Introduction
Glioma refers to malignant tumors that are
frequently found in the central nervous system (1), accounting for 45% of primary
intracranial tumors with an incidence rate of ~5/100,000 per year
(2). Despite the significant advance
in the surgical and medical treatment of this disease, the
prognosis for glioma patients is still poor (3). Therefore, the study of the
pathophysiology of glioma will help to develop new therapeutic
strategies.
Syndecan-1 (SDC1, also known as CD138) is a major
member of syndecans, which belong to the transmembrane heparan
sulphate proteoglycan (HSPG) family. HSPGs consist of an
extracellular domain carrying glycosaminoglycan (GAG) chains, a
transmembrane domain and a highly conserved cytoplasmic domain
(4). Studies have shown that SDC1
participates in diverse cellular processes, including cell
proliferation (5,6), migration (7,8), invasion
and angiogenesis (9,10). SDC1 expression is dysregulated in many
human cancers such as glioma and its expression is related to poor
prognosis and high metastatic potential of these cancers (11–20).
Syndecan binding protein (SDCBP, also known as syntenin), a
scaffold protein containing two PDZ domains, has been identified as
a molecule binding to the cytoplasmic domain of SDC (21). Recent studies on glioma have also
demonstrated that SDCBP is an important mediator of invasion
(22,23). SDC1 may form a functional complex with
SDCBP to play key roles in regulating tumor cell adhesion and
migration, as well as the biogenesis of exosomes (24,25). The
association of SDC1 and SDCBP in glioma has not be investigated.
Rac1, as a member of Rho GTPase family proteins, it regulates cell
migration, reorganization of cytoskeletal as well as cell
proliferation (26). The migration of
glioma cells was suppressed by the inactivation of Rac1 (27). The regulatory function of SDC1 and
SDCBP on Rac1 activity has been reported. Heparanase stimulated the
clustering of SDC1 and enhanced cell spreading, thus promoting cell
adhesion and spreading, by PKC, Src and Rac1 (28). SDCBP knockdown suppressed Rac1
activity and the invasion of breast cancer cells (29). It is not known whether SDC1/SDCBP
expression affects Rac1 activity in glioma.
In the present study, we compared SDC1 and SDCBP
expression among high grade gliomas, low grade gliomas and
non-tumorous brain tissues, and explored the correlation between
SDC1 and SDCBP expression in glioma tissues. We also manipulated
the expression of SDC1 in U251 glioma cells and studied the
functions of SDC1 and SDCBP expression, cell migration as well as
Rac1 activity. This study may provide an experimental basis for the
identification of novel therapeutic target in glioma.
Materials and methods
Tissue specimens
Archival formalin-fixed and paraffin-embedded tissue
specimens were obtained at the First People's Hospital of Zunyi and
the First Affiliated Hospital of Zunyi Medical College (Zunyi,
Guizhou, China). Thirty-two patients with glioma admitted from 2014
to 2016 were enrolled in this study. The glioma was confirmed by
pathological examination after operation. These patients included 7
cases of World Health Organization (WHO) level I glioma, 7
specimens of grade II glioma, 9 specimens of level III glioma as
well as 9 specimens of level IV glioma [glioblastoma multiforme
(GBM)]. The glioma patients did not receive any surgical treatment,
preoperative radiotherapy and/or chemotherapy before surgery.
Non-neoplastic brain tissues were available at the inside
decompression of five patients who received temporal lobe resection
for epilepsy. The study was approved by the Clinical Research
Ethics Committee of the First People's Hospital of Zunyi and the
First Affiliated Hospital of Zunyi Medical College (Zunyi). All
participants gave their written concent agreement.
Immunohistochemical analysis
Tissue specimens were sectioned, deparaffinized and
hydrated. After antigen retrieval, slices were incubated with
rabbit anti-human SDC1 (dilution, 1:100; cat. no. ab128936) or
SDCBP monoclonal antibody (dilution, 1:50; cat. no. ab133267) (both
from Abcam, Cambridge, MA, USA) instantly at 4°C. Then washed with
phosphate-buffered saline (PBS), the Donkey anti-Rabbit IgG H&L
secondary polyclonal antibody (dilution, 1:1,000; cat. no. ab98488;
Abcam) was applied for half an hour at 23°C. The slices were
stained with diaminobenzidine (DAB; Zhongshan, Beijing, China) and
counterstained with hematoxylin. Images were captured with Leica
Application Suite 4.6.0 program (Leica, Deerfield, IL, USA) at
magnification, ×200. Five fields were chosen at random for each
slice. Intensity of immunoreactivity (IRS) was scored as: IRS =
percentage of positive cells (PP) + staining intensity (SI). The SI
was 0, 1, 2 and 3, respectively, when the staining was negative,
weak, intermediate and powerful. The score of PP was 0, 1, 2, 3 and
4, respectively, when the percentage was <5, 5–25, 26–50, 51–75
and >75%. The IRS was set as 0–1, negative (−); 2 weakly
positive (+); 3–4 moderately positive (++); ≥5 strongly positive
(+++). The evaluation of staining results was carried out by two
investigators independently.
Cell culture and transfection
U251 cells were from China Center for Type Culture
Collection of Wuhan University (CCTCC; Wuhan, China) and cultured
in Eagle's Minimum Essential Medium (MEM; HyClone, Logan, UT, USA)
coupled with 10% fetal bovine serum (FBS; Gibco, Carlsbad, CA,
USA). U251 cells were infected with SDC1 expression plasmid
(SDC1-OE), control plasmid (vector) (Shanghai Genechem Co., Ltd.,
Shanghai, China), SDC1 siRNA (5-AGGAGGAAUUCUAUGCCUGA-3) or control
siRNA (GenePharma Co., Ltd., Shanghai, China) with Lipofectamine
2000 (Invitrogen, Carlsbad, CA, USA) following the manufacturer's
instruction. Two days after infection, transfected cells were
harvested and SDC1 expression was evaluated by western blot
analysis.
