Introduction
Fatty acid synthase (FASN) is a multi-enzyme that
catalyzes the de novo synthesis of palmitate (C16:0, a
long-chain saturated fatty acid) from acetyl-CoA and malonyl-CoA,
in the presence of NADPH (1). FASN is
not only a key factor in the role of fatty acid biosynthesis for
energy storage (2,3), but also its expression level increases
significantly in adipose tissues and a variety of human carcinomas,
including liver, breast, prostate, lung, endometrium, ovary, colon
and pancreatic cancer (4–13). This prominent difference of FASN
expression between normal and neoplastic tissues makes FASN a
potential diagnostic tumor marker (14).
Breast cancer is the most common type of cancer and
a leading cause of cancer-associated mortalities among females,
with a common feature of abnormal cell apoptosis in its development
(15). In addition, high levels of
FASN expression has been demonstrated to be associated with poor
clinical outcome in breast carcinomas, suggesting that FASN
expression and tumor aggressiveness are closely associated
(4,16). It was identified that obesity may
serve a crucial role in the incidence and progression of breast
cancer (17). According to the close
association between FASN, obesity and breast cancer, the studies of
FASN inhibitors have indicated their role as targets for
chemotherapy in breast cancer and a novel strategy for
antineoplastic intervention (18). In
fact, previous studies demonstrated that certain synthetic and
natural FASN inhibitors, including C75, desoxyrhaponticin,
rhaponticin and α-mangostin may lead to selective cytotoxicity in
FASN over-expressing cancer cell lines (18–20). This
result suggested again that the pharmacological inhibition of FASN
may represent a potential target for drug development.
In previous studies, a number of dietary
polyphenols, including α-mangostin, resveratrol, curcumin and
quercetin exhibited high inhibitory activity against FASN (21–30).
Although the detailed mechanism of the inhibitory effect of
polyphenols on FASN was not fully understood, the structure
activity association analysis demonstrated that the flavonoids
containing two hydroxyl groups in the B ring and 5, 7-hydroxyl
groups in the A ring with C-2, 3 double bond were the most potent
inhibitors on FASN (31). Patuletin
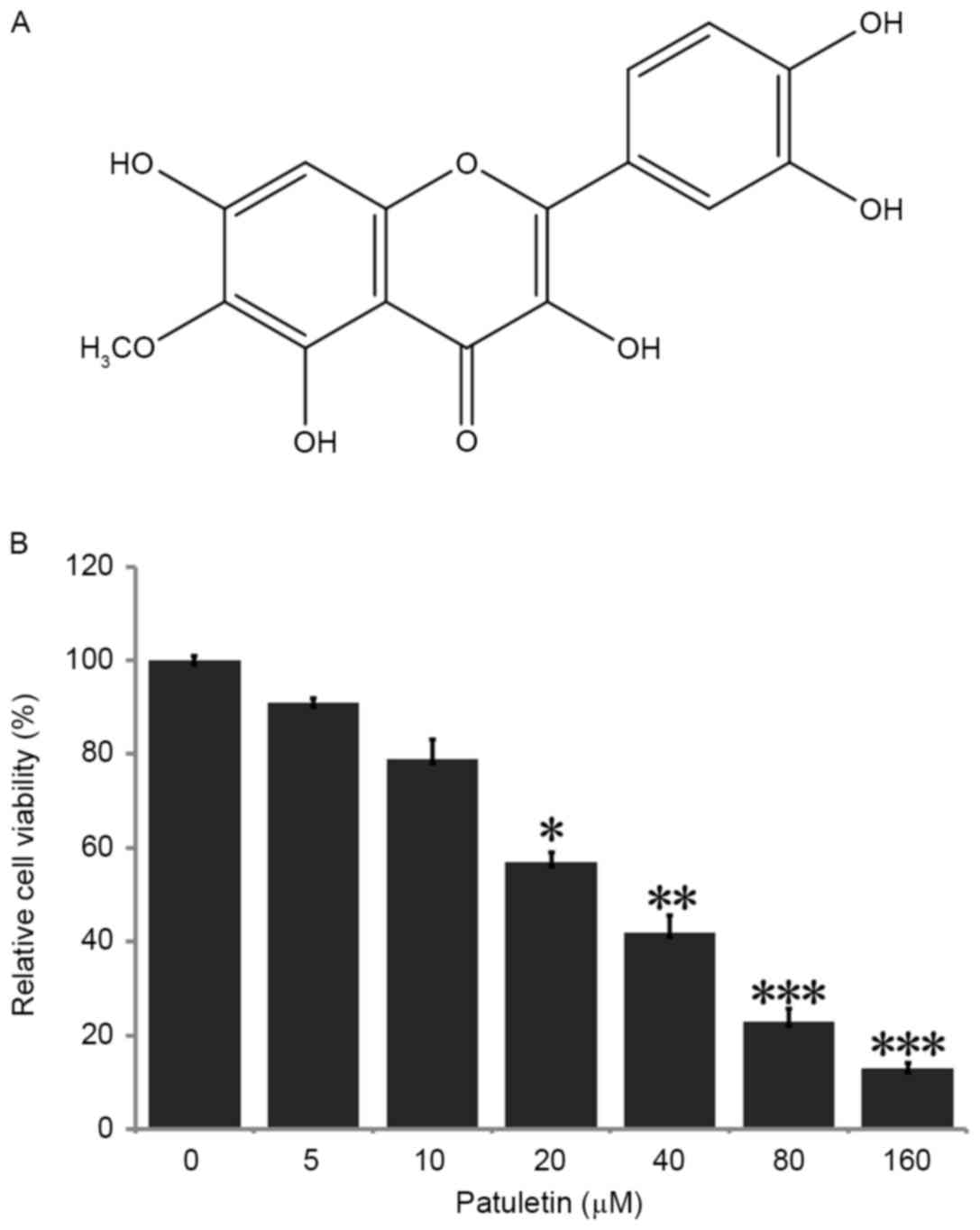
(3,5,7,3′,4′-pentahydroxy-6-methoxy-flavone) (Fig. 1A), a natural flavonoid primarily
present in the genus Eriocaulon (32), exhibits anti-inflammatory,
anti-oxidant and anti-bacterial properties (33–36).
