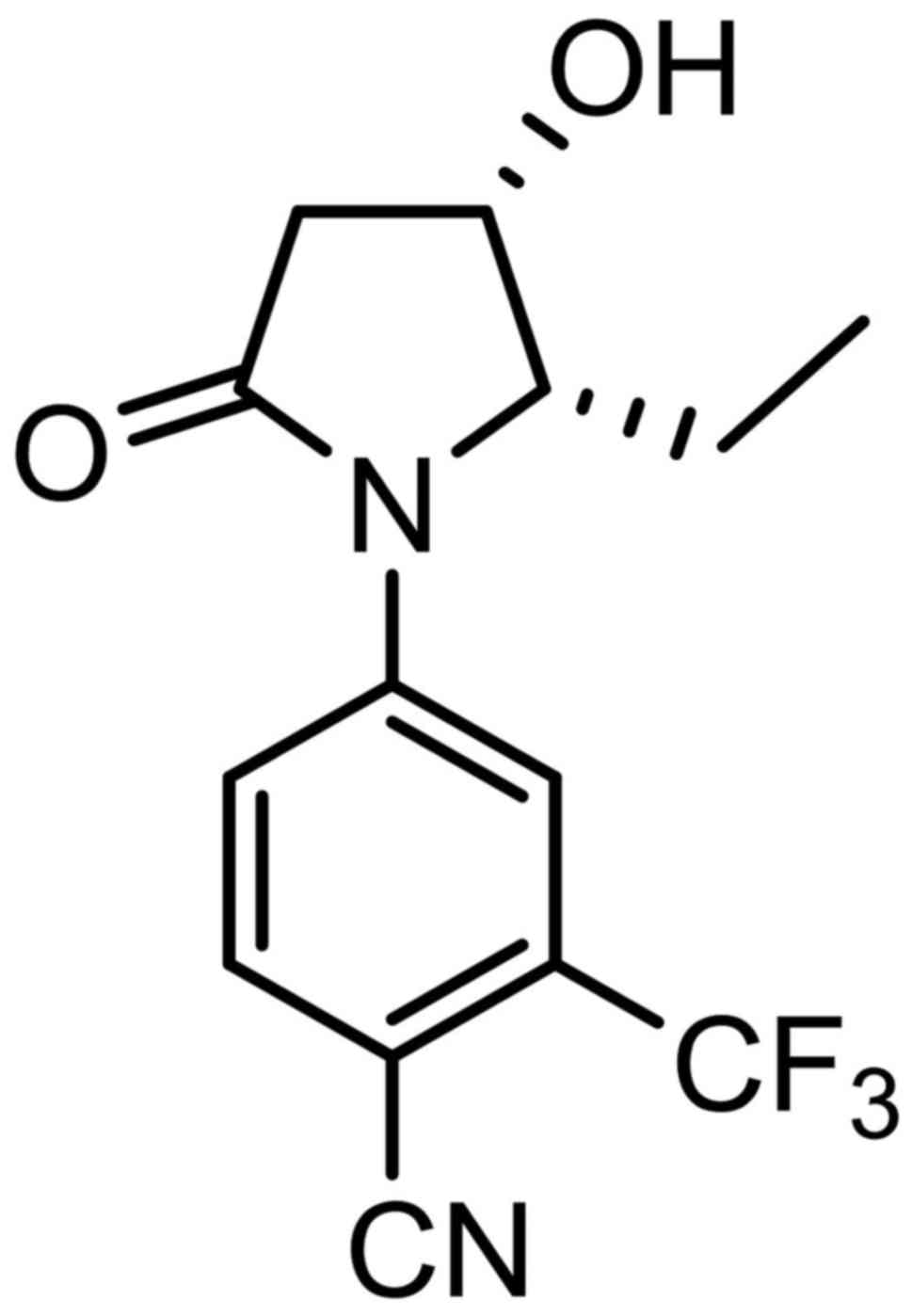
Introduction
Cancer cachexia is defined as a multifactorial
disorder associated with an ongoing loss of skeletal muscle mass
with or without loss of fat mass, which leads to a progressive
functional impairment that cannot be fully reversed with
conventional nutritional supplementation (1). The symptoms appear in up to 80% of
patients with cancer and account for at least 20% of
cancer-associated deaths (1,2). Cancer cachexia has been reported to be
caused by nutritional deficiencies, metabolic disorders, and
inflammatory disorders (3–6). Presently, however, there is no approved
therapy for the treatment or prevention of cancer cachexia.
Therefore, an effective therapy for cancer cachexia is an unmet
medical need. There are several potential therapeutic approaches
for cancer cachexia (7–10). Androgen is a steroid hormone with
multiple physiological functions including promotion of growth
hormone release, appetite stimulation, anabolic actions,
stimulatory effects on the central nervous system (CNS), and
regulation of energy homeostasis. Low plasma androgen levels are
often observed in patients with terminal cancer (11,12).
Therefore, androgen-based treatments might be a potential therapy
for cancer cachexia. However, testosterone replacement therapy has
unavoidable side effects such as myocardial infarction, heart
failure, stroke, depression, and aggression (13,14).
In a previous study, we reported the
4-(pyrrolidin-1-yl) benzonitrile derivative as a selective androgen
receptor modulator (SARM)-1c (15).
Unlike testosterone, SARM-1c exhibited anabolic effects on the
levator ani muscles without excessively increasing prostate weight
in rat Hershberger assays (15). We
further improved the pharmacokinetic (PK) profile by modifying the
structure and identified 4-[(2S,S)
−2-ethyl-3-hydroxy-5-oxopyrrolidin-1-yl]-2-(trifluoromethyl)
benzonitrile (SARM-2f) as a novel SARM (Fig. 1). In the present study, we examined
the effects of SARM-2f on the weight of skeletal muscles (i.e., the
gastrocnemius and soleus muscles), body weight, body composition,
and anorexia in a castrated rat model and three cancer cachexia
models.