Western blot analysis
The cells were lysed with radioimmunoprecipitation
assay buffer (Beyotime, Shanghai, China). Besides, concentrations
of protein were quantified with BCA assay kit (Thermo Fisher
Scientific, Rockford, IL, USA). The extracted protein was
influenced by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) before being transferred onto the
membrane of a polyvinylidene difluoride (PVDF). Immune-detection of
specific proteins was performed with enhanced chemiluminescence
(ECL; Millipore, Bradford, PA, USA) following the incubation of
primary antibodies (rabbit anti-human SDC1; dilution, 1:200; cat.
no. ab128936) or SDCBP monoclonal antibody (dilution, 1:500; cat.
no. ab133267) and donkey anti-rabbit IgG H&L (HRP) (dilution,
1:1,000; cat. no. ab6802). All the important antibodies and
secondary antibodies were purchased from Abcam. Rabbit anti-human
β-actin (dilution, 1:1,000; cat. no. ab8227) or GAPDH monoclonal
antibody (dilution, 1:1,000; cat. no. ab9485) served as a loading
control. The Image Pro Plus 6.0 was used to quantify immunoblot
intensities (Media Cybernetics, Bethesda, MD, USA). The experiments
were performed three times.
Transwell assays
The migration ability of cells was assessed by
Transwell assay. U251 cells were transfected with plasmids and
siRNAs as described above. Forty-eight hours after transfection,
U251 cells (3×104/well) were seeded onto the upper
chamber (8-µm pore size; Corning, Lowell, MA, USA) containing MEM.
The lower chamber contained MEM with 10% FBS. After being incubated
for 24 h, non-migrated cells were completely removed, then stain
and fix the remaining cells with crystal violet. The cell numbers
were calculated by using a microscope (Nikon, Tokyo, Japan) in five
sections for each group. Carry out the experiment in triplicate and
repeat it for three times independently.
Scratch-wound healing assay
Cells migration capacity was assessed with a
scratch-wound healing assay. Cells were transfected with plasmids
and siRNAs and grown to >90% confluence. A linear wound was
created by scraping the cell monolayer with a sterile 200 µl
pipette tip. Then, cells were then cleaned up with MEM three times
and cultured for 0 and 12 h before being photographed with a
microscope (Nikon). The formula is the percentage of wound closure:
(original wound area actual wound area)/area of the original wound
×100. The experiment was repeated three times.
Pull-down assay of GST-PAK1-binding
domain (GST-PBD)
U251 cells in 10 cm culture dishes were transfected
with plasmids or siRNAs as described above. At 48 h
post-transfection, GST-PBD pull-down assay was carried out using
active Rac1 pull-down and detection kit (Thermo Fisher Scientific)
following the manufacturer's protocol. The precipitated samples
were analyzed via western blotting.
Statistical analysis
SPSS software was used to carry out statistical
analysis (version 17.0; SPSS Inc., Chicago, IL, USA). Ordered
variables in different groups were analyzed with Kruskal-Wallis H
test. Mann-Whitney U test as well as Bonferroni modification
(α=0.0167) was used to compare the difference between two groups.
Associations between two factors were assessed by Spearmans
correlation analysis. One-way ANOVA followed by LSD-t experiment
(for the same variance) or Tamhane's T2 experiment (for different
variance) was used to analyze the mean values for different groups
(α=0.05). P-values <0.05 were regarded as statistically
significant.
Results
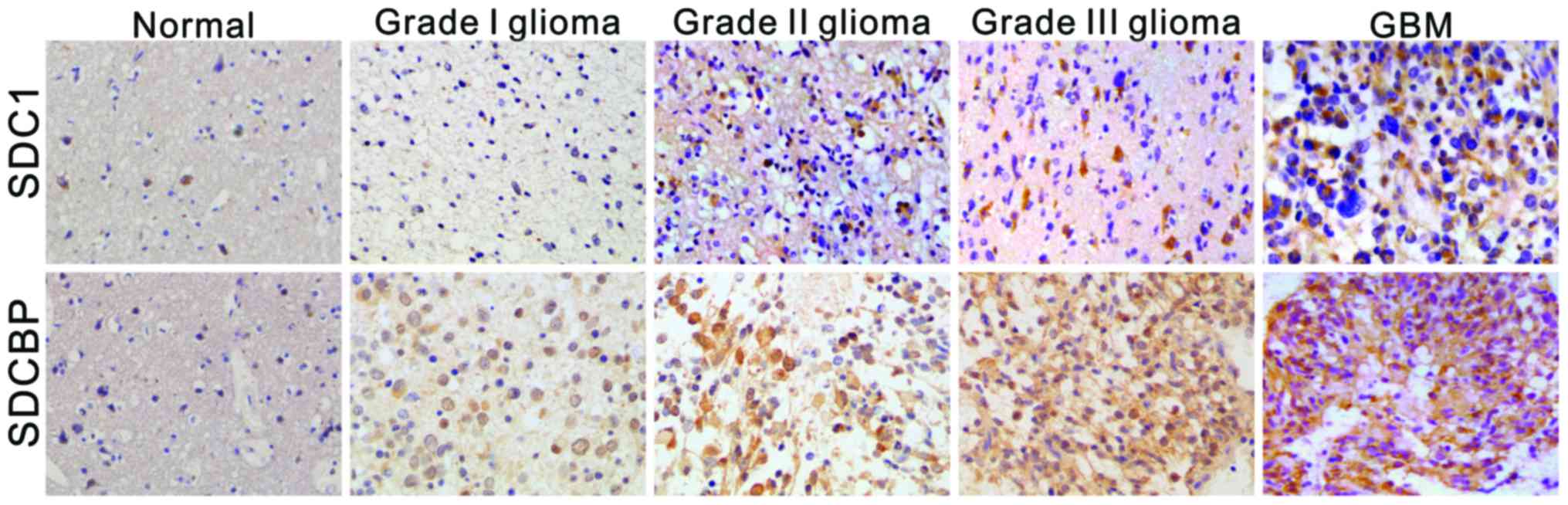
SDC1 and SDCBP are upregulated in
human glioma tissues
Immunohistochemical analysis was carried out in 32
glioma and 5 non-tumorous brain tissues with antibodies against
SDC1 or SDCBP (Fig. 1). SDC1 mainly
located in the cell membrane and cytoplasm, while SDCBP localized
in the cytoplasm. In non-tumorous brain tissues, the signals were
weak and the positive-staining cells were scattered. In low grade
glioma tissues (grade I–II), the signals were weak to moderate.
