Introduction
Colorectal cancer is one of the most prevalent
gastrointestinal cancers in the world, accounting for 9% of adult
malignancies (1,2). Unfortunately, an astounding number of
colorectal cancer patients have advanced stages diagnosed at
initial diagnosis (3). Although
surgical resection remains the most curative treatment for
colorectal cancer and improves the prognosis of patients, its
effect is still limited because patients finally develop relapse or
metastasis (4,5). Therefore, the improvement of new
diagnostic and treatment strategy for colorectal cancer is urgently
required.
In mammals, microRNAs (miRNAs) are a class of small,
non-protein-coding potential, single-stranded RNAs which suppress
the expression of target genes through directly binding to the
3′-untranslated region (UTR) of mature mRNAs (6,7). Notably,
miRNAs have been reported to be differently expressed in human
cancers and play crucial roles in cell growth, migration,
apoptosis, angiogenesis, invasion, metastasis and so forth
(8–12). miRNAs serve as oncogene or tumor
suppressor in various human cancers, involving colorectal cancer
(13–15). Specifically, miR-646 is a novel miRNAs
which was first discovered to be mammalian specific (16). Previous studies reported that miR-646
functioned as an anti-tumor miRNA which abrogated human lung cancer
or renal caner progression (17,18).
Importantly, one genomic deletion was encountered involving
miR-646/AK309218 in copy number variants (CNVs) of colorectal
cancer (19). In CpG Island
Methylator Phenotype positive (CIMP+) colorectal tumors, miR-646
was downregulated compared with other tumor markers (20). However, the role of miR-646 in
colorectal cancer is yet to be elucidated.
NOB1, as an essential protein encoding the Nin one
binding protein by two-hybrid screening, is originally discovered
in Saccharomyces cerevisiae and involved in the proteasome
as well as ribosome biogenesis (21–23). The
human NOB1 gene, localized in the nucleus, is mainly expressed in
the prostate, colorectal and lung (24–26). NOB1
plays pivotal role of modulating some physiological and
pathological functions (27).
Upregulation of the NOB1 gene is identified to elicit various human
cancers. In human ovarian cancer, downregulation of NOB1 inhibits
cell proliferation (28). Similarly,
the disregulated expression of NOB1 is identified in papillary
thyroid carcinoma and involved in tumorigenesis (29). Suppressed NOB1 blocked various human
cancer progressions, including colorectal cancer (30), non-small cell lung cancer (31), oral squamous cell carcinoma (31), laryngeal cancer (31) and so forth.
In the present study, we demonstrated that miR-646
serves as a tumor-suppressive miRNA in colorectal cancer, which
blunted colorectal cancer cell proliferation and migration.
Furthermore, miR-646 attenuated NOB1 protein level through directly
binding its 3′UTR. Notably, we further investigated that
downregulation of NOB1 also depressed colorectal cancer
progression.
Materials and methods
Clinical sample
A total of 15 colorectal cancer tissues and normal
tissues were obtained for patients who underwent surgical
resections at the first Department of General Surgery, Shidong
Hospital (Shanghai, China). The fresh tissues were frozen in liquid
nitrogen to protect the protein or RNA away from degradation. The
use of human tissues was approved by the ethics committee of
Shidong Hospital.
Cell culture
The human colorectal cancer cell lines HCT116,
SW480, SW620 and normal adult human intestine cell lines NCM460
were obtained from Cell Bank of the Chinese Academy of Sciences
(Shanghai, China). HCT116, SW480, SW620 were cultured in
serum-containing medium as recommended in a humidified 5%
CO2 atmosphere at 37°C. The normal adult human intestine
cell lines NCM460 was cultured in M3 base media as previously
described (32).
Cell transfection and vector
construction
According to manufacturer's protocol, miR-646/mock
(GenePharma, Shanghai, China) and scrambled shRNA (stem-loop-stem
structure) against NOB1 (sh-NOB1)/sh-con (Guangzhou RiboBio Co.,
Ltd., Guangzhou, China) were transiently transfected in SW480 or
SW620 cells using Lipofectamine 2000 (Invitrogen, Carlsbad, CA,
USA) at a final concentration of 50 nM. The cells were harvested at
48 h after transfection. The effects of gene silencing were
measured via western blotting and real-time PCR analysis.
The sh-NOB1 sequence was designed and synthesized
(sh-NOB1: 5′-AAGGTTAAGGTGAGCTCAT-3′). The sh-con sequence was
designed and synthesized: 5′-AATTCTCCGAACGTGTCACGT-3′. The
full-length NOB1 (NM_014062.2) cDNA containing plasmid was ordered
from IBSbio (IBS Solutions Co. Ltd, Shanghai, China). The complete
NOB1 cDNA was reverse transcribed by PCR and ligated into the
pcDNA3.1 mammalian expression vector (Invitrogen).
MTT assay
Five days after lentivirus infection, SW480 or SW620
cells were trypsinized, resuspended, seeded in a 96-well plate with
a density of 2×103 cells/well and incubated at 37°C. The
number of viable cells was measured at daily intervals (days 1–5).
At each time-point, 20 µl of 5 mg/ml MTT (Dingguo Biotechnology,
Beijing, China) was added and incubation was continued for 4 h. At
the end of the incubation period, the medium was removed carefully
and 150 µl of acidified isopropanol (in 0.01 M HCl) was added. The
plates were agitated and the absorbance was measured at 490 nm on
the spectrophotometer Biotek ELx800 (Beijing, China). Each data
point was collected from five parallel wells.
Colony formation assay
The SW480 or SW620 cells were seeded in 6-well
plates (8×102 cells/well) (in three duplicate wells) and
cultured at 37°C in 5% CO2. After two weeks, the cells
were washed with PBS once and fixed with paraformaldehyde for 30
min and washed with PBS and stained with Giemsa for 20 min. ddH2O
was used to wash the cells three times to obtain a clean
background. The number of colonies and cell number in each colony
were counted and statistically analyzed.
Wound-healing assay
SW480 or SW620 cells were cultured in 25
cm2 culture flask to 80–90% confluency, collected by
digestion and centrifugation, and then seeded into six-well plates
at 2.5×105 cells per well. miR-646 mimic/mock or
sh-NOB1/sh-con was transfected into SW480 or SW620 cells. The
floating cells were removed by washing with PBS, and the width of
scratch was observed at 0 and 48 h using inverted microscope
(50-fold). Each experiment was repeated three times.
Quantitative real-time PCR
(qRT-PCR)
Total RNA from tissues or cells were extracted using
Trizol reagent (Invitrogen) according to the manufacturer's
protocol. cDNAs were synthesized using a PrimeScript RT reagent kit
(Takara Bio, Inc., Otsu, Japan). qRT-PCR was performed with KAKA
SYBR FAST qPCR kit (Kapa Biosystems, Inc., Wilmington, MA, USA)
using a 7900HT Fast Real-Time PCR system (Applied Biosystems Japan
Ltd., Tokyo, Japan). The expression level were normalized to
endogenous small nuclear RNA U6 or GAPDH. The 2−ΔΔCt
method was used to analyze the expression level relative to the
endogenous control. Following primers were used: NOB1-Forward,
5′-ATCTGCCCTACAAGCCTAAAC-3′; NOB1-Reverse,
5′-TCCTCCTCCTCCTCCTCAC-3′; U6-Forward, 5′CTCGCTTCGGCAGCACA3′;
U6-Revers, 5′AACGCTTCACGAATTTGCGT3′; GAPDH-Forward,
5′-GAAGGTGAAGGTCGGAGTC-3′; GAPDH-Reverse,
5′-GAAGATGGTGATGGGATTTC-3′.
Protein extracting and western blot
analysis
The cells were lysed using RIPA buffer plus protease
inhibitors and phosphatase inhibitors. For western blot analysis,
25 µg of protein extracts were loaded to 12% sodium
dodecylsulfate-polyacrylamide gel electrophoresis gels and
transferred to nitrocellulose membranes. The membranes were
incubated with a primary antibody at 4° overnight, and incubated
with a secondary antibody in 1 h with room temperature. The
expression of GAPDH was used as loading control. The information of
antibodies were listed as follow: NOB1 (1:10,000; Abcam, Cambridge,
MA, USA) or GAPDH (1:5,000; Santa Cruz Biotechnology, Inc., Dallas,
TX, USA).
Luciferase reporter assay
Luciferase reporter assay using the one step
directed cloning kit (Novoprotein, Shanghai, China). The
3′untranslated regions (3′UTR) of NOB1 mRNA containing the intact
miR-646 recognition sequences were PCR-amplified and subcloned into
the Sac I and Sal I sites of pmirGLO vector. The wild-type plasmid
was created containing the 3′-UTR of NOB1 with complementary
sequence of miR-646 (NOB1 3′-UTR wt), and mutant plasmid with the
mutation sequence without complementary sequence of miR-646 (NOB1
3′-UTR mut1, mut2 and mut3). Following primers were used: (Forward)
NOB1-3′-UTR wild-F, 5′-CAAGCTTAGCGAGTTCCCGCAGGCAAAT-3′; (Reverse)
NOB1-3′-UTR wild-R, 5′-CTCTAGACATGATCTCTGGGCACAC-3′;
Statistical analysis
All data were analyzed using the Graphpad 6.0
statistical software. Results are expressed as means ± SD from at
least 3 independent experiments. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-646 is downregulated in colorectal
cancer tissues and cell lines
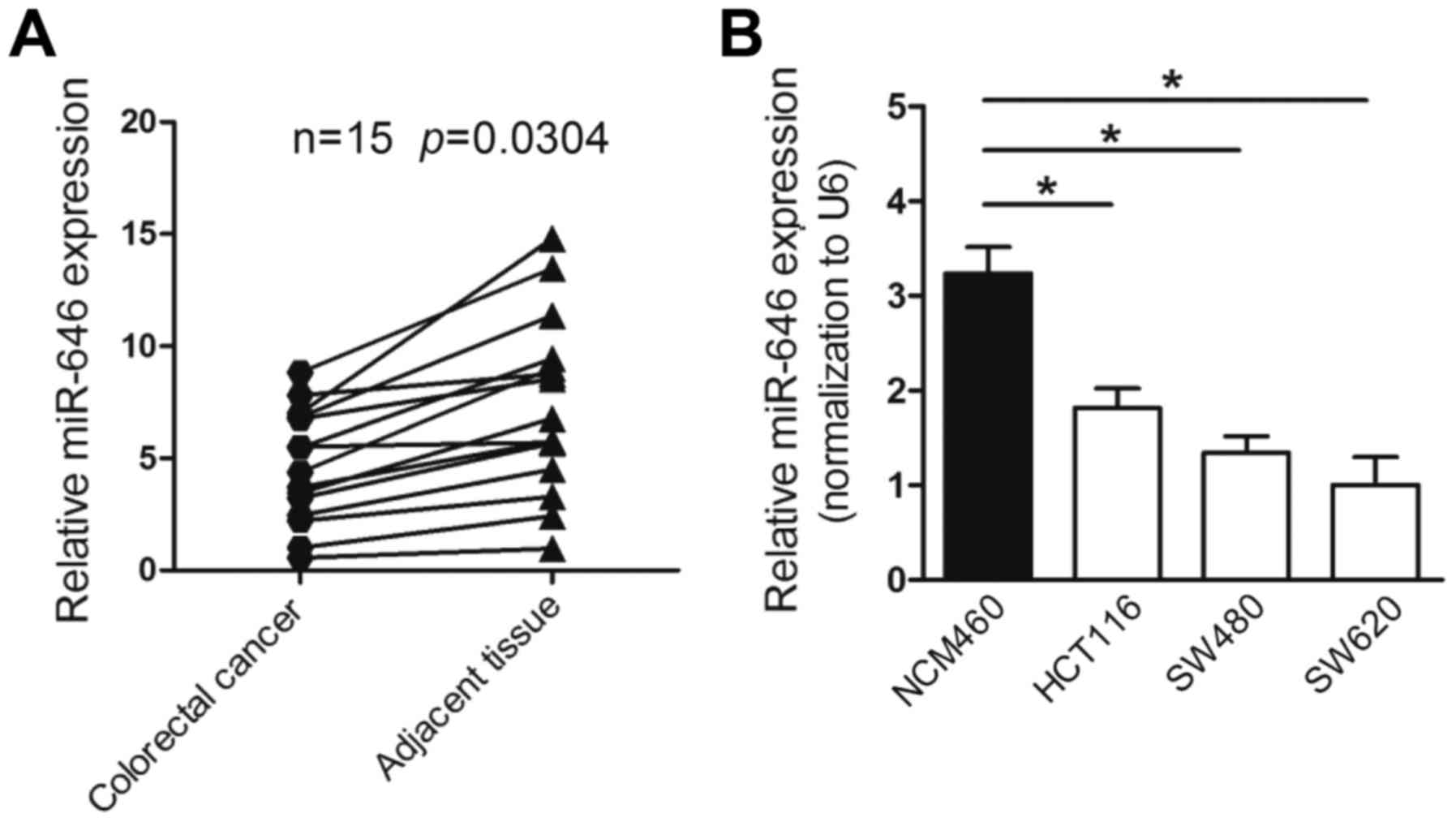
To explore the expression of miR-646 in colorectal
cancer, we first performed quantitative real-time PCR (qRT-PCR) to
detect miR-646 mRNA level in colorectal cancer tissues and
non-tumorous tissues from 15 colorectal cancer patients. The
results proved that the expression of miR-646 was remarked
downregulated in colorectal cancer compared to adjacent
non-cancerous tissues (P<0.05) (Fig.
1A). Furthermore, the mRNA level of miR-646 was lower-expressed
in various colorectal cancer cell lines, involving HCT116, SW480
and SW620 compared to normal adult human intestine cell lines
NCM460 (Fig. 1B).
Taken together, the results above indicated that the
level of miR-646 was reduced in colorectal cancer tissues and cell
lines.
miR-646 attenuates colorectal cancer
proliferation and migration in vitro
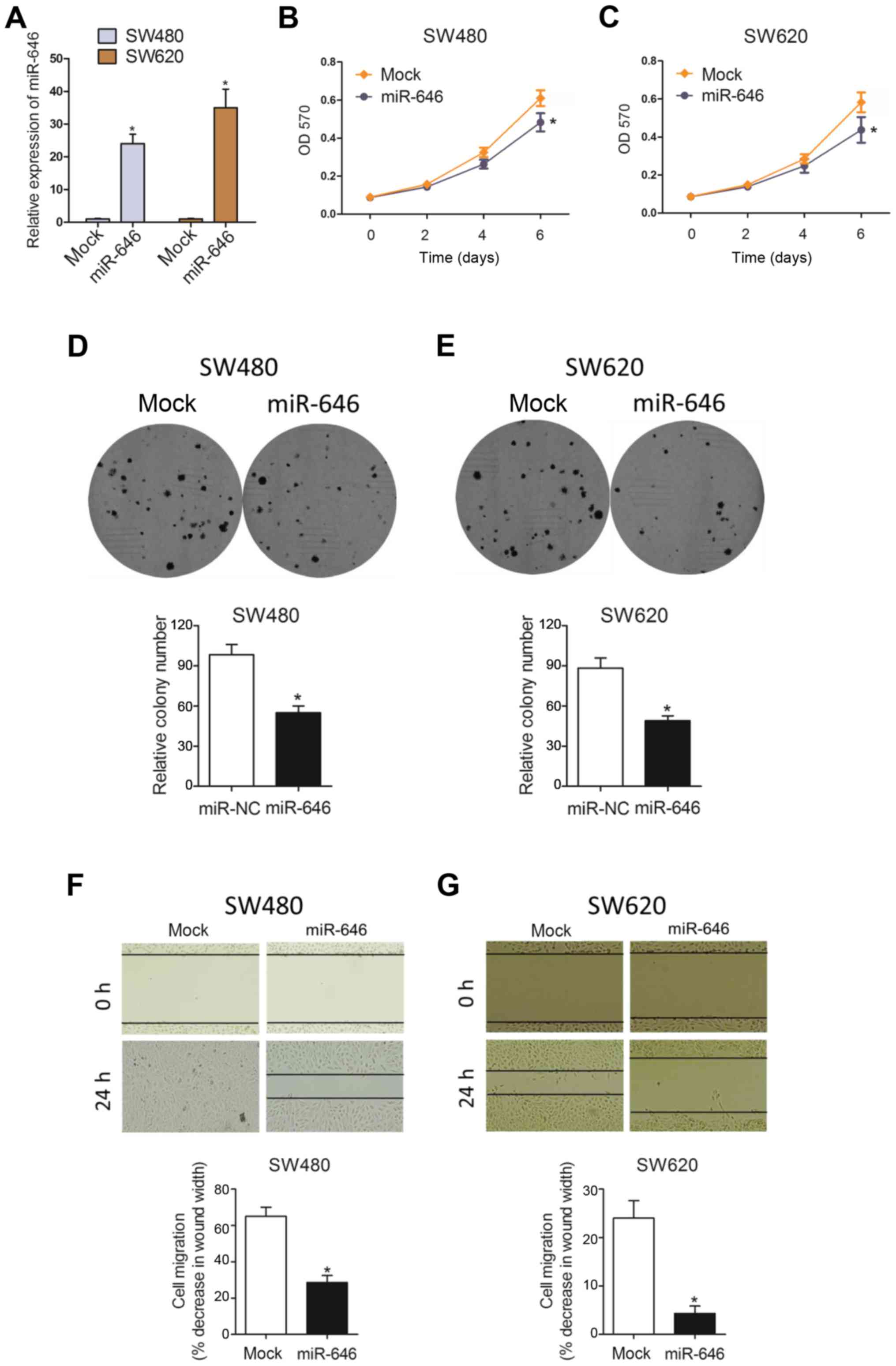
To examine the phenotypic role of miR-646 in
colorectal cancer progression, we constructed colorectal cancer
cell lines (SW480 and SW620) with miR-646 mimics to overexpress
miR-646 expression(Fig. 2A). As
Fig. 2B and C showed, MTT assay
revealed that overexpressed miR-646 (miR-646) suppressed colorectal
cancer cell proliferation in both SW480 and SW620 cell lines
compared to the control group (mock). Similarly, we also
substantiated that miR-646 attenuated cell proliferation in both
cells using colony formation assay (Fig.
2D and E). In addition, compared to mock group, miR-646 mimic
significantly abolished colorectal cancer cell migration through
wound-healing assay (Fig. 2F and
G).
Taken together, all data above suggested that
miR-646 inhibited colorectal cancer cell proliferation and
migration.
miR-646 directly targets NOB1
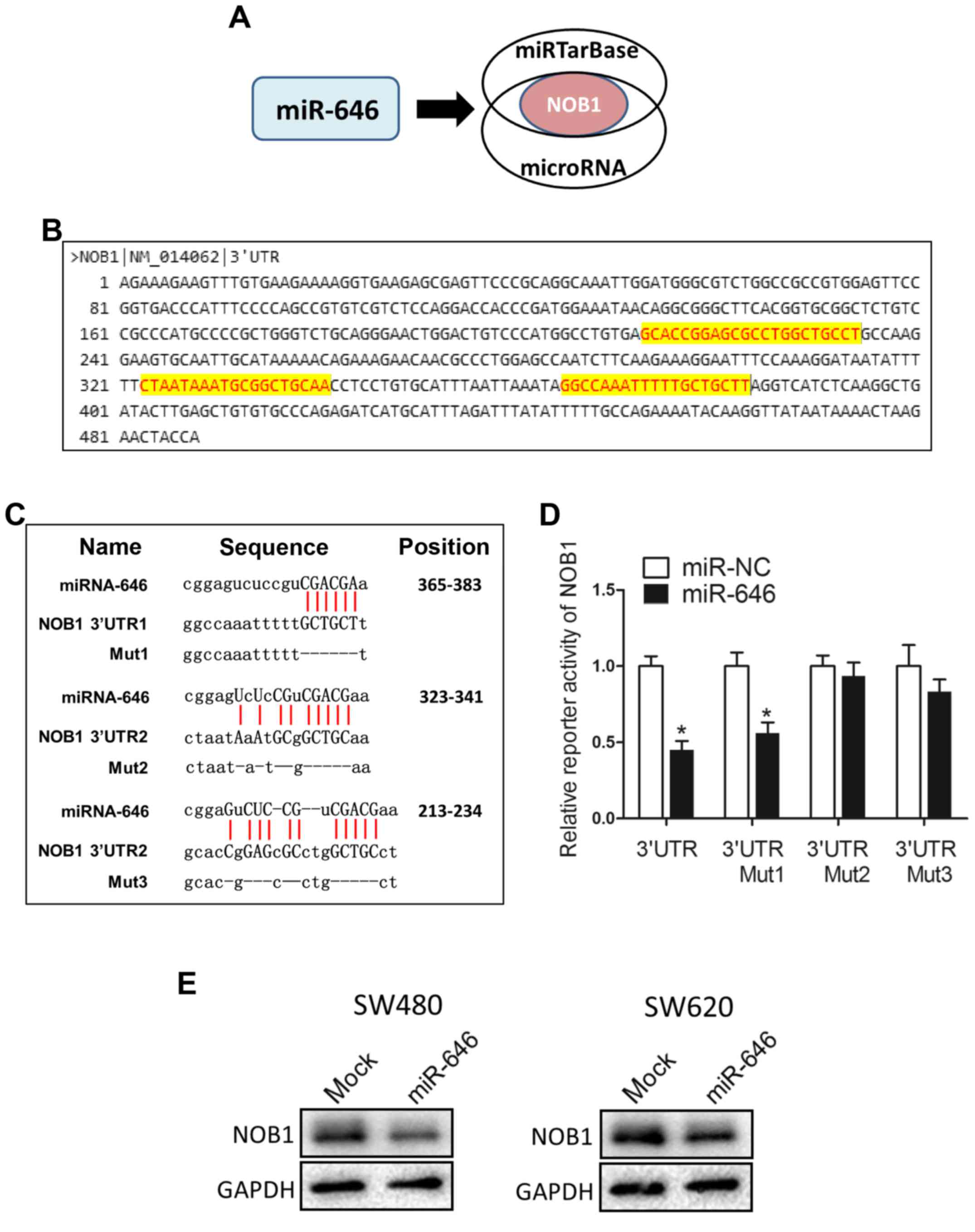
To dissect the detailed mechanism underlying
miR-646-mediated depression of colorectal cancer progression, we
used two different miRNA target-predicting algorithms (miRTarBase
and microRNA) to select the potential target genes of miR-646 and
then focused on only one candidate target genes related with cell
proliferation and migration (NOB1) (Fig.
3A). Fig. 3B showed that NOB1
included three putative miR-646 binding sites in its 3′UTR region.
We constructed luciferase reporters through cloning the 3′-UTR of
NOB1 wild-type as well as NOB1 mutant-type via the deletion
complementary sequence located at 365–383 (Mut1), 323–341 (Mut2)
and 213–234 (Mut3) (Fig. 3C).
Dual-luciferase reporter assays elucidated that the coexpression of
miR-646 in 293T cells significantly reduced the firefly luciferase
reporter activity of the NOB1 wild-type 3′-UTR but barely impacted
the mutant 3′-UTR (Fig. 3D).
Interestingly, for reporters with a single mutant 3′-UTR, only that
of the NOB1 mut1 was also downregulated by miR-646 (Fig. 3D). This result attested that wild-type
and mut2/3 3′-UTR of NOB1 were targeted by miR-646. In addition, we
performed WB to confirm that overexpressed miR-646 blunted NOB1
protein level into both colorectal cancer cell lines (Fig. 3E).
Taken together, these results suggested that miR-646
abrogated NOB1 protein and activity through directly targeting the
3′-UTR of NOB1.
Sh-NOB1 deletes cell proliferation and
migration in colorectal cancer
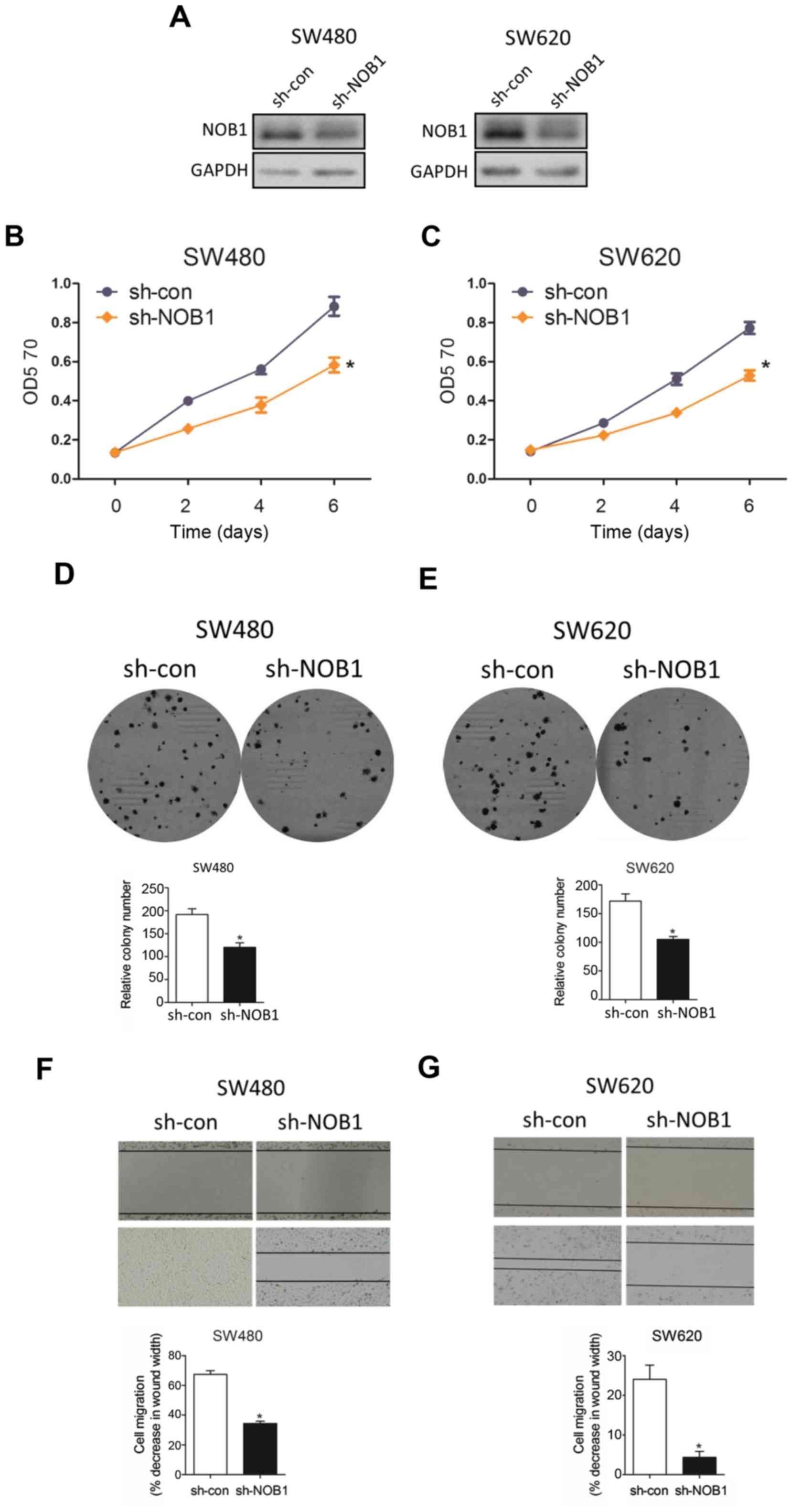
In an attempt to visualize the functional role of
NOB1 in colorectal cancer, we knocked down NOB1 expression with
sh-NOB1 and WB analysis substantiated that sh-NOB1 potently
interfered NOB1 protein in both cell lines (Fig. 4A). As expected, sh-NOB1 minimized cell
proliferation with MTT assay (Fig.
4B) and colony formation assay (Fig.
4C). In parallel, sh-NOB1 substantially depressed cell
migration capability through wound-healing assay (Fig. 4D), consistent with the phenotype
caused by overexpressed miR-646 in colorectal cancer.
Taken together, all data above suggested that
sh-NOB1 attenuated colorectal cancer cell progression.
Discussion
In the present work, we reported that miR-646
expression was downregulated in colorectal cancer tissues and cell
lines compared to that in surrounding non-cancerous tissues as well
as normal human intestine cells. Functional studies certified that
overexpressed miR-646 obviously suppressed cell growth and cell
migration via MTT, colorectaly formation and wound healing assay in
SW480 and SW620 cell lines. Importantly, miR-646 targeted
downstream gene NOB1 through directing binding a regulatory element
in its coding region. Furthermore, knocking-down NOB1 expression
was responsible for the tumor-suppressive effect of miR-646.
Collectively, all these findings supported the conclusion that bona
fide miR-646 served as a key modulator of NOB1 expression in human
colorectal cancer cells, which revealed a novel mechanism of
colorectal cancer progression.
Altered miRNAs expression has been reported to be
associated with tumor progression and overall survival in patients
with colorectal cancer. A series of miRNAs have been shown to be
anti-miRNAs in colorectal cancer. Zhang et al identified
that miR-600 abolished cell proliferation, migration and invasion
by targeting p53 in mutant p53-expressing human colorectal cancer
cell lines (33). Another study
reported that miR-219-5p attenuated the proliferation and invasion
of colorectal cancer cells by targeting calcyphosin (34). Recently, miR-145, acting as a classic
anti-miRNA, could inhibited human colorectal cancer cell migration
and invasion via PAK4-dependent pathway (35). On the contrary, an astounding number
of miRNAs have been reproted to be an oncogene in colorectal
cancer. Specifically, miR-141 facilitated colon cancer cell
proliferation by inhibiting MAP2K4 (36). Liu et al demonstrated that
miR-19a enhanced colorectal cancer proliferation and migration by
targeting TIA1 (37). Lu et al
miR-146a induced cell migration and invasion in human colorectal
cancer via carboxypeptidase M/src-FAK pathway (38). In brief, aberrant miRNAs could
function as tumor suppressors or oncogenes involving multiple
biological functions in colorectal cancer development.
A bulk of evidence indicated that miR-646 acted as a
tumor suppressor in various human cancers (17,18).
Specially, upregulated miR-646, which was a prognosis factor for
overall survival of lung cancer, not only decreased EGFR pathway,
but also reduced cell proliferation and metastasis of lung cancer
(17). On the other hand, miR-646
functioned as an anti-tumor miRNA to repress cell growth and
migration in renal cancer (18).
Consistently, our present findings also demonstrated that miR-646
was downregulated in colorectal cancer tissues and cell lines to
block tumor progression through downregulating specific oncogenic
targets-NOB1. Therefore, miR-646 could be considered as a vital
tumor suppressor in vast majority of human cancers.
Previous studies elucidated that NOB1 was found to
be upregulated in various human tumors and could be the potential
target for several miRNAs (39–41).
Specifically, NOB1, as the target gene of miR-326, was associated
with poor prognosis in glioma and gastric cancer. In cervical
cancer, NOB1 reversed the effect of miR-139-3p on inhibiting cell
metastasis while inducing cell apoptosis. In our work, we reported
that downregulation of NOB1, as a target of miR-646, attenuated
cell growth and migration of colorectal cancer.
In conclusion, our report substantiated that miR-646
acts as a suppressive non-coding RNA to mediate colorectal cancer
cell progression through targeting NOB1 expression. Our finding
provided a new therapeutic approach of colorectal cancer.
References
|
1
|
Boyle P and Langman JS: ABC of colorectal
cancer: Epidemiology. BMJ. 321:805–808. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics, 2008. CA Cancer J Clin.
58:71–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Walsh JM and Terdiman JP: Colorectal
cancer screening: Scientific review. JAMA. 289:1288–1296. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kang H, O'Connell JB, Maggard MA, Sack J
and Ko CY: A 10-year outcomes evaluation of mucinous and
signet-ring cell carcinoma of the colon and rectum. Dis Colon
Rectum. 48:1161–1168. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cunningham D, Atkin W, Lenz HJ, Lynch HT,
Minsky B, Nordlinger B and Starling N: Colorectal cancer. Lancet.
375:1030–1047. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bagga S, Bracht J, Hunter S, Massirer K,
Holtz J, Eachus R and Pasquinelli AE: Regulation by let-7 and lin-4
miRNAs results in target mRNA degradation. Cell. 122:553–563. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
White NM, Bao TT, Grigull J, Youssef YM,
Girgis A, Diamandis M, Fatoohi E, Metias M, Honey RJ, Stewart R, et
al: miRNA profiling for clear cell renal cell carcinoma: Biomarker
discovery and identification of potential controls and consequences
of miRNA dysregulation. J Urol. 186:1077–1083. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nelson KM and Weiss GJ: MicroRNAs and
cancer: Past, present, and potential future. Mol Cancer Ther.
7:3655–3660. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Osanto S, Qin Y, Buermans HP, Berkers J,
Lerut E, Goeman JJ and van Poppel H: Genome-wide microRNA
expression analysis of clear cell renal cell carcinoma by next
generation deep sequencing. PLoS One. 7:e382982012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer. 96
Suppl:R40–R44. 2007.PubMed/NCBI
|
|
13
|
Li C, Xu N, Li YQ, Wang Y and Zhu ZT:
Inhibition of SW620 human colon cancer cells by upregulating
miRNA-145. World J Gastroenterol. 22:2771–2778. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li B, Song Y, Liu TJ, Cui YB, Jiang Y, Xie
ZS and Xie SL: miRNA-22 suppresses colon cancer cell migration and
invasion by inhibiting the expression of T-cell lymphoma invasion
and metastasis 1 and matrix metalloproteinases 2 and 9. Oncol Rep.
29:1932–1938. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ahmed FE: miRNA as markers for the
diagnostic screening of colon cancer. Expert Rev Anticancer Ther.
14:463–485. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Finnerty JR, Wang WX, Hébert SS, Wilfred
BR, Mao G and Nelson PT: The miR-15/107 group of microRNA genes:
Evolutionary biology, cellular functions, and roles in human
diseases. J Mol Biol. 402:491–509. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pan Y, Chen Y, Ma D, Ji Z, Cao F, Chen Z,
Ning Y and Bai C: miR-646 is a key negative regulator of EGFR
pathway in lung cancer. Exp Lung Res. 42:286–295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li W, Liu M, Feng Y, Xu YF, Huang YF, Che
JP, Wang GC, Yao XD and Zheng JH: Downregulated miR-646 in clear
cell renal carcinoma correlated with tumour metastasis by targeting
the nin one binding protein (NOB1). Br J Cancer. 111:1188–1200.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Venkatachalam R, Verwiel ET, Kamping EJ,
Hoenselaar E, Görgens H, Schackert HK, van Krieken JH, Ligtenberg
MJ, Hoogerbrugge N, van Kessel AG and Kuiper RP: Identification of
candidate predisposing copy number variants in familial and
early-onset colorectal cancer patients. Int J Cancer.
129:1635–1642. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Slattery ML, Wolff E, Hoffman MD, Pellatt
DF, Milash B and Wolff RK: MicroRNAs and colon and rectal cancer:
Differential expression by tumor location and subtype. Genes
Chromosomes Cancer. 50:196–206. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tone Y, Tanahashi N, Tanaka K, Fujimuro M,
Yokosawa H and Toh-e A: Nob1p, a new essential protein, associates
with the 26S proteasome of growing saccharomyces cerevisiae cells.
