Introduction
Much evidence demonstrates dysregulation in the
PI3K/AKT signaling pathway in colorectal cancer (CRC) (1). It seems that in some cases activation of
AKT downstream is an initial step in the development of CRC. On the
other hand, PI3K/AKT activation is found in more advanced stages of
CRC carcinogenesis, which in many cases is caused by PTEN protein
loss or downregulation.
Phosphatase and tensin homolog deleted on chromosome
ten (PTEN) is an important suppressor gene. Protein product of the
PTEN gene is a dual-specific phosphatase which plays a crucial role
in the signal transduction from the membrane receptors to the
intracellular downstream cascades. Its role in the development of
many cancers was doubtlessly confirmed with a mechanism of action
via the catalysis of phosphate detachment from (3–5)-tri-phospho-inositol (PIP3) in the
PI3K/AKT signaling pathway (2,3).
In our previous studies we demonstrated that PTEN
activity may be diminished as early as in the phase of an
intestinal glandular polyp and is frequently observed in CRC
(4,5).
These observations justify the following questions: How much the
loss of PTEN activates AKT signaling and how it is reflected in the
levels and states of its signaling protein members.
This problem is very important in the context of
cytotoxic therapy in CRC which is administered concurrently with
directed therapy aimed to compensate molecular disturbances in the
activity of the PI3K/AKT/PTEN pathway. Identification of the
characteristic pattern of signaling protein expression and
phosphorylation states could be a predictive marker for target
therapies based on specific signaling kinase inhibitors. The
simultaneous quantitative or at least semi-quantitative assessment
of many proteins with preservation of information about their
spatial location is within the interest of the system biology.
Unfortunately, it suffers from a lack of appropriate methodology to
achieve these goals. Some hope with severe restrictions is placed
on immunofluorescence techniques. However, it is possible to
visualize only 3–4 proteins with these methods. Significantly
greater possibilities are connected with the use of antibodies
conjugated with semiconducting quantum dots (QDs) which are within
the interest of nanotechnology.
Quantum dots are semiconducting nanocrystals sized
2–10 nm which became a promising research, diagnostic and possibly
therapeutic tool due to their unique possibilities of light
absorption and emission in the process known as the quantum
confinement effect. By changing the composition and/or the size of
QDs emission spectrum may be freely modified starting from
ultraviolet (UV) to infrared (IR). Simultaneously QDs are
characterized by a very broad absorption spectrum and a narrow
emission spectrum, which allows synchronic excitation of several
QDs by the light of the same wavelength. Due to all these
properties, QDs could be used in the precise detection of a
plethora of biomolecules such as nucleic acids, carbohydrates,
enzymes and antibodies (6–8).
The aim of the present study was to visualize
dysregulation of signaling protein members of PI3K/AKT/PTEN pathway
in CRC in response to the loss of activity of its major suppressor
that is PTEN using QD-conjugated antibodies. In the future
perspective the presented method could be utilized in visualization
of subtle changes of concentrations and activation states of
crucial regulatory proteins in other signaling cascades which are
important in oncogenesis.
Materials and methods
Study group
A total of fifty patients diagnosed with
adenocarcinoma of the large intestine after major surgery were
enrolled in the study. Clinico-pathological characteristics of the
study group are presented in Table I.
Immediately after the excision intestinal biopsy samples with
tumors were fixed in neutral buffered formalin with the addition of
commercially-available pan-phosphatase inhibitor PhosSTOP
(Phosphatase Inhibitor Coctail tablets; Roche, Mannheim, Germany).
Preparations were dissected longitudinally to ensure quick
penetration of the fixative to the tumor tissues. During the
grossing process the representative tumor samples were taken by the
pathologist then dehydrated and embedded in paraffin.
 | Table I.Clinicopathological characteristics of
the study group. |
Table I.