However, no anti-neoplastic effects of patuletin have been
identified at present. Increased expression of FASN has previously
been demonstrated in different breast cancer cells (37). Among them, human breast cancer SK-BR-3
cells exhibited higher expression levels of FASN compared with
other breast cancer cells, including MCF-7 and MDA-MB-231 cells.
The present study aimed to identify for the first time that
patuletin induces apoptosis in FASN over-expressing human breast
cancer SK-BR-3 cells.
Materials and methods
Reagents
Acetyl-Coenzyme A (CoA), Malonyl-CoA, dexamethasone,
NADPH, ethyl acetate (EtOAc), chloroform, methanol, MTT,
3-isobutyl-1-methylxanthine (IBMX), EDTA and DTT were all purchased
from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Annexin
V-fluorescein isothiocyanate (FITC) Apoptosis Detection kit was
purchased from BD Biosciences (San Jose, CA, USA). Dulbecco's
modified Eagle's medium (DMEM), fetal bovine serum, PBS,
penicillin-streptomycin, trypsin-EDTA, dimethyl sulfoxide (DMSO),
TRIzol, SuperScript III First-Strand Synthesis system were
purchased from Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
Rabbit antibodies against FASN (cat no. 3180) and β-actin (cat no.
4967) were purchased from Cell Signaling Technology, Inc. (Danvers,
MA, USA).
Extraction and isolation of
patuletin
The whole plants of Eriocaulon buergerianum
were collected in Zhejiang (China) by the research group. Patuletin
from air-dried whole plants of E. buergerianum (3.0 kg) was
extracted with 95% ethanol at room temperature. The concentrated
crude extract was dissolved in H2O and partitioned with
EtOAc. The EtOAc portion (225 g) was chromatographed on a silica
gel column eluting with a chloroform-methanol gradient system to
yield patuletin (300 mg). Isolated patuletin was ≥98% pure as
determined by HPLC-UV (Agilent Technologies, Inc., Santa Clara, CA,
USA).
Cell culture
The human breast cancer SK-BR-3 cell line was
purchased from the Cell Bank of the Chinese Academy of Sciences
(Shanghai, China). Cells were incubated at 5% CO2 and
37°C in a medium containing 89% DMEM (high glucose), 10% bovine
fetal serum and 100 U/ml penicillin-streptomycin. For passage, the
cells were digested by 0.25% trypsin-EDTA every 4 days.
Cell viability assay
SK-BR-3 cells were seeded in 96-well plate firstly,
at a density about 5×103 cells/well and then treated
with purified patuletin in different concentrations (5, 10, 20, 40,
80 or 160 µM) for 24 h. Thereafter, 5 mg/ml MTT solution was added
into each well and incubated for 4 h at 37°C. Then, the medium with
MTT was aspirated, 200 µl DMSO/well was added to the wells and the
cells were incubated for 15 min. Finally, the concentration was
measured at 492 nm by a microplate spectrophotometer (BioTek China,
Beijing, China). PBS was used as blank control, and cells without
patuletin treatment were used as negative control.
Cell lysis and immunoblotting
Cells were lysed as previously described (38). Protein concentration of cell lysates
was measured by the Pierce BCA protein assay kit using bovine serum
albumin as a standard control. 50 µg protein was loaded per lane,
separated by SDS-PAGE (12% gel), and then electrophoretically
transferred to polyvinylidene difluoride membranes (EMD Millipore,
Billerica, MA, USA). Then the protein samples were blocked with 5%
skimmed milk for 1–2 h at room temperature to prevent nonspecific
antibody binding, and probed with primary antibodies against FASN
and β-actin at a dilution of 1:1,000 overnight at 4°C.
Subsequently, membranes were washed twice with TBST (10 mM Tris, 10
mM NaCl, 0.1% Tween-20), and incubated 1 h at room temperature with
corresponding peroxidase conjugated secondary antibody (cat no.
7074) and developed with a commercial enhanced chemiluminescence
kit (West Pico chemiluminescent substrate; GE Healthcare
Bio-Sciences, Pittsburgh, PA, USA) according to the manufacturer's
protocol. Blots were probed with an antibody against β-actin as the
control.