Materials and methods
Ethical statement
The animal protocol for this study was approved by
the Animal Ethics Committee of Takeda Pharmaceutical Company Ltd.
In addition, all procedures were performed according to protocols
approved by the Institutional Animal Care and Use Committee of the
Pharmaceutical Research Division of Takeda Pharmaceutical Company
Limited. The protocols were in accordance with the ethical
standards laid down in the 1964 Declaration of Helsinki and its
later amendments.
Animals and compounds
CD (Sprague-Dawley, SD, IGS) rats were purchased
from Charles River Laboratories Japan (Yokohama, Japan). F344
immunodeficient rats (5- or 15-week-old) and BALB/c mice (5- or
6-weeks-old) were obtained from CLEA Japan, Inc. (Tokyo Japan). The
animals were maintained under a 12-h light/dark cycle at a constant
temperature of 23±2°C while food and water were provided ad
libitum. SARM-2f (4-((2S,
3S)-2-ethyl-3-hydroxy-5-oxopyrrolidin-1-yl)-2-(trifluoromethyl)benzonitrile)
was synthesized by Takeda Pharmaceutical Company Ltd., and
suspended in 0.5% methylcellulose. Tumor necrosis factor (TNF)-α
(WAKO Pure Chemical Industries, Ltd., Osaka, Japan) was dissolved
in saline (Otsuka, Tokushima, Japan), whereas testosterone
propionate (TP; Tokyo Chemical Industry, Tokyo, Japan) was
dissolved in 20% benzyl benzoate and 80% corn oil (BBoil).
Castrated rat model
Mature 12-week-old male SD (ID) IGS rats were
castrated under isoflurane anesthesia. Then, 8 weeks later, they
were divided into six groups (n=10) based on body weight (day 0).
The animals were then administered the vehicle (0.5%
methylcellulose, orally, p.o.; once daily, QD) and BBoil
(subcutaneously, s.c., thrice weekly), SARM-2f (0.3, 3, or 30
mg/kg, p.o., QD) and BBoil (s.c., thrice weekly), or TP (2.5 mg/kg,
s.c., thrice weekly) and 0.5% methylcellulose (p.o., QD) for 4
weeks. Body weight was measured twice weekly, whereas the body
composition was determined using nuclear magnetic resonance
spectroscopy (EchoMRI-700, Echo Medical Systems, Houston, TX, USA)
on day 0 and 27. On day 28, the skeletal muscles and sex accessory
organs were removed and weighed. The organs were immersed in saline
until weighed to prevent them from drying out.
TNFα-induced cachexia mouse model
Seven-week-old BALB/c male mice were castrated under
anesthesia (day 0). Intraperitoneal (i.p.) injections of TNFα (5
µg/day twice daily, BID) were administered to the mice starting
from day 7 and continued for 2 weeks. The animals were
simultaneously treated with SARM-2f (10 or 30 mg/kg, p.o., BID) and
BBoil (s.c., QD) or TP (30 mg/kg, s.c., QD) and 0.5%
methylcellulose (p.o., BID). The body weight and amount of food
consumed were measured twice weekly. In addition, carcass weight
(i.e., the weight of the mouse after removal of all internal
organs) and wet organ weight were measured at the end of the
study.
G361 cancer cachexia model
Fifteen-week-old F344 immunodeficient rats were used
for this study. Under isoflurane anesthesia, 5×106 G361
human melanoma cells (American Tissue Culture Collection, ATCC,
Manassas, VA, USA) were implanted subcutaneously into one flank of
each rat (day 0). The rats were then administered the vehicle [0.5%
methylcellulose (p.o., QD)] and BBoil (s.c., thrice weekly),
SARM-2f (3 mg/kg, p.o., QD) and BBoil (s.c., thrice weekly), or TP
(2.5 mg/kg, s.c., thrice weekly) and 0.5% methylcellulose (p.o.,
QD) for 4 weeks starting from day 7. The tumor size and body weight
were measured twice a week. The tumor volume (expressed in
mm3) was calculated using the formula 0.5 × a ×
b2, where ‘a’ is the largest diameter, and ‘b’ is
largest diameter perpendicular to ‘a.’ Lean body mass was measured
using an EchoMRI-700 on days 7 and 35. The muscles were removed and
measured at the end of the study.
C26 cancer cachexia model
Six-week-old male BALB/c mice were used for this
model. Briefly, 5×105 C26 mouse carcinoma cells were
subcutaneously implanted into one flank of each mouse on the day of
castration (day 0). The mice were then administered the vehicle
[0.5% methylcellulose (p.o., BID)] and BBoil (s.c., QD), SARM-2f
(10, 30, or 100 mg/kg, p.o., BID) and BBoil (s.c., QD), or TP (30
mg/kg, s.c., QD) and 0.5% methylcellulose (p.o., BID) for 2 weeks
starting from day 11. The tumor size and body weight were measured
twice weekly. The tumor volume was expressed as described above.
Carcass, muscle, and organ weights were measured at the end of the
study.
Statistical analysis
Differences between the control and treatment groups
were analyzed using the Student's t-test, Williams' test, or
Dunnett's test.