Clinicopathological characteristics of
the study group.
| Feature | Value |
|---|
| Age, years | 67 (range,
42–85) |
| Sex |
|
| Male | 27 |
|
Female | 23 |
| Location |
|
|
Colon | 32 |
|
Rectum | 18 |
| Histologic type |
|
|
Adenocarcinoma | 55 |
|
Mucinosum | 5 |
| Dysplasia grade |
|
| G1 | 13 |
| G2 | 29 |
| G3 | 8 |
| TNM staging |
|
| I | 14 |
| II | 17 |
| III | 9 |
| IV | 10 |
Preparing QD-conjugated
antibodies
During QD-conjugation of antibodies
commercially-available reagent kits were used (QDots Antibody
Labeling kit, Molecular Probes; Invitrogen, Carlsbad, CA, USA).
Quantum dots with maxima of emission located at 525, 565 and 585 nm
were utilized in the present study. The procedure of antibody
labeling was carried out according to the manufacturer's
instructions and consisted of the following five steps: i) Antibody
concentration and buffer exchange in order to eliminate other
proteins as well as azide ions used as an antibody preservative,
ii) modification of the carbohydrate domain of the antibody using
galactosidase enzyme, iii) azide domain connection to the modified
antibody, iv) purification of the activated antibody amd v)
conjugation of the activated antibody with QD molecule using
dibenzo-cyclo-octane (DIBO) residue as an interconnector.
Final concentration of the conjugated antibodies was
estimated to be about 1 mg/ml according to the manufacturer's
specifications assuming 2 µmol/l molarity of the obtained conjugate
(9).
Immunofluorescence
Formalin-fixed paraffin-embedded tumor samples were
cut with a microtome into 5 µm-thick histological sections and
fixed on microscope subject glasses. The slides were rehydrated in
xylene, graded alcohols and finally in phosphate-buffered saline
(PBS). Next the process of antigen retrieval was carried out in
citric buffer for 30 min in a microwave oven. Finally, the remains
of citric buffers were washed out using PBS and the slides were
exposed to QD-labeled antibodies. In order to demonstrate the
concentration and spatial location of subject proteins (PTEN,
phosphorylated PDK1 and partially-phosphorylated AKT) specific
QD-labeled monoclonal antibodies were used. The brief
characteristics of the used antibodies and their concentrations are
presented in the Table II.
| Table II.Summary and the characteristics of
primary antibodies used in the study. |
Table II.
Summary and the characteristics of
primary antibodies used in the study.
| Target | Antibody
type/clone | Host | Antibody
dilution | Manufacturer |
|---|
| PTEN (pan) | Monoclonal 138G6 | Rabbit IgG | 1:500 | Cell Signaling
Technology, Inc. |
| Phospho-Akt
Thr308 | Monoclonal D52E6 | Rabbit IgG | 1:1,500 | Cell Signaling
Technology, Inc., no. 13038 |
| PhosphoPDK1
Ser241 | Monoclonal C49H2 | Rabbit IgG | 1:1,000 | Cell Signaling
Technology, Inc., no. 3438 |
Incubation of the tumor sections with primary
antibodies was performed for 12 h in a humidified chamber in the
fridge at the temperature of 6°C. After the incubation antibody
solutions were washed out with PBS and the preparations were
mounted with fluorescence-preserving mounting medium dedicated to
QDs (Qmount Qdot Mounting Medium; Invitrogen).
Fluorescence acquisition and
fluorescence signal assessment
Visualization of the fluorescence signal of the
QD-labeled antibodies was performed with epi-fluorescence
microscope (AxioImager M.2; Carl Zeiss, Jena, Germany) equipped
with a set of excitation/emission filters with spectral
characteristics compatible with used QDs band-pass filters with the
transmission bandwidth of 20 nm (Chroma, Augsburg, Germany),
Plan-apochromat objectives with magnification of 63× NA=0.95 and
100× NA=1.3 oil immersion as well as AxioCam MRm 1.4 Mega pixel
monochrome camera (Carl Zeiss).