Cell apoptosis assay
Cell apoptosis detection was performed using an
Annexin V-FITC Apoptosis Detection kit (BD Biosciences) according
to the manufacturer's protocol. First, cells were collected after
24 h treatment with patuletin at different concentrations (20, 40
and 80 µM). Then, cells (1×106 cells/tube) were washed
twice with cold PBS and resuspended in 100 µl 1X binding buffer
(Biomiga Inc., San Diego, CA, USA). Cell suspension was incubated
with 5 µl Annexin V-FITC and 10 µl propidium iodide (PI) for 15 min
at room temperature and kept in a dark place. Immediately following
that, 400 µl 1X binding buffer was added and the cells were
analyzed by a CellQuest Pro software (FACSstation 6.0; BD
Biosciences) in a BD FACSCalibur™ flow cytometer (BD
Biosciences) within 1 h. Those cells stained with Annexin
V+/PI− were early apoptotic cells and those
stained with Annexin V+/PI+ were late
apoptotic cells.
FASN gene expression analysis
FASN gene expression analysis was performed in
SK-BR-3 cells treated with patuletin at different concentrations
(5, 10, 20, 40, 80 and 160 µM) for 24 h. Cells were washed with PBS
twice for RNA extraction. Total RNA was isolated from SK-BR-3 cells
using TRIzol reagent (Thermo Fisher Scientific, Inc.), following
the manufacturer's protocol. A total of ~2 µg RNA was reverse
transcribed into complementary DNA (cDNA) using SuperScript III
First-Strand Synthesis system (Thermo Fisher Scientific, Inc.),
from the control and treated cells. Polymerase chain reaction (PCR)
was performed in 20 µl of the final volume, using primers for
analyses of the FASN and β-actin genes. The conditions for PCR were
as follows: Initial denaturation at 95°C for 5 min and followed by
45 cycles (95°C for 15 sec, 55°C for 15 sec, 72°C for 20 sec).
(FASN sense, 5′-TATGCTTCTTCGTGCAGCAGTT-3′ and antisense,
5′-GCTGCCACACGCTCCTCTAG-3′; β-actin sense,
5′-AAAGACCTGTACGCCAACACAGTGCTGTCTGG-3′ and antisense,
5′-CGTCATACTCCTGCTTGCTGATCCACATCTGC-3′) The β-actin gene, which is
a housekeeping gene, was used as an internal control, and samples
without reverse transcription were used as negative control.
Quantitative PCR was performed in 25 µl final volume containing 2
µl cDNA, SYBR Green Master Mix (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) on a 7500 Real-time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). FASN gene and β-actin
gene expression levels were determined with the comparative
Cq method in triplicate experiments (39).
Intracellular fatty acids assay
SK-BR-3 cells were collected after 24 h treatment
with patuletin at different concentrations (20, 40, 80 µM,
respectively). Then, cells were washed twice with cold PBS and
extracted by homogenization with pure chloroform containing 1%
Triton X-100 (Sigma-Aldrich; Merck KGaA). The extract was
centrifuged at 10,800 × g for 5–10 min at 4°C, to collect the
organic phase. Next, the organic phase was air and vacuum dried to
remove chloroform. The dissolved dried lipids were applied to
detect the amount of intracellular fatty acid by Fatty Acid Assay
kit (BioVision, Inc., Milpitas, CA, USA), following the
manufacturer's protocol. The fatty acids concentration was measured
at 570 nm by a microplate spectrophotometer.
Cell FASN activity assay
Intracellular FASN activity was assessed as
described previously (40). SK-BR-3
cells were harvested and collected in cold assay buffer containing
100 mM potassium phosphate buffer, 1 mM EDTA, 0.6 mM PMSF and 1 mM
dithiolthreitol (pH 7.0). Then, the cell suspension was centrifuged
at 10,800 × g for 30 min at 4°C, and the supernatant was collected
for the overall reaction assay. A total of 25 ml supernatant was
added into the reaction mix containing 25 mM
KH2PO4-K2HPO4 buffer,
0.25 mM EDTA, 0.25 mM dithiothreitol, 30 mM acetyl-CoA, 100 mM
malonyl-CoA, 350 mM NADPH (pH 7.0) to a total volume of 200 ml. The
protein content in the supernatant was determined using a
bicinchoninic acid (BCA) assay (Pierce; Thermo Fisher Scientific,
Inc.) and results were expressed as the specific activity of FASN
at the same protein concentration.
Quantification of fatty acid
Following treatment with patuletin at the
corresponding concentrations (0, 20, 40 and 80 µM), cells were
harvested using trypsin-EDTA, washed twice with PBS, and stored at
−80°C. The amount of intracellular fatty acid was determined with a
Free Fatty Acid Assay kit (Sigma-Aldrich; Merck KGaA), according to
the manufacturer's protocol.
Statistical analysis
All values are presented as mean ± standard
deviation. To determine if differences between experimental and
control groups existed in cancer cell viability, apoptosis, FASN
gene expression and activity and in intracellular fatty acids
concentration, the results were evaluated and analyzed by one-way
analysis of variance and the Dunnett's post-hoc test using GraphPad
software (version 5.0; GraphPad Software, Inc., La Jolla, CA, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Inhibitory effects of patuletin on the
viability of SK-BR-3 cells
To identify whether patuletin affected the viability
of the breast cancer SK-BR-3 cell line, cells were treated with
0–160 µM patuletin for 24 h, and following this, the ability of
cell survival was examined by MTT assay. As demonstrated in
Fig. 1B, SK-BR-3 cell viability was
reduced significantly subsequent to treatment with 20, 40, 80 and
160 µM patuletin. When compared with the negative control (0 µM
patuletin), cell survival rate was markedly reduced to 13%
following treatment with 160 µM patuletin. Patuletin demonstrated
high inhibition of cell population in a dose-dependent manner, with
a half-maximal inhibitory concentration (IC50) value of
24 µM.