Results
Effects of SARM-2f on body weight and
skeletal muscles of castrated rats
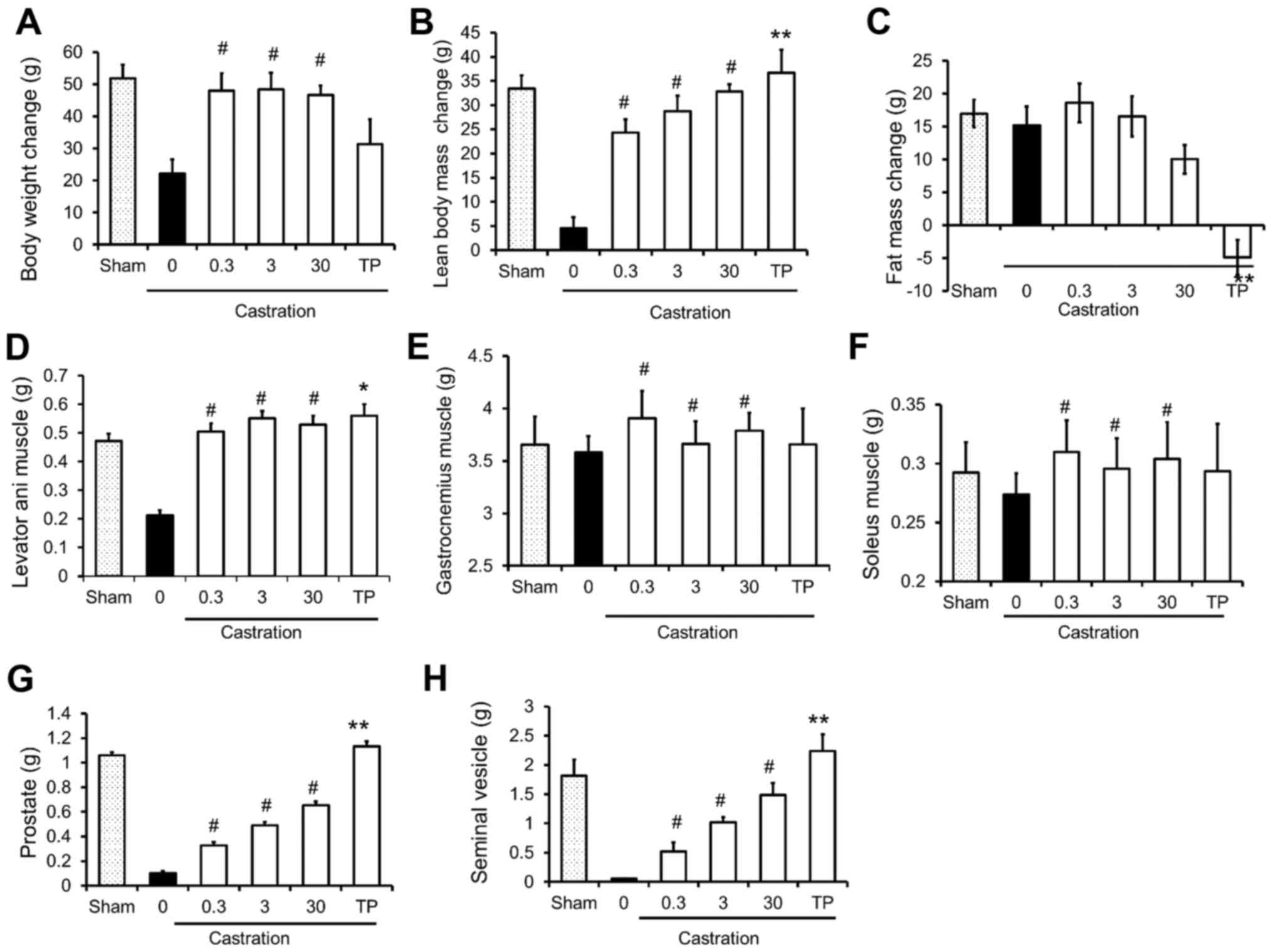
The effects of SARM-2f on castrated rats are
presented in Fig. 2. Castration
decreased the body weight, lean body mass, and the weight of the
levator ani muscle, prostate, and seminal vesicle (Fig. 2A, B, D, G, and H); however, it did not
influence fat mass or the weight of the gastrocnemius or soleus
muscles (Fig. 2C-F). Treatment with
SARM-2f at doses of 0.3, 3, and 30 mg/kg significantly increased
the body weight gain (2.2-fold, 2.2-fold, and 2.1-fold,
respectively), lean body mass (5.4-fold, 6.3-fold, and 7.2-fold,
respectively), and weight of the levator ani (2.3-fold, 2.6-fold,
and 2.6-fold, respectively), gastrocnemius (1.1-fold, 1.02-fold,
and 1.06-fold, respectively), and soleus (1.1-fold, 1.1-fold, and
1.1-fold, respectively) muscles of the castrated rats (Fig. 2A, B, and D-F). However, SARM-2f
treatment did not influence the fat mass (Fig. 2C). Treatment with TP at a dose of 2.5
mg/kg increased the levator ani muscle weight (2.6-fold) but not
the body weight (Fig. 2A and D). The
TP treatment significantly reduced the fat mass (Fig. 2C). Treatment with SARM-2f at doses of
0.3, 3, and 30 mg/kg increased the weights of the prostate
(3.2-fold, 4.8-fold, and 6.4-fold, respectively) and seminal
vesicle (9.8-fold, 19-fold, and 28-fold, respectively) in a
dose-dependent manner in the castrated rats (Fig. 2G and H). However, the prostate and
seminal vesicle weights of the SARM-2f-treated castrated rats were
lower than the respective weights of the non-castrated group
(Fig. 2G and H). In contrast, TP
increased the weights of the prostate (11-fold) and seminal vesicle
(42-fold) in the castrated rats, whichwere higher than the
respective weights in the non-castrated group (Fig. 2G and H). These results demonstrate the
anabolic effect and tissue specificity of SARM-2f.
Recovery of food intake rate and body
weight by SARM-2f in TNFα-induced cachexia mouse model
We examined the effects of SARM-2f on
inflammation-related cachexia in the TNFα-induced cachexia mouse
model. TNFα caused body weight loss in castrated mice (Fig. 3A). However, SARM-2f at doses of 10 and
30 mg/kg as well as TP at 30 mg/kg increased the body weight in the
TNFα-treated mice (Fig. 3A). SARM-2f
at doses of 10 and 30 mg/kg increased the carcass weight (1.1-fold
and 1.1-fold, respectively), and food consumption rate (1.1-fold
and 1.1-fold, respectively) in TNFα-treated mice (Fig. 3B and C). TP increased the carcass
weight (1.1-fold) and food consumption rate (1.1-fold) in
TNFα-treated mice (Fig. 3B and
C).
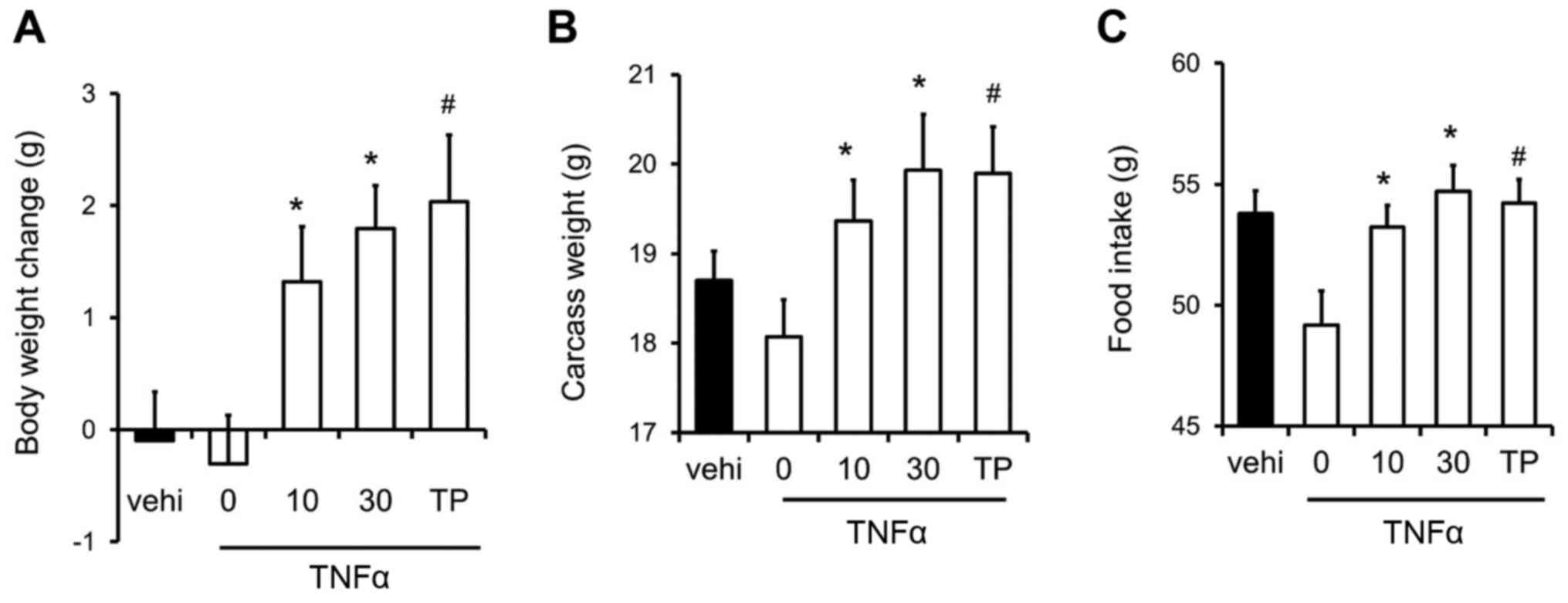 | Figure 3.Recovery of body weight and food
intake in mice with tumor necrosis factor (TNF)-α-induced cachexia.
(A) Body weight change, (B) carcass weight, and (C) food intake in
mice treated with vehicle, selective androgen receptor modulator
(SARM)-2f (10 or 30 mg/kg; oral, p.o.; twice daily BID), or
testosterone propionate (TP, 30 mg/kg, subcutaneous, s.c.; daily,
QD). Body weight change and food intake indicate the change between
day 7 and day 21. Data are mean ± standard error of the mean (SEM,
n=9). #P<0.05 (Dunnett's test) compared to
TNFα-treated castrated mice. *P<0.05 (Student's t-test) compared
to the TNFα-treated castrated mice. |
Prevention of skeletal muscle and body
weight loss by SARM-2f in G361-tumor-bearing rats
The growth of the G361 human melanoma cells that
were subcutaneously grafted into the F344 immunodeficient rats
caused progressive body weight loss and a decrease in lean body
mass in the rats (Fig 4A and B).
Treatment with SARM-2f or TP did not change the body weight and
lean body mass (Fig. 4A and B).
However, both SARM-2f and TP treatment increased the weight of the
levator ani muscle (1.6-fold and 1.7-fold, respectively) (Fig. 4C). TP but not SARM-2f increased the
prostate and seminal vesicle weights (Fig. 4D and E). The tumor growth was
unaffected by either treatment (Fig.
4F).