Gene. 243:37–45. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wild T, Horvath P, Wyler E, Widmann B,
Badertscher L, Zemp I, Kozak K, Csucs G, Lund E and Kutay U: A
protein inventory of human ribosome biogenesis reveals an essential
function of exportin 5 in 60S subunit export. PLoS Biol.
8:e10005222010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wyler E, Zimmermann M, Widmann B, Gstaiger
M, Pfannstiel J, Kutay U and Zemp I: Tandem affinity purification
combined with inducible shRNA expression as a tool to study the
maturation of macromolecular assemblies. RNA. 17:189–200. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Y, Huang H, Yuan B, Zhuang LY, Luo TP
and Zhang Q: Lentivirus-mediated knockdown of NOB1 suppresses the
proliferation of colon cancer cells. Z Gastroenterol. 52:429–435.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu K, Chen HL, Gu MM and You QS:
Relationship between NOB1 expression and prognosis of resected
non-small cell lung cancer. Int J Biol Markers. 30:e43–e48. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang X, Zhang D, Qu F, Hong Y, Cao J, Pan
X, Li L, Huang Y, Huang H, Yin L, et al: Knockdown of NOB1
expression inhibits the malignant transformation of human prostate
cancer cells. Mol Cell Biochem. 396:1–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Veith T, Martin R, Wurm JP, Weis BL,
Duchardt-Ferner E, Safferthal C, Hennig R, Mirus O, Bohnsack MT,
Wöhnert J and Schleiff E: Structural and functional analysis of the
archaeal endonuclease Nob1. Nucleic Acids Res. 40:3259–3274. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin Y, Peng S, Yu H, Teng H and Cui M:
RNAi-mediated downregulation of NOB1 suppresses the growth and
colony-formation ability of human ovarian cancer cells. Med Oncol.
29:311–317. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lin S, Meng W, Zhang W, Liu J, Wang P, Xue
S and Chen G: Expression of the NOB1 gene and its clinical
significance in papillary thyroid carcinoma. J Int Med Res.
41:568–572. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
He XW, Feng T, Yin QL, Jian YW and Liu T:
NOB1 is essential for the survival of RKO colorectal cancer cells.
World J Gastroenterol. 21:868–877. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang W, Zhong W, Xu J, Su B, Huang G, Du
J and Liu Q: Lentivirus-mediated gene silencing of NOB1 suppresses
non-small cell lung cancer cell proliferation. Oncol Rep.
34:1510–1516. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Moyer MP, Manzano LA, Merriman RL,
Stauffer JS and Tanzer LR: NCM460, a normal human colon mucosal
epithelial cell line. In Vitro Cell Dev Biol Anim. 32:315–317.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang P, Zuo Z, Wu A, Shang W, Bi R, Jin
Q, Wu J and Jiang L: miR-600 inhibits cell proliferation, migration
and invasion by targeting p53 in mutant p53-expressing human
colorectal cancer cell lines. Oncol Lett. 13:1789–1796.
2017.PubMed/NCBI
|
|
34
|
Wang Q, Zhu L, Jiang Y, Xu J, Wang F and
He Z: miR-219-5p suppresses the proliferation and invasion of
colorectal cancer cells by targeting calcyphosin. Oncol Lett.
13:1319–1324. 2017.PubMed/NCBI
|
|
35
|
Sheng N, Tan G, You W, Chen H, Gong J,
Chen D, Zhang H and Wang Z: MiR-145 inhibits human colorectal
cancer cell migration and invasion via PAK4-dependent pathway.
Cancer Med. 6:1331–1340. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ding L, Yu LL, Han N and Zhang BT: miR-141
promotes colon cancer cell proliferation by inhibiting MAP2K4.
Oncol Lett. 13:1665–1671. 2017.PubMed/NCBI
|
|
37
|
Liu Y, Liu R, Yang F, Cheng R, Chen X, Cui
S, Gu Y, Sun W, You C, Liu Z, et al: miR-19a promotes colorectal
cancer proliferation and migration by targeting TIA1. Mol Cancer.
16:532017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lu D, Yao Q, Zhan C, Le-Meng Z, Liu H, Cai
Y, Tu C, Li X, Zou Y and Zhang S: MicroRNA-146a promote cell
migration and invasion in human colorectal cancer via
carboxypeptidase M/src-FAK pathway. Oncotarget. 8:22674–22684.
2017.PubMed/NCBI
|
|
39
|
Zhou J, Xu T, Yan Y, Qin R, Wang H, Zhang
X, Huang Y, Wang Y, Lu Y, Fu D and Chen J: MicroRNA-326 functions
as a tumor suppressor in glioma by targeting the Nin one binding
protein (NOB1). PLoS One. 8:e684692013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Huang P, Xi J and Liu S: MiR-139-3p
induces cell apoptosis and inhibits metastasis of cervical cancer
by targeting NOB1. Biomed Pharmacother. 83:850–856. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ji S, Zhang B, Kong Y, Ma F and Hua Y:
MiR-326 inhibits gastric cancer cell growth through down regulating
NOB1. Oncol Res. 25:853–861. 2017. View Article : Google Scholar : PubMed/NCBI
|