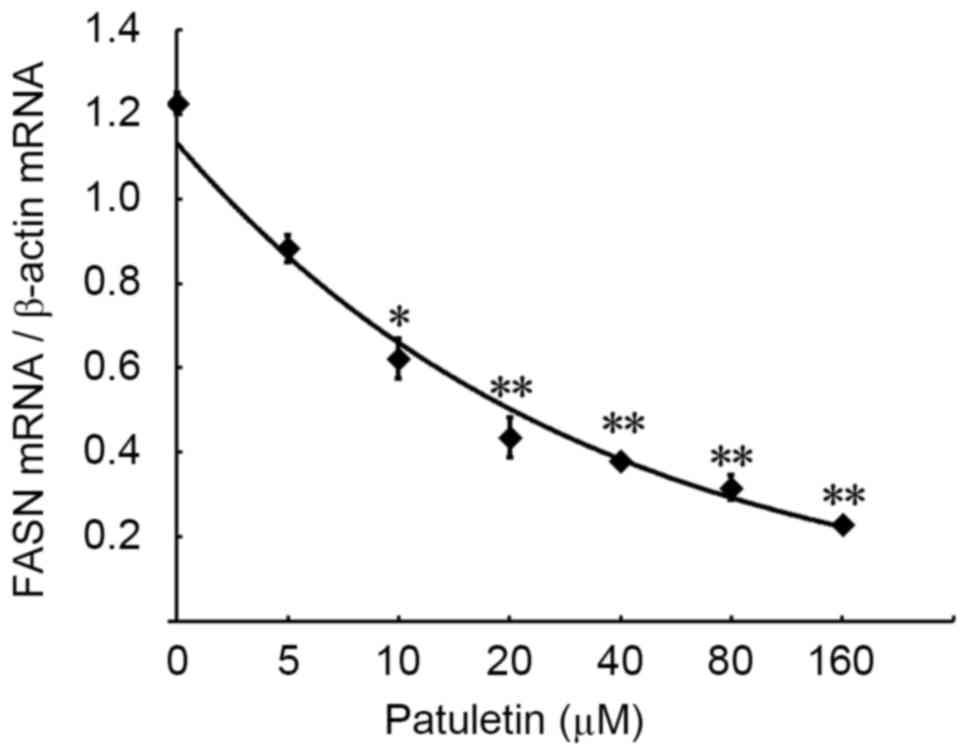
Patuletin reduces gene expression of
FASN in SK-BR-3 cells
The effect of patuletin on FASN gene expression in
SK-BR-3 cells was measured by PCR and reverse transcription-PCR. As
demonstrated in Fig. 2, treatment of
SK-BR-3 cells for 24 h with increasing concentrations of patuletin
(from 5–160 µM) resulted in a significant reduction in FASN mRNA
expression.
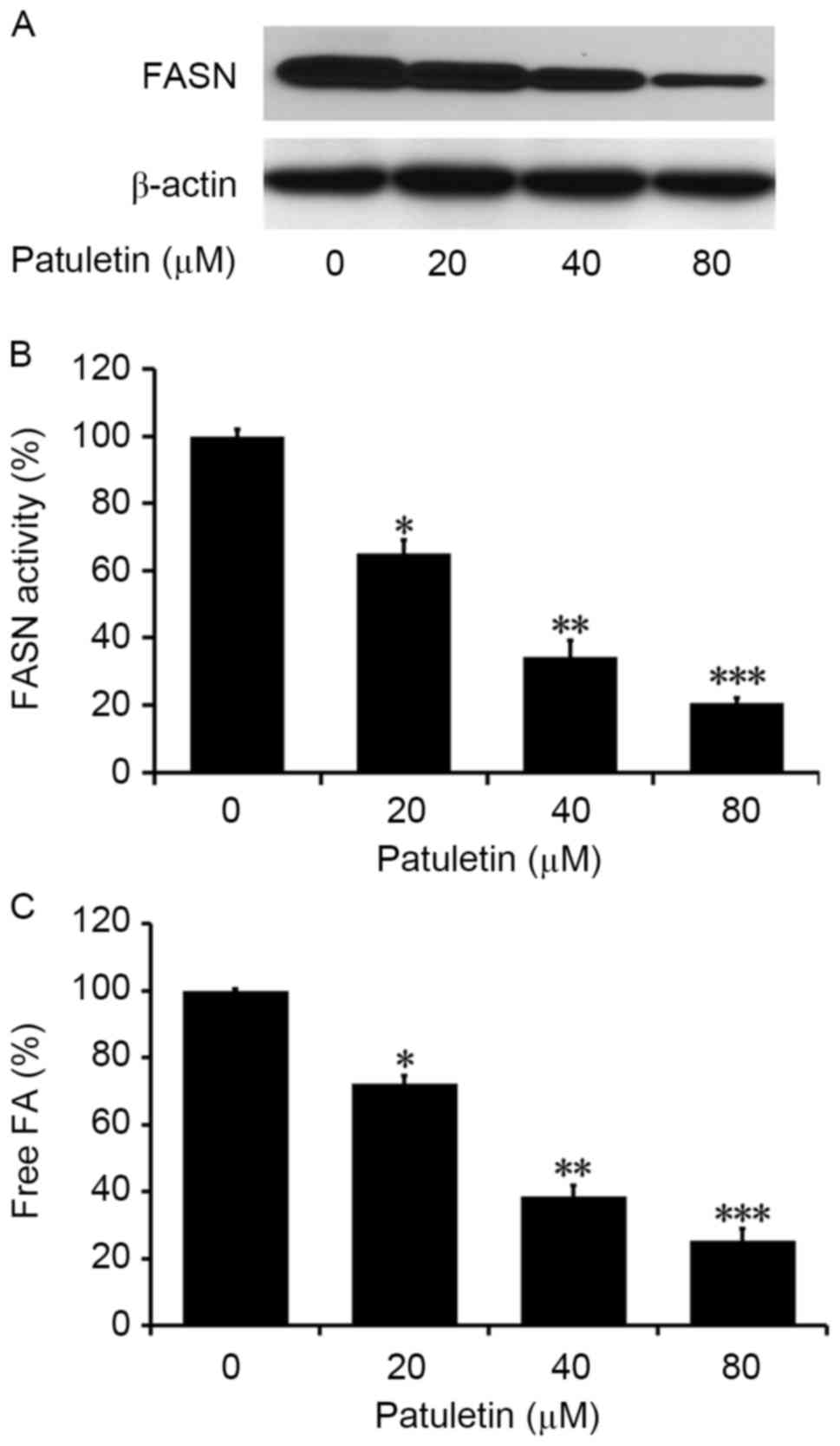
Patuletin inhibits intracellular FASN
activity in SK-BR-3 cells
The effect of patuletin on the FASN activity in
SK-BR-3 cells was measured by western blotting analysis. As
demonstrated in Fig. 3A, SK-BR-3
cells treated with patuletin for 24 h exhibited much lower levels
of FASN compared with the control. When SK-BR-3 cells were treated
with patuletin at different concentrations for 24 h, intracellular
FASN activity was significantly reduced to 65.0, 34.3 and 20.6%,
respectively (P<0.05, P<0.01 and P<0.001, respectively;
