Introduction
Thyroid cancer is the most prevalent type of
endocrine neoplasm globally (1).
Papillary thyroid carcinoma (PTC) is the most common type of
thyroid cancer accounting for ~80% of all thyroid cancer cases
worldwide (2). The incidence of PTC
has increased over the last four decades (3). The majority of patients with PTC have a
good prognosis, and the 5-year survival can achieve >95%
(2). However, a small fraction of
patients develop PTC with aggressive features. These patients may
develop local and distant metastasis, thus the prognosis of these
cases is unsatisfactory and the 10-year survival falls to 10%
(4,5).
Despite the efforts to develop novel treatments for aggressive PTC,
these patients have not benefited much (6). Therefore, a better understanding of the
mechanisms underlying tumor progression in aggressive PTC is
required.
It has been reported that epithelial-mesenchymal
transition (EMT) is common in PTC and contributes to PTC metastasis
(7). EMT has been extensively studied
in tumor metastasis in recent years. The process of EMT is
characterized by the loss of cell-to-cell contact, the remodeling
of cytoskeleton and the gain of a migratory phenotype (8). EMT can be induced by various signals
within the tumor microenvironment, which activate tumor cell
intrinsic transcription factors, including Snail1, Snail2, Twist 1
and zinc finger E-box binding homeobox 1 (ZEB1) (9). The defining feature of EMT is the loss
of E-cadherin and the acquisition of Vimentin (10). Numerous EMT regulators, including
transforming growth factor (TGF)-β, Snail1, Snail2 and Twist have
been analyzed in PTC, and are associated with PTC metastasis
(11,12).
Discoidin domain receptor tyrosine kinase 2 (DDR2)
is a member of the receptor tyrosine kinase (RTK) family that can
be activated by collagen I, II, III and X (13,14).
Multiple studies have demonstrated that DDR2 is implicated in
several cancer cell behaviors, including tumor angiogenesis, cell
adhesion and matrix remodeling (15–17).
Recent studies have revealed that DDR2 upregulation is predictive
of poor prognosis in breast cancer, hepatocellular carcinoma (HCC),
head and neck squamous cell carcinoma, and gastric cancer. The
possible mechanism is that DDR2 upregulation induces EMT in these
cancer cells (15,18–20).
Rodrigues et al (21)
identified that DDR2 overexpression in aneuploidy PTC was
associated with mortality from disease and distant metastasis.
However, the expression profile of DDR2 in tumor tissues of PTC
patients with local metastasis was not reported.
In the present research, the expression levels of
DDR2 were detected in PTC tissues with local metastasis and human
cell lines, and the effect of DDR2 on EMT was investigated in PTC
cells. The present study aimed to identify the function of DDR2 on
EMT in PTC and to reveal the underlying mechanism.
Materials and methods
Cell lines and reagents
The human normal follicular cell line Nthy-ori 3–1,
and human PTC cell lines BCPAP, TPC-1 and GLAG-66 were obtained
from the Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). Cells were maintained in RPMI 1640 medium
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) with 10%
fetal bovine serum (FBS, Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and 1% penicillin-streptomycin at 37°C in a
humidified atmosphere with 5% CO2.
Primary antibodies against DDR2 (cat no. 12133;
dilution, 1:1,000), GAPDH (cat no. 5174; dilution, 1:2,000),
E-cadherin (cat no. 3195; dilution, 1:1,000), Vimentin (cat no.
5741; dilution, 1:1,000), phosphorylated (P)-extracellular
signal-regulated kinase (ERK)1/2 (cat no. 4370, dilution, 1:1,000),
ERK1/2 (cat no. 4695; dilution, 1:1,000), Snail1 (cat no. 3879,
dilution, 1:1,000), Snail2 (cat no. 9586; dilution, 1:1,000),
Twist1 (cat no. 46702; dilution, 1:1000) and ZEB1 (cat no. 3396;
dilution, 1:1,000) were purchased from Cell Signaling Technology,
Inc. (Danvers, MA, USA). The human recombinant TGF-β was purchased
from PeproTech, Inc. (Rocky Hill, NJ, USA). Human collagen I was
purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Tissue samples
Human PTC tumor tissues (TT) and adjacent non-tumor
tissues (NT) from 10 patients with PTC with local metastasis were
obtained from the Department of Head and Neck Surgery, Zhejiang
Cancer Hospital (Zhejiang, China). In total, 6 female patients and
4 male patients aged 45 to 73 years (mean age, 64.3 years) were
included in the present study. No patients received previous
neoadjuvant radiotherapy or chemotherapy. The clinical diagnosis
was confirmed by pathological analysis. The present study was
approved the Medical Ethics committee of Zhejiang Cancer Hospital
and written informed consent was obtained from all patients.
Total RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA from cell and tissue samples was extracted
using TRIzol® reagent (Takara Bio, Inc., Otsu, Japan).
The cDNA was synthesized by Primescript RT master mix (Takara Bio,
Inc.). The temperature protocols were as follows: 37°C for 30 min,
85°C for 4 sec and 4°C for 2 min. The RT-qPCR was performed using
the ABI PRISM 7300 Sequence Detector (Applied Biosystems; Thermo
Fisher Scientific, Inc.) with the SYBR-Green PCR kit (Takara Bio,
Inc., Otsu, Japan). The thermal cycling conditions for RT-qPCR was
as follows: One cycle of 2 min at 95°C, 40 cycles of 15 sec at
95°C, 15 sec at 58°C and 30 sec at 72°C. The relative mRNA changes
were calculated by 2−ΔΔCq method with GAPDH as an
internal control (22). The primer
sequences were as follows: DDR2 forward, 5′-CTCCCAGAATTTGCTCCAG-3′
and reverse, 5′-GCCACATCTTTTCCTGAGA-3′; E-cadherin forward,
5′-CGAGAGCTACACGTTCACGG-3′ and reverse,
5′-GGGTGTCGAGGGAAAAATAGG-3′; Vimentin forward,
5′-GACGCCATCAACACCGAGTT-3′ and reverse,
5′-CTTTGTCGTTGGTTAGCTGGT-3′; Snail1 forward,
5′-GCTCCACAAGCACCAAGAGT-3′ and reverse, 5′-ATTCCATGGCAGTGAGAAGG-3′;
GAPDH forward, 5′-GGAGCGAGATCCCTCCAAAAT-3′, and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′.
DDR2 overexpression
The pEGFP-DDR2 cDNA lentiviral vector was obtained
from Shanghai GeneChem Co., Ltd. (Shanghai, China). The vector used
was GV218 vector, with the elements of Ubi-MCS-EGFP-IRES-Puromycin
(Shanghai GeneChem Co., Ltd., Shanghai, China). The colon site was
AgeI/AgeI. The recombinant GV218-DDR2 was transfected into HEK293T
cells using Lipofectamine™ 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.). The supernants containing the lentivirus
were collected 72 h after transfection. The lentiviral transfection
was conducted following the manufacturer's protocol. Briefly, the
cells were seeded on a 24-weel plate and cultured overnight. Prior
to transfection, the lentivirus was diluted in Enhanced Infection
Solution (Eni.S, Shanghai GeneChem Co., Ltd.) to the concentration
of 1×107 TU/ml, and polybrene (Shanghai GeneChem Co.,
Ltd.) was diluted in complete medium to the final concentration of
50 µg/ml. To perform transfection, 300 µl fresh medium mixed with
10 µl lentivirus and 10 µl polybrene were added to the plates and
cultured for 8 h. Then the medium was substituted with fresh
medium. The cells were then incubated at 37°C for an additional 96
h.
Small interfering RNA (siRNA)
transfection
The siRNA was transfected in to PTC cells using
Lipofectamine RNAiMAX Regent (Invitrogen; Thermo Fisher Scientific,
Inc.) following the manufacturer's protocol. The siRNA of DDR2
(cat. no. sc-39922), Snail1 (cat. no. sc-39922), ERK2 (cat. no.
sc-35335) and scramble siRNAs (cat. no. sc-37007) were obtained
from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA). The scramble
siRNAs were used as a control. The concentration of all the siRNAs
used was 100 nM/l.
Wound healing assay
The cell migration was assessed by performing wound
healing assay. A total of 2×104 cells suspended in RPMI
1640 medium with 10% FBS were seeded on each side of a wound
healing culture insert (Ibidi, Munich, Germany). The culture
inserts were removed to create a cell-free area of ~500 µm
following cultivation at 37°C and 5% CO2 for 24 h.
Afterwards, the cells were incubated with 1 ml serum-free RPMI 1640
medium for another 48 h. Cell migration was imaged under an Olympus
BX51 microscope (Olympus Corporation, Tokyo, Japan).
Transwell migration and invasion
assays
Transwell migration assay was performed in 12-well
plates with BioCoat control inserts with 8-µm diameter pores (BD
Biosciences, Franklin Lakes, NJ, USA). A total of 1×105
cells suspended in 500 µl serum-free RPMI 1640 medium were seeded
in the upper chamber. The lower chamber was filled with RPMI 1640
medium with 10% FBS. After 24 h of incubation, cells on the upper
membrane were wiped out, while the cells on the opposite side were
stained with crystal violet (MCE China, Shanghai, China) for 5 min
at room temperature and imaged at magnification, ×40 under an
Olympus BX51 microscope. As for the Transwell invasion assay, a
BioCoat Matrigel invasion chamber (BD Biosciences) was utilized
following the same protocols of the Transwell migration assay. To
induce EMT, 6-well plates were coated with collagen I at the
concentration of 2 mg/ml, and TPC-1 cells and TPC-1 with DDR2 siRNA
transfection were incubated with 5 ng/ml TGF-β for 24 h at 37°C.
The ERK1/2 inhibitor U0126 was obtained from Selleck Chemicals
(Shanghai, China) and used at the concentration of 1 µM at 37°C for
24 h.
Statistical analysis
All the data are expressed as the mean ± standard
deviation. Statistical significance was established using SPSS
version 16.0 statistical software package (SPSS, Inc., Chicago, IL,
USA). The differences between groups were analyzed using the
Student's t-test and one-way analysis of variance with post hoc
contrasts by Student-Newman-Keuls test. P<0.05 was considered to
indicate a statistically significant difference.
Results
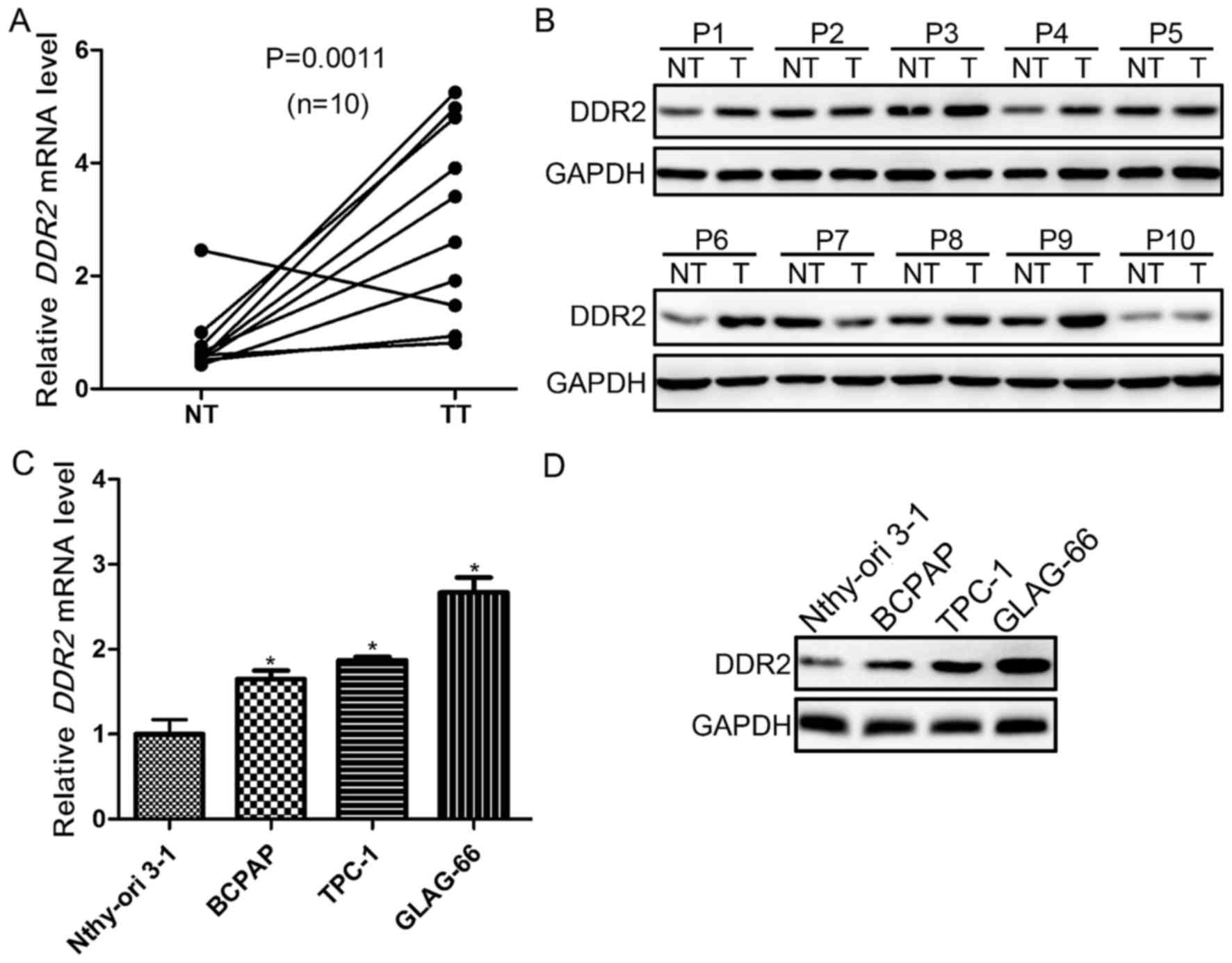
Upregulation of DDR2 is frequent in
PTC with local metastasis and human PTC cell lines
To explore the role of DDR2 in PTC with local
metastasis, its expression was evaluated in tumor and adjacent
non-tumor tissues from 10 patients with PTC with local metastasis
using RT-qPCR, and western blotting. It was demonstrated that DDR2
was upregulated in 9/10 tumor samples. The expression of DDR2 was
significantly higher in tumor compared with non-tumor tissues
(P=0.0011; Fig. 1A). The protein
expression levels were then detected in the tumor and non-tumor
tissues. The DDR2 protein level was markedly higher in PTC tumor
tissues compared with non-cancerous tissues (Fig. 1B). Furthermore, the DDR2 expression in
normal follicular cell line (Nthy-ori 3-1) and PTC cell lines
(BAPCP, TPC-1 and GLAG-66) was also determined using RT-qPCR, and
western blotting. The DDR2 expression in PTC cell lines was
significantly upregulated compared with Nthy-ori 3-1 cells at the
mRNA and protein level (Fig. 1C and
D).
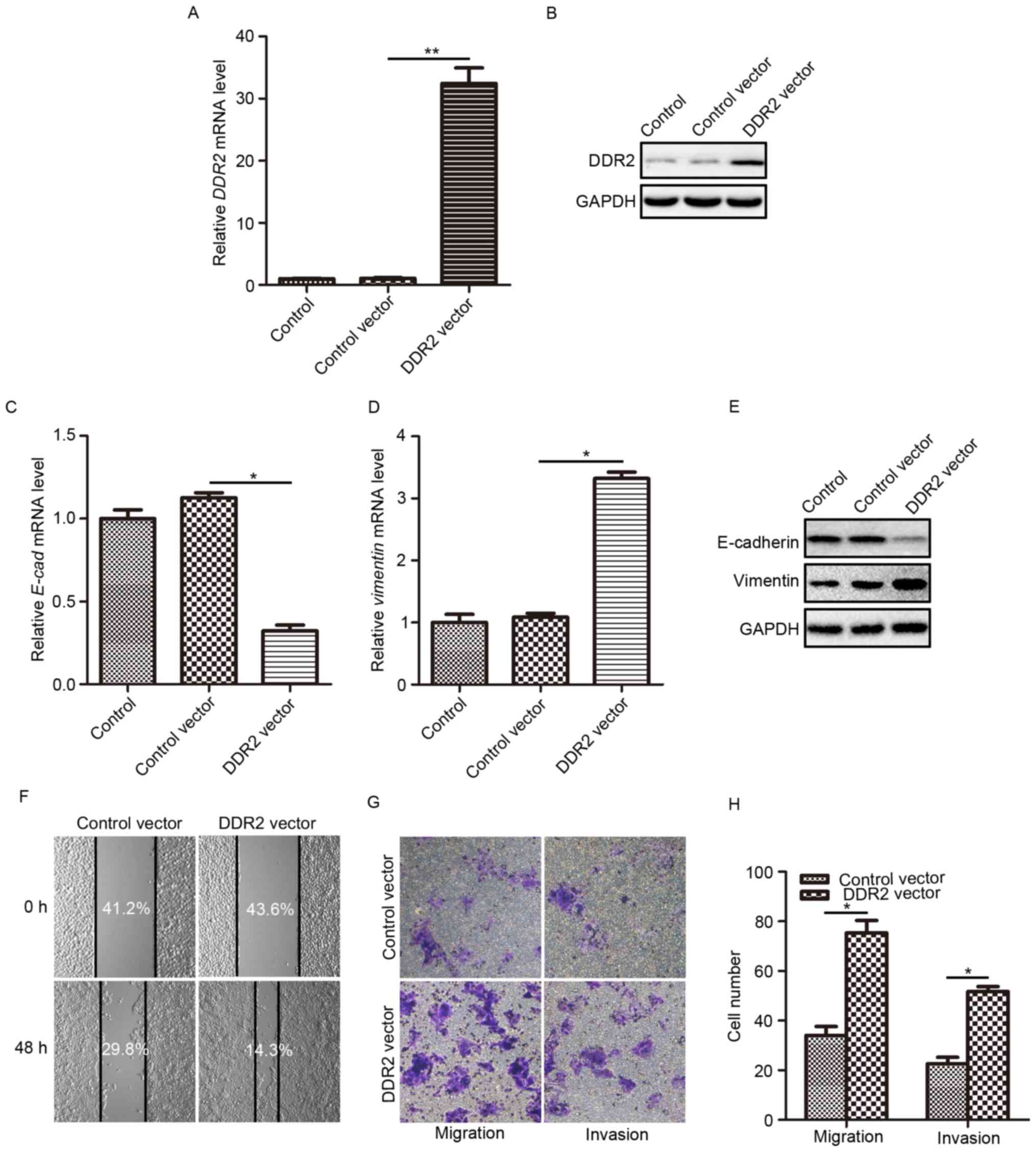
Overexpression of DDR2 induces EMT,
and promotes invasion and migration of TPC-1 cells
According to the research by Xie et al
(18), the upregulation of DDR2 may
induce EMT in HCC cells. To fully understand how DDR2 affects the
behaviors of PTC cells and to explore whether DDR2 upregulation
contributes to PTC cell EMT, the effects DDR2 overexpression on
cell invasion and migration were evaluated. TPC-1 cells were
infected with lentivirus carrying the human DDR2-expressing vector.
The overexpression efficiency was verified using RT-qPCR and
western blotting. It was demonstrated that the transduction of the
DDR2 vector caused a significant elevation in DDR2 mRNA expression
(Fig. 2A) and protein level (Fig. 2B) compared with the control, and
control vector groups. Then, the E-cadherin and Vimentin expression
levels were determined in TPC-1 cells with elevated DDR2. The
epithelial cell marker E-cadherin mRNA level was significantly
reduced (Fig. 2C) and the mesenchymal
cell marker Vimentin mRNA level was significantly increased
(Fig. 2D) following DDR2
upregulation. The changes in E-cadherin and Vimentin protein level
were consistent with that at the mRNA level (Fig. 2E).
Subsequently, the migration and invasion capacity of
TPC-1 cells following DDR2 upregulation were assessed using wound
healing and Transwell assays. The wound healing assay demonstrated
that following incubation with serum-free medium for 48 h, cell
migration was markedly enhanced by DDR2 upregulation (Fig. 2F). The migration assay and invasion
assay conducted using the Transwell chamber system also revealed
that DDR2 upregulation significantly promoted TPC-1 cell migration,
and invasion (Fig. 2G and H) compared
with the control vector-treated cells.
The aforementioned results demonstrated that DDR2
upregulation induced EMT in TPC-1 cells, which facilitated PTC cell
migration and invasion.
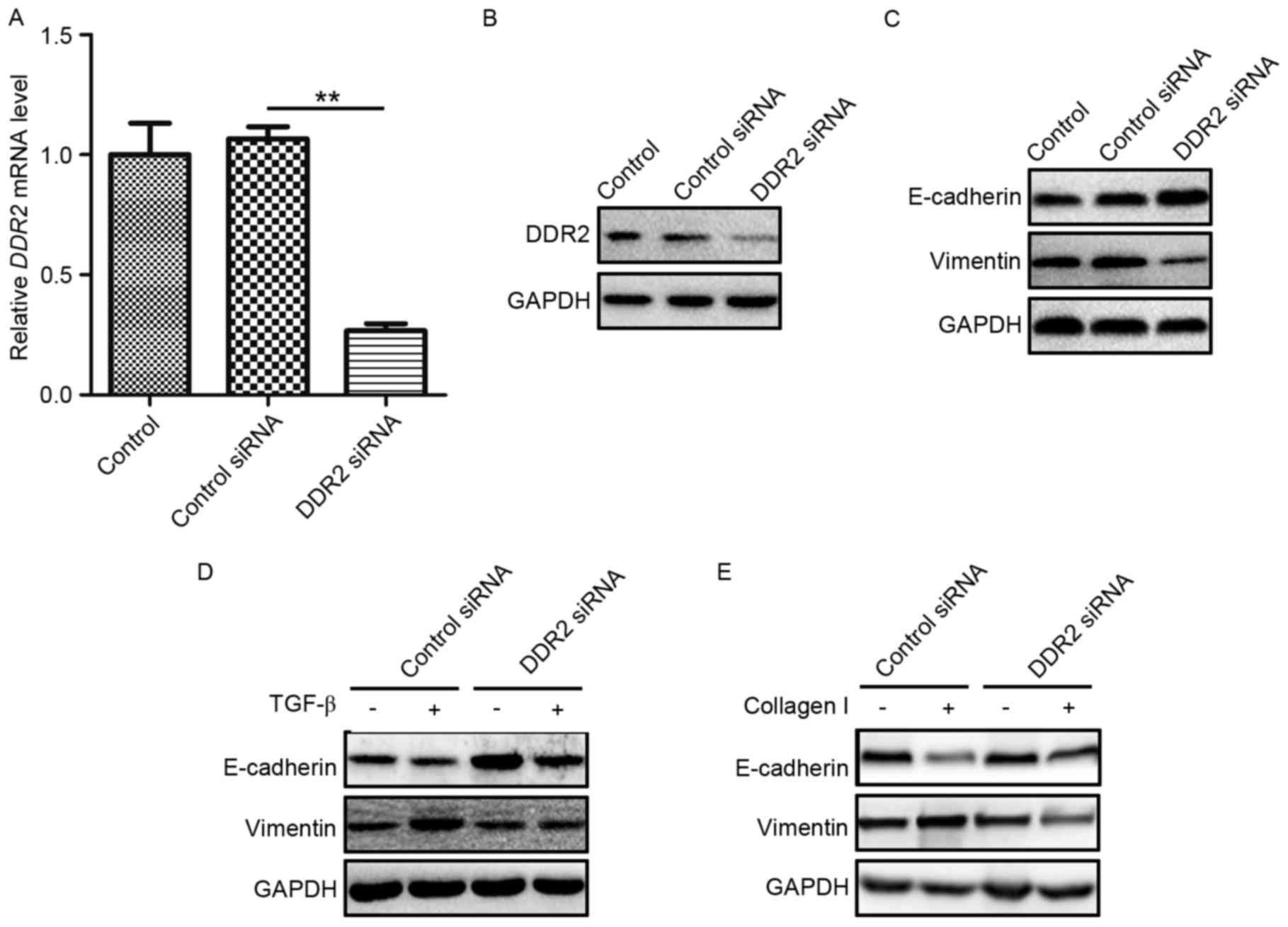
Downregulation of DDR2 reverses TGF-β-
and collagen I-induced EMT in TPC-1 cells
As DDR2 has been reported to possess a prominent
role in the pathway of TGF-β-induced EMT, and as a RTK, DDR2 may
directly bind to collagen I and induce EMT (15), in the present study, whether DDR2 was
a key regulator in the TGF-β-and collagen I-induced EMT was
evaluated in PTC. TPC-1 cells were transfected with DDR2 siRNA or a
scrambled non-specific siRNA as a negative control. The knockdown
efficiency of DDR2 siRNA was verified by RT-qPCR and western
blotting (Fig. 3A and B). E-cadherin
and Vimentin protein levels were then detected in TPC-1 cells
transfected with DDR2 siRNA. DDR2 siRNA markedly increased
E-cadherin and reduced Vimentin levels (Fig. 3C).
To assess the function of DDR2 in TGF-β-induced EMT
in PTC cells, TPC-1 cells transfected with DDR2 siRNA were treated
with recombinant TGF-β (5 ng/ml) for 24 h and western blotting was
performed using lysates from the cells. The silencing of DDR2 by
siRNA reversed TGF-β-induced EMT in PTC cells (Fig. 3D). To determine the role in DDR2 in
collagen I-induced EMT in PTC cells, TPC-1 cells were transfected
with DDR2 siRNA and cultured on plates pre-treated with collagen I
(2 mg/ml) for 24 h. The western blotting results demonstrated that
DDR2 siRNA reversed the decrease in E-cadherin and the increase in
Vimentin protein levels (Fig.
3E).
These results demonstrated that the downregulation
of DDR2 reversed TGF-β- and collagen I-induced EMT in PTC
cells.
Overexpression of DDR2 increases
Snail1 protein expression
To explore the underlying mechanism by which DDR2
regulated EMT in PTC cells, the protein expression levels of four
classical EMT transcription factors (Snail1, Snail2, Twist1 and
ZEB1) were determined in TPC-1 cells with increased and decreased
DDR2 expression. It was demonstrated DDR2 upregulation increased
the Snail1 protein level, while the silencing of DDR2 reduced the
Snail1 protein level (Fig. 4A and B).
The changes of DDR2 had no significant effect on the levels of
Snail2, Twist1 and ZEB1. The RT-qPCR results revealed that the
overexpression or depletion of DDR2 did not significantly affect
Snail1 mRNA expression (Fig. 4C and
D).
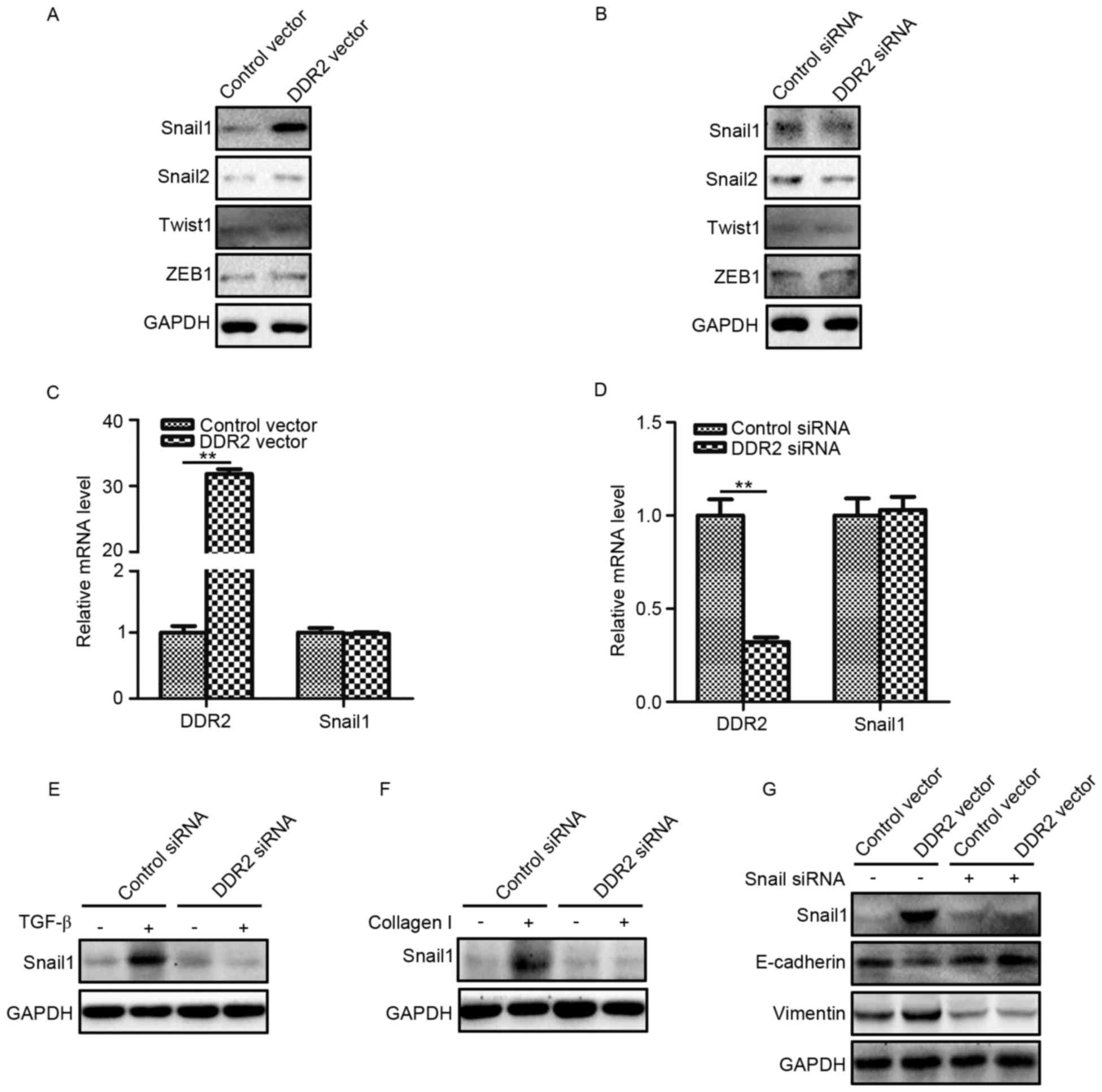 | Figure 4.Overexpression of DDR2 increases
Snail1 protein expression. (A) TPC-1 cells transfected with DDR2
vector or control vector were subjected to Snail1, Snail2, Twist1
and ZEB1. (B) TPC-1 cells transfected with DDR2 siRNA or control
siRNA were subjected to western blotting for Snail1, Snail2, Twist1
and ZEB1 (C) RT-qPCR results of Snail1 in TPC-1 cells transfected
with DDR2 vector or control vector. (D) RT-qPCR results of Snail1
in TPC-1 cells transfected with DDR2 siRNA or control siRNA. (E)
TPC-1 cells transfected with DDR2 siRNA or control siRNA were
incubated with 5 ng/ml of TGF-β for 48 h. The Snail1 protein level
was detected by western blotting. (F) TPC-1 cells transfected with
DDR2 siRNA or control siRNA were incubated with 2 µg/ml collagen I
for 48 h. The Snail1 protein level was detected by western
blotting. (G) TPC-1 cells transfected with DDR2 vector or control
vector were transduced with Snail1 siRNA. The protein level of
Snail1, E-cadherin and Vimentin was detected by western blotting.
**P<0.01. RT-qPCR, reverse transcription-quantitative polymerase
chain reaction; DDR2, discoidin domain receptor tyrosine kinase 2;
si, small interfering; TGF, transforming growth factor; ZEB, zinc
finger E-box binding homeobox 1. |
The expression of Snail1 in TGF-β- and collagen
I-treated TPC-1 cells transduced with DDR2 siRNA were then
determined. The results demonstrated that the depletion of DDR2
inhibited TGF-β- and collagen I-induced Snail1 elevation (Fig. 4E and F). To further confirm that
Snail1 was the key transcription factor in DDR2 mediated EMT in PTC
cells, Snail1 siRNA was adopted to downregulate Snail1 expression
in TPC-1 cells transfected with DDR2 vector. It was revealed that
Snail1 siRNA abrogated DDR2 overexpression-induced E-cadherin
decreases and Vimentin increases (Fig.
4G).
These results suggested that the Snail1 protein
level increased by DDR2 may be responsible for DDR2-induced EMT in
PTC cells.
Inhibition of ERK2 reverses
DDR2-induced Snail1 upregulation and EMT
Zhang et al (15) reported that ERK2 may directly
phosphorylate Snail1, thus lead to Snail1 nuclear accumulation and
increase the Snail1 protein half-life. ERK activation in TPC-1
cells with DDR2 overexpression or depletion was investigated. It
was demonstrated that p-ERK1/2 was activated in TPC-1 cells with
increased DDR2 (Fig. 5A). DDR2
depletion in TPC-1 cells resulted in a downregulation of p-ERK1/2
(Fig. 5B). The inhibition of p-ERK1/2
by p-ERK1/2 inhibitor U0126 reversed Snail1 upregulation, and the
subsequent E-cadherin decrease and Vimentin increase induced by
DDR2 overexpression. To further confirm the role of ERK2 in
DDR2-induced Snail1 upregulation, ERK2 siRNA was transfected into
TPC-1 cells with DDR2 overexpression. ERK2 depletion attenuated the
increase in Snail1, Vimentin and the decrease in E-cadherin.
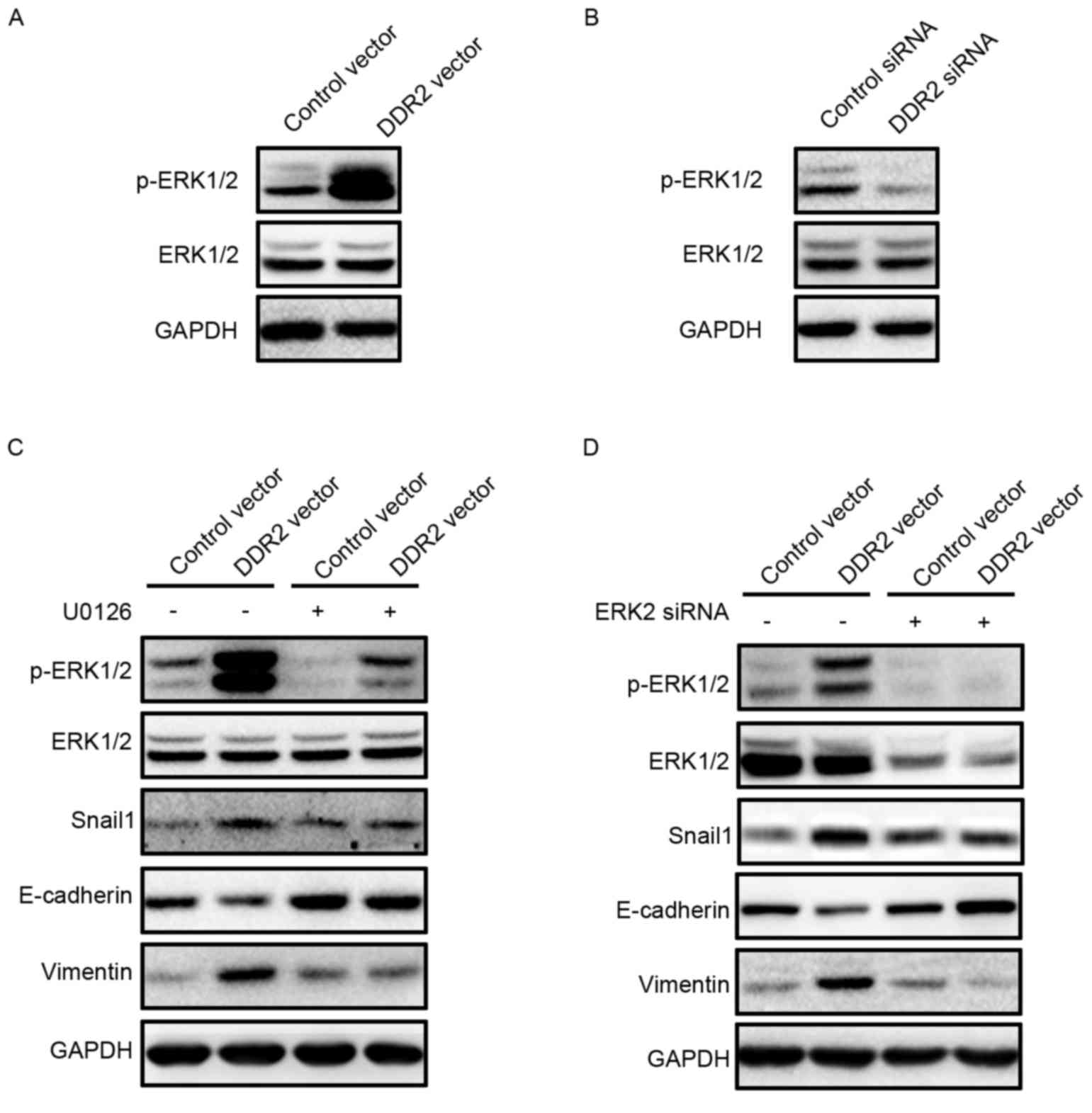 | Figure 5.Inhibition of ERK2 reverses
DDR2-induced Snail1 upregulation and EMT. (A) TPC-1 cells
transfected with DDR2 vector or control vector were subjected to
western blotting for p-ERK1/2. (B) TPC-1 cells transfected with
DDR2 siRNA or control siRNA were subjected to western blotting for
p-ERK1/2. (C) TPC-1 cells transfected with DDR2 vector or control
vector were incubated with 1 µM of U0126 for 24 h. The p-ERK1/2,
Snail1, E-cadherin and Vimentin protein level was detected by
western blotting. (D) TPC-1 cells transfected with DDR2 vector or
control vector were transfected with ERK2 siRNA. The p-ERK1/2,
Snail1, E-cadherin and Vimentin protein level was detected by
western blotting. RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; DDR2, discoidin domain receptor tyrosine
kinase 2; si, small interfering; TGF, transforming growth factor;
p, phosphorylated; ERK, extracellular signal-regulated kinase. |
These data suggested that DDR2 upregulation
activated ERK2, which increased the Snail1 protein level and thus
promoted EMT in PTC cells.
Discussion
In the present study, it was observed that DDR2 was
upregulated in the PTC tumor tissues with local metastasis and
human PTC cell lines when compared with non-tumor tissue samples,
and a normal follicular cell line. The upregulation of DDR2 induced
EMT in PTC cells, and promoted cell migration and invasion. The
downregulation of DDR2 reversed TGF-β- and collagen I-induced EMT.
The underlying mechanism beneath DDR2-induced EMT in PTC cells was
also investigated in the present study. It was revealed that DDR2
overexpression significantly increased Snail1 protein expression
without affecting its mRNA expression. The inhibition of Snail1 was
sufficient to abrogate DDR2 overexpression-induced PTC cell EMT. In
addition, the activation of ERK2 was revealed to increase Snail1
mRNA expression level.
As a RTK, DDR2 has been identified to be upregulated
in a series of cancer cells and associated with poor prognosis in
numerous cancer types such as breast cancer (15), HCC (18), head and neck squamous cell carcinoma
(19) and gastric cancer (20). Zhang et al (15) reported that DDR2 was expressed in
human invasive ductal breast cancer and correlated with a worse
survival rate. In addition, Xie et al (18) demonstrated that DDR2 was highly
expressed in HCC tumor tissues compared with in non-cancerous
tissues. The DDR2 overexpression was predictive of short overall
survival and disease-free survival in patients with HCC (18). In head and neck squamous cell
carcinoma, DDR2 has been identified as upregulated in tumor tissues
(19). The frequency and expression
intensity of DDR2 were associated with the tumor pathological stage
and lymph node metastasis (19). In a
newly published study by Wang et al (20), high DDR2 expression in gastric cancer
was identified to be associated with multiple tumor locations and
intestinal-type gastric cancer. To the best of our knowledge, there
are no previous studies regarding the expression profile of DDR2 in
tumor tissues of PTC with local metastasis. A previous study by
Rodrigues et al (21)
demonstrated that DDR2 was overexpressed in aneuploidy PTC, and was
associated with disease-associated mortality and distant
metastasis. In the current study, it was revealed that DDR2
overexpression was a frequent event in tumor tissues of PTC with
local metastasis and human PTC cell lines. However, the correlation
between DDR2 overexpression and prognosis was not analyzed, due to
lack of long-term follow-up data, which should be further
investigated.
DDR2 was an essential mediator of TGF-β1- and
collagen-induced EMT (23,24). In tumor tissues, DDR2 overexpression
is often positively correlated with Vimentin and negatively with
E-cadherin (19,20). DDR2 overexpression in tumor cells has
previously been demonstrated to be sufficient in inducing
morphological changes, and promoting enhanced migration and
invasion, which are essential biological processes in tumor
metastasis (23,25). The increased migration and invasion
capacity are dependent on the secretion of matrix
metalloproteinases (MMP) (26,27). MMP-2
and MMP-9 are two important subsets of downstream target genes of
DDR2 signaling (16). Increased DDR2
may promote the generation and secretion of MMP-2, and MMP-9 in HCC
and head and neck squamous cell carcinoma (18,19). In
the present study, it was observed that the overexpression of DDR2
decreased E-cadherin and increased Vimentin expression levels, and
promoted the migration and invasion of TPC-1 cells. It was
speculated that the induced EMT, and enhanced migration and
invasion were also the results of elevated MMP secretion.
The induced EMT following DDR2 overexpression
observed in this study occurred partially due to the activation of
ERK2 and the subsequent increase in Snail1 (23). DDR2 overexpression was identified to
be associated with ERK activation and increased Snail1 protein
level in tumor tissues. The present results suggest that the
interaction between DDR2 and collagen I stimulated ERK2 activity,
which then directly phosphorylated Snail1, leading to nuclear
accumulation, and increased the Snail1 protein half-life. ERK2
activation is essential for Snail1 stabilization and EMT induction
(28). DDR2 stabilizes Snail1 protein
expression post-transcriptionally without affecting the
transcription of Snail1 (15). This
was consistent with the findings in the current study whereby
overexpression of DDR2 did not change the mRNA expression of
Snail1, but increased its protein expression.
In conclusion, the present study revealed that
overexpression of DDR2 in tumor tissues of PTC with local
metastasis and human PTC cell lines was a frequent event.
Furthermore, the results suggest that the upregulation of DDR2
induced EMT, and promoted cell migration and invasion of PTC cells
by activating the ERK2/Snail1 signaling pathway, making it a
promising therapeutic target for reducing PTC local and distant
metastasis.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen AY, Jemal A and Ward EM: Increasing
incidence of differentiated thyroid cancer in the United States,
1988–2005. Cancer. 115:3801–3807. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jung CK, Little MP, Lubin JH, Brenner AV,
Wells SA Jr, Sigurdson AJ and Nikiforov YE: The increase in thyroid
cancer incidence during the last four decades is accompanied by a
high frequency of BRAF mutations and a sharp increase in RAS
mutations. J Clin Endocrinol Metab. 99:E276–E285. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sipos JA and Mazzaferri EL: Thyroid cancer
epidemiology and prognostic variables. Clin Oncol (R Coll Radiol).
22:395–404. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kondo T, Ezzat S and Asa SL: Pathogenetic
mechanisms in thyroid follicular-cell neoplasia. Nat Rev Cancer.
6:292–306. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luster M, Weber T and Verburg FA:
Differentiated thyroid cancer-personalized therapies to prevent
overtreatment. Nat Rev Endocrinol. 10:563–574. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vasko V, Espinosa AV, Scouten W, He H,
Auer H, Liyanarachchi S, Larin A, Savchenko V, Francis GL, de la
Chapelle A, et al: Gene expression and functional evidence of
epithelial-to-mesenchymal transition in papillary thyroid carcinoma
invasion. Proc Natl Acad Sci USA. 104:pp. 2803–2808. 2007;
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zavadil J and Böttinger EP: TGF-beta and
epithelial-to-mesenchymal transitions. Oncogene. 24:5764–5774.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shook D and Keller R: Mechanisms,
mechanics and function of epithelial-mesenchymal transitions in
early development. Mech Dev. 120:1351–1383. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lv N, Shan Z, Gao Y, Guan H, Fan C, Wang H
and Teng W: Twist1 regulates the epithelial-mesenchymal transition
via the NF-kappaB pathway in papillary thyroid carcinoma.
Endocrine. 51:469–477. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Buehler D, Hardin H, Shan W,
Montemayor-Garcia C, Rush PS, Asioli S, Chen H and Lloyd RV:
Expression of epithelial-mesenchymal transition regulators SNAI2
and TWIST1 in thyroid carcinomas. Mod Pathol. 26:54–61. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Leitinger B and Kwan AP: The discoidin
domain receptor DDR2 is a receptor for type X collagen. Matrix
Biol. 25:355–364. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Carafoli F, Bihan D, Stathopoulos S,
Konitsiotis AD, Kvansakul M, Farndale RW, Leitinger B and
Hohenester E: Crystallographic insight into collagen recognition by
discoidin domain receptor 2. Structure. 17:1573–1581. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang K, Corsa CA, Ponik SM, Prior JL,
Piwnica-Worms D, Eliceiri KW, Keely PJ and Longmore GD: The
collagen receptor discoidin domain receptor 2 stabilizes SNAIL1 to
facilitate breast cancer metastasis. Nat Cell Biol. 15:677–687.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim D, Ko P, You E and Rhee S: The
intracellular juxtamembrane domain of discoidin domain receptor 2
(DDR2) is essential for receptor activation and DDR2-mediated
cancer progression. Int J Cancer. 135:2547–2557. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu H, Bihan D, Chang F, Huang PH, Farndale
RW and Leitinger B: Discoidin domain receptors promote α1β1-and
α2β1-integrin mediated cell adhesion to collagen by enhancing
integrin activation. PLoS One. 7:e522092012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xie B, Lin W, Ye J, Wang X, Zhang B, Xiong
S, Li H and Tan G: DDR2 facilitates hepatocellular carcinoma
invasion and metastasis via activating ERK signaling and
stabilizing SNAIL1. J Exp Clin Cancer Res. 34:1012015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu J, Lu W, Zhang S, Zhu C, Ren T, Zhu T,
Zhao H, Liu Y and Su J: Overexpression of DDR2 contributes to cell
invasion and migration in head and neck squamous cell carcinoma.
Cancer Biol Ther. 15:612–622. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang YG, Xu L, Jia RR, Wu Q, Wang T, Wei
J, Ma JL, Shi M and Li ZS: DDR2 induces gastric cancer cell
activities via activating mTORC2 signaling and is associated with
clinicopathological characteristics of gastric cancer. Dig Dis Sci.
61:2272–2283. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rodrigues R, Roque L, Espadinha C, Pinto
A, Domingues R, Dinis J, Catarino A, Pereira T and Leite V:
Comparative genomic hybridization, BRAF, RAS, RET, and oligo-array
analysis in aneuploid papillary thyroid carcinomas. Oncol Rep.
18:917–926. 2007.PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Walsh LA, Nawshad A and Medici D:
Discoidin domain receptor 2 is a critical regulator of
epithelial-mesenchymal transition. Matrix Biol. 30:243–247. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Provenzano PP, Inman DR, Eliceiri KW and
Keely PJ: Matrix density-induced mechanoregulation of breast cell
phenotype, signaling and gene expression through a FAK-ERK linkage.
Oncogene. 28:4326–4343. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang ZL, Fan ZQ, Jiang HD and Qu JM:
Selective Cox-2 inhibitor celecoxib induces epithelial-mesenchymal
transition in human lung cancer cells via activating MEK-ERK
signaling. Carcinogenesis. 34:638–646. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Olaso E, Ikeda K, Eng FJ, Xu L, Wang LH,
Lin HC and Friedman SL: DDR2 receptor promotes MMP-2-mediated
proliferation and invasion by hepatic stellate cells. J Clin
Invest. 108:1369–1378. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Labrador JP, Azcoitia V, Tuckermann J, Lin
C, Olaso E, Mañes S, Brückner K, Goergen JL, Lemke G, Yancopoulos
G, et al: The collagen receptor DDR2 regulates proliferation and
its elimination leads to dwarfism. EMBO Rep. 2:446–452. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Strippoli R, Benedicto I, Pérez Lozano ML,
Cerezo A, López-Cabrera M and del Pozo MA:
Epithelial-to-mesenchymal transition of peritoneal mesothelial
cells is regulated by an ERK/NF-kappaB/Snail1 pathway. Dis Model
Mech. 1:264–274. 2008. View Article : Google Scholar : PubMed/NCBI
|