Introduction
Human osteosarcoma is a leading cause of
tumor-associated mortality in children and young adults; currently
there is an ~70% five-year survival rate following treatment with
combination chemotherapy (1,2). However, a significant proportion of
patients with osteosarcoma exhibit a poor response to chemotherapy,
in particular to cisplatin treatment, and suffer a high risk of
local relapse or metastasis following intensive combination
chemotherapy (3).
MicroRNAs (miRNAs) are small non-coding RNAs that
are 19–21 nucleotides long and are highly conserved (4). miRNAs serve regulatory roles by
modulating gene expression at the posttranscriptional level by
binding the 5′-untranslated region (5′-UTR), coding sequences and
3′-UTR of target mRNA (5). miRNAs
directly regulate >60% of human protein coding genes or
non-coding genes, indicating their crucial roles in a wide range of
biological processes, including embryogenesis, development,
differentiation and apoptosis (6).
Previous studies have demonstrated that miRNAs also are involved in
tumorigenesis (6–8). In osteosarcoma, miR-21 was revealed to
be highly upregulated (9). The
neutralization of miR-21 suppressed invasion and migration in MG63
cells (10). The expression profile
of miR-34a was also significantly altered in osteosarcoma (11). As a regulated target of p53, miR-34a
inhibits p53-mediated cell cycle arrest, proliferation, apoptosis
and migration in osteosarcoma cells (12). Compared with osteoblasts, in
osteosarcoma cell lines, miR-199a-3p was overexpressed and
associated with a decrease in cell growth, and with G1
phase cell cycle arrest in a p53-independent manner (13). Notably, miRNAs are also involved in
the induction of chemoresistance. miR-132 and miR-140 have
previously been revealed to serve critical roles in the induction
of chemoresistance, which suggests the necessity of furthering
current understanding of the molecular mechanisms underlying this
disease (14,15).
miR-133b has previously been reported to be a
muscle-specific miRNA that serves a regulatory role in the
development of skeletal muscle (16).
In a recent study, miR-133b was identified to be downregulated in
osteosarcoma tissues, compared with the adjacent tissue (17). Subsequently, further research
demonstrated its critical role in promoting cell proliferation,
migration, invasion and apoptosis (18). However, the role of miR-133b in the
chemoresistance of osteosarcoma remains unclear.
Based on previous results, the present study
hypothesized that miR-133b may serve critical roles in regulating
the chemoresistance of osteosarcoma. In order to validate this
hypothesis, the present study evaluated the expression levels of
miR-133b in cisplatin-resistant MG63 cells and normal MG63 cells
using reverse transcription-quantitative polymerase chain reaction
(RT-qPCR). The present study aimed to reveal miR-133b as a novel
therapeutic target for treating chemoresistance.
Materials and methods
Cell culture and induction of the
cisplatin-resistant sub-line
The MG63 human osteosarcoma cell line (no. CRL-1427;
American Type Culture Collection, Manassas, VA, USA) was maintained
in Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.). The
cisplatin-resistant sub-line of MG63 (MG63-DDP) was derived from
the original MG63 cell line by continuous exposure to cisplatin
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). The initial dose
of cisplatin was 0.1 µM; after 72 h, the media was removed and
cells were allowed to recover for a further 72 h. This continuous
period lasted for six months. Cells were subsequently maintained in
the presence of a half-concentration of the inducing dose, 0.05 µM
cisplatin.
Proliferation assay (MTT)
Cells (5×103) were seeded into 96-well
plates and allowed to attach to the plate overnight at 37°C.
Following 0.2 µM cisplatin treatment, the MTT reagent was added to
cells and incubated for 4 h at 37°C according to the manufacturer's
instructions. Subsequently, dimethylsulphoxide was added to the
cells and mixed for 5 min according to manufacturer's instructions.
Absorbance at 595 nm was measured using a BioTek Synergy (BioTek
Instruments, Inc., Winooski, VT, USA).
Clonogenic survival assay
The clonogenic assay, which is used to determine the
effectiveness of cytotoxic agents, including chemotherapeutic
agents, was performed to determine the sensitivity of cells to
cisplatin. Cells (1×105) were seeded in a 6-well plate
and allowed to attach overnight at 37°C, and maintained with 0.25,
0.5, 0.75, 1, 1.5 or 2 µM cisplatin for 14–21 days. Subsequently,
the colonies were observed by fixation with 4% paraformaldehyde and
staining with methanol (25% v/v) substituted with crystal violet
(0.05% w/v) for 30 min, prior to being washed with 1X PBS. Colonies
>40 µm in diameter were counted using a X71 (U-RFL-T)
fluorescence microscope (Olympus, Melville, NY, USA).
Evaluation of cisplatin-DNA
adducts
Cells treated with the half maximal inhibitory
concentration (IC50) cisplatin (~1.36 µM) were fixed
using methanol at room temperature for 30 min, and then subjected
to proteolytic digestion with 100 µg/ml pepsin and 50 µg/ml
proteinase K at 37°C for 10 min. To block the non-specific binding
sites, fixed cells were incubated with PBS supplemented with 5%
(w/v) bovine serum albumin and 5% FBS for 30 min. Subsequently, PBS
was removed from the cells using a pipette without washing. The
antibody (MABE416; Millipore, Billerica, MA, U.S.A.) against
cisplatin-GpG DNA adducts, was added at dilution of 1:2,000 and
incubated at 37°C for 3 h. The primary antibody-DNA adducts complex
was detected using an anti-rat Cy5-labeled secondary antibody, goat
anti-Rat IgG (H+L) Cross-Adsorbed Secondary Antibody (cat. no.
