
leukemia cells through modulation of cell cycle progression in
acute myeloid leukemia. Carcinogenesis. 31:2012–2021. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu LL, Zhang DH, Mao X, Zhang XH and
Zhang B: Over-expression of FoxM1 is associated with adverse
prognosis and FLT3-ITD in acute myeloid leukemia. Biochem Biophys
Res Commun. 446:280–285. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu XR, Chen YH, Liu DM, Sha JJ, Xuan HQ,
Bo JJ and Huang YR: Increased expression of forkhead box M1 protein
is associated with poor prognosis in clear cell renal cell
carcinoma. Med Oncol. 30:3462013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takata A, Takiguchi S, Okada K, Takahashi
T, Kurokawa Y, Yamasaki M, Miyata H, Nakajima K, Mori M and Doki Y:
Clinicopathological and prognostic significance of FOXM1 expression
in esophageal squamous cell carcinoma. Anticancer Res.
34:2427–2432. 2014.PubMed/NCBI
|
|
9
|
Wang Y, Wen L, Zhao SH, Ai ZH, Guo JZ and
Liu WC: FoxM1 expression is significantly associated with
cisplatin-based chemotherapy resistance and poor prognosis in
advanced non-small cell lung cancer patients. Lung Cancer.
79:173–179. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang C, Xie D, Cui J, Li Q, Gao Y and Xie
K: FOXM1c promotes pancreatic cancer epithelial-to-mesenchymal
transition and metastasis via upregulation of expression of the
urokinase plasminogen activator system. Clin Cancer Res.
20:1477–1488. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cai Y, Balli D, Ustiyan V, Fulford L,
Hiller A, Misetic V, Zhang Y, Paluch AM, Waltz SE, Kasper S and
Kalin TV: Foxm1 expression in prostate epithelial cells is
essential for prostate carcinogenesis. J Biol Chem.
288:22527–22541. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wen N, Wang Y, Wen L, Zhao SH, Ai ZH, Wang
Y, Wu B, Lu HX, Yang H, Liu WC and Li Y: Overexpression of FOXM1
predicts poor prognosis and promotes cancer cell proliferation,
migration and invasion in epithelial ovarian cancer. J Transl Med.
12:1342014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kong FF, Qu ZQ, Yuan HH, Wang JY, Zhao M,
Guo YH, Shi J, Gong XD, Zhu YL, Liu F, et al: Overexpression of
FOXM1 is associated with EMT and is a predictor of poor prognosis
in non-small cell lung cancer. Oncol Rep. 31:2660–2668. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gonzalez DM and Medici D: Signaling
mechanisms of the epithelial-mesenchymal transition. Sci Signal.
7:re82014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Weber CE, Li NY, Wai PY and Kuo PC:
Epithelial-mesenchymal transition, TGF-β, and osteopontin in wound
healing and tissue remodeling after injury. J Burn Care Res.