High grade gliomas (grade III–IV) had the strongest staining
intensity and the highest positive-staining cell number. The
positive rate of SDC1 in non-tumorous brain tissues, low level
gliomas and high level gliomas were 20.0% (1/5), 50.0% (7/14) and
88.9% (16/18), respectively. The positive rate of SDCBP in
non-tumorous brain tissues, low level gliomas as well as high level
gliomas were 0% (0/5), 42.9% (6/14) and 88.9% (16/18),
respectively. By Kruskal-Wallis H test (Table I), we found that the expression of
SDC1 and SDCBP showed significant difference among the three
groups. According to the findings of Mann-Whitney U test
experiment, there was an obvious difference in the expression of
SDC1 as well as SDCBP between high grade gliomas and control brain
tissues, as well as between high grade gliomas and low grade
gliomas. There was no significant difference of SDC1 and SDCBP
expression between low level gliomas and control brain tissues.
Moreover, SDC1 and SDCBP expression was increased with the
malignant grade of glioma, by Spearmans correlation analysis (SDC1,
rs=0.576, P=0.001; SDCBP, rs=0.661,
P<0.001).
 | Table I.Results of immunohistochemical
staining. |
Table I.
Results of immunohistochemical
staining.
|
|
|
|
|
| Kruskal-Wallis H
test | Mann-Whitney U
test |
|---|
|
|
|
|
|
|
|
|
|---|
| Protein | Expression | Control (n=5) | I–II (n=14) | III–IV (n=18) | H | P-value |
| Z | P-value |
|---|
| SDC1 | − | 4 | 7 | 2 | 12.784 | 0.002a | III–IV vs.
control | −2.852 | 0.003c |
|
| + | 1 | 4 | 4 |
|
| III–IV vs.
I–II | −2.806 | 0.004c |
|
| ++ | 0 | 2 | 5 |
|
| I–II vs.
control | −1.251 | 0.305 |
|
| +++ | 0 | 1 | 7 |
|
|
|
|
|
| SDCBP | − | 5 | 8 | 2 | 16.27 |
<0.001b | III–IV vs.
control | −3.193 | 0.001c |
|
| + | 0 | 3 | 2 |
|
| III–IV vs.
I–II | −3.120 | 0.001c |
|
| ++ | 0 | 2 | 6 |
|
| I–II vs.
control | −1.689 | 0.175 |
|
| +++ | 0 | 1 | 8 |
|
|
|
|
|
SDC1 expression was positively related
to SDCBP expression in glioma tissues
Spearman experiment was then employed to assess the
relationship between SDC1 expression as well as SDCBP expression in
glioma tissues (Table II). The
results showed a positive relation between protein levels of SDC1
and SDCBP among the glioma tissues (rs=0.628,
P=0.001).
| Table II.Correlation analysis between the
protein levels of SDC1 and SDCBP in glioma tissues. |
Table II.
Correlation analysis between the
protein levels of SDC1 and SDCBP in glioma tissues.
|
| SDCBP |
|
|
|---|
|
|
|
|
|
|---|
| SDC1 | Positive | Negative | rs | P-value |
|---|
| Positive | 20 | 3 | 0.628 | 0.001a |
| Negative | 2 | 7 |
|
|
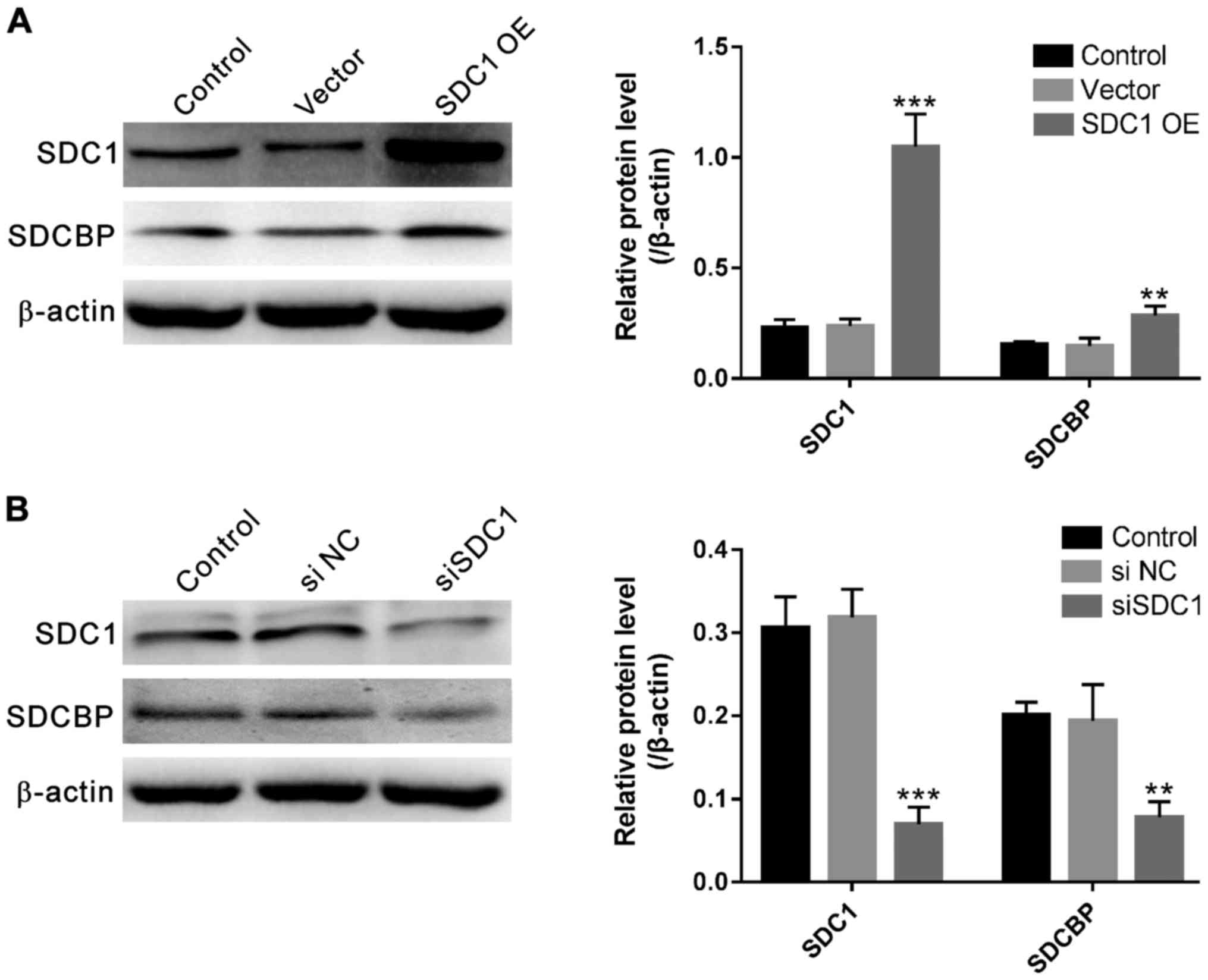
SDC1 affects SDCBP expression in U251
cells
U251 cells were infected with SDC1 expression
plasmid (SDC1-OE), control plasmid (vector), SDC1 siRNA (siSDC1) or
control siRNA (siNC). Forty-eight hours after the infection, SDC1
as well as SDCBP's protein levels were determined by western
blotting. As shown in Fig. 2A, SDC1
was significantly increased in U251 cells transfected with SDC1-OE
(1.049±0.146) as compared to control cells (0.228±0.038) or cells
transfected with vector (0.237±0.031) (P<0.001). The levels of
protein in SDCBP were also increased in cells transfected with
SDC1-OE (SDC1-OE, 0.284±0.044; control, 0.153±0.013; vector,
0.146±0.036; P=0.004). The protein levels of SDC1 and SDCBP in
cells transfected with vector had no significant difference
compared to control cells. On the contrary, SDC1 siRNA transfection
significantly decreased the expression of SDC1 (SDC1-OE,
0.069±0.021; control, 0.306±0.037; vector, 0.318±0.033; P<0.001)
and SDCBP (siSDC1, 0.077±0.019; control, 0.201±0.015; siNC,
0.193±0.044; P=0.004) (Fig. 2B). siNC
transfection had no effects on the expression of the proteins as
compared to control cells. These data suggested the association
between the protein expression of SDC1 and SDCBP in glioma
cells.