Fluorescence signal measurement was made in a
region-based manner. At least 20 manually selected points
corresponding to submembrane or cytoplasmic regions of individual
neoplastic cells were sampled and averaged for the particular
region and case. As a measure of antibody concentrations the mean
pixel values ranging from 0 (no fluorescence) to 255 (maximal
fluorescence signal strength) was reported and included in the
further analysis.
Statistical analysis
Statistical analysis of the results was performed
using Statistica software v8.0, (Statsoft Inc., Palo Alto, CA,
USA). The comparisons of the mean values of the analyzed parameters
were performed using Student's t-test. Interrelationships between
fluorescence signals were investigated by calculating of Pearson's
correlation coefficient (R). P<0.05 was considered to indicate a
statistically significant difference.
Ethical approval
The present study was conducted in accordance with
the guidelines of the Declaration of Helsinki and its subsequent
amendments, and informed consent was obtained from all patients.
Project of the study was approved by institutional review board of
the Medical University of Silesia, Katowice (approval no.
KNW/0022KBI/178/09).
Results
In the cases with a complete loss or a diminished
expression of the PTEN protein (defined as pixel values of PTEN
immunofluorescence less than median) fluorescence signals
corresponding to phospho-PDK1 (Ser241) and phospho-AKT (Thr308)
were higher than in the group with normal PTEN expression (above
median; t-Student test P=0.002 and P=0.01 respectively). Maxima of
fluorescence signals corresponding to phospho-PDK1 and phospho-AKT
were localized in a submembrane cellular compartment and were
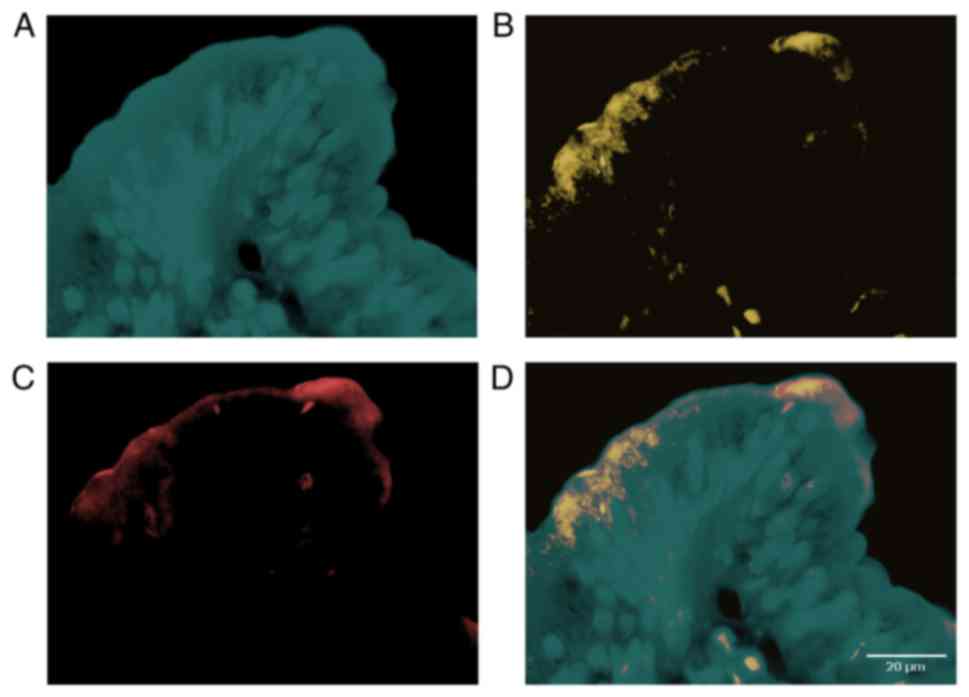
colocalized with each other (Fig. 1).