33:311–318. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yin SY, Peng AP, Huang LT, Wang YT, Lan CW
and Yang NS: The phytochemical shikonin stimulates
epithelial-mesenchymal transition (EMT) in skin wound healing. Evid
Based Complement Alternat Med. 2013:2627962013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Grant CM and Kyprianou N: Epithelial
mesenchymal transition (EMT) in prostate growth and tumor
progression. Transl Androl Urol. 2:202–211. 2013.PubMed/NCBI
|
|
18
|
Ombrato L and Malanchi I: The EMT
universe: Space between cancer cell dissemination and metastasis
initiation. Crit Rev Oncog. 19:349–361. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kwasnicki A, Jeevan D, Braun A, Murali R
and Jhanwar-Uniyal M: Involvement of mTOR signaling pathways in
regulating growth and dissemination of metastatic brain tumors via
EMT. Anticancer Res. 35:689–696. 2015.PubMed/NCBI
|
|
20
|
Yoshida T, Ozawa Y, Kimura T, Sato Y,
Kuznetsov G, Xu S, Uesugi M, Agoulnik S, Taylor N, Funahashi Y and
Matsui J: Eribulin mesilate suppresses experimental metastasis of
breast cancer cells by reversing phenotype from
epithelial-mesenchymal transition (EMT) to mesenchymal-epithelial
transition (MET) states. Br J Cancer. 110:1497–1505. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ueno H, Shinto E, Kajiwara Y, Fukazawa S,
Shimazaki H, Yamamoto J and Hase K: Prognostic impact of
histological categorisation of epithelial-mesenchymal transition in
colorectal cancer. Br J Cancer. 111:2082–2090. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Miao L, Xiong X, Lin Y, Cheng Y, Lu J,
Zhang J and Cheng N: Down-regulation of FoxM1 leads to the
inhibition of the epithelial-mesenchymal transition in gastric
cancer cells. Cancer Genet. 207:75–82. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bosman FT, Carneiro F, Hruban RH and
Theise ND: WHO Classification of Tumours of the Digestive System.
4th. IARC Press; Lyon: 2010
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hu J and Verkman AS: Increased migration
and metastatic potential of tumor cells expressing aquaporin water
channels. FASEB J. 20:1892–1894. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu D, Zhang Z and Kong CZ: High FOXM1
expression was associated with bladder carcinogenesis. Tumour Biol.
34:1131–1138. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xia JT, Wang H, Liang LJ, Peng BG, Wu ZF,
Chen LZ, Xue L, Li Z and Li W: Overexpression of FOXM1 is
associated with poor prognosis and clinicopathologic stage of
pancreatic ductal adenocarcinoma. Pancreas. 41:629–635. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang K, Jiang L, Hu Y, Yu J, Chen H, Yao Y
and Zhu X: Short hairpin RNA-mediated gene knockdown of FOXM1
inhibits the proliferation and metastasis of human colon cancer
cells through reversal of epithelial-to-mesenchymal transformation.
J Exp Clin Cancer Res. 34:402015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Quail DF and Joyce JA: Microenvironmental
regulation of tumor progression and metastasis. Nat Med.
19:1423–1437. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Theys J, Jutten B, Habets R, Paesmans K,
Groot AJ, Lambin P, Wouters BG, Lammering G and Vooijs M:
E-Cadherin loss associated with EMT promotes radioresistance in
human tumor cells. Radiother Oncol. 99:392–397. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen L, Muñoz-Antonia T and Cress WD:
Trim28 contributes to EMT via regulation of E-cadherin and
N-cadherin in lung cancer cell lines. PLoS One. 9:e1010402014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mohamet L, Hawkins K and Ward CM: Loss of
function of e-cadherin in embryonic stem cells and the relevance to
models of tumorigenesis. J Oncol. 2011:3526162011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
He X, Chen Z, Jia M and Zhao X:
Downregulated E-cadherin expression indicates worse prognosis in
Asian patients with colorectal cancer: Evidence from meta-analysis.
PLoS One. 8:e708582013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yun JA, Kim SH, Hong HK, Yun SH, Kim HC,
Chun HK, Cho YB and Lee WY: Loss of E-Cadherin expression is
associated with a poor prognosis in stage III colorectal cancer.
Oncology. 86:318–328. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang C, Qiu Z, Wang L, Peng Z, Jia Z,
Logsdon CD, Le X, Wei D, Huang S and Xie K: A novel FoxM1-caveolin
signaling pathway promotes pancreatic cancer invasion and
metastasis. Cancer Res. 72:655–665. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Meng FD, Wei JC, Qu K, Wang ZX, Wu QF, Tai
MH, Liu HC, Zhang RY and Liu C: FoxM1 overexpression promotes
epithelial-mesenchymal transition and metastasis of hepatocellular
carcinoma. World J Gastroenterol. 21:196–213. 2015. View Article : Google Scholar : PubMed/NCBI
|