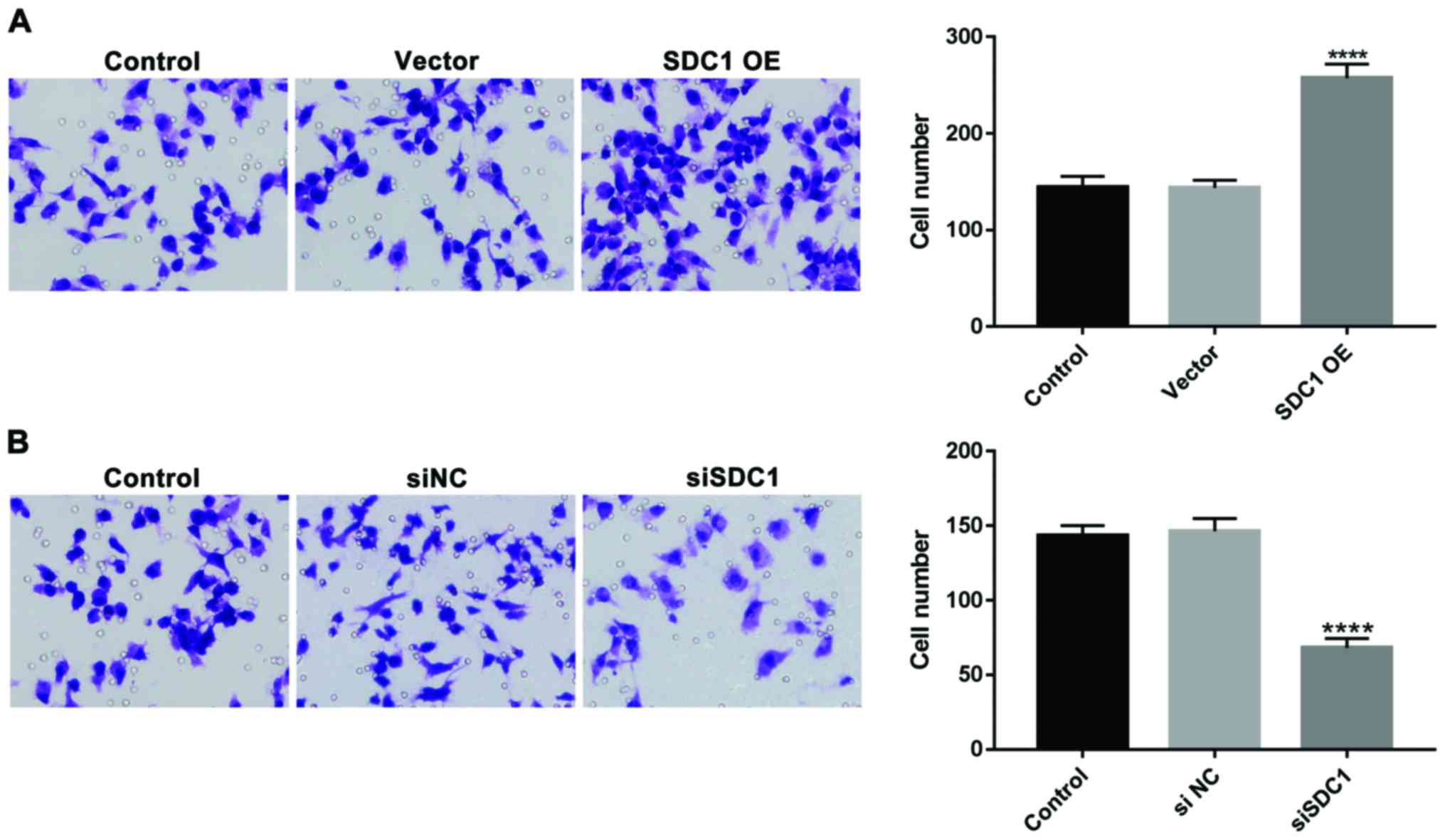
Effects of SDC1 expression on the
migration ability of U251 cells
To determine whether SDC1 influences the migration
of U251 cells, Transwell assay (Fig.
3) and Scratch-wound healing assay (Fig. 4) were performed. U251 cells
transfected with SDC1-OE showed a significant increase in cell
migration compared to vector-transfected cells or control cells
(SDC1-OE, 257±15; control, 144±11; vector, 144±8; P<0.001).
Complementary to the results from SDC1 overexpression, SDC1
knockdown remarkably reduced the migration of U251 cells (siSDC1,
68±6; control, 143±7; siNC, 146±9; P<0.001).
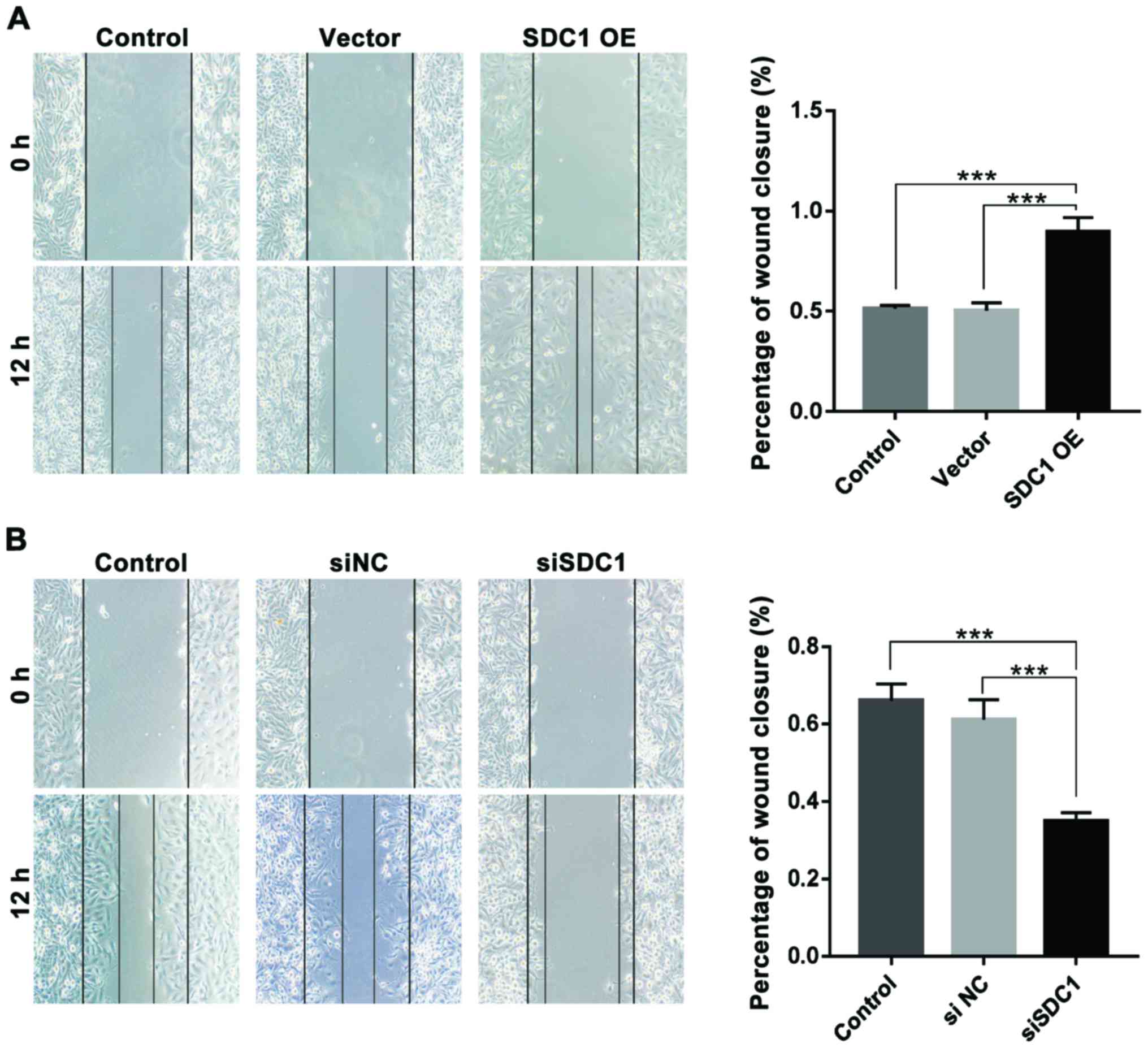
Scratch-wound healing assay showed that SDC1
overexpression significantly enhanced the migration of U251 cells
(SDC1-OE, 89.74±6.94%; control, 51.32±1.56%; vector, 50.36±3.78;
P<0.001) (Fig. 4A). On the
contrary, U251 cells transfected with siSDC1 displayed a notable
decrease in the migration (siSDC1, 34.89±2.19%; control,
66.06±4.29%; siNC, 61.16±5.16%; P<0.001) (Fig. 4C).
Effects of SDC1 expression on the
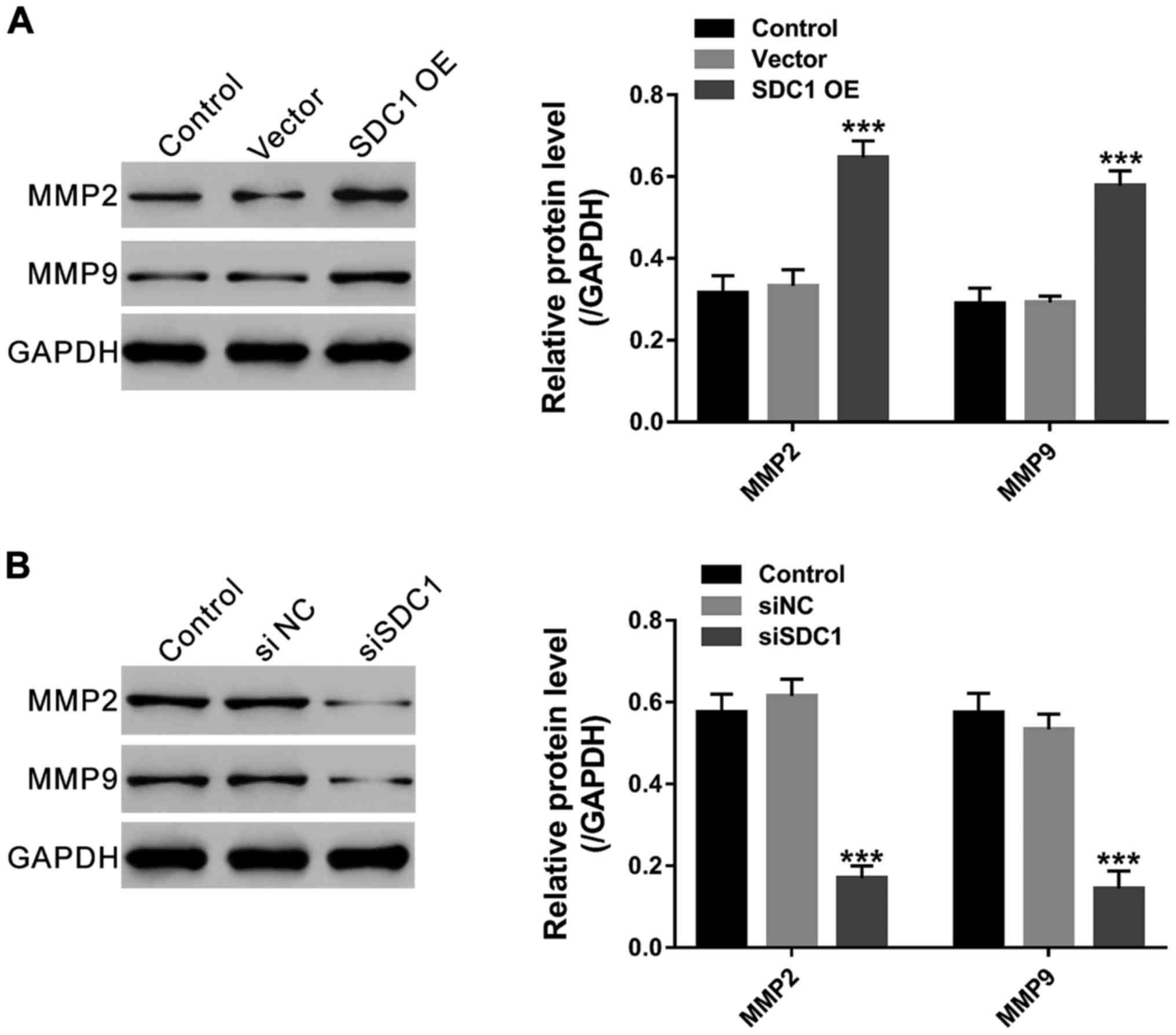
expression of matrix metalloproteinase 2 (MMP2) and MMP9
The expression of MMP2 and MMP9, two major MMP
associated with cell migration (30),
was detected. As illustrated in Fig.
5, MMP2 as well as MMP9 expression was significantly increased
in SDC1 overexpressed cells, but remarkably decreased in SDC1
knockdown cells.
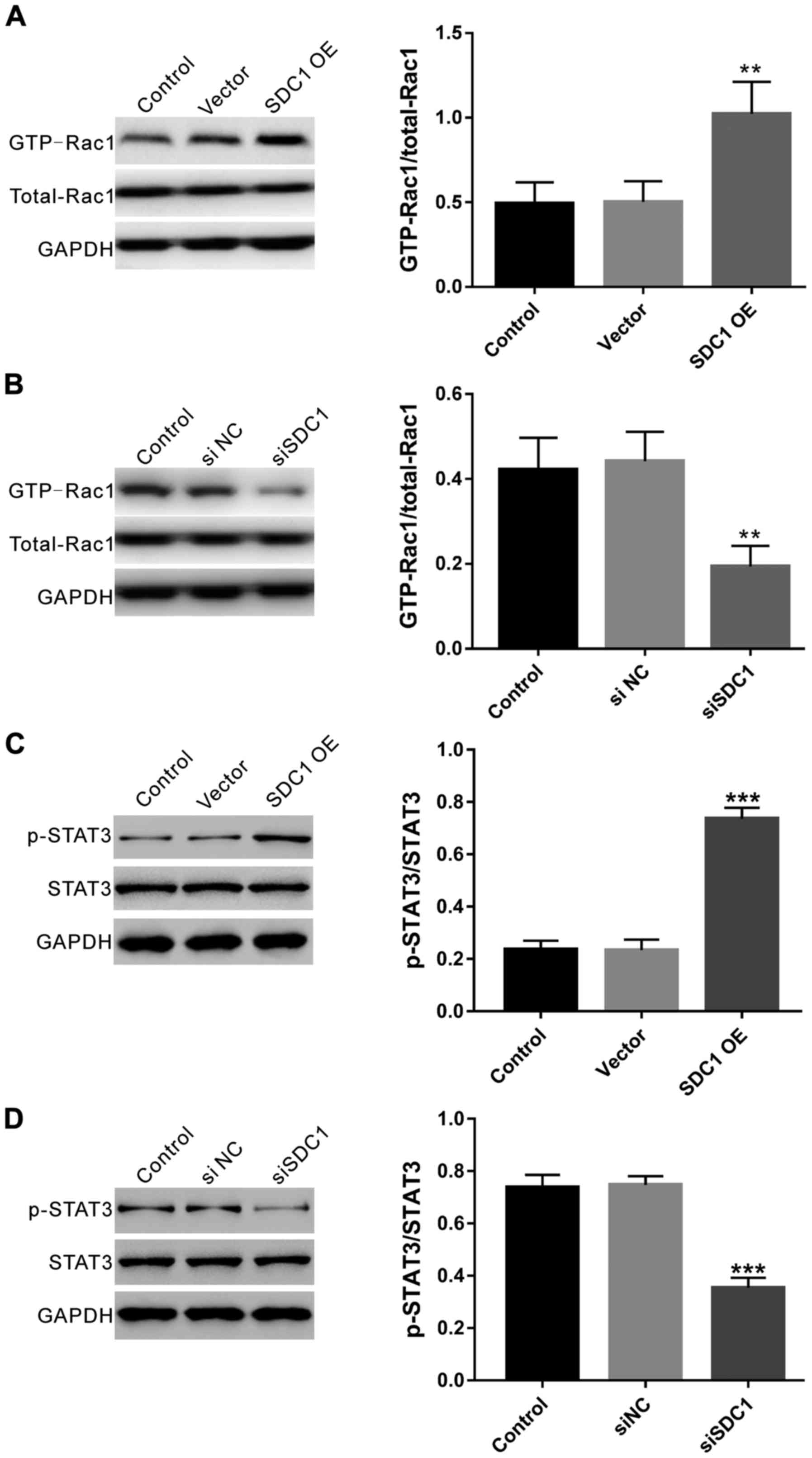
Effects of SDC1 expression on Rac1
activity and STAT3 phosphorylation
GST-PBD pull-down assay was conducted to test Rac1
activity, which is critical for glioma cell migration (27). According to Fig. 6A, ectopic expression of SDC1 notably
induced Rac1 activity (SDC1-OE, 1.02±0.19; control, 0.49±0.12;
vector, 0.50±0.12; P=0.007), while Rac1 expression was not affected
by SDC1. Complementary data were obtained in SDC1 knockdown cells
(siSDC1, 0.197±0.048; control, 0.422±0.074; siNC, 0.442±0.070;
P=0.006). Judging from these data, SDC1 may influence glioma by
inducing Rac1 activity.