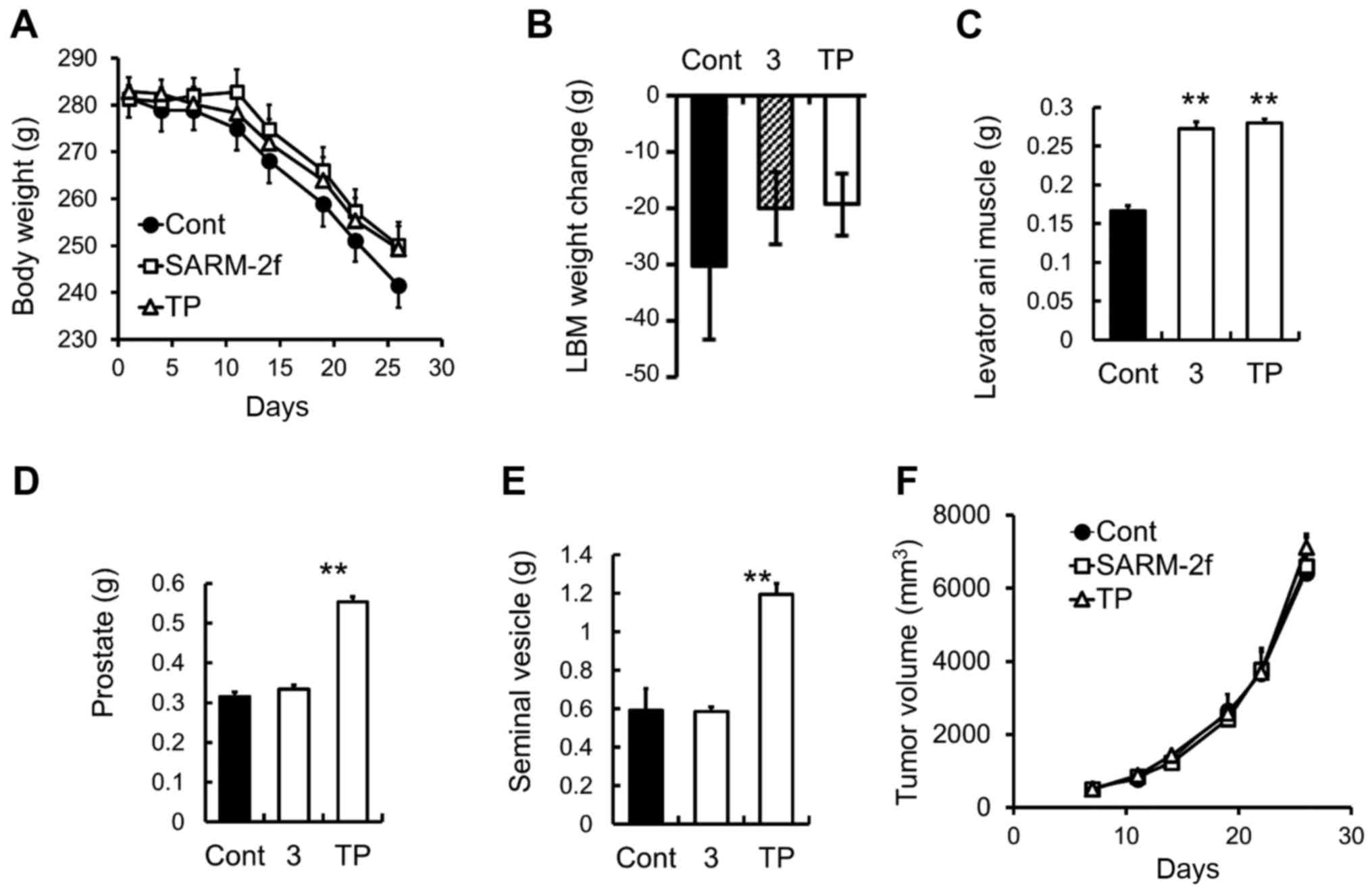 | Figure 4.Body and muscle weight in
G361-tumor-bearing rats (A) Body weight was measured twice weekly.
(B) Lean body mass (LBM) change between day 7 and day 35 by
treatment with vehicle, selective androgen receptor modulator
(SARM)-2f (3 mg/kg; oral, p.o.; daily, QD), or testosterone
propionate (TP, 2.5 mg/kg, subcutaneous, s.c.; thrice weekly).
Weights of (C) levator ani muscle, (D) prostate, and (E) seminal
vesicle were measured at the end of the study. (F) Tumor volume was
measured twice weekly as described in the methods. Data are mean ±
standard error of the mean (SEM, n=9–10). *P<0.05 (Student's
t-test) compared to the vehicle control group. |
Prevention of body weight loss by
SARM-2f in C26-tumor-induced cachexia mouse model
The mouse allograft model of C26 adenocarcinoma is a
well-established model for investigating body weight loss caused by
cancer cachexia (16–18). We found that SARM-2f at doses of 10,
30, and 100 mg/kg increased the body (1.1-fold, 1.1-fold, and
1.1-fold, respectively), carcass (1.1-fold, 1.1-fold, and 1.1-fold,
respectively), and levator ani muscle weights (3.3-fold, 2.9-fold,
and 3.8-fold respectively) in the castrated mice (Fig. 5A-C). No effect was observed on the
ventral prostate weight at a dose of 10 mg/kg (Fig. 5D). Meanwhile, TP increased the body
weight (1.2-fold), carcass weight (1.2-fold), and the weights of
the levator ani muscle (4.6-fold), ventral prostate (3.8-fold), and
seminal vesicle (19-fold) (Fig.
5A-E). Neither SARM-2f nor TP had any effects on tumor growth
in this model (Fig. 5F).