We also found a positive correlation between fluorescence signals
corresponding to phospho-PDK1 and phospho-AKT (R=0.65 P=0.047) as
well as a negative correlation between PTEN signal and the
above-mentioned signaling proteins (R=−0.47, P=0.05; and R=−0.51,
P=0.055, respectively) in the group of all the examined
samples.
The summary of the obtained results is presented in
Table III.
| Table III.Fluorescent signal strength of the
analyzed signaling proteins in relation to the expression of the
PTEN protein. |
Table III.
Fluorescent signal strength of the
analyzed signaling proteins in relation to the expression of the
PTEN protein.
|
| PTEN expression |
|---|
|
|
|
|---|
| Marker | Negative (n=26) | Positive (n=24) |
|---|
| PTEN (pan) | Mean=156.29
(49–250)a |
|
| MarkerMedian=167 |
|
| SD=57.1 |
|
| Mean=108.08
(49–167) | Mean=206.5
(173–250) |
|
| Median=105 | Median=211.5 |
|
| SD=33.58 | SD=21.68 |
| phospho-AKT
(Thr308) | Mean=75.92
(28–153) | Mean=49.04
(18–117) |
|
| SD=27.98 | SD=29.16 |
| phospho-PDK1
(Ser241) | Mean=74.16
(43–107) | Mean=61.00
(34–101) |
|
| SD=16.47 | SD=18.25 |
Discussion
A dynamic development of imaging techniques
utilizing QDs in medicine and biological sciences has been observed
since the end of the 20th century, especially from the moment of
the first synthesis of water-soluble QDs with biologically-active
coating molecules. Since then QDs with adequately modified surface
have been used in labeling of cell surface and intracellular
structures in live cells or fixed specimens. Unique features of QDs
connected with wideband excitation spectra and a narrow emission of
excited light allowed to use their functionalized conjugates in
detection and visualization of virtually all biomolecules, cells
and tissues both in vivo and in vitro. In medicine
QDs are seen as modern markers which could be used to visualize and
track spreading of neoplastic cells or viruses in the human body
(7,10,11).
Quantum dots also possess a therapeutic potential in oncology
(12,13). It seems that QDs are more precise and
stable markers than widely used organic chromogens (6).
By increasing the diameter of the core of the QD the
wavelength of emitted light can be increased. Based on this
phenomenon, a broad range of QDs with different spectral
characteristics can be easily synthesized in one technological
process. Using a single monochromatic excitation source all of
these QDs can be excited producing fluorescence light from UV to IR
in narrow, non-overlapping bands eliminating the problem of
cross-talk between channels (6).
In life sciences the following are used in
fluorescence microscopy: Cadmium selenide (core) and cadmium
sulfide (shell) QDs (CdSe/ZnS) with emission between 450–650 nm.
The second type of QDs with emission of longer wavelengths between
500 and 750 nm are cadmium telluride QDs (CdTe) (6,8). Broad
absorption spectra of QDs allow at the same time to effectively
excite them using single light source emitting in the range of UV,
violet or blue light equipped with a short-pass excitation filter.
Simultaneously, a narrow emission spectrum with almost arbitrary
shaping of its width and placement within both visible and IR light
allows creating many non-overlapping acquisition channels with the
aid of standard single-band interference emission filters with a
bandwidth of 20 or 40 nm. In the present experiment only three
simultaneous channels were used, however it confirms the concept in
the field of bioimaging of protein concentration and activation
state with preserving information about their spatial location in
single cells.
Many cancers possess elevated levels of
PtdIns(3,4,5)P(3), the second messenger that induces
activation of the protein kinase PKB/AKT and thereby stimulates
cell proliferation, growth, and survival. The importance of this
pathway in tumorigenesis has been highlighted by the finding that
PTEN, the lipid phosphatase that breaks down PtdIns(3,4,5)P(3) to
PtdIns(4,5)P(2), is
frequently mutated in human cancer. Cells lacking PTEN possess
elevated levels of PtdIns(3,4,5)P(3) correlated with PKB activation (14,15).