Signal transducer and activator of transcription 3
(STAT3) has been identified to be essential for activation of Rac1
for the regulation of cell proliferation and migration (31). As shown in Fig. 6B, SDC1 overexpression notably
increased the levels of phosphorylated STAT3 (SDC1-OE, 0.74±0.04;
control, 0.24±0.04; vector, 0.23±0.03; P<0.0001), while
complementary data were obtained in SDC1 knockdown cells (siSDC1,
0.35±0.04; control, 0.74±0.05; siNC, 0.75±0.03; P<0.0001).
Discussion
SDC1 is the most studied member of the HPSGs.
Deregulated SDC1 expression has been used as a prognostic factor in
human malignancies. Decreased SDC1 expression is a negative
prognostic factor in colorectal (13), head and neck (14) and hepatocellular carcinoma (18). In contrast, strong stromal SDC1
expression has been shown to correlate with a poor prognosis in
patients with glioma (12), prostate
(11), breast (17), ovarian (19) and gastric cancer (20). Studies have shown that SDC1 binds to
its partners through the cytoplasmic domain, thus involving in cell
proliferation (5,6), migration (7,8), invasion
and angiogenesis (9,10). The EFYA motif is crucial for the
interaction of SDC and PDZ-domain proteins, including SDCBP
(21). SDCBP was overexpressed in
various tumor samples (32–35). In the present study, by
immunohistochemcal staining, the expression levels of SDC1 and
SDCBP in glioma tissues were higher than in normal brain tissues,
and the expression levels of both proteins were increased with the
malignant grade of glioma, consistently with previous findings
(12,35). Importantly, the protein levels of SDC1
and SDCBP were positively correlated in glioma tissues. We then
overexpressed or knocked down SDC1 expression in U251 glioma cells,
and found that SDCBP expression was substantially affected by the
manipulation of SDC1 expression. These data suggested an
association between SDC1 and SDCBP during glioma tumorigenesis.
Migration is the critical process for metastasis of
cancer. SDC1 is actively involved in cell migration (7,8) and
angiogenesis (9,10). SDCBP can promote the invasion of
glioma cells (22,23). MMP2 and MMP9 are major MMPs associated
with cell migration (30). In the
present study, ectopic expression of SDC1 in glioma cells
significantly enhanced their migration ability and the expression
of MMP2/9 as indicated by Transwell assay and western blotting,
respectively, which provided evidence that SDC1 expression was
associated with metastatic potential of glioma.
Rac1 and its downstream effector STAT3 have been
suggested to be a part of regulating cell migration (27,31).
Previous studies have shown that SDCBP and SDC1 could regulate Rac1
activity (28,29). Here, we found that ectopic expression
of SDC1 in U251 cells significantly increased SDCBP expression,
Rac1 activity and STAT3 phosphorylation. Complementary data were
obtained in SDC1 knockdown cells. These findings indicated that
SDC1 induced SDCBP expression and Rac1/STAT3 pathway, and that the
association between SDC1 and SDCBP may promote the development of
glioma.
In conclusion, SDC1 may promote the migration of
glioma cells by forming a complex with SDCBP and activating the
Rac1 signaling pathway. SDC1 and SDCBP may be clinical markers for
glioma progression, prognosis, and treatment outcome evaluation.
However, tumor metastasis is a complicated process, which require a
series of orderly steps (36).
Further detailed investigation is needed to explore how SDC1 and
SDCBP interact with tumor microenvironment and participate in
glioma carcinogenesis.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (no. 81660421), the High-level
Personnel Funded Projects, the Science and Technology Benefits of
Zunyi and the First People's Hospital of Zunyi Joint Research and
Development of Science and Technology Projects [grant no.
(2011)24].
References
|
1
|
Louis DN, Perry A, Reifenberger G, von
Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD,
Kleihues P and Ellison DW: The 2016 World Health Organization
classification of tumors of the central nervous system: A summary.
Acta Neuropathol. 131:803–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dolecek TA, Propp JM, Stroup NE and
Kruchko C: CBTRUS statistical report: Primary brain and central
nervous system tumors diagnosed in the United States in 2005–2009.
Neuro Oncol. 14 Suppl 5:v1–v49. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johnson DR and O'Neill BP: Glioblastoma
survival in the United States before and during the temozolomide
era. J Neurooncol. 107:359–364. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bernfield M, Götte M, Park PW, Reizes O,
Fitzgerald ML, Lincecum J and Zako M: Functions of cell surface
heparan sulfate proteoglycans. Annu Rev Biochem. 68:729–777. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Beauvais DM, Burbach BJ and Rapraeger AC:
The syndecan-1 ectodomain regulates alphavbeta3 integrin activity
in human mammary carcinoma cells. J Cell Biol. 167:171–181. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu W, Litwack ED, Stanley MJ, Langford
JK, Lander AD and Sanderson RD: Heparan sulfate proteoglycans as
adhesive and anti-invasive molecules. Syndecans and glypican have
distinct functions. J Biol Chem. 273:22825–22832. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Beauvais DM and Rapraeger AC:
Syndecan-1-mediated cell spreading requires signaling by
alphavbeta3 integrins in human breast carcinoma cells. Exp Cell
Res. 286:219–232. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stepp MA, Daley WP, Bernstein AM,
Pal-Ghosh S, Tadvalkar G, Shashurin A, Palsen S, Jurjus RA and
Larsen M: Syndecan-1 regulates cell migration and fibronectin
fibril assembly. Exp Cell Res. 316:2322–2339. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Beauvais DM, Ell BJ, McWhorter AR and
Rapraeger AC: Syndecan-1 regulates alphavbeta3 and alphavbeta5
integrin activation during angiogenesis and is blocked by
synstatin, a novel peptide inhibitor. J Exp Med. 206:691–705. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Purushothaman A, Uyama T, Kobayashi F,
Yamada S, Sugahara K, Rapraeger AC and Sanderson RD:
Heparanase-enhanced shedding of syndecan-1 by myeloma cells
promotes endothelial invasion and angiogenesis. Blood.