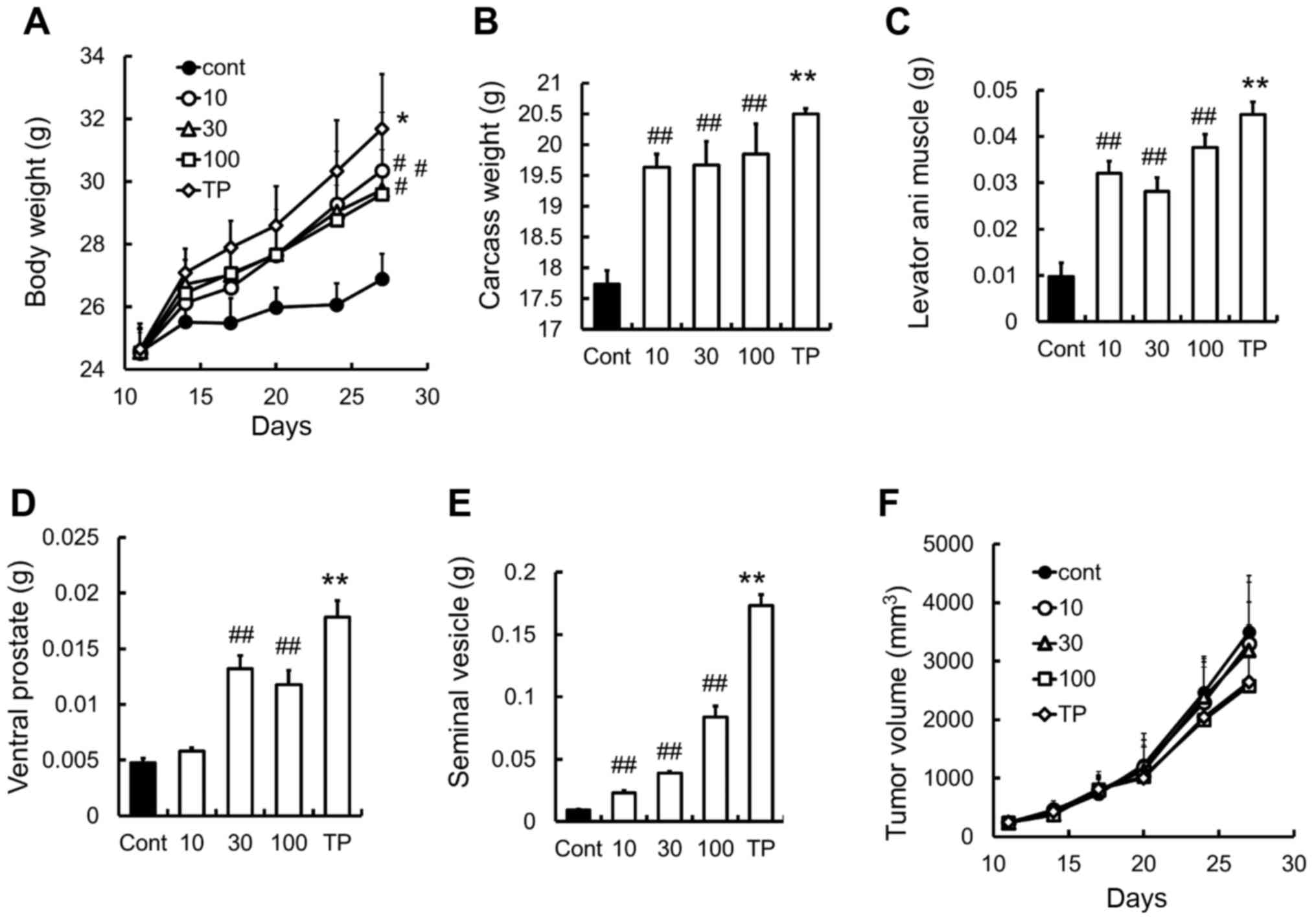 | Figure 5.Prevention of body weight loss in C26
cancer cachexia model (A) Body weight was measured twice weekly.
(B) Carcass weight and the weights of (C) levator ani muscle, (D)
ventral prostate, and (E) seminal vesicle were measured after
treatment with selective androgen receptor modulator (SARM)-2f (10,
30, or 100 mg/kg; oral, p.o.; twice daily, BID) or testosterone
propionate (TP, 30 mg/kg, subcutaneous, s.c.; once daily, QD) on
day 27. (F) Tumor volume was measured twice weekly as described in
methods. Data are mean ± standard error of the mean (SEM, n=7).
#P<0.05 and ##P<0.01 (Dunnett's test)
compared to control group. **P<0.01 (Student's t-test) compared
to control group. |
Discussion
We observed that SARM-2f increased the weights of
the gastrocnemius and soleus muscles in the castrated rats. In
addition, it prevented body weight loss in the TNFα-treated model
and the C26 tumor model, and muscle wasting in the C26 and G361
tumor models. Moreover, it improved anorexia in the TNFα-treated
model. It has been reported that testosterone increases
gastrocnemius muscle weight in rats (19). However, our data showed that a 2.5
mg/kg TP dose increased the weights of the levator ani muscle and
the prostate but had no effects on the weights of the gastrocnemius
and soleus muscles without forced exercise in the castrated male
rat model.
The anabolic effect of testosterone on the skeletal
muscles is controversial and considered dose-dependent (20). Interestingly, SARM-2f increased the
weights of both the gastrocnemius and soleus muscles in our study.
Currently, the difference between the anabolic effects of TP and
those of SARM-2f is unclear. However, the tissue distribution and
tissue-selective gene expression profiles of testosterone may
differ from those of SARM-2f and, therefore, the effects of the two
drugs on balancing the synthesis and breakdown of skeletal muscle
protein may differ.
Several mechanisms have been proposed to explain the
effects of SARM-2f on body weight and body composition in cancer
cachexia models. One possible mechanism is the direct activation of
androgen receptors in the muscles while another is likely mediated
by activation of androgen receptors in other tissues such as CNS,
which may affect behavior, amount of exercise, and appetite
(21). We previously demonstrated
that at a dose of 0.5 mg/kg QD (p.o.), SARM-2f enhanced the sexual
behavior of castrated rats (22). In
the present study, SARM-2f increased the food intake of mice with
TNFα-induced cachexia. These findings suggest that SARM-2f might
have CNS effects that improve cachexia.
The other possible mechanism is the suppression of
inflammation, and inflammatory cytokines such as TNFα, interleukin
(IL)-1, and IL-6 have been reported to be involved in cancer
cachexia (1,5,23,24). Moreover, nandrolone decanoate, an
anabolic steroidal androgen, has been shown to regulate
inflammatory processes in skeletal muscles (2). Therefore, further studies are required
to elucidate the mechanism underlying the effect of SARM-2f on
cancer cachexia.
The causes of cancer cachexia are poorly understood
because of its heterogeneity, which possibly stems from the
heterogeneity and complexity of cancer. Several cancer cachexia
models such as ectopic or orthotopic transplanted xenograft and
allograft models have been reported (17). However, no animal model completely
mimics the complexity of human cancer cachexia (17). Several factors such as diversity of
cancer, use of chemotherapy, exercise, or nutritional support
should be taken into consideration, and suitable experimental
models reflecting the clinical situation should be used for
accurate evaluation of compounds for the treatment of cancer
cachexia.
Despite evidence of the androgen-induced improvement
of muscle wasting and anorexia, androgen replacement therapy is not
a standard treatment for cancer cachexia (25–27).