Fig. 3B). This suggests that
intracellular FASN activity was significantly suppressed by
patuletin, and that the inhibition was dose-dependent.
Patuletin reduced intracellular fatty
acids in SK-BR-3 cells
The levels of intracellular fatty acids in SK-BR-3
cells treated with 20, 40 and 80 µM patuletin was measured by Fatty
Acids Assay kit. As demonstrated in Fig.
3C, compared with the control (0 µM patuletin), the levels of
intracellular fatty acids in treated cells significantly decreased
to 63.8, 52.4 and 31.2% (P<0.05, P<0.01 and P<0.001,
respectively).
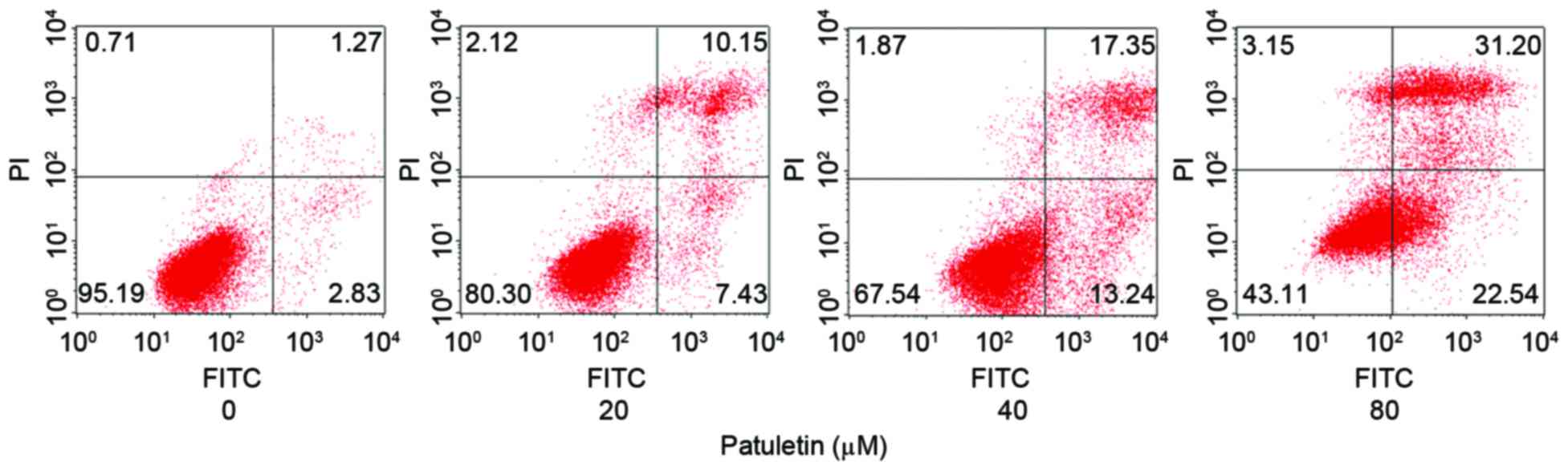
Patuletin induced SK-BR-3 cells
apoptosis
The apoptotic rate of SK-BR-3 cells following 20, 40
and 80 µM patuletin treatment for 24 h was measured using a Annexin
V-FITC Apoptosis Detection kit and analyzed by flow cytometry. As
demonstrated in Fig. 4, patuletin
markedly induced SK-BR-3 cell apoptosis in a dose-dependent
manner.