115:2449–2457. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zellweger T, Ninck C, Mirlacher M,
Annefeld M, Glass AG, Gasser TC, Mihatsch MJ, Gelmann EP and
Bubendorf L: Tissue microarray analysis reveals prognostic
significance of syndecan-1 expression in prostate cancer. Prostate.
55:20–29. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu Y, Yuan J, Zhang Z, Lin L and Xu S:
Syndecan-1 expression in human glioma is correlated with advanced
tumor progression and poor prognosis. Mol Biol Rep. 39:8979–8985.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wei HT, Guo EN, Dong BG and Chen LS:
Prognostic and clinical significance of syndecan-1 in colorectal
cancer: A meta-analysis. BMC Gastroenterol. 15:1522015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Anttonen A, Kajanti M, Heikkilä P,
Jalkanen M and Joensuu H: Syndecan-1 expression has prognostic
significance in head and neck carcinoma. Br J Cancer. 79:558–564.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Joensuu H, Anttonen A, Eriksson M,
Mäkitaro R, Alfthan H, Kinnula V and Leppä S: Soluble syndecan-1
and serum basic fibroblast growth factor are new prognostic factors
in lung cancer. Cancer Res. 62:5210–5217. 2002.PubMed/NCBI
|
|
16
|
Jary M, Lecomte T, Bouché O, Kim S, Dobi
E, Queiroz L, Ghiringhelli F, Etienne H, Léger J, Godet Y, et al:
Prognostic value of baseline seric Syndecan-1 in initially
unresectable metastatic colorectal cancer patients: A simple
biological score. Int J Cancer. 139:2325–2335. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Barbareschi M, Maisonneuve P, Aldovini D,
Cangi MG, Pecciarini L, Mauri Angelo F, Veronese S, Caffo O,
Lucenti A, Palma PD, et al: High syndecan-1 expression in breast
carcinoma is related to an aggressive phenotype and to poorer
prognosis. Cancer. 98:474–483. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Matsumoto A, Ono M, Fujimoto Y, Gallo RL,
Bernfield M and Kohgo Y: Reduced expression of syndecan-1 in human
hepatocellular carcinoma with high metastatic potential. Int J
Cancer. 74:482–491. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kusumoto T, Kodama J, Seki N, Nakamura K,
Hongo A and Hiramatsu Y: Clinical significance of syndecan-1 and
versican expression in human epithelial ovarian cancer. Oncol Rep.
23:917–925. 2010.PubMed/NCBI
|
|
20
|
Wiksten JP, Lundin J, Nordling S, Lundin
M, Kokkola A, von Boguslawski K and Haglund C: Epithelial and
stromal syndecan-1 expression as predictor of outcome in patients
with gastric cancer. Int J Cancer. 95:1–6. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Grootjans JJ, Zimmermann P, Reekmans G,
Smets A, Degeest G, Dürr J and David G: Syntenin, a PDZ protein
that binds syndecan cytoplasmic domains. Proc Natl Acad Sci USA.
94:pp. 13683–13688. 1997, View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kegelman TP, Das SK, Hu B, Bacolod MD,
Fuller CE, Menezes ME, Emdad L, Dasgupta S, Baldwin AS, Bruce JN,
et al: MDA-9/syntenin is a key regulator of glioma pathogenesis.
Neuro Oncol. 16:50–61. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhong D, Ran JH, Tang WY, Zhang XD, Tan Y,
Chen GJ, Li XS and Yan Y: Mda-9/syntenin promotes human brain
glioma migration through focal adhesion kinase (FAK)-JNK and
FAK-AKT signaling. Asian Pac J Cancer Prev. 13:2897–2901. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sulka B, Lortat-Jacob H, Terreux R,
Letourneur F and Rousselle P: Tyrosine dephosphorylation of the
syndecan-1 PDZ binding domain regulates syntenin-1 recruitment. J
Biol Chem. 284:10659–10671. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Baietti MF, Zhang Z, Mortier E, Melchior
A, Degeest G, Geeraerts A, Ivarsson Y, Depoortere F, Coomans C,
Vermeiren E, et al: Syndecan-syntenin-ALIX regulates the biogenesis
of exosomes. Nat Cell Biol. 14:677–685. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Parri M and Chiarugi P: Rac and Rho
GTPases in cancer cell motility control. Cell Commun Signal.
8:232010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sun Z, Zhang B, Wang C, Fu T, Li L, Wu Q,
Cai Y and Wang J: Forkhead box P3 regulates ARHGAP15 expression and
affects migration of glioma cells through the Rac1 signaling
pathway. Cancer Sci. 108:61–72. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Levy-Adam F, Feld S, Suss-Toby E,
Vlodavsky I and Ilan N: Heparanase facilitates cell adhesion and
spreading by clustering of cell surface heparan sulfate
proteoglycans. PLoS One. 3:e23192008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hwangbo C, Park J and Lee JH:
mda-9/Syntenin protein positively regulates the activation of Akt
protein by facilitating integrin-linked kinase adaptor function
during adhesion to type I collagen. J Biol Chem. 286:33601–33612.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nabeshima K, Inoue T, Shimao Y and
Sameshima T: Matrix metalloproteinases in tumor invasion: Role for
cell migration. Pathol Int. 52:255–264. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Arulanandam R, Geletu M, Feracci H and
Raptis L: Activated Rac1 requires gp130 for Stat3 activation, cell
proliferation and migration. Exp Cell Res. 316:875–886. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Oyesanya RA, Bhatia S, Menezes ME, Dumur
CI, Singh KP, Bae S, Troyer DA, Wells RB, Sauter ER, Sidransky D,
et al: MDA-9/Syntenin regulates differentiation and angiogenesis
programs in head and neck squamous cell carcinoma. Oncoscience.
1:725–737. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Helmke BM, Polychronidis M, Benner A,
Thome M, Arribas J and Deichmann M: Melanoma metastasis is
associated with enhanced expression of the syntenin gene. Oncol
Rep. 12:221–228. 2004.PubMed/NCBI
|
|
34
|
Yang Y, Hong Q, Shi P, Liu Z, Luo J and
Shao Z: Elevated expression of syntenin in breast cancer is
correlated with lymph node metastasis and poor patient survival.
Breast Cancer Res. 15:R502013. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhong D, Tang W, Wu H, Yang G and Yan Y:
Expression of syntenin in human glioma and its clinical
significance: Report of 34 cases. Di 3 Jun Yi Da Xue Xue Bao.
32:597–600. 2010.(In Chinese).
|
|
36
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|