Testosterone may be a viable therapeutic option for cancer
cachexia. However, clinical trials with large sample sizes are
needed to confirm whether testosterone is an efficacious treatment
for improving body composition, physical function, quality of life,
and cancer survival of patients with cachexia. A double-blind,
placebo-controlled, interventional study investigating this is
currently in progress (NCT00878995). The major concern associated
with testosterone use is the unavoidable side effects such as
cardiovascular disorders (14,15), which
may be partly attributable to estrogen derived from testosterone
(13,28). However, SARM-2f is not a substrate of
aromatase, which is the enzyme that converts testosterone to
estradiol. Therefore, unlike testosterone therapy, treatment with
SARM-2f will not result in the unwanted side effects of
estrogen.
SARMs have been widely investigated in preclinical
animal models of muscle wasting, hypogonadism, osteoporosis, and
other conditions (29–34). Enobosarm is one of the SARMs developed
for treating muscle-wasting diseases. It showed tissue-selectivity
similar to SARM-2f in the Hershberger assay (35). To the best of our knowledge, enobosarm
has not been reported to be effective in cancer cachexia models;
however, it has been reported to increase lean body mass and
improve muscle function in patients with non-small cell lung
carcinoma (NSCLC) in a Phase III clinical trial (36,37).
However, in another clinical trial on NSCLC, enobosarm did not
improve stair climb power in patients with cachexia (36,37).
Therefore, larger clinical trials should be conducted to confirm
the therapeutic potential of enobosarm.
In conclusion, we demonstrated that SARM-2f
increased body and skeletal muscle weight without excessively
stimulating sex accessory organs. It also improved anorexia in rat
and mouse cachexia models, suggesting that it may be an effective
therapeutic option for treating cachexia in patients with
cancer.
Acknowledgements
We would like to thank Jin Kouno and Tsuneo Masaki
for providing technical assistance at Takeda Pharmaceutical Company
Limited. The present study was funded by Takeda Pharmaceutical
Company Ltd.
Glossary
Abbreviations
Abbreviations:
|
SARM
|
selective androgen receptor
modulator
|
|
TP
|
testosterone propionate
|
|
TNFα
|
tumor necrosis factor alpha
|
|
BB
|
benzyl benzoate
|
|
LBM
|
lean body mass
|
References
|
1
|
Fearon K, Strasser F, Anker SD, Bosaeus I,
Bruera E, Fainsinger RL, Jatoi A, Loprinzi C, MacDonald N,
Mantovani G, et al: Definition and classification of cancer
cachexia: An international consensus. Lancet Oncol. 12:489–495.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Argilés JM, Busquets S, Stemmler B and
López-Soriano FJ: Cancer cachexia: Understanding the molecular
basis. Nat Rev Cancer. 14:754–762. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fearon KC, Glass DJ and Guttridge DC:
Cancer cachexia: Mediators, signaling, and metabolic pathways. Cell
Metab. 16:153–166. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cooper C, Burden ST, Cheng H and
Molassiotis A: Understanding and managing cancer-related weight
loss and anorexia: Insights from a systematic review of qualitative
research. J Cachexia Sarcopenia Muscle. 6:99–111. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tuca A, Jimenez-Fonseca P and Gascón P:
Clinical evaluation and optimal management of cancer cachexia. Crit
Rev Oncol Hematol. 88:625–636. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Porporato PE: Understanding cachexia as a
cancer metabolism syndrome. Oncogenesis. 5:e2002016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Coss CC, Bohl CE and Dalton JT: Cancer
cachexia therapy: A key weapon in the fight against cancer. Curr
Opin Clin Nutr Metab Care. 14:268–273. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dodson S, Baracos VE, Jatoi A, Evans WJ,
Cella D, Dalton JT and Steiner MS: Muscle wasting in cancer
cachexia: Clinical implications, diagnosis, and emerging treatment
strategies. Annu Rev Med. 62:265–279. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Madeddu C, Mantovani G, Gramignano G and
Macciò A: Advances in pharmacologic strategies for cancer cachexia.
Expert Opin Pharmacother. 16:2163–2177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Molfino A, Amabile MI, Fanelli Rossi F and
Muscaritoli M: Novel therapeutic options for cachexia and
sarcopenia. Expert Opin Biol Ther. 16:1239–1244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Strasser F, Palmer JL, Schover LR, Yusuf
SW, Pisters K, Vassilopoulou-Sellin R, DeGracia B, Willey JS and
Bruera E: The impact of hypogonadism and autonomic dysfunction on
fatigue, emotional function, and sexual desire in male patients
with advanced cancer: A pilot study. Cancer. 107:2949–2957. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Burney BO, Hayes TG, Smiechowska J,
Cardwell G, Papusha V, Bhargava P, Konda B, Auchus RJ and Garcia
JM: Low testosterone levels and increased inflammatory markers in
patients with cancer and relationship with cachexia. J Clin
Endocrinol Metab. 97:E700–E709. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Basaria S, Coviello AD, Travison TG,
Storer TW, Farwell WR, Jette AM, Eder R, Tennstedt S, Ulloor J,
Zhang A, et al: Adverse events associated with testosterone
administration. N Engl J Med. 363:109–122. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vigen R, O'Donnell CI, Barón AE, Grunwald
GK, Maddox TM, Bradley SM, Barqawi A, Woning G, Wierman ME,
Plomondon ME, et al: Association of testosterone therapy with
mortality, myocardial infarction, and stroke in men with low
testosterone levels. JAMA. 10:1829–1836. 2013. View Article : Google Scholar
|
|
15
|
Aikawa K, Miyawaki T, Hitaka T, Imai YN,
Hara T, Miyazaki J, Yamaoka M, Kusaka M, Kanzaki N, Tasaka A, et
al: Synthesis and biological evaluation of novel selective androgen
receptor modulators (SARMs). Part I. Bioorg Med Chem. 23:2568–2578.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dwarkasing JT, Boekschoten MV, Argilès JM,
van Dijk M, Busquets S, Penna F, Toledo M, Laviano A, Witkamp RF
and van Norren K: Differences in food intake of tumor-bearing
cachectic mice are associated with hypothalamic serotonin
signaling. J Cachexia Sarcopenia Muscle. 6:84–94. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ballarò R, Costelli P and Penna F: Animal
models for cancer cachexia. Curr Opin Support Palliat Care.
