Introduction
The 14-3-3 protein family are highly conserved
soluble acidic proteins (1). There
are 7 isoforms (including β, ε, η, γ, τ, ζ and σ) in the 14-3-3
family, with molecular masses ranging between 28 and 33 kDa
(1). The 14-3-3 proteins may function
as adaptor molecules or scaffolds of protein-protein interactions,
or act as localization anchors, controlling the subcellular
localization of proteins, or as enzyme activity regulators,
increasing or suppressing their catalytic activity (2). As a member of the 14-3-3 protein family,
14-3-3ε may integrate a number of signaling pathways by interacting
with distinct proteins; therefore, 14-3-3ε serves functions in the
regulation of the cell cycle (3,4) and
apoptosis (5–8). Previously, 14-3-3ε has been reported to
bind with heat shock factor 1 (HSF1) through the extracellular
signal regulated kinase/glycogen synthase kinase 3/14-3-3ε pathway,
thus suppressing HSF1 activity during cellular proliferation
(9). Zhong et al (6) demonstrated that inhibiting the
interaction of 14-3-3ε with chloride intracellular channel protein
4 enhanced autophagy and contributed to mitochondrial and ER
stress-induced apoptosis under starvation conditions. In addition,
14-3-3ε has an association with the occurrence and development of a
number of types of tumor; for example, the expression of 14-3-3ε
has been identified to be downregulated in gastric cancer (10) and pediatric astrocytoma (11), whereas it is upregulated in
hepatocellular carcinoma (12,13), renal
cancer (14) and breast cancer
(15).
Previous studies have revealed that certain isoforms
of 14-3-3 protein family may inhibit cell apoptosis (16–20). In
the present study, it was demonstrated that 14-3-3ε was a
differentially expressed nuclear matrix component in MG-63 cells
during apoptosis. The nuclear matrix is composed of the nuclear
lamina, the nucleolar remnants and an internal nuclear mesh of
fibers of an unknown constitution that it is possible to visualize
by electron microscopy (21).
Alteration of nuclear matrix protein composition was demonstrated
to be involved in the apoptotic process (22). The present study revealed that 14-3-3ε
was an altered nuclear matrix protein, suggesting that 14-3-3ε may
be an important regulator in curcumin induced apoptosis. However,
the regulatory mechanism of 14-3-3ε in cell carcinogenesis and
apoptosis remains unknown. In the present study, further
investigations were performed on the expression and localization of
14-3-3ε and its co-localization with apoptosis-associated proteins
in order to improve the current understanding of 14-3-3ε function
in the regulation of cellular carcinogenesis and apoptosis.
Materials and methods
Materials
Human osteosarcoma MG-63 cells were purchased from
Xiangya Central Experiment Laboratory (Changsha, China). Curcumin
standards were purchased from Fuzhou Min Jian Medicine Inspection
Service (Fuzhou, China).
Cell culture and induction of
apoptosis
Human osteosarcoma MG-63 cells were cultured in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific Inc., Waltham,
MA, USA) containing 10% fetal bovine serum (Zheijiang Tianhang
Biotechnology Co., Inc., Hangzhou, China), 100 U/ml penicillin and
100 U/ml streptomycin, at 37°C in a humidified atmosphere
containing 5% CO2. Induction of apoptosis was performed
following a method described previously (22). Cells in the exponential phase were
digested with 0.25% trypsin (Xiamen Lulong Biotech Development Co.,
Inc., Xiamen, China) and transferred to new plates. After 24 h, the
medium was replaced with fresh RPMI-1640 medium with or without 7.5
µg/ml curcumin. Cells were incubated with curcumin at 37°C for 0–48
h and collected at the corresponding time.
Selective extraction of nuclear matrix
proteins (NMPs)
NMPs were extracted following a modified method
described previously by Michishita et al (23). Selected cells were washed with PBS
three times. Subsequently, cells were resuspended with cytoskeleton
extraction buffer CSK100 [10 mM Pipes, pH 6.8, 300 mM sucrose, 100
mM NaCl, 3 mM MgCl2, 1 mM EGTA, 1 mM
phenylmethylsulfonyl fluoride (PMSF) and 0.5% Triton X-100] and
incubated on ice for 10 min. Lysates were centrifuged at 900 × g
for 5 min at 4°C, and the resulting supernatant was discarded. The
precipitate was washed with cytoskeleton extraction buffer CSK50
(10 mM Pipes, pH 6.8, 300 mM sucrose, 50 mM NaCl, 3 mM
MgCl2, 1 mM EGTA, 1 mM PMSF and 0.5% Triton X-100)
twice. CSK50 containing 400 U/ml DNase I (Sangon Biotech Co., Ltd.,
Shanghai, China) was added to the precipitate prior to incubation
at room temperature for 30 min. Subsequently, 1 M
(NH4)2SO4 was added dropwise to the tube to a
final concentration of 0.25 M. After 15 min of incubation at room
temperature, NMPs were obtained by centrifugation at 900 × g for 5
min at 4°C. NMPs were washed with CSK50 once. The supernatant was
removed and the proteins were stored at −80°C until use.
Two-dimensional (2D) gel
electrophoresis and mass spectrometry analysis
NMPs were re-dissolved in 2D sample buffer which
containing 7 M urea, 2 M Tthiourea, 4% CHAPS, 40 mM Tris-base, 2%
pharmalyte (pH 3–10), 40 mM DTT, 1 mM PMSF, 1 mM EDTA. The 2D PAGE
process was performed following a previously described method
(24). For first dimension
separation, immobilized pH gradient (IPG) strips (11 cm, pH 4–7)
were used and a total of 350 µg proteins were loaded, followed by
isoelectric focusing electrophoresis according to the optimized
procedure. IPG strips were rehydrated with the sample buffