Introduction
Prostate cancer (PC) has the highest rate of
malignancy in males and is recorded as the second leading cause of
cancer-associated mortality in Europe and the USA (1). The incidence of PC has markedly
increased in the majority of developed countries, which may be due
to the prevalence of a Western lifestyle and the age distribution
of the populations (1,2). The prostate-specific antigen (PSA) blood
test is a widely used screening test for PC. However, the
prevalence of PC is markedly higher than has been traditionally
believed, and a proposed lowering of the PSA threshold for biopsies
may result in the increased over diagnosis and over treatment of PC
(3–5).
Surgery and radio therapy are effective for the treatment of
localized disease, but ~30% of treated patients with PC experience
disease relapses (6–8). Therefore, it is desirable to find a
biomarker to detect early cancer and to be able to reflect the
prognosis for patients and the therapeutic effect.
Clinically high-grade PC with Gleason scores of
between 8 and 10 exhibits rapid growth and is more likely to
exhibit spread beyond the prostate (9). The majority of patients with advanced
disease respond poorly to androgen deprivation therapy and
typically acquire a castration-resistant phenotype (6,7,10). There are limited treatment options for
these patients, including docetaxel combined with prednisone, which
are able to provide a minimal effect (11,12).
Castration-resistant PC progresses aggressively and, eventually,
patients succumb. Therefore, the development of novel therapies, on
the basis of the underlying molecular mechanisms of PC progression,
is required.
We previously analyzed gene expression profiles of
high-grade PC, using a cDNA microarray combined with laser
microbeam microdissection, to enrich populations of cancer cells
(13,14). The results of the microarray data are
available from the National Center for Biotechnology Information
(NCBI) Gene Expression Omnibus (accession no. GSE6811 and
GSE45016).
In the present study, shisa family member 2 (SHISA2)
overexpression was identified in clinical high-grade PC with
Gleason scores (GSs) between 8 and 10. In addition, SHISA2 was
positively involved in cell proliferation and the progression of
high-grade PC. The results of the present study may provide novel
insights into the underlying molecular mechanisms of PC progression
and provide insights to aid the development of novel therapeutic
drugs and a potential diagnostic biomarker for PC.
Materials and methods
Patients and tissue samples
A total of 114 frozen or paraffin-embedded tissue
samples were obtained from 114 male patients (mean age, 70 years;
age range 57–85 years) with PC undergoing open prostatectomy or
prostatic needle biopsy at Kochi Medical School Hospital (Nankoku,
Japan) with appropriate written informed consent. Patient
information was previously described by Satake et al
(14). All of the surgical specimens
were diagnosed and scored by a single pathologist using the Gleason
Grading system as described previously (13,14), and
GSs were between 6 and 10. High-grade PCs included in the surgical
specimens of PCs were characterized and identified by criteria as
previously described (14,15). All protocols were approved by the
Ethical Committee of Kochi University.
Cell lines
The LNCaP, 22Rv1, DU145 and PC-3 cell lines were
obtained from the American Type Culture Collection (Manassas, VA,
USA). LNCaP is a human prostate carcinoma epithelial cell line
derived from a left supraclavicular lymph node metastasis. 22Rv1 is
a human prostate carcinoma epithelial cell line derived from a
xenograft that was serially propagated in mice following
castration-induced regression. DU145 is a human prostate carcinoma
epithelial cell line derived from a brain metastasis. PC-3 is a
human prostate carcinoma epithelial cell line derived from a bone
metastasis. All cell lines were cultured between 3 and 10 passages,
following purchase as monolayers, in Dulbecco's modified Eagle's
medium (DMEM; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
supplemented with 10% fetal bovine serum (Thermo Fisher Scientific,
Inc.) and 1% antimycotic solution (Thermo Fisher Scientific, Inc.).
Cells were maintained in incubators at 37°C in a humidified
atmosphere containing 5% CO2.
Reverse
transcription-semi-quantitative polymerase chain reaction
(RT-sqPCR)
Purification of PC cells and normal prostatic
epithelial cells from frozen PC tissues was described previously
(13,14). Total RNA from tissues and PC cell
lines was extracted using the RNeasy kit (Qiagen, Inc., Valencia,
CA, USA) and RNase-Free DNase Set (Qiagen, Inc.), according to the
manufacturer's protocol. Subsequently, RNA was reverse transcribed
to single-stranded cDNA using an oligo d(T)12-18 primer and Super
Script Reverse Transcriptase II (Invitrogen; Thermo Fisher
Scientific, Inc.) at 42°C for 50 min. Appropriate dilutions of each
single-stranded cDNA was prepared and the cDNA content was
normalized to that of β-actin (ACTB). PCR was then performed
using single-stranded cDNA as the PCR template. Primer sequences
were as follows: SHISA2 forward, 5′-ATTGTTGGCTCCGTGTTTGT-3′
and reverse, 5′-ACTGTTGGTGTGAGGGAAGG-3′; and ACTB forward,
5′-TTGGCTTGACTCAGGATTTA-3′ and reverse, 5′-ATGCTATCACCTCCCCTGTG-3′.
PCR conditions were as follows: Initial denaturation at 98°C for 10
sec, followed by a number of cycles (21 cycles for ACTB; 28
cycles for SHISA2) of denaturation at 98°C for 10 sec,
annealing at 55°C for 5 sec and elongation at 72°C for 45 sec on a
TGradient Thermocycler (Biometra GmbH, Göttingen, Germany). PCR
products were analyzed by 2% agarose gel electrophoresis followed
by ethidium bromide staining.
Generating SHISA2 antibody
The NH2-terminal peptide (CDNDRQQGAGEPGRA) and
COOH-terminal peptide (HTNSEQKMYPAVTV) of the human SHISA2 protein
were synthesized and used to immunize rabbits. Immune sera were
purified on affinity columns packed with CNBr-activated Sepharose
4B (GE Healthcare, Chicago, IL, USA) conjugating each of the
peptide antigens.
Immunohistochemical analysis
A total of 114 frozen or paraffin-embedded tissue
samples were obtained from 114 PC patients undergoing prostatectomy
or prostatic needle biopsy at Kochi Medical School Hospital
(Nankoku, Japan) with appropriate informed consent. Frozen samples
were embedded in Tissue-Tek OTC Compound (Sakura Finetek USA, Inc.,
Torrance, CA, USA) immediately after tissue procurement and stored
at −80°C until their use. Frozen sections (4-µm thick) were cut.
For paraffin-embedded samples, the tissues were fixed in 10%
formalin neutral buffer solution for 1 day at room temperature. The
tissue specimens were embedded in paraffin wax (Merck KGaA,
Darmstadt, Germany) after fixation. Paraffin sections (2-µm thick)
were cut. All of the surgical specimens were diagnosed and scored
by a single pathologist. An immunohistochemical study was performed
using the Ventana automated immunohistochemical system (Discovery;
Ventana Medical Systems, Inc., Tucson, AZ, USA). Serial sections
were incubated with a 1:100 diluted solution of purified
anti-SHISA2 antibody (generated as described above) and a 1:100
diluted solution of anti-Wnt-5a antibody (catalog no. ab86720;
3D10; Abcam, Cambridge, UK) for 16 min at 37°C. All sections were
independently scored in a blinded manner by a clinical pathologist.
The intensity of SHISA2 staining was assessed using an
immunohistochemical score of the tumor on the basis of strength:
Negative, weak, moderate or strong staining. Comparisons between
the SHISA2 expression levels in three groups were analyzed and the
groups were formulated depending on the GS of the patient (group 1,
GS 6; group 2, GS 7; and group 3, GS between 8 and 10). Prostate
cancers with a Gleason score of 6 usually have rather good
prognoses. By contrast, those with Gleason scores of 8–10 tend to
be advanced disease that are unlikely to be cured. Additionally,
those with Gleason score 7 are intermediate in clinical
aggressiveness.
Construction of the SHISA2 expression
vector
Full-length human SHISA2 cDNA (accession no.
NM_001007538) was amplified using cDNA derived from human embryonic
293 cells and primers containing hemagglutinin (HA)-tag sequences
in the COOH-terminus, and were inserted into the pIRESneo3 vector
(Clontech, Mountain view, CA, USA). Primer sequences were as
follows: SHISA2 forward, 5′-AGGGTGGTGCCATGTGGG-3′ and
reverse, 5′-TTAAGCGTAATCTGGAACATCGTATGGGTATACAGTCACCGCTGG-3′.
Subcellular localization
PC-3 cells (1×105) per well were seeded
onto a chamber slide and cultured to 50% confluence. Cells were
transfected with the pIRESneo3-SHISA2-HA expression vector using
FuGENE6 reagent (Roche Diagnostics, Basal, Switzerland) according
to the manufacturer's protocol. Cells were fixed with 4%
paraformaldehyde for 15 min at room temperature and incubated with
PBS containing 0.1% Triton X-100 for 2 min at room temperature, and
subsequently stained with a standard protocol. Primary antibodies
used were rat anti-HA monoclonal antibody (mAb) (3F10; catalog no.
12158167001; Roche Diagnostics) and mouse anti-inositol-requiring
enzyme 1 (IRE1) mAb, as an endoplasmic reticulum (ER) marker (9F2;
catalog no. ab96481; Abcam), and all antibodies were used in a
1:250 dilution and incubated for 60 min at RT. Subsequently, cells
were incubated with the following secondary antibodies: Alexa Fluor
(AF) 568-conjugated anti-mouse immunoglobulin G (catalog no.
A11004; Thermo Fisher Scientific, Inc.) and AF 488-conjugated
anti-rat immunoglobulin G (catalog no. A11006; Thermo Fisher
Scientific, Inc.), and were used in a 1:250 dilution and incubated
for 60 min at RT. Cells were analyzed using a FV500 confocal
microscope (Olympus, Tokyo, Japan).
Western blot analysis
PC-3 cells (1×106) were seeded onto a
10-cm dish and cultured to 50% confluence. Cells were transfected
with 10 µg of the pIRESneo3 empty vector or pIRESneo3-SHISA2-HA
expression vector using FuGENE6 reagent (Roche Diagnostics),
according to the manufacturer's protocol. Cells were lysed using IP
Lysis Buffer (Thermo Fisher Scientific, Inc.) and protein
concentration was determined by the Bio-Rad Protein Assay (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Cell lysates (30 µ (30 es
and culture medium (DMEM without FBS) were separated on SDS-PAGE
(12% gels), transferred to polyvinylidene difluoride (PVDF)
membranes, and subsequently blocked with 5% milk for 30 min at RT.
Membranes were then incubated with anti-HA mAb or anti-SHISA2 pAb
for 60 min at RT. Signals were visualized using the Enhanced
Chemiluminescence Plus Western Blotting Detection system (GE
Healthcare). The anti-ACTB antibody (dilution, 1:5,000; clone
AC-15; Sigma-Aldrich; Merck KGaA) served as a loading control. ACTB
was used as a loading control (cell lysate). Silver stain was used
as a loading control (medium).
Construction of short hairpin RNA
(shRNA) expression vectors
Plasmids designed to express shRNA were prepared by
cloning double-stranded oligonucleotides into the pBAsi-hU6 Neo
vector (Takara Bio, Inc., Otsu, Japan) according to the
manufacturer's protocol. Target sequences for SHISA2 were as
follows: Sense strand sequence SHISA2-si1,
5′-CCGTGTTTGTCGCCTTTAT-3′; SHISA2-si2, 5′-CCAGATTGTGCCACATCAA-3′;
and negative control short interfering (si)RNA (siCONTROL),
5′-GCGCGCTTTGTAGGATTCG-3′, as described previously (12–14).
Cell viability assay
PC-3 cells (2×105), that highly expressed
SHISA2, were seeded onto a 6-well plate and cultured to 50%
confluence. Cells were transfected with shRNA expression vectors
(SHISA2-si1, SHISA2-si2 and siCONTROL) using FuGENE6 (Roche
Diagnostics), according to the manufacturer's protocol and cultured
in DMEM containing 800 µg/ml geneticin (Thermo Fisher Scientific,
Inc.) for 14 days. Cells were fixed with 100% methanol and stained
with 0.1% crystal violet-H2O for the colony formation
assay. Cell viability was determined using Cell Counting Kit-8
(Dojindo Molecular Technologies, Kumamoto, Japan) 10 days after
transfection according to the manufacturer's protocol. Absorbance
was determined at 450 nm as a reference with a Microplate Reader
iMark (Bio-Rad Laboratories, Inc.). These experiments were
performed in triplicate. Preliminarily, the knockdown effects of
the shRNA-expression vectors on endogenous SHISA2 expression were
validated 7 days after transfection by western blot analysis as
aforementioned.
Generation of SHISA2-overexpressing
cells and cell proliferation assay
PC-3 cells (1×106) were seeded onto a
10-cm dish and cultured to 50% confluence. Cells were transfected
with 4 µg of the pIRESneo3 empty vector or pIRESneo3-SHISA2-HA
expression vector using FuGENE6 reagent (Roche Diagnostics),
according to the manufacturer's protocol and selected for by
culturing in an DMEM containing 400 µg/ml geneticin (Thermo Fisher
Scientific, Inc.) for 16 days, and subsequently discrete colonies
were collected. Stable clones were maintained in DMEM, as
aforementioned, and assayed for exogenous SHISA2 expression by
western blot analysis as aforementioned. The proliferation of PC-3
cells that stably expressed SHISA2 (PC-3-SHISA2 clone mixture) and
those transfected with pIRESneo3 empty vector (PC-3-Mock clone
mixture) were examined using Cell-Counting Kit-8 (Dojindo Molecular
Technologies, Inc.). PC-3-SHISA2 and PC-3-Mock cells were seeded at
a concentration of 3×103 cells/well onto 48-well plates.