10:281–287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Talbert EE, Metzger GA, He WA and
Guttridge DC: Modeling human cancer cachexia in colon 26
tumor-bearing adult mice. J Cachexia Sarcopenia Muscle. 5:321–328.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ustünel I, Akkoyunlu G and Demir R: The
effect of testosterone on gastrocnemius muscle fibres in growing
and adult male and female rats: A histochemical, morphometric and
ultrastructural study. Anat Histol Embryol. 32:70–79. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Urban RJ: Growth hormone and testosterone:
Anabolic effects on muscle. Horm Res Paediatr. 76 Suppl 1:S81–S83.
2011. View Article : Google Scholar
|
|
21
|
De Gendt K and Verhoeven G: Tissue- and
cell-specific functions of the androgen receptor revealed through
conditional knockout models in mice. Mol Cell Endocrinol.
352:13–25. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Aikawa K, Asano M, Ono K, Habuka N, Yano
J, Wilson K, Fujita H, Kandori H, Hara T, Morimoto M, et al:
Synthesis and biological evaluation of novel selective androgen
receptor modulators (SARMs) Part III: Discovery of
4-(5-oxopyrrolidine-1-yl)benzonitrile derivative 2f as a clinical
candidate. Bioorg Med Che. 25:3330–3349.. 2017. View Article : Google Scholar
|
|
23
|
Argiles JM, Lopez-Soriano FJ and Busquets
S: Counteracting inflammation: A promising therapy in cachexia.
Crit Rev Oncog. 17:253–262. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Thompson RW, McClung JM, Baltgalvis KA,
Davis JM and Carson JA: Modulation of overload-induced inflammation
by aging and anabolic steroid administration. Exp Gerontol.
41:1136–1148. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Srinath R and Dobs A: Enobosarm (GTx-024,
S-22): A potential treatment for cachexia. Future Oncol.
10:187–194. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dillon EL, Basra G, Horstman AM, Casperson
SL, Randolph KM, Durham WJ, Urban RJ, Diaz-Arrastia C, Levine L,
Hatch SS, et al: Cancer cachexia and anabolic interventions: A case
report. J Cachexia Sarcopenia Muscle. 3:253–263. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Del Fabbro E, Garcia JM, Dev R, Hui D,
Williams J, Engineer D, Palmer JL, Schover L and Bruera E:
Testosterone replacement for fatigue in hypogonadal ambulatory
males with advanced cancer: A preliminary double-blind
placebo-controlled trial. Support Care Cancer. 21:2599–2607. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
de Vooqt HJ, Smith PH, Pavone-Macaluso M,
de Pauw M and Suciu S: Cardiovascular side effects of
diethylstilbestrol, cyproterone acetate, medroxyprogesterone
acetate and estramustine phosphate used for the treatment of
advanced prostatic cancer: Results from European Organization for
Research on Treatment of Cancer Trials 30761 and 30762. J Urol.
135:303–307. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ponnusamy S, Sullivan RD, Thiyagarajan T,
Tillmann H, Getzenberg RH and Narayanan R: Tissue selective
androgen receptor modulators (SARMs) increase pelvic floor muscle
mass in ovariectomized mice. J Cell Biochem. 118:640–646. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chisamore MJ, Gentile MA, Dillon GM, Baran
M, Gambone C, Riley S, Schmidt A, Flores O, Wilkinson H and Alves
SE: A novel selective androgen receptor modulator (SARM) MK-4541
exerts anti-androgenic activity in the prostate cancer xenograft
R-3327G and anabolic activity on skeletal muscle mass &
function in castrated mice. J Steroid Biochem Mol Biol. 163:88–97.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Blanqué R, Lepescheux L, Auberval M, Minet
D, Merciris D, Cottereaux C, Clément-Lacroix P, Delerive P and
Namour F: Characterization of GLPG0492, a selective androgen
receptor modulator, in a mouse model of hindlimb immobilization.
BMC Musculoslelet Disord. 15:2912014. View Article : Google Scholar
|
|
32
|
Akita K, Harada K, Ichihara J, Takata N,
Takahashi Y and Saito K: A novel selective androgen receptor
modulator, NEP28, is efficacious in muscle and brain without
serious side effects on prostate. Eur J Pharmacol. 720:107–114.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang X, Allan GF, Tannenbaum P, Sbriscia
T, Linton O, Lai MT, Haynes-Johnson D, Bhattacharjee S, Lundeen SG
and Sui Z: Pharmacological characterization of an imidazolopyrazole
as novel selective androgen receptor modulator. J Steroid Biochem
Mol Biol. 134:51–58. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Narayanan R, Mohler ML, Bohl CE, Miller DD
and Dalton JT: Selective androgen receptor modulators in
preclinical and clinical development. Nucl Recept Signal.
6:e0102008.PubMed/NCBI
|
|
35
|
Narayanan R, Coss CC, Yepuru M, Kearbey
JD, Miller DD and Dalton JT: Steroidal androgens and nonsteroidal,
tissue-selective androgen receptor modulator, S-22, regulate
androgen receptor function through distinct genomic and nongenomic
signaling pathways. Mol Endocrinol. 22:2448–2465. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Crawford J, Prado CM, Johnston MA, Gralla
RJ, Taylor RP, Hancock ML and Dalton JT: Study design and rationale
for phase 3 clinical development program of enobosarm, a selective
androgen receptor modulator, for the prevention and treatment of
muscle wasting in cancer patients (POWER Trials). Curr Oncol Rep.
18:372016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Garber K: No longer going to waste. Nat
Biotechnol. 34:458–461. 2016. View Article : Google Scholar : PubMed/NCBI
|