A10525, 1:5,000 dilution; Thermo Fisher Scientific, Inc.). Cells
were subsequently incubated in 1 µg/ml DAPI and PBS for 10 min at
room temperature for nuclear counterstaining. Images were acquired
using an Olympus X71 fluorescence microscope (Olympus, Tokyo,
Japan).
Transfection of pre-miR™ miRNA
precursors (mimics) and anti-miR™ miRNA inhibitors into MG63 or
MG63-DDP cells
Mimics and anti-miRNA mimics for miR-133b were
purchased from Ambion; Thermo Fisher Scientific, Inc. MG63 cells
were transfected with miR-133b mimics (MG63/miR-133b mimics) or
scrambled miR-133b mimics (MG63/vector) using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Briefly, 100 nM miRNA
was mixed with 8 µl Lipofectamine® 2000 and incubated at
room temperature for 30 min. MG63-DDP cells were transfected with
anti-miR-133b mimics (MG63-DDP/anti-miR-133b) or scrambled
anti-miR-133b mimics (MG63-DDP/vector), following the
aforementioned protocol.
RNA extraction and RT-qPCR
RNA was extracted from the original MG63,
MG63/vector, MG63/miR-133b mimics, MG63-DDP, MG63-DDP/anti-miR-133b
or MG63-DDP/vector cells treated in the previously described
conditions, using the mirVana™ miRNA Isolation kit (Ambion; Thermo
Fisher Scientific, Inc.) and was treated with DNase I to eliminate
the contaminated genomic DNA. The detecting primers, M-MLV Reverse
Transcriptase, and PowerUpä SYBR® Green Master Mix were
also all supplied by Ambion; Thermo Fisher Scientifc, Inc. The
primer sequences were as follows: miR-133b forward,
5′-AAAGGACCCCAACAACCAGCAA-3 and reverse,
5′-TTGCTGGTTGTTGGGGTCCTTT-3′; and U6 small nuclear (sn)RNA forward,
5′-CTCGCTTCGGCAGCACAT ATA CT-3 and reverse,
5′-ACGCTTCACGAATTTGCGTGTC-3. The relative expression level for each
miRNA was determined using the comparative Cq method following
normalization to the level of U6 RNA. Individual samples were run
in triplicate on the Applied Biosystems ABI 7500 PCR system (Thermo
Fisher Scientific, Inc.) (19).
Scratch wound assay
A total of 1×105 cells were seeded in a
24-well plate and allowed to adhere overnight at 37°C. On the
second day, a scratch was made on a 100%-confluent monolayer of
cells using a sterile 200 µl disposable pipette tip, and cells were
washed with 3 ml PBS to remove debris. Images to evaluate cell
proliferation were captured using the Olympus X71 microscope
(Olympus) at 0 and 24 h following the scratch.
Invasion assay
To investigate the invasion ability of cells,
1×105 cells were seeded into the upper chamber of a
Transwell chamber with a Matrigel-coated membrane (Corning Inc.,
Corning, NY, USA). Medium without FBS but with cisplatin was added
to the upper chamber, while medium supplemented with 2% FBS and
cisplatin was applied to the lower chamber. The cells were
subsequently incubated at 37°C for 24 h. Cells that did not invade
through the pores were removed using a cotton swab. Cells on the
lower surface of the membrane were fixed with 4% paraformaldehyde,
stained with 0.05% crystal violet for 15 min at room temperature
and counted using an Olympus X71 microscope (Olympus).
Statistical analysis
Data in the present study are expressed as the mean
± standard deviation of a minimum of three independent experiments.
Analyses were performed using SPSS software version 17.0 (SPSS,
Inc., Chicago, IL, USA). An unpaired Student's t-test was used for
comparisons between two groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
MG63 cells with a cisplatin-resistant
phenotype demonstrate a higher miR-133b expression level, lower
proliferation and clonogenic ability
In order to investigate the miR-133b expression
profile in cisplatin-resistant osteosarcoma cells,
cisplatin-resistant MG63 (MG63-DDP) cells were generated by
long-term treatment with cisplatin, as mentioned previously. The
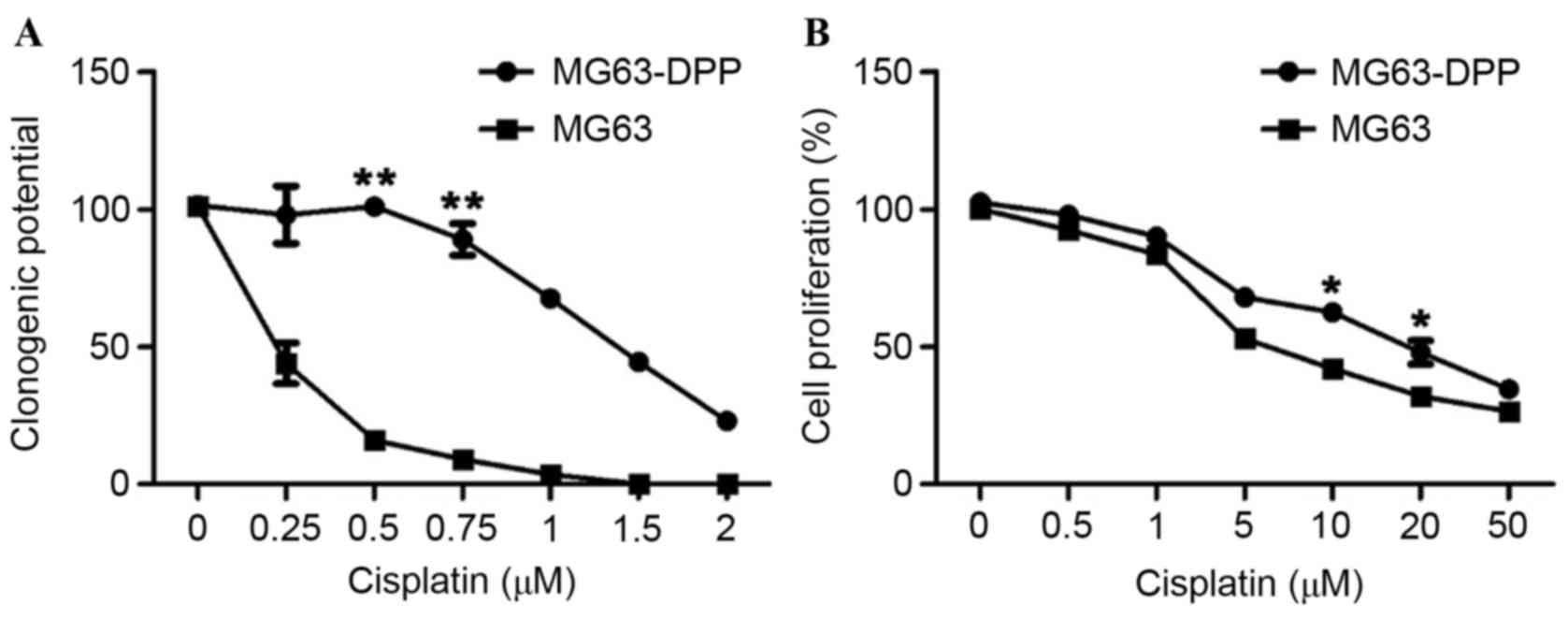
clonogenic assay determined that MG63-DDP cells are significantly
more resistant to cisplatin treatment compared with ordinary cells
(Fig. 1A; IC50, ~1.2 µM
for MG63-DDP cells; IC50, ~0.2 µM for MG63 cells). A
short-term proliferation assay was also performed to evaluate the
effects of cisplatin on cell division. Notably, a significant
difference in the level of proliferation at 48 h was observed
between MG63 and MG63-DDP cells (P<0.05); This indicated that
the cisplatin-resistant phenotype induced a difference in
clonogenic ability (Fig. 1B).
According to this observation, 0.2 µM cisplatin was selected for
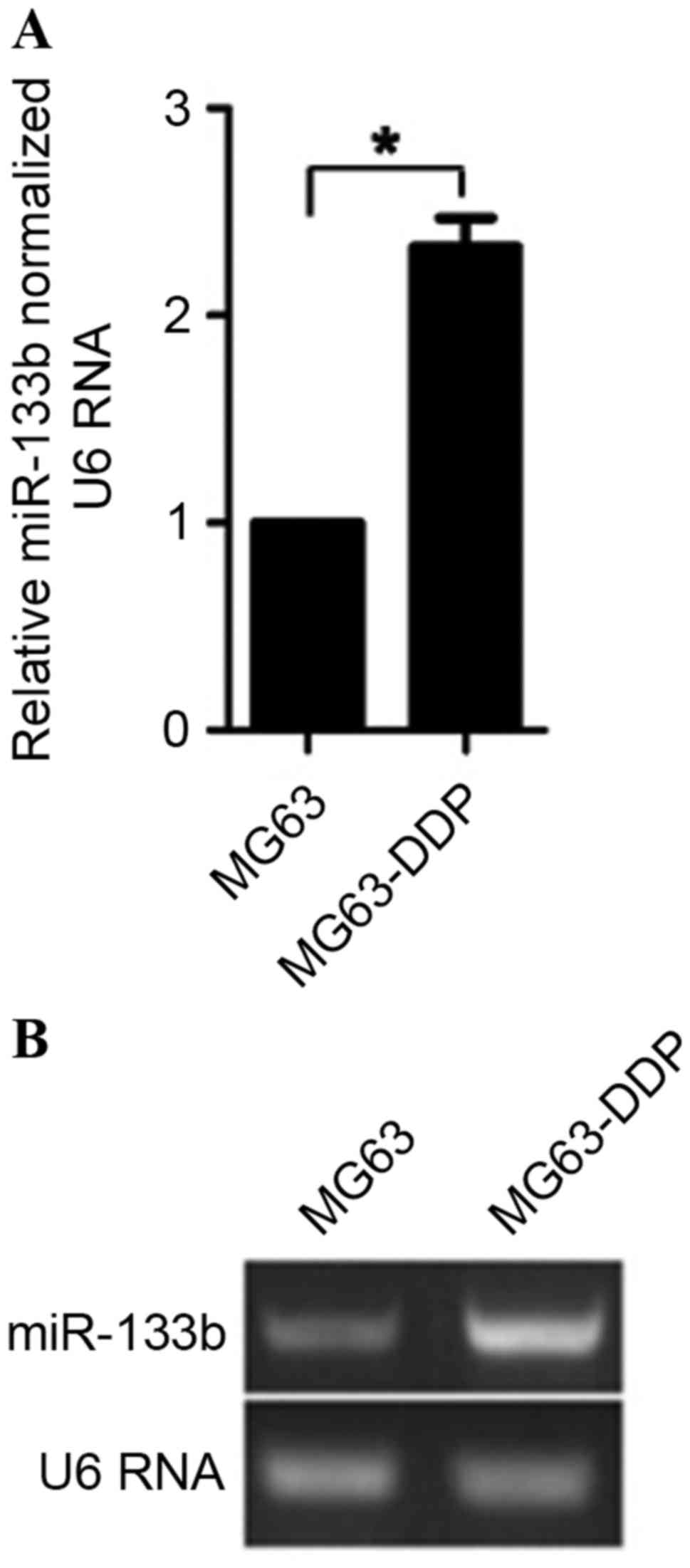
further study. To determine the difference in expression between
MG63-DDP cells and MG63 cells, semiquantitative and quantitative
RT-PCR were performed. The results demonstrated that MG63-DDP cells
expressed ~2.3-fold higher levels of miR-133b, compared with
ordinary MG63 cells (Fig. 2). This
suggests that the upregulation of miR-133b may have induced this
difference.
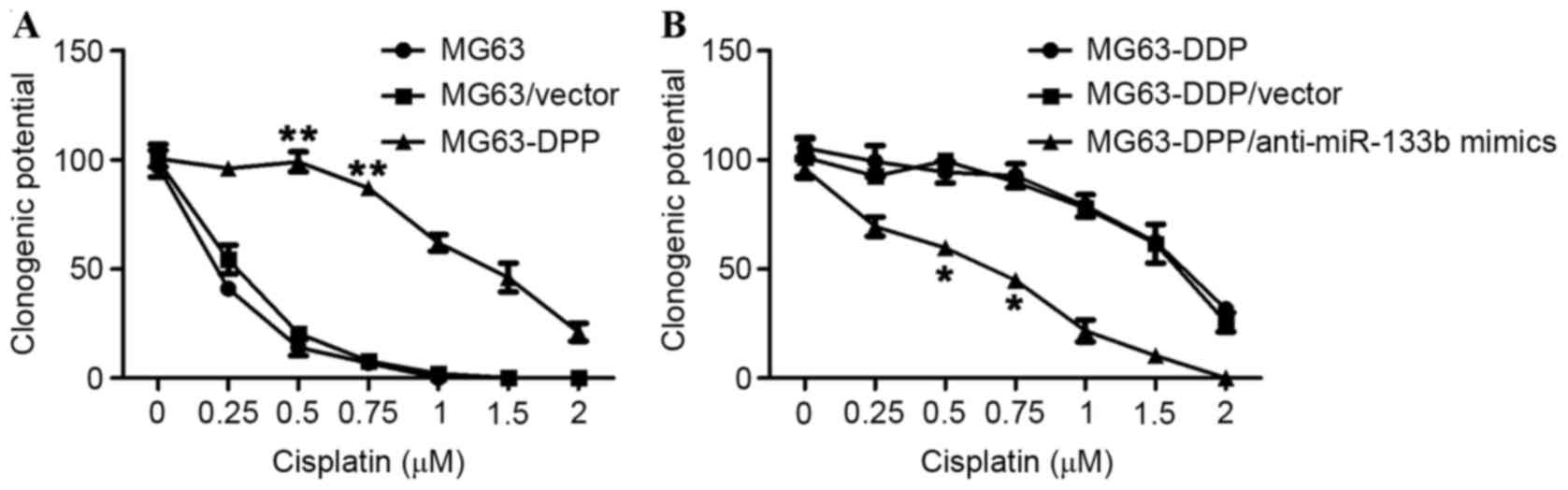
miR-133b mimic is sufficient to induce
a cisplatin-resistant phenotype in MG63 cells
As the aforementioned result demonstrated
upregulation of miR-133b in MG63-DDP cells, miR-133b may serve as a
potential effector for inducing a cisplatin-resistant phenotype.
Thus, the present study determined the efficacy of using miR-133b
mimics to induce a cisplatin-resistant phenotype, independent of
long-term treatment with cisplatin. The expressing vectors
containing the coding sequence of pre-miR-133b were transfected
into MG63 cells for 72 h, and subsequently evaluated using a
clonogenic assay. As hypothesized, promotion of the clonogenic
ability of MG63/pre-miR-133b cells was observed, which confirmed
the effect of miR-133b on the induction of a cisplatin-resistant
phenotype in MG63 cells (Fig. 3).
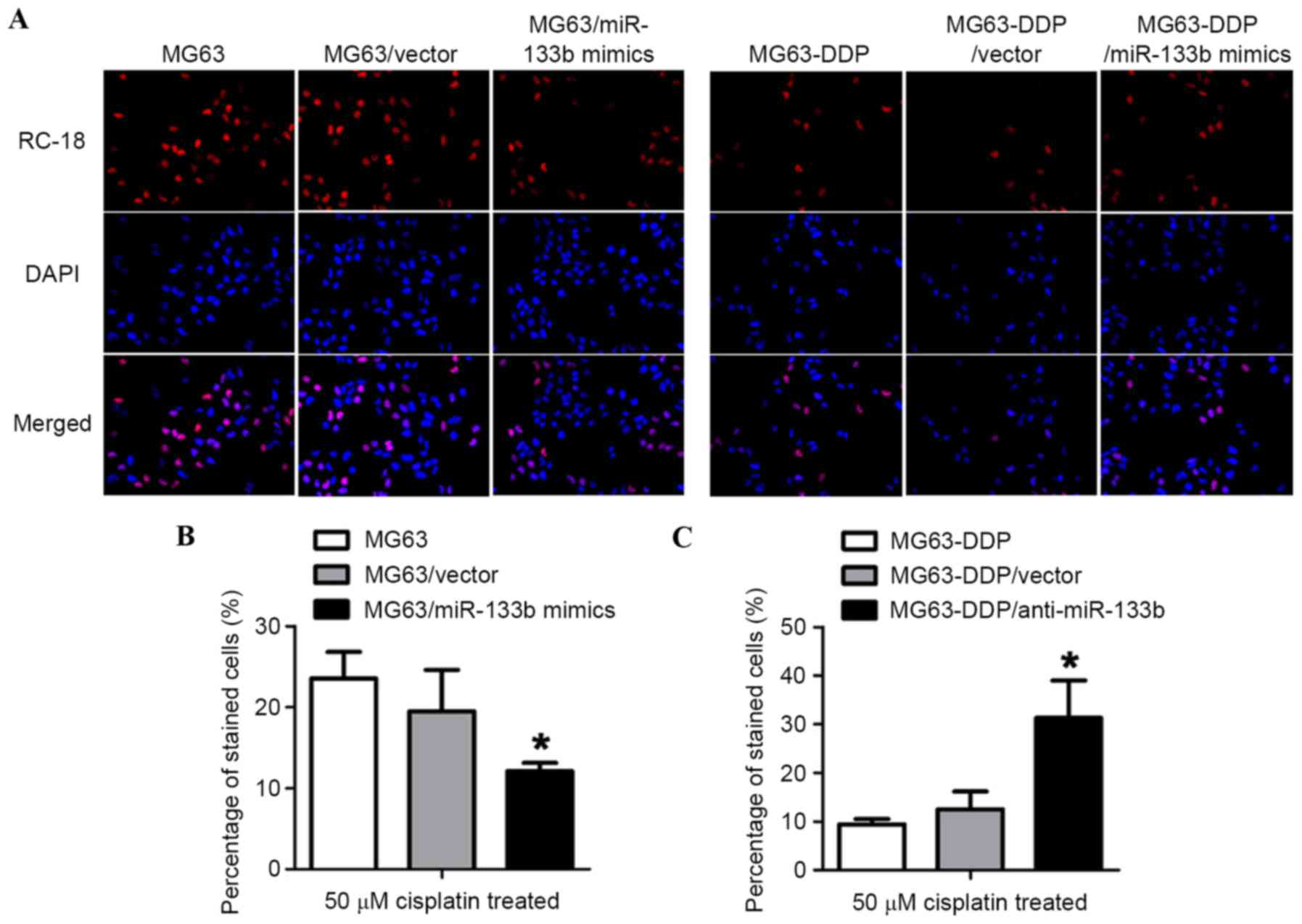
Epigenetic expression of miR-133b
promoted a decrease in cisplatin-1,2-intrastrand d (GpG)
crosslinking (GpG) DNA adduct formation and apoptotic ratio under
cisplatin stress
To qualitatively determine the level of DNA adducts
in the nuclear DNA of MG63-DDP cells, the cisplatin-treated cells
were stained for Platinum-GpG (Pt-(GpG)) cross-links in DNA using
the RC-18 antibody. It was revealed that the epigenic expression of
miR-133b decreased cisplatin-DNA adduct formation compared with
MG63 or MG63/vector cells (Fig. 4A,
left panel). As hypothesized, the introduction of anti-miR-133b
mimics markedly increased cisplatin-DNA adduct formation (Fig. 4A, right panel), which suggested that
miR-133b inhibited the accumulation of Pt-DNA lesions. To further
confirm the role of miR-133b in the cisplatin-resistant phenotype,
miR-133b mimics and the antisense strand of miR-133b mimics
(anti-miR-133b) were synthesized and introduced into the respective
cells. To investigate death rate, CFSE/PI dual staining was
performed. Compared with MG63 or MG63/vector cells, MG63 cells
transfected with miR-133b mimics (MG63/miR-133b mimics)
demonstrated a lower cell death ratio, indicating the cell's
cisplatin resistance (Fig. 4A).
Consistently, in MG63-DDP cells, the removal of miR-133b by
antisense strand miR-133b mimics revealed an increased sensitivity
to cisplatin compared with MG63-DDP and MG63-DDP/vector cells
(Fig. 4B and C). Taken together, the
expression levels of miR-133b in MG63 cells was sufficient to
induce cisplatin resistance, and disturbance of miR-133b expression
was also sufficient to make cisplatin-resistant MG63 cells more
sensitive to cisplatin.
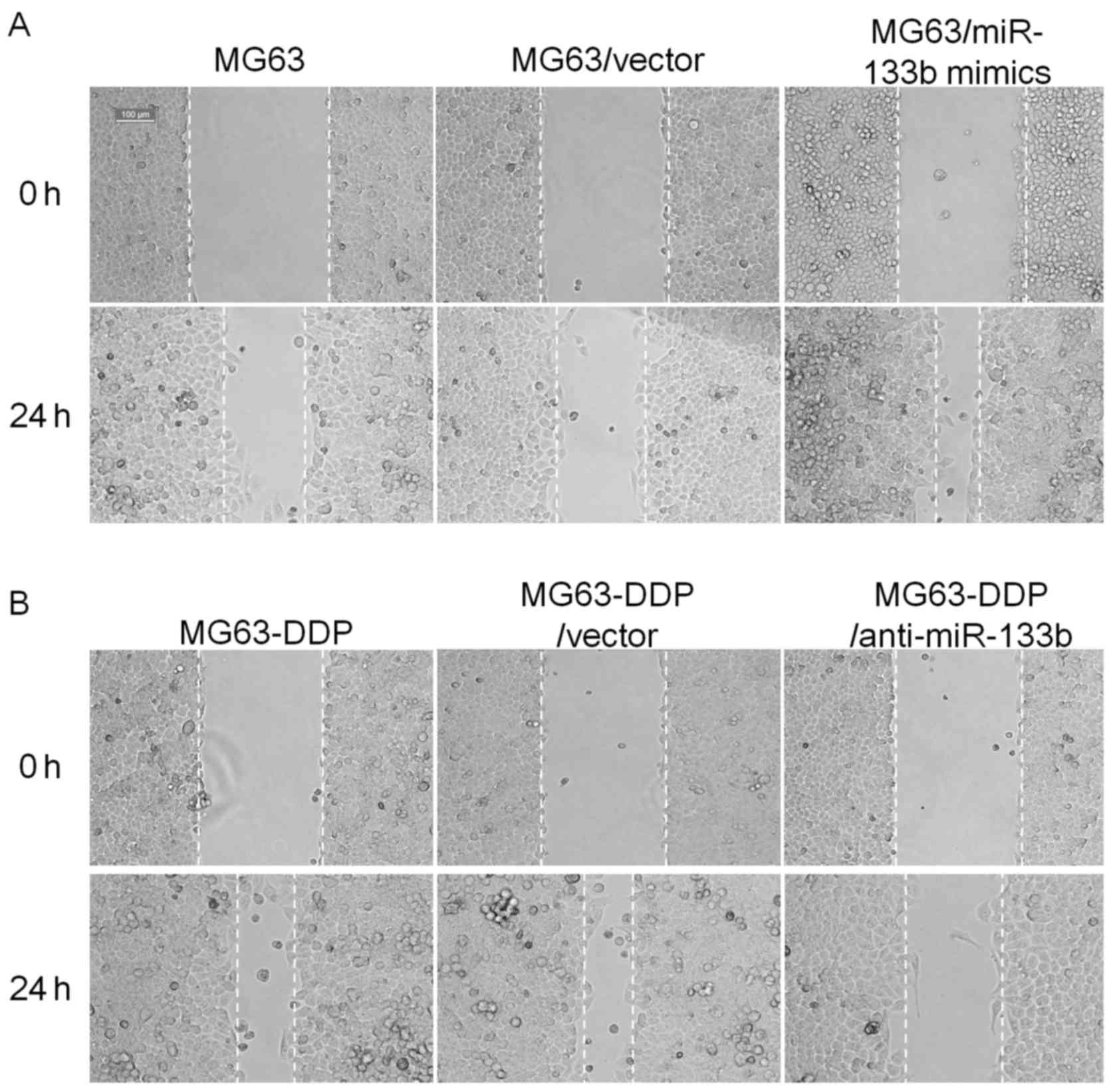
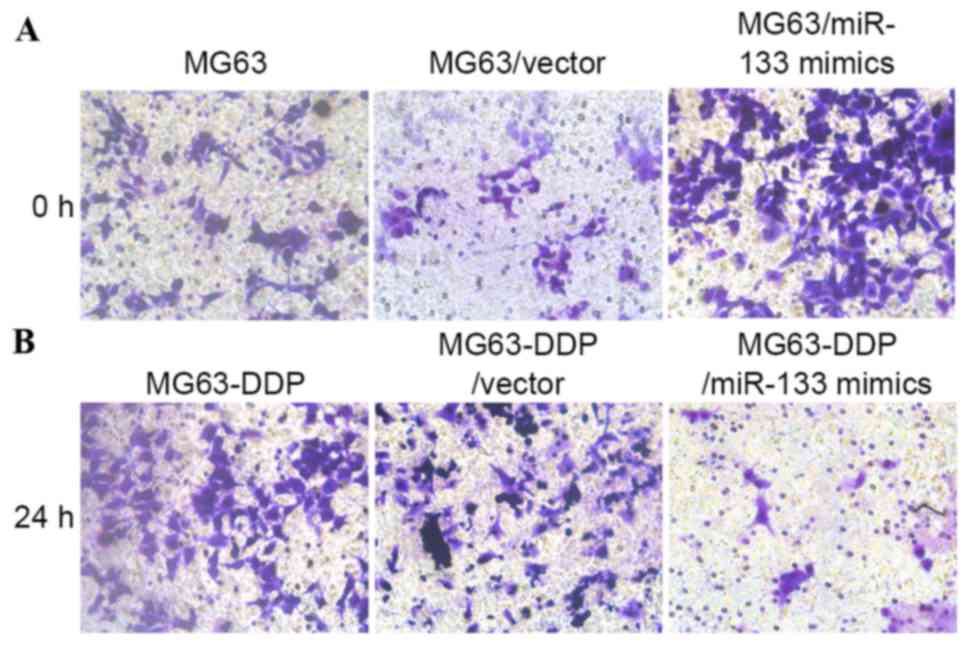
miR-133b expression level promotes
migration and invasion under cisplatin stress
To clarify the effect of miR-133b expression on the
migration and invasion of MG63 or MG63-DDP cells, two experiments
were performed. As presented in Fig.
5, the scratch wound assay demonstrated that cell migration was
markedly increased in MG63/miR-133b mimics compared with the
original MG63 or MG63/vector cells at 24 h, respectively.
Conversely, in MG63-DDP cells, neutralization of miR-133b by the
introduction of anti-miR-133b mimics inhibited their migration and
invasion. Furthermore, the Transwell migration assay demonstrated
that the MG63/miR-133b mimics and the MG63-DDP/anti-miR-133b cells
traversed the matrix gel membrane further than the control groups
(Fig. 6).
Discussion
Cisplatin is one of the most commonly used agents in
chemotherapy due to its high efficiency, mild side effects and easy
administration (20). However, the
failure of cisplatin treatment is often observed due to
chemoresistance, which motivates the search for novel strategies to
enhance cell sensitivity to cisplatin (21). One strategy is to suppress the p38α
MAPK pathway that is responsible for desensitizing cancer cells to
cisplatin treatment (22). A previous
study by Wang et al (23)
demonstrated that downregulation of P28GANK gene expression
may sensitize osteosarcoma cells to cisplatin treatment via the
subsequent downregulation of multi-drug resistance gene 1 and
B-cell lymphoma 2 (23). A previous
study further confirmed that cisplatin resistance primarily
resulted from an increase in the enzymatic activity of glutathione
S-transferase P1 (24). The high
mobility group box 1 protein-mediated autophagy is a signaling
pathway that induces cisplatin-resistance in osteosarcoma cells
(25). An alternative strategy is to
stimulate the transcription start site pathway to enhance the
efficacy, which is directly associated with the enhancement of
chemotherapeutic effects (26).
miRNAs are known to be contributors to tumor
malignancy. Emerging evidence has revealed that miRNAs also serve
an important role in the induction of chemoresistance in
osteosarcoma (27). It has previously
been revealed that overexpression of miR-126 desensitizes
osteosarcoma cells to cisplatin by inhibiting apoptosis under
epigallocatechin-3-gallate treatment (28). Compared with adjacent tissues,
osteosarcoma cells demonstrated a higher expression level of
miR-33a, which promoted osteosarcoma cell resistance to cisplatin
by downregulating TWIST protein in vitro (29). miR-221 has previously been reported to
induce cell survival and cisplatin resistance via the
phosphoinositide-3 kinase/protein kinase B signaling pathway in
human osteosarcoma cells (30).
The present study demonstrated that overexpressing
miR-133b in human MG63 osteosarcoma cells enhanced their resistance
to cisplatin by inhibiting cell death induced by cisplatin,
migration and invasion. Upregulation of miR-133b resulted from
long-term cisplatin treatment. Taken together, these results
indicate that miR-133b serves an important role in
cisplatin-induced chemoresistance in osteosarcoma cells by
supporting tumor cell survival and cisplatin resistance. Previous
studies revealed that miR-133b is typically specifically expressed
in muscle tissue (31); it has also
been identified to be abnormally downregulated in numerous types of
cancer cells, which indicates its potential role in tumorigenesis
(32). By targeting the epidermal
growth factor receptor, the expression level of miR-133b negatively
regulates cell proliferation, migration and invasion in prostate
cancer cell lines (33). However, the
biological roles of miR-133b in induction of chemoresistance in
osteosarcoma are not yet clear. The present study demonstrated that
cisplatin treatment induced the upregulation of miR-133b. Similar
effects were observed when miR-133b mimics were delivered to MG63
cells during cisplatin treatment. Furthermore, the introduction of
anti-miR-133b mimics sensitized the MG63 cells to cisplatin, also
suggesting that miR-133b is sufficient for inducing cisplatin
resistance in MG63 cells.
In conclusion, miR-133b expression levels are
upregulated in human MG63 osteosarcoma cells following long-term
cisplatin treatment. Cisplatin treatment induces the overexpression
of miR-133b in MG63 cells, leading to the inhibition of
cisplatin-induced cell death, and promotion of migration and
invasion under cisplatin stress. The present study provided a novel
insight into the underlying mechanism of chemoresistance to
cisplatin, and miR-133b upregulation exhibits potential as a
biomarker of chemoresistance in osteosarcoma.
References
|
1
|
Fink-Puches R, Zenahlik P, Bäck B, Smolle
J, Kerl H and Cerroni L: Primary cutaneous lymphomas: Applicability
of current classification schemes (European Organization for
Research and Treatment of Cancer, World Health Organization) based
on clinicopathologic features observed in a large group of
patients. Blood. 99:800–805. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kager L, Zoubek A, Pötschger U, Kastner U,
Flege S, Kempf-Bielack B, Branscheid D, Kotz R, Salzer-Kuntschik M,
Winkelmann W, et al: Primary metastatic osteosarcoma: Presentation
and outcome of patients treated on neoadjuvant Cooperative
Osteosarcoma Study Group protocols. J Clin Oncol. 21:2011–2018.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bacci G, Briccoli A, Rocca M, Ferrari S,
Donati D, Longhi A, Bertoni F, Bacchini P, Giacomini S, Forni C, et
al: Neoadjuvant chemotherapy for osteosarcoma of the extremities
with metastases at presentation: Recent experience at the Rizzoli
Institute in 57 patients treated with cisplatin, doxorubicin, and a
high dose of methotrexate and ifosfamide. Ann Oncol. 14:1126–1134.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thomas M, Lieberman J and Lal A:
Desperately seeking microRNA targets. Nat Struct Mol Biol.
17:1169–1174. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sassen S, Miska EA and Caldas C: MicroRNA:
Implications for cancer. Virchows Arch. 452:1–10. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schetter AJ and Harris CC: Alterations of
microRNAs contribute to colon carcinogenesis. Semin Oncol.
38:734–742. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schetter AJ, Okayama H and Harris CC: The
role of microRNAs in colorectal cancer. Cancer J. 18:244–252. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hayes CN and Chayama K: MicroRNAs as
biomarkers for liver disease and hepatocellular carcinoma. Int J
Mol Sci. 17:2802016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vanas V, Haigl B, Stockhammer V and
Sutterlüty-Fall H: MicroRNA-21 increases proliferation and
cisplatin sensitivity of osteosarcoma-derived cells. PLoS One.
11:e01610232016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yuan J, Chen L, Chen X, Sun W and Zhou X:
Identification of serum microRNA-21 as a biomarker for
chemosensitivity and prognosis in human osteosarcoma. J Int Med
Res. 40:2090–2097. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yan K, Gao J, Yang T, Ma Q, Qiu X, Fan Q
and Ma B: MicroRNA-34a inhibits the proliferation and metastasis of
osteosarcoma both in vitro and in vivo. PLoS One. 7:e337782012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
He C, Xiong J, Xu X, Lu W, Liu L, Xiao D
and Wang D: Functional elucidation of miR-34 in osteosarcoma cells
and primary tumor samples. Biochem Biophys Res Commun. 388:35–40.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Duan Z, Choy E, Harmon D, Liu X, Susa M,
Mankin H and Hornicek F: MicroRNA-199a-3p is downregulated in human
osteosarcoma and regulates cell proliferation and migration. Mol
Cancer Ther. 10:1337–1345. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gougelet A, Pissaloux D, Besse A, Perez J,
Duc A, Dutour A, Blay JY and Alberti L: Micro-RNA profiles in
osteosarcoma as a predictive tool for ifosfamide response. Int J
Cancer. 129:680–690. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Song B, Wang Y, Xi Y, Kudo K, Bruheim S,
Botchkina GI, Gavin E, Wan Y, Formentini A, Kornmann M, et al:
Mechanism of chemoresistance mediated by miR-140 in human
osteosarcoma and colon cancer cells. Oncogene. 28:4065–4074. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Haas JD, Nistala K, Petermann F, Saran N,
Chennupati V, Schmitz S, Korn T, Wedderburn LR, Förster R, Krueger
A and Prinz I: Expression of miRNAs iR-133b and miR-206 in the
II17a/f locus is co-regulated with IL-17 production in αβ and γδ T
cells. PLoS One. 6:e201712011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Novello C, Pazzaglia L, Cingolani C, Conti
A, Quattrini I, Manara MC, Tognon M, Picci P and Benassi MS: miRNA
expression profile in human osteosarcoma: Role of miR-1 and
miR-133b in proliferation and cell cycle control. Int J Oncol.
42:667–675. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao H, Li M, Li L, Yang X, Lan G and
Zhang Y: miR-133b is down-regulated in human osteosarcoma and
inhibits osteosarcoma cells proliferation, migration, and invasion
and promotes apoptosis. PLoS One. 8:e835712013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shen DW, Pouliot LM, Hall MD and Gottesman
MM: Cisplatin resistance: A cellular self-defense mechanism
resulting from multiple epigenetic and genetic changes. Pharmacol
Rev. 64:706–721. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Grossi V, Peserico A, Tezil T and Simone
C: p38α MAPK pathway: A key factor in colorectal cancer therapy and
chemoresistance. World J Gastroenterol. 20:9744–9758. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang G, Rong J, Zhou Z and Duo J: Novel
gene P28GANK confers multidrug resistance by modulating the
expression of MDR-1, Bcl-2 and Bax in osteosarcoma cells. Mol Biol
(Mosk). 44:1010–1017. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pasello M, Michelacci F, Scionti I,
Hattinger CM, Zuntini M, Caccuri AM, Scotlandi K, Picci P and Serra
M: Overcoming glutathione S-transferase P1-related cisplatin
resistance in osteosarcoma. Cancer Res. 68:6661–6668. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang J, Liu K, Yu Y, Xie M, Kang R,
Vernon P, Cao L, Tang D and Ni J: Targeting HMGB1-mediated
autophagy as a novel therapeutic strategy for osteosarcoma.
Autophagy. 8:275–277. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dai H, Huang Y, Li Y, Meng G, Wang Y and
Guo QN: TSSC3 overexpression associates with growth inhibition,
apoptosis induction and enhances chemotherapeutic effects in human
osteosarcoma. Carcinogenesis. 33:30–40. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu X, Wells A, Padilla MT, Kato K, Kim KC
and Lin Y: Asignaling pathway consisting of miR-551b, catalase and
MUC1 contributes to acquired apoptosis resistance and
chemoresistance. Carcinogenesis. 35:2457–2466. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiang L, Tao C, He A and He X:
Overexpression of miR-126 sensitizes osteosarcoma cells to
apoptosis induced by epigallocatechin-3-gallate. World J Surg
Oncol. 12:3832014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou Y, Huang Z, Wu S, Zang X, Liu M and
Shi J: miR-33a is up-regulated in chemoresistant osteosarcoma and
promotes osteosarcoma cell resistance to cisplatin by
down-regulatin TWIST. J Exp Clin Cancer Res. 33:122014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao G, Cai C, Yang T, Qiu X, Liao B, Li
W, Ji Z, Zhao J, Zhao H, Guo M, et al: MicroRNA-221 induces cell
survival and cisplatin resistance through P13 K/Akt pathway in
human osteosarcoma. PLoS One. 8:e539062013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen JF, Mandel EM, Thomson JM, Wu Q,
Callis TE, Hammond SM, Conlon FL and Wang DZ: The role of
microRNA-1 and microRNA-133 in skeletal muscle proliferation and
differentiation. Nat Genet. 38:228–233. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen X, Wu B, Xu Z, Li S, Tan S, Liu X and
Wang K: Downregulation of miR-133b predict progression and poor
prognosis in patients with urothelial carcinoma of bladder. Cancer
Med. 5:1856–1862. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tao J, Wu D, Xu B, Qian W, Li P, Lu Q, Yin
C and Zhang W: microRNA-133 inhibits cell proliferation, migration
and invasion in prostate cancer cells by targeting the epidermal
growth factor receptor. Oncol Rep. 27:1967–1975. 2012.PubMed/NCBI
|