The assay was performed every 24 h for 5 days, according to the
manufacturer's protocol. In addition, 22Rv1-stable transformants
were generated in the same way.
cDNA microarray analysis and data acquisition. PC-3
cells (1×106) were seeded onto a 10-cm dish and cultured
to 50% confluence. Cells were transfected with pBA-siSHISA2 or
pBA-siCONTROL using FuGENE6 (Roche Diagnostics), according to the
manufacturer's protocol. At 7 days after transfection, total RNA
was extracted using the RNeasy Mini kit (Qiagen, Inc.) according to
the manufacturer's protocol. GeneChip array data were compared
using the Kurabo custom analysis service (Kurabo Industries Ltd.,
Osaka, Japan) as described previously (14). Briefly, total RNA was reverse
transcribed to cDNA using a T7 oligo d(T) primer (Affymetrix;
Thermo Fisher Scientific, Inc.). The cDNA synthesis product was
used in an in vitro transcription (IVT) reaction involving
T7 RNA polymerase (Thermo Fisher Scientific, Inc.). A
biotin-labeled ribonucleotide mix (Thermo Fisher Scientific, Inc.)
was used in the first cycle of IVT amplification. Labeled aRNA
products were fragmented, loaded onto the GeneChip Human Genome
U133 Plus 2.0 array (Thermo Fisher Scientific, Inc.) and hybridized
according to the manufacturer's protocol.
Streptavidin-phycoerythrin (Thermo Fisher Scientific, Inc.) was
used as a fluorescent conjugate to detect hybridized target
sequences. Raw intensity data from the GeneChip array were analyzed
using the GeneChip Operating Software 1.4 (Thermo Fisher
Scientific, Inc.).
Statistical analysis
Results are expressed as the mean ± standard
deviation or frequency. Data analysis was performed using the
statistical software package JMP (version 7; SAS Institute Inc.,
Cary, NC, USA). Spearman's univariate correlation analysis was
conducted between SHISA2 levels and GSs for non-parametric
variables. A comparison of three groups (siCONTROL, SHISA2-si1 and
SHISA2-si2) was analyzed statistically using One-way ANOVA and
Tukey's test. Cell proliferation assay was performed as triplicate
and Statistical significance was calculated using Student's t-test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
SHISA2 overexpression in high-grade PC
cells
Gene expression profiles of high-grade PC cells,
purified from clinical PC tissues with high GSs (between 8 and 10),
were analyzed (NCBI Gene Expression Omnibus; accession no.
GSE45016) (14). Among the genes that
were transactivated in high-grade PC cells, compared with normal
prostate epithelial cells (NP), SHISA2 was focused on in the
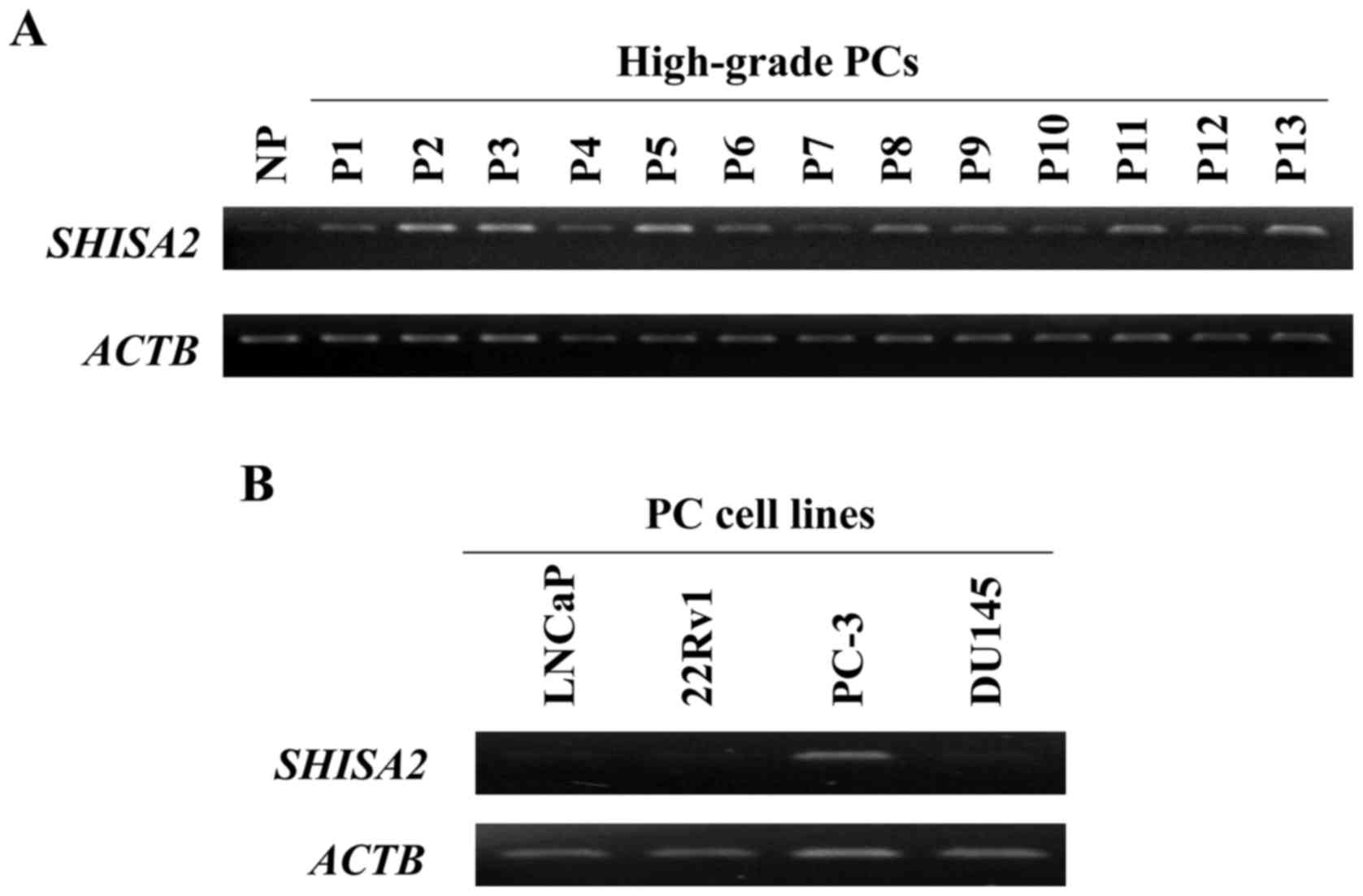
present study. RT-sqPCR validated the fact that SHISA2
expression was increased in 13 clinical high-grade PC cell samples
compared with that in NP cells (Fig.
1). Additionally, SHISA2 was expressed only in PC-3
cells (androgen-independent PC cell line) among 4 human PC cell
lines.
Expression and subcellular
localization of SHISA2 protein in PC cells
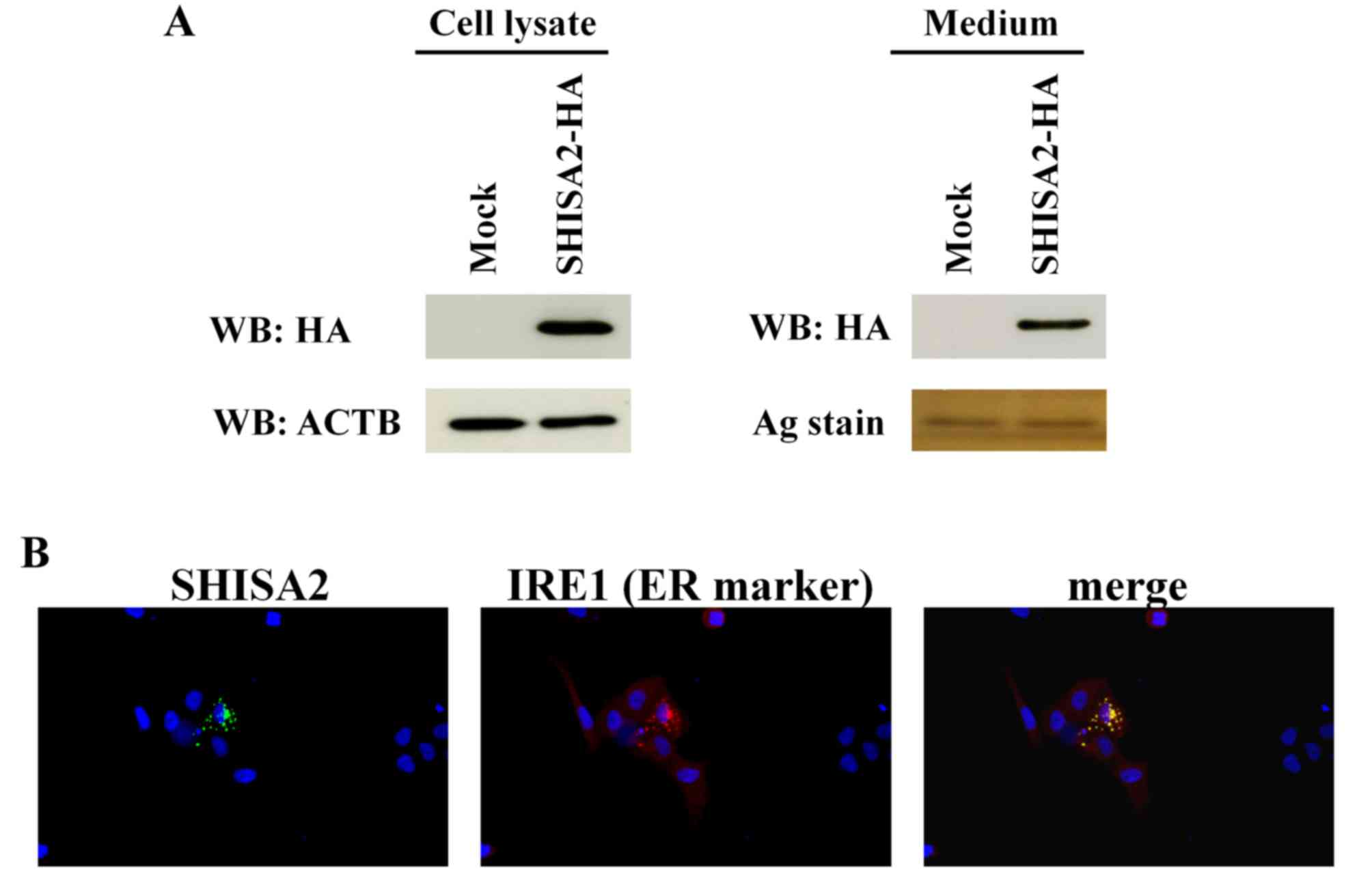
To determine the expression of SHISA2 protein in PC
cells, the pIRESneo3-SHISA2-HA expression vector was transfected
into PC-3 cells. SHISA2 protein was detected in the cell lysate and
culture medium using western blot analysis (Fig. 2A). Subcellular localization of SHISA2
protein was validated in the ER of PC-3 cells using
double-fluorescent immunostaining with anti-HA antibody and
anti-IRE1 antibody (ER marker) (Fig.
2B).
Immunohistochemical analysis of SHISA2
in clinical PC tissues
To validate the overexpression of SHISA2 protein in
clinical PC cells, immunohistochemical analysis was performed on
114 clinical PC tissues with various GSs, using a polyclonal
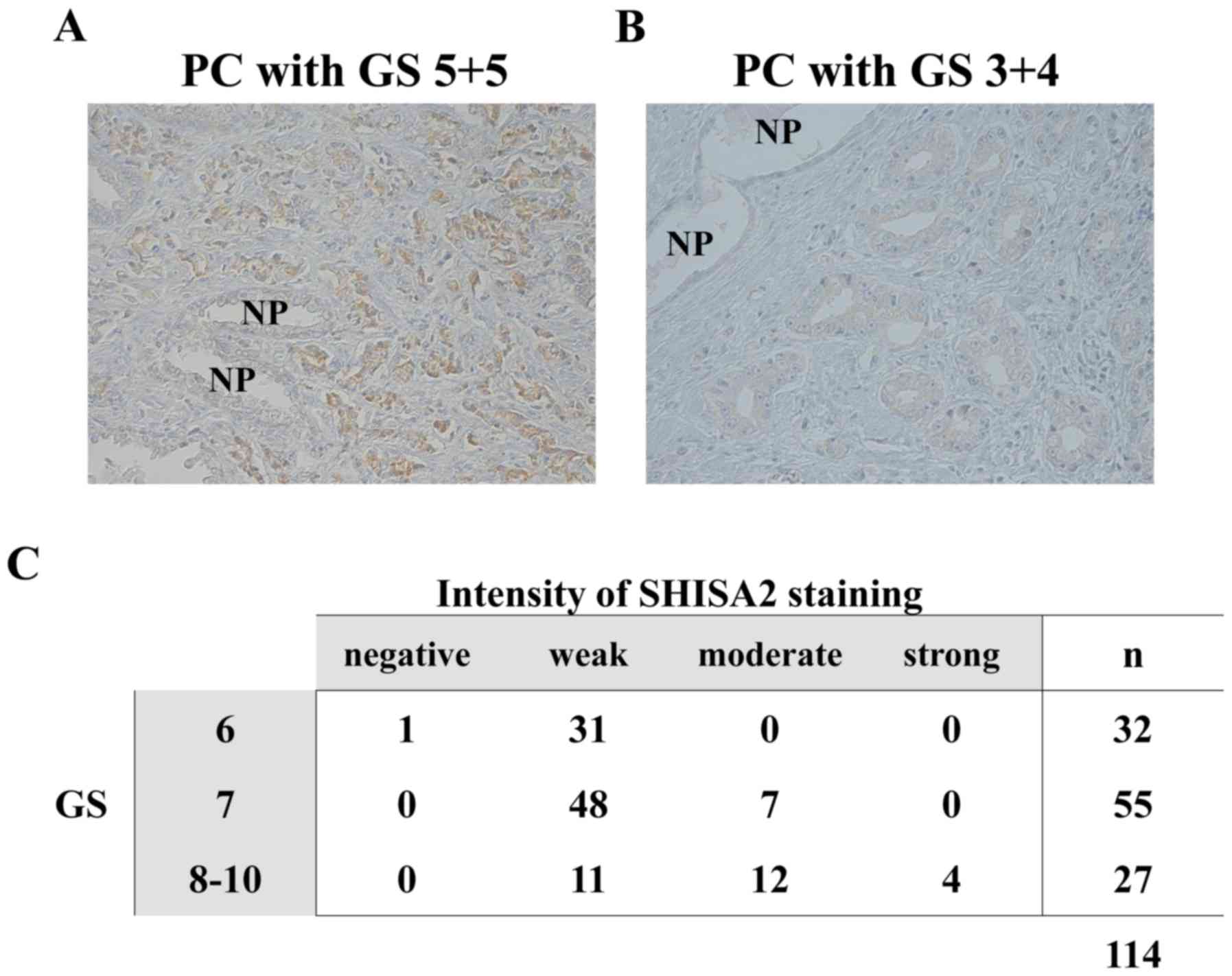
antibody specific to human SHISA2. As presented in Fig. 3A, a strong immunohistochemical signal
for SHISA2 was determined predominantly in the cytoplasm of PC
cells with GS 5+5=10, whereas PC cells with GS 3+4=7 revealed weak
immune reactivity to anti-SHISA2 antibody (Fig. 3B). Adjacent NP epithelium in the same
patient revealed no or very weak signal for SHISA2. Overall, the
results demonstrated that SHISA2 was expressed in high-grade
PCs.
To investigate the clinical and pathological
significance of SHISA2 expression in PC tissues, the association
between SHISA2 expression and GS in 114 PC tissues with various GSs
was analyzed. As presented in Fig.
3C, a significant association was observed between SHISA2
expression levels and a high GS, as determined using the univariate
Spearman's rank correlation analysis (r=0.5287; P<0.0001).
Knockdown of SHISA2 expression by
shRNA decreases PC cell numbers
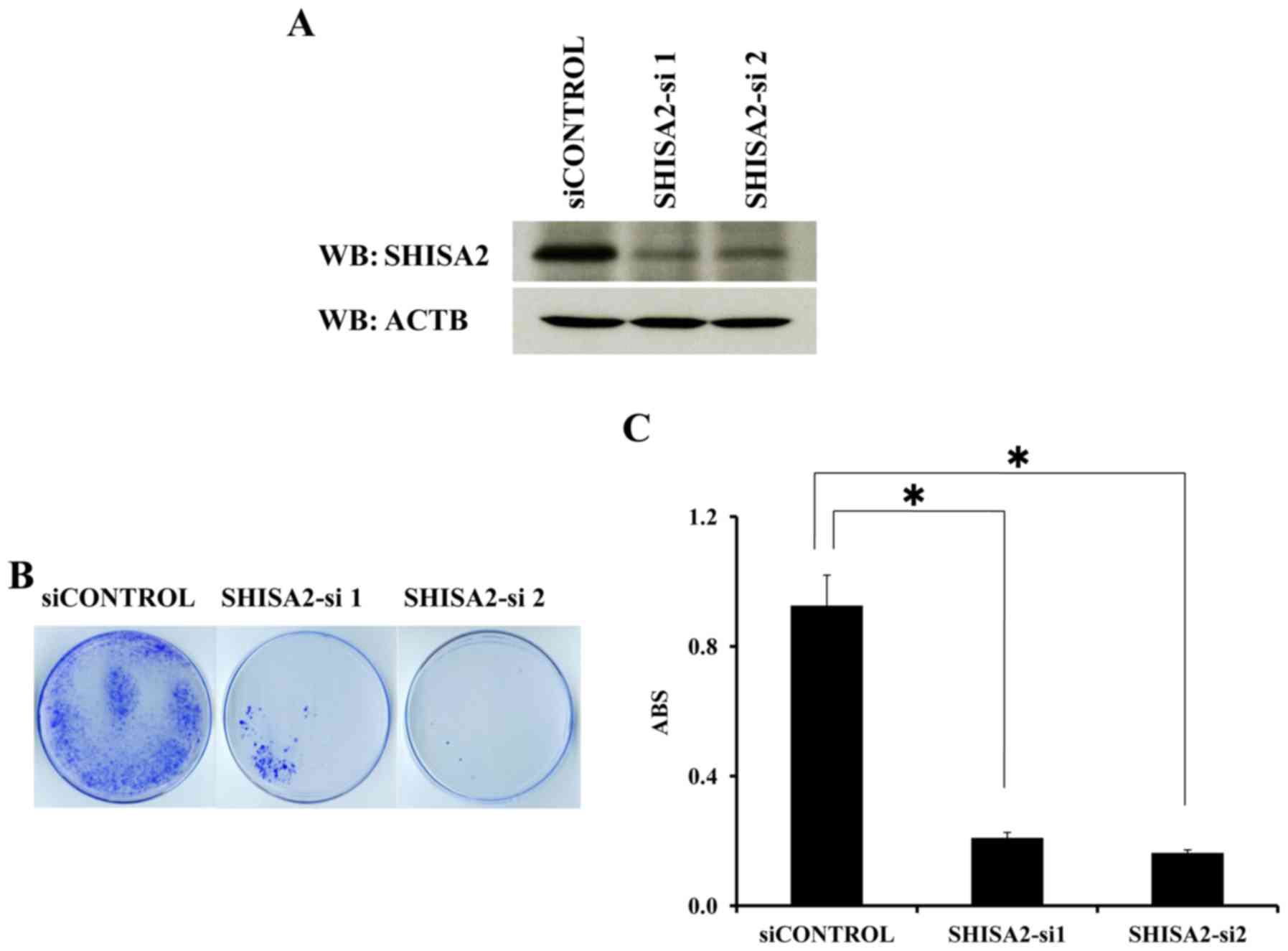
To examine the biological roles of SHISA2
overexpressed in PC cells, shRNA expression vectors (SHISA2-si1,
SHISA2-si2 and siCONTROL) were constructed and transfected into
PC-3 cells expressing endogenous SHISA2 (Fig. 1B). SHISA2-si1 and SHISA2-si2 exhibited
a marked knockdown effect on endogenous SHISA2 protein, as
determined using western blot analysis (Fig. 4A). Furthermore, transfection resulted
in a decreased number of PC colonies and viable PC cells, as
determined using a cell viability assay (P<0.01, Tukey's test).
By contrast, transfection of the negative control (siCONTROL)
exhibited a limited or no effect on SHISA2 expression and
did not affect the viability of PC-3 cells (Fig. 4B and C).
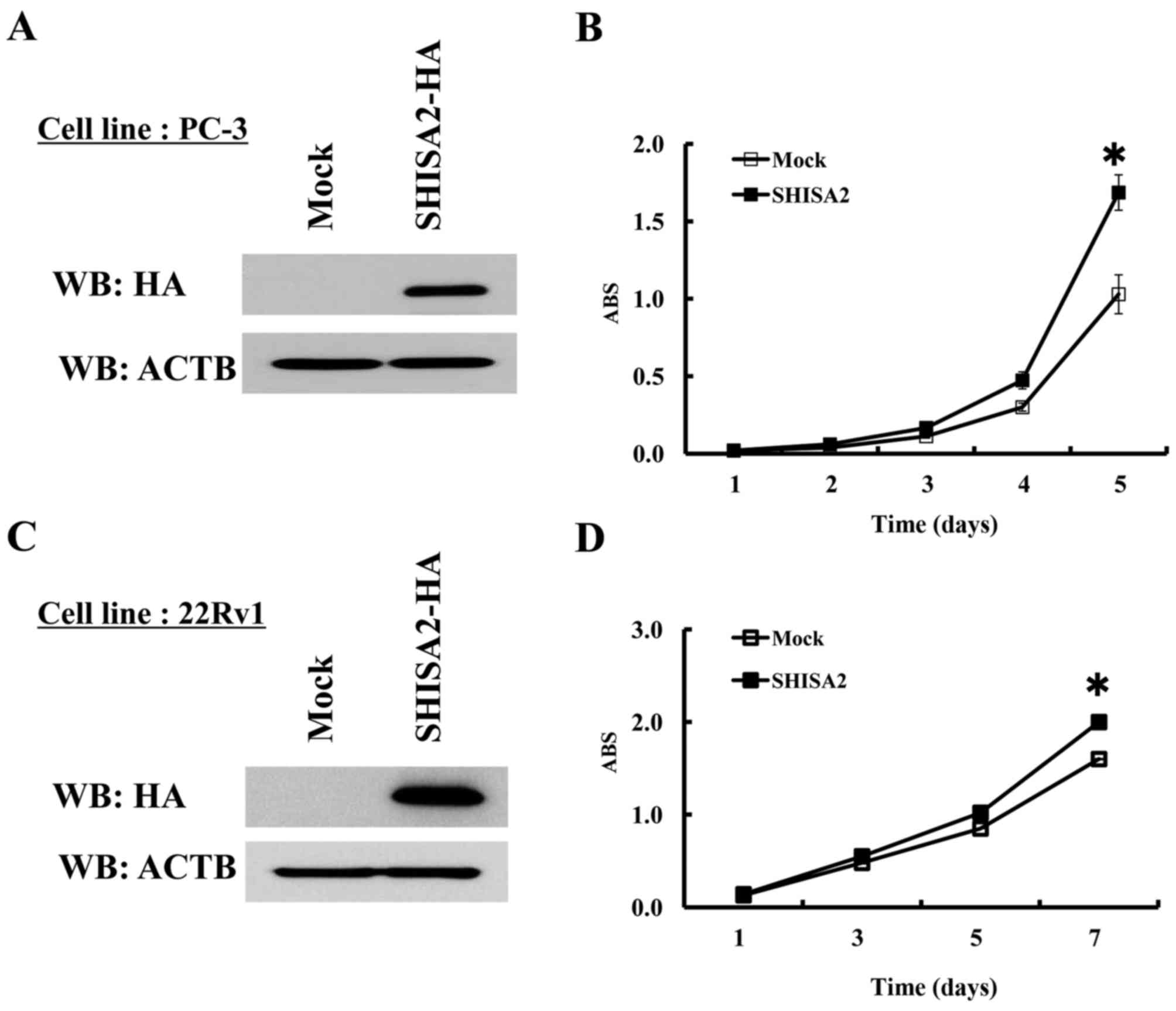
SHISA2 overexpression promotes PC cell
proliferation
To identify the potential oncogenic function of
SHISA2, a stable transformant was established from PC-3 cells,
which constitutively expressed exogenous SHISA2 (Fig. 5A). In addition, control PC-3 cells
were prepared, which were transfected with the empty vector (Mock),
and the proliferation was compared between the two. Western blot
analysis determined an increased level of exogenous SHISA2
expression in the stable clone. A cell proliferation assay revealed
that PC-3-SHISA2-overexpressing clones grew more rapidly compared
with PC-3-mock clones (P<0.01, Student's t-test), indicating
that SHISA2 overexpression promoted PC cell proliferation (Fig. 5B). Additionally, a stable transformant
was established from 22Rv1 cells, in which SHISA2 is not expressed
endogenously (Fig. 5C). As with PC-3,
a cell proliferation assay revealed that
22Rv1-SHISA2-overexpressing clones grew more rapidly compared with
22Rv1-mock clones (P<0.01, Student's t-test) (Fig. 5D).
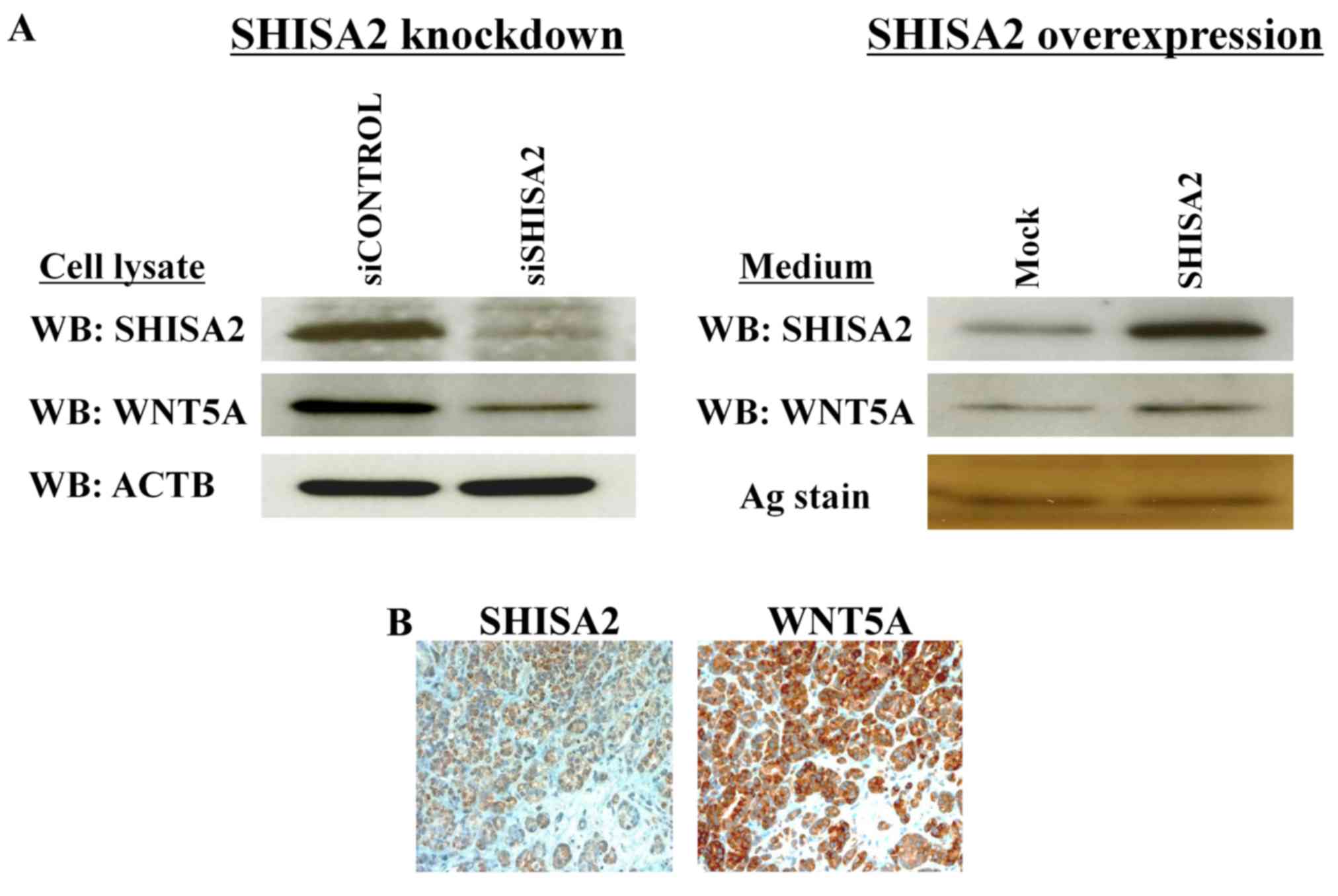
SHISA2 affects WNT5A production
The physiological and pathological functions of
SHISA in humans and human cancer types remains unknown. To
understand the association between SHISA2 overexpression and the
aggressiveness of clinical high-grade PCs, gene expression analysis
was performed using a cDNA microarray. Gene expression patterns of
PC-3 cells transfected with the two shRNA expression vectors
(siSHISA2 and siCONTROL) were compared. The microarray data are
available from the NCBI Gene Expression Omnibus (accession no.
GSE48833).
The results of the microarray analysis, with a focus
on SHISA2-knockdown in PC-3 cells, identified that WNTs and FGFs
were downregulated (Table I). Western
blot analysis identified marked downregulation of WNT5A by
SHISA2-knockdown, and upregulation of WNT5A by
SHISA2-overexpression (Fig. 6A).
Furthermore, it was validated that SHISA2 was co-expressed with
WNT5A in clinical PC tissues using immunohistochemical analysis
(Fig. 6B).
 | Table I.Downregulation of WNTs and FGFs by
SHISA2-knockdown. |
Table I.
Downregulation of WNTs and FGFs by
SHISA2-knockdown.
| Gene symbol | Gene name | Log ratio |
|---|
| WNT5A | Wingless-type MMTV
integration site family, member 5A | −3.6 |
| FGF20 | Fibroblast growth
factor 20 | −3.6 |
| WNT5A | Wingless-type MMTV
integration site family, member 5A | −3 |
| WNT6 | Wingless-type MMTV
integration site family, member 6 | −2.2 |
| FGF18 | Fibroblast growth
factor 18 | −1.8 |
| FGF18 | Fibroblast growth
factor 18 | −0.9 |
| WNT10B | Wingless-type MMTV
integration site family, member 10B | −0.8 |
| FGF18 | Fibroblast growth
factor 18 | −0.8 |
| FGF11 | Fibroblast growth
factor 11 | −0.5 |
| FGF21 | Fibroblast growth
factor 21 | −0.4 |
| FGF3 | Fibroblast growth
factor 3 | −0.2 |
Discussion
To the best of our knowledge, the present study is
the first to identify a novel molecular target and diagnostic
biomarker, SHISA2, as an overexpressed gene in high-grade PC
cells. To characterize the molecular features of clinical
high-grade PCs, gene expression profiles were analyzed using
genome-wide cDNA microarrays, combined with micro dissection, and a
number of trans-activated genes in clinical high-grade PCs were
identified (14,15). In the present study, in order to
obtain molecular cancer drug targets and diagnostic biomarkers
highly specific to PC, candidate genes were selected on the basis
of the following criteria: i) Genes that were transactivated in PC
cells relative to NP epithelial cells (to identify cancer-specific
molecular targets and biomarkers); ii) genes that encode secreted
protein (to identify serum biomarkers); and iii)
androgen-independent genes (to identify biomarkers distinct from
the androgen receptor signaling pathway, including PSA).
SHISA2 was focused on as it met the aforementioned
conditions.
Shisa was initially identified by Yamamoto et
al (16) as a novel antagonist
against Wnt and FGF signaling for head formation in Xenopus.
‘Shisa’ was termed on the basis of a type of sculpture that is
common to southern Japan (Okinawa) and exhibits a large head,
similar to the Egyptian sphinx. Shisa is a transcription
factor-type molecule that physically interacts with immature forms
of the Wnt receptor Frizzled and the FGF receptor within the ER,
and inhibits posttranslational maturation and trafficking to the
cell surface (16). In vertebrates,
the Shisa family has nine known members. Human SHISA2 is an
orthologue of the Xenopus SHISA2. The latter serves an
essential role in segmental patterning during Xenopus
somitogenesis (17). However, the
physiological and pathological functions of SHISA2 in humans
and human cancer types remains unknown.
In the present study, it was identified that
clinically high-grade PC cellsoverexpressed SHISA2.
Knockdown of SHISA2 expression by siRNA resulted in the marked
suppression of PC cell viability. By contrast, exogenous SHISA2
expression in transfected cells promoted PC cell proliferation. The
results of the present study suggested that SHISA2 was involved in
the viability and proliferation of PC cells. Furthermore, secreted
SHISA2 protein in the culture medium was detected. It was
hypothesized that, if secreted SHISA2 protein promotes the
viability and proliferation of PC cells in an autocrine or
paracrine manner, SHISA2 may be detected in the serum of patients
with PC as a useful diagnostic biomarker in predicting PC
progression. In addition, neutralization of SHISA2 by highly
specific antibodies may be used in a therapeutic strategy to treat
PC.
Notably, as a result of the cDNA microarray analysis
in the present study, the expression of WNT5A was identified to be
downregulated by SHISA2-knockdown. Previous studies have
demonstrated that WNT5A is highly upregulated in PC tissue from
patients with advanced PC, and WNT5A overexpression was associated
with the aggressiveness and poor prognosis of PC (18,19).
Additionally, it has been identified that WNT5A contributes to
gastric cancer progression by increasing metastatic potential and
that WNT5A upregulates laminin subunit γ2 to mediate cancer cell
aggressiveness (20,21). Furthermore, western blot analysis
validated a significant downregulation of WNT5A by SHISA2-knockdown
and the upregulation of WNT5A by SHISA2 overexpression.
Immunohistochemical analysis revealed the co-expression of SHISA2
and WNT5A in serial sectionsof clinical PC tissues, which suggested
that SHISA2 affects WNT5A production.
The WNT signaling pathway through which WNT5A was
regulated by SHISA2 may be essential in the cell viability and
proliferation of PC, although the underlying molecular mechanisms
remain unknown and further investigations are required. The
detection of SHISA2 and its inhibition may provide a novel target
to develop a diagnostic biomarker and/or a molecular target in the
treatment of PC.
Acknowledgements
The present study was supported by the Suzuki
Foundation for Urological Medicine, by a Grant-in-Aid for
Scientific Research (grant no. 21791508) and by the Japanese
Society for the Promotion of Science (grant no. 23791762).
References
|
1
|
Grönberg H: Prostate cancer epidemiology.
Lancet. 361:859–864. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hsing AW and Devesa SS: Trends and
patterns of prostate cancer: What do they suggest? Epidemiol Rev.
23:3–13. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Roddam AW, Hamdy FC, Allen NE and Price
CP: UK Prostate Cancer Risk Management Programme: The impact of
reducing the prostate-specific antigen threshold and including
isoform reflex tests on the performance characteristics of a
prostate-cancer detection programme. BJU Int. 100:514–517. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Müntener M, Kunz U, Eichler K, Puhan M,
Schmid DM, Sulser T and Strebel RT: Lowering the PSAthreshold for
prostate biopsy from 4 to 2.5 ng/ml: Influence on cancer
characteristics and number of men needed to biopt. Urol Int.
84:141–146. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shao YH, Albertsen PC, Roberts CB, Lin Y,
Mehta AR, Stein MN, DiPaola RS and Lu-Yao GL: Risk profiles and
treatment patterns among men diagnosed as having prostate cancer
and a prostate-specific antigen level below 4.0 ng/ml. Arch Intern
Med. 170:1256–1261. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Feldman BJ and Feldman D: The development
of androgen-independent prostate cancer. Nat Rev Cancer. 1:34–45.
2001. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Scher HI and Sawyers CL: Biology of
progressive, castration-resistant prostate cancer: Directed
therapies targeting the androgen-receptor signaling axis. J Clin
Oncol. 23:8253–8261. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Han M, Partin AW, Piantadosi S, Epstein JI
and Walsh PC: Era specific biochemical recurrence-free survival
following radical prostatectomy for clinically localized prostate
cancer. J Urol. 166:416–419. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hurwitz MD, Schultz D, Richie JP, Wein AJ,
Whittington R, Malkowicz SB and D'Amico AV: Radical prostatectomy
for high-gradeprostate cancer. Urology. 68:367–370. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Grossmann ME, Huang H and Tindall DJ:
Androgen receptor signaling in androgen refractory prostate cancer.
J Natl Cancer Inst. 93:1687–1697. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tannock IF, de Wit R, Berry WR, Horti J,
Pluzanska A, Chi KN, Oudard S, Theodore C, James ND, Turesson I, et
al: Docetaxel plus predonisone or mitoxantrone plus predonisone for
advanced prostate cancer. N Engl J Med. 351:1502–1512. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Petrylak DP, Tangen CM, Hussain MH, Lara
PN Jr, Jones JA, Taplin ME, Burch PA, Berry D, Moinpour C, Kohli M,
et al: Docetaxel and estramustine compared with mitoxantrone and
predonisone for advanced refractory prostate cancer. N Engl J Med.
351:1513–1520. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tamura K, Furihata M, Tsunoda T, Ashida S,
Takata R, Obara W, Yoshioka H, Daigo Y, Nasu Y, Kumon H, et al:
Molecular features of hormone-refractory prostate cancer cells by
genome-wide gene expression profiles. Cancer Res. 67:5117–5125.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Satake H, Tamura K, Furihata M, Anchi T,
Sakoda H, Kawada C, Iiyama T, Ashida S and Shuin T: The
ubiquitin-like molecule interferon-stimulated gene 15 is
overexpressed in human prostate cancer. Oncol Rep. 23:11–16.
2010.PubMed/NCBI
|
|
15
|
Anchi T, Tamura K, Furihata M, Satake H,
Sakoda H, Kawada C, Kamei M, Shimamoto T, Fukuhara H, Fukata S, et
al: SNRPE is involved in cell proliferation and progression of high
grade prostate cancer through the regulation of androgen receptor
expression. Oncol Lett. 3:264–268. 2012.PubMed/NCBI
|
|
16
|
Yamamoto A, Nagano T, Takehara S, Hibi M
and Aizawa S: Shisa promotes head formation through the inhibition
of receptor protein maturation for the caudalizing factors, Wnt and
FGF. Cell. 120:223–235. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nagano T, Takehara S, Takahashi M, Aizawa
S and Yamamoto A: Shisa2 promotes the maturation of somatic
precursors and transition to the segmental fate in Xenopus embryos.
Development. 133:4643–4654. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yamamoto H, Oue N, Sato A, Hasegawa Y,
Yamamoto H, Matsubara A, Yasui W and Kikuchi A: Wnt5a signaling is
involved in the aggressiveness of prostate cancer and expression of
metalloproteinase. Oncogene. 29:2036–2046. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Thiele S, Rauner M, Goettsch C, Rachner
TD, Benad P, Fuessel S, Erdmann K, Hamann C, Baretton GB, Wirth MP,
et al: Expression profile of WNT molecules in prostate cancer and
its regulation by aminobisphosphonates. J Cell Biochem.
112:1593–1600. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yamamoto H, Kitadai Y, Yamamoto H, Oue N,
Ohdan H, Yasui W and Kikuchi A: Laminin gamma2 mediates
Wnt5a-induced invasion of gastric cancer cells. Gastroenterology.
137:242–252. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hanaki H, Yamamoto H, Sakane H, Matsumoto
S, Ohdan H, Sato A and Kikuchi A: An anti-Wnt5a antibody suppresses
metastasis of gastric cancer cells in vivo by inhibiting
receptor-mediated endocytosis. Mol Cancer Ther. 11:298–307. 2012.
View Article : Google Scholar : PubMed/NCBI
|