3-Phosphoinositide-dependent kinase 1 (PDK1) is the
first node of the PI3K/AKT/PTEN signal cascade and is required for
activation of AKT. PIP(3) recruits
PDK1 and AKT to the cell membrane through interactions with their
pleckstrin homology domains, allowing PDK1 to activate AKT by
phosphorylating it at residue threonine-308 (16).
It was demonstrated in breast cancer patients that
increased PDK1 copy number is associated with upstream pathway
lesions (ERBB2 amplification, PTEN loss, or PIK3CA mutation), as
well as patient survival. The examination of an independent set of
breast cancers and tumor cell lines derived from multiple forms of
human cancers also found increased PDK1 protein levels associated
with such upstream pathway lesions. In human mammary cells, PDK1
enhanced the ability of upstream lesions to signal to AKT,
stimulate cell growth and migration, and rendered cells more
resistant to PDK1 and PI3K inhibition (17).
The role of PTEN in the CRC pathology has been
postulated by many authors. The loss of PTEN expression is
postulated to be an early event in CRC carcinogenesis which is
described as early as in precancerous lesions-large intestine
adenomatous polyps (adenomas) (4) and
its frequency increases in more advanced stages (5). An association between PTEN status and
histological grade, tumor size and clinical outcome was also
demonstrated by other authors (18–20).
In our present experiment we tried to assess the
functional state of the top-most elements of PI3K/AKT/PTEN pathway
in CRC. They include gate-keeping, pacemaker gene PTEN and its two
effector proteins PDK1 and AKT.
In the analyzed samples of large intestine
adenocarcinoma we found that a lower concentration of PTEN in the
cytoplasm neighboring the cell membrane correlates with the
accumulation of activated by phosphorylation forms of downstream
signaling proteins: Phospho-PDK1-Ser241 and phospho-AKT-Thr308.
This observation is indirect evidence of tri-phosphatydyl-inositol
(PIP3) concentration due to the loss of phosphatase activity of
PTEN in cancer cells described by other authors as separate
observations. To our best knowledge, this is the first report
demonstrating the above-described changes concomitantly in the same
cellular compartments of cancer cells as a response to the loss of
PTEN activity.
In patients with a diminished or complete loss of
the PTEN expression in the tumor tissue we found increased levels
of activated/phosphorylated forms of PDK1 (Phospho-PDK1-Ser241) and
AKT (Phospho-AKT-Thr308) proteins which are responsible for a
permanent activation of the PI3K/AKT/PTEN pathway in some cases of
CRC.
Application of QDs in fluorescence bioimaging is a
more flexible and precise method in comparison to the traditionally
utilized organic chromophores. Due to their unique properties QDs
form a new class of optical markers successfully applied in life
sciences imaging with specific antibodies allowing simultaneous
visualization of location, concentration and activity states of
specified signaling proteins in cellular compartments.
References
|
1
|
Davies EJ, Marsh Durban V, Meniel V,
Williams GT and Clarke AR: PTEN loss and KRAS activation leads to
the formation of serrated adenomas and metastatic carcinoma in the
mouse intestine. J Pathol. 233:27–38. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carnero A, Blanco-Aparicio C, Renner O,
Link W and Leal JF: The PTEN/PI3K/AKT signalling pathway in cancer,
therapeutic implications. Curr Cancer Drug Targets. 8:187–198.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Leslie NR and Downes CP: PTEN function:
How normal cells control it and tumour cells lose it. Biochem J.