Discussion
The present study focuses on the effects of
patuletin on FASN gene expression and activity in human breast
cancer SK-BR-3 cells. To the best of our knowledge, it was
demonstrated for the first time that patuletin dose-dependently
decreases the gene and protein expression levels of FASN and its
activity in the human breast cancer SK-BR-3 cell line. In addition,
this natural flavone markedly inhibits cell proliferation and
induces apoptosis in SK-BR-3 cells.
Natural polyphenolic compounds include of a wide
variety of biologically active compounds, a number of which have
been suggested to exhibit antineoplastic properties (40–42).
However, the anti-cancer activity of patuletin has not yet been
examined, to the best of our knowledge. In the present study, the
inhibition of FASN activity was associated with the apoptosis of
cancer cells, which suggested that efficient FASN inhibitors may be
potential target drugs for the treatment of cancer. It was also
demonstrated that the natural polyphenolic compound patuletin may
inhibit intracellular FASN activity, and therefore induce breast
cancer cell apoptosis.
FASN is a key multi-enzyme that catalyzes fatty acid
synthesis. The expression level of FASN is relatively low in the
majority of normal tissues, however; increased expression of FASN
has been identified in human breast cancer cells, particularly in
SK-BR-3 cells (37). According to
previous studies on tumor proliferation, FASN may contribute to the
generation of tumor cell membranes (43). Therefore, FASN inhibitors such as C75
and orlistat are promising potential anti-cancer drugs for the
prevention and/or treatment of a variety of cancers such as
cervical, prostate, leukemia and colon cancer (44–46). It is
essential to identify more effective FASN inhibitors that may be
applied practically as chemotherapeutic drugs.
The present study identified that patuletin not only
downregulated mRNA and protein expression of FASN, but also
demonstrated a high inhibitory activity on intracellular FASN. The
decrease of intracellular FASN activity and fatty acids levels in
SK-BR-3 cells revealed that patuletin acted on FASN as an
inhibitory target. The intracellular activity of FASN directly
affected the amount of intracellular fatty acids as FASN serves a
key role in de novo fatty acid biosynthesis.
Like certain FASN inhibitors such as C75 and
cerulenin (18), patuletin has been
suggested to induce apoptosis of breast cancer cells. The results
demonstrated that the apoptotic ratio of patuletin treated SK-BR-3
cells increased from 4.10% (control) to 53.74% (80 µM patuletin).
The mechanism of cancer cell apoptosis through the inhibition of
intracellular FASN expression may be explained by accumulating
malonyl-CoA, which was considered as a trigger of cancer cell death
and apoptosis (47,48). The present study also concluded that
the signal pathways in cancer cell apoptosis exhibit close
associations with the inhibition of FASN, therefore FASN inhibitors
may be ideal drugs for the treatment of cancer.
In conclusion, patuletin induced apoptosis in breast
cancer SK-BR-3 cells via inhibiting intracellular FASN activity and
downregulating the mRNA and protein expression levels of FASN. As
patuletin demonstrated a significant promotion of apoptosis in
SK-BR-3 cells, it exhibits potential for application as an
anti-cancer drug candidate for the treatment of human breast
cancers.
Acknowledgements
The present study was sponsored by Natural Science
Foundation of Heilongjiang Province (grant no. C201439),
Heilongjiang Postdoctoral Fund (grant no. LBH-Z13142) and China
Postdoctoral Science Foundation (grant no. 2014M551267).
References
|
1
|
Wakil SJ: Fatty-acid synthase, a
proficient multifunctional enzyme. Biochemistry. 28:4523–4530.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Milgraum LZ, Witters LA, Pasternack GR and
Kuhajda FP: Enzymes of the fatty acid synthesis pathway are highly
expressed in in situ breast carcinoma. Clin Cancer Res.
3:2115–2120. 1997.PubMed/NCBI
|
|
3
|
Aggarwal BB and Shishodia S: Molecular
targets of dietary agents for prevention and therapy of cancer.
Biochem Pharmacol. 71:1397–1421. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Alo' PL, Visca P, Marci A, Mangoni A,
Botti C and Di Tondo U: Expression of fatty acid synthase (FAS) as
a predictor of recurrence in stage I breast carcinoma patients.
Cancer. 77:474–482. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Swinnen JV, Roskams T, Joniau S, Van
Poppel H, Oyen R, Baert L, Heyns W and Verhoeven G: Overexpression
of fatty acid synthase is an early and common event in the
development of prostate cancer. Int J Cancer. 98:19–22. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pizer ES, Lax SF, Kuhajda FP, Pasternack
GR and Kurman RJ: Fatty acid synthase expression in endometrial
carcinoma: Correlation with cell proliferation and hormone
receptors. Cancer. 83:528–537. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gansler TS, Hardman W III, Hunt DA,
Schaffel S and Hennigar RA: Increased expression of fatty acid
synthase (OA-519) in ovarian neoplasms predicts shorter survival.
Hum Pathol. 28:686–692. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rashid A, Pizer ES, Moga M, Milgraum LZ,
Zahurak M, Pasternack GR, Kuhajda FP and Hamilton SR: Elevated
expression of fatty acid synthase and fatty acid synthetic activity
in colorectal neoplasia. Am J Pathol. 150:201–208. 1997.PubMed/NCBI
|
|
9
|
Orita H, Coulter J, Tully E, Kuhajda FP
and Gabrielson E: Inhibiting fatty acid synthase for
chemoprevention of chemically induced lung tumors. Clin Cancer Res.
14:2458–2464. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Nie F, Ouyang J, Wang X and Ma X:
Inhibitory effects of sea buckthorn procyanidins on fatty acid
synthase and MDA-MB-231 cells. Tumor Biol. 35:9563–9569. 2014.
View Article : Google Scholar
|
|
11
|
Alo PL, Amini M, Piro F, Pizzuti L,
Sebastiani V, Botti C, Murari R, Zotti G and Di Tondo U:
Immunohistochemical expression and prognostic significance of fatty
acid synthase in pancreatic carcinoma. Anticancer Res.
27:2523–2527. 2007.PubMed/NCBI
|
|
12
|
Fan H, Tian W and Ma X: Curcumin induces
apoptosis of HepG2 cells via inhibiting fatty acid synthase. Target
Oncol. 9:279–286. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Y, Tian WX and Ma XF: Inhibitory
effects of onion (Allium cepa L.) extract on proliferation
of cancer cells and adipocytes via inhibiting fatty acid synthase.
Asian Pac J Cancer Prev. 13:5573–5579. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Walter K, Hong SM, Nyhan S, Canto M,
Fedarko N, Klein A, Griffith M, Omura N, Medghalchi S, Kuhajda F
and Goggins M: Serum fatty acid synthase as a marker of pancreatic
neoplasia. Cancer Epidemiol Biomarkers Prev. 18:2380–2385. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ginsburg OM and Love RR: Breast cancer: A
neglected disease for the majority of affected women worldwide.
Breast J. 17:289–295. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Swinnen JV, Heemers H, Deboel L, Foufelle
F, Heyns W and Verhoeven G: Stimulation of tumor-associated fatty
acid synthase expression by growth factor activation of the sterol
regulatory element-binding protein pathway. Oncogene. 19:5173–5181.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Prieto-Hontoria PL, Pérez-Matute P,
Fernández-Galilea M, Bustos M, Martínez JA and Moreno-Aliaga MJ:
Role of obesity-associated dysfunctional adipose tissue in cancer:
A molecular nutrition approach. Biochim Biophys Acta. 1807:664–678.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kuhajda FP: Fatty acid synthase and
cancer: New application of an old pathway. Cancer Res.
66:5977–5980. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li P, Tian W, Wang X and Ma X: Inhibitory
effect of desoxyrhaponticin and rhaponticin, two natural stilbene
glycosides from the Tibetan nutritional food Rheum
tanguticum Maxim. ex Balf., on fatty acid synthase and human
breast cancer cells. Food Funct. 5:251–256. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li P, Tian W and Ma X: Alpha-mangostin
inhibits intracellular fatty acid synthase and induces apoptosis in
breast cancer cells. Mol Cancer. 13:1382014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fan H, Wu D, Tian W and Ma X: Inhibitory
effects of tannic acid on fatty acid synthase and 3T3-L1
preadipocyte. Biochim Biophys Acta. 1831:1260–1266. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu D, Ma X and Tian W: Pomegranate husk
extract, punicalagin and ellagic acid inhibit fatty acid synthase
and adipogenesis of 3T3-L1 adipocyte. J Funct Food. 5:633–641.
2013. View Article : Google Scholar
|
|
23
|
Quan X, Wang Y, Ma X, Liang Y, Tian W, Ma
Q, Jiang H and Zhao Y: α-Mangostin induces apoptosis and suppresses
differentiation of 3T3-L1 cells via inhibiting fatty acid synthase.
PLoS One. 7:e333762012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jiang HZ, Ma QY, Fan HJ, Liang WJ, Huang
SZ, Dai HF, Wang PC, Ma XF and Zhao YX: Fatty acid synthase
inhibitors isolated from Punica granatum L. J Braz Chem Soc.
23:889–893. 2012. View Article : Google Scholar
|
|
25
|
Jiang HZ, Quan XF, Tian WX, Hu JM, Wang
PC, Huang SZ, Cheng ZQ, Liang WJ, Zhou J, Ma XF and Zhao YX: Fatty
acid synthase inhibitors of phenolic constituents isolated from
Garcinia mangostana. Bioorg Med Chem Lett. 20:6045–6047.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liang Y, Tian W and Ma X: Inhibitory
effects of grape skin extract and resveratrol on fatty acid
synthase. BMC Complement Altern Med. 13:3612013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nie F, Liang Y, Xun H, Sun J, He F and Ma
X: Inhibitory effects of tannic acid in the early stage of 3T3-L1
preadipocytes differentiation by down-regulating PPARγ expression.
Food Funct. 6:894–901. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiang B, Liang Y, Sun X, Liu X, Tian W and
Ma X: Potent inhibitory effect of chinese dietary spices on fatty
acid synthase. Plant Foods Hum Nutr. 70:257–262. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zeng XF, Li WW, Fan HJ, Wang XY, Pan J,
Wang ZR, Ma S, Li LL, Ma XF and Yang SY: Discovery of novel fatty
acid synthase (FAS) inhibitors based on the structure of ketoaceyl
synthase (KS) domain. Bioorg Med Chem Lett. 21:4742–4744. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao YX, Liang WJ, Fan HJ, Ma QY, Tian WX,
Dai HF, Jiang HZ, Li N and Ma XF: Fatty acid synthase inhibitors
from the hulls of Nephelium lappaceum L. Carbohydrate Res.
346:1302–1306. 2011. View Article : Google Scholar
|
|
31
|
Tian WX: Inhibition of fatty acid synthase
by polyphenols. Curr Med Chem. 13:967–977. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yasukawa K and Kasahara Y: Effects of
flavonoids from French marigold (Florets of Tagetes patula
L.) on acute inflammation model. Int J Inflam. 2013:3094932013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Koleckar V, Brojerova E, Rehakova Z,
Kubikova K, Cervenka F, Kuca K, Jun D, Hronek M, Opletalova V and
Opletal L: In vitro antiplatelet activity of flavonoids from
Leuzea carthamoides. Drug Chem Toxicol. 31:27–35. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fang JJ, Ye G, Chen WL and Zhao WM:
Antibacterial phenolic components from Eriocaulon
buergerianum. Phytochemistry. 69:1279–1286. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Könczöl A, Engel R, Szabó K, Hornok K,
Tóth S, Béni Z, Prechl A, Máthé I and Balogh Tibor G: Topical
analgesic, anti-inflammatory and antioxidant properties of
Oxybaphus nyctagineus: Phytochemical characterization of
active fractions. J Ethnopharmacol. 155:776–784. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li S, Mao W, Cao X, Liang S, Ding Z and Li
N: Inhibition of rat lens aldose reductase by quercetagetin and
patuletin. Yan Ke Xue Bao. 7(29–30): 331991.
|
|
37
|
Yoon S, Lee MY, Park SW, Moon JS, Koh YK,
Ahn YH, Park BW and Kim KS: Up-regulation of acetyl-CoA carboxylase
alpha and fatty acid synthase by human epidermal growth factor
receptor 2 at the translational level in breast cancer cells. J
Biol Chem. 282:26122–26131. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Uddin S, Ah-Kang J, Ulaszek J, Mahmud D
and Wickrema A: Differentiation stage-specific activation of p38
mitogen-activated protein kinase isoforms in primary human
erythroid cells. P Natl Acad Sci USA. 101:147–152. 2004. View Article : Google Scholar
|
|
39
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fan H, Liang Y, Jiang B, Li X, Xun H, Sun
J, He W, Lau HT and Ma X: Curcumin inhibits intracellular fatty
acid synthase and induces apoptosis in human breast cancer
MDA-MB-231 cells. Oncol Rep. 35:2651–2656. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Adaramoye O, Erguen B, Nitzsche B, Höpfner
M, Jung K and Rabien A: Punicalagin, a polyphenol from pomegranate
fruit, induces growth inhibition and apoptosis in human PC-3 and
LNCaP cells. Chem Biol Interact. 274:100–106. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Szeja W, Grynkiewicz G and Rusin A:
Isoflavones, their glycosides and glycoconjugates. synthesis and
biological activity. Curr Org Chem. 21:218–235. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Menendez JA, Mehmi I, Atlas E, Colomer R
and Lupu R: Novel signaling molecules implicated in
tumor-associated fatty acid synthase-dependent breast cancer cell
proliferation and survival: Role of exogenous dietary fatty acids,
p53-p21WAF1/CIP1, ERK1/2 MAPK, p27KIP1,
BRCA1, and NF-κB. Int J Oncol. 24:591–608. 2004.PubMed/NCBI
|
|
44
|
Menendez JA and Lupu R: Fatty acid
synthase and the lipogenic phenotype in cancer pathogenesis. Nat
Rev Cancer. 7:763–777. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cioccoloni G, Bonmassar L, Pagani E,
Caporali S, Fuggetta MP, Bonmassar E, D'Atri S and Aquino A:
Influence of fatty acid synthase inhibitor orlistat on the DNA
repair enzyme O6-methylguanine-DNA methyltransferase in human
normal or malignant cells in vitro. Int J Oncol. 47:764–772. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Rae C, Haberkorn U, Babich JW and Mairs
RJ: Inhibition of fatty acid synthase sensitizes prostate cancer
cells to radiotherapy. Radiat Res. 184:482–493. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Pizer ES, Thupari J, Han WF, Pinn ML,
Chrest FJ, Frehywot GL, Townsend CA and Kuhajda FP:
Malonyl-coenzyme-A is a potential mediator of cytotoxicity induced
by fatty-acid synthase inhibition in human breast cancer cells and
xenografts. Cancer Res. 60:213–218. 2000.PubMed/NCBI
|
|
48
|
Zhou W, Simpson PJ, McFadden JM, Townsend
CA, Medghalchi SM, Vadlamudi A, Pinn ML, Ronnett GV and Kuhajda FP:
Fatty acid synthase inhibition triggers apoptosis during S phase in
human cancer cells. Cancer Res. 63:7330–7337. 2003.PubMed/NCBI
|