382:1–11. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Waniczek D, Śnietura M, Młynarczyk-Liszka
J, Pigłowski W, Kopeć A, Lange D, Rudzki M and Arendt J: PTEN
expression profiles in colorectal adenocarcinoma and its
precancerous lesions. Pol J Pathol. 64:15–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Waniczek D, Snietura M, Kopec A,
Scieglinska D, Piglowski W, Lorenc Z, Muc-Wierzgon M and
Nowakowska-Zajdel E: A novel quantitative method of pten expression
assessment in tumor tissue. J Biol Regul Homeost Agents. 30:79–90.
2016.PubMed/NCBI
|
|
6
|
Smith AM, Duan H, Mohs AM and Nie S:
Bioconjugated quantum dots for in vivo molecular and cellular
imaging. Adv Drug Deliv Rev. 60:1226–1240. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fang M, Peng CW, Pang DW and Li Y: Quantum
dots for cancer research: Current status, remaining issues, and
future perspectives. Cancer Biol Med. 9:151–163. 2012.PubMed/NCBI
|
|
8
|
Iga AM, Robertson JH, Winslet MC and
Seifalian AM: Clinical potential of quantum dots. J Biomed
Biotechnol. 10:760872007.
|
|
9
|
Molecular Probes Inc., . Separating the
conjugated antibody from unconjugated antibodyQdot antibody
conjugation kits. Molecular Probes, Inc., . pp. 7–8. 2008
|
|
10
|
Wang Y and Chen L: Quantum dots, lighting
up the research and development of nanomedicine. Nanomedicine.
7:385–402. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu SL, Wang ZG, Zhang ZL and Pang DW:
Tracking single viruses infecting their host cells using quantum
dots. Chem Soc Rev. 45:1211–1224. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tholouli E, Sweeney E, Barrow E, Clay V,
Hoyland JA and Byers RJ: Quantum dots light up pathology. J Pathol.
216:275–285. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao J, Qiu X, Wang Z, Pan J, Chen J and
Han J: Application of quantum dots as vectors in targeted survivin
gene siRNA delivery. Onco Targets Ther. 3:303–309. 2013. View Article : Google Scholar
|
|
14
|
Bayascas JR, Leslie NR, Parsons R, Fleming
S and Alessi DR: Hypomorphic mutation of PDK1 suppresses
tumorigenesis in PTEN(+/-) mice. Curr Biol. 15:1839–1846. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Haas-Kogan D, Shalev N, Wong M, Mills G,
Yount G and Stokoe D: Protein kinase B (PKB/Akt) activity is
elevated in glioblastoma cells due to mutation of the tumor
suppressor PTEN/MMAC. Curr Biol. 8:1195–1198. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bayascas JR: Dissecting the role of the
3-phosphoinositide-dependent protein kinase-1 (PDK1) signalling
pathways. Cell Cycle. 7:2978–2982. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Maurer M, Su T, Saal LH, Koujak S, Hopkins
BD, Barkley CR, Wu J, Nandula S, Dutta B, Xie Y, et al:
3-Phosphoinositide-dependent kinase 1 potentiates upstream lesions
on the phosphatidylinositol 3-kinase pathway in breast carcinoma.
Cancer Res. 69:6299–6306. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yazdani Y, Farazmandfar T, Azadeh H and
Zekavatian Z: The prognostic effect of PTEN expression status in
colorectal cancer development and evaluation of factors affecting
it: miR-21 and promoter methylation. J Biomed Sci. 23:92016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Colakoglu T, Yildirim S, Kayaselcuk F,
Nursal TZ, Ezer A, Noyan T, Karakayali H and Haberal M:
Clinicopathological significance of PTEN loss and the
phosphoinositide 3-kinase/Akt pathway in sporadic colorectal
neoplasms: Is PTEN loss predictor of local recurrence? Am J Surg.
195:719–725. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hsu CP, Kao TY, Chang WL, Nieh S, Wang HL
and Chung YC: Clinical significance of tumor suppressor PTEN in
colorectal carcinoma. Eur J Surg Oncol. 37:140–147. 2011.
View Article : Google Scholar : PubMed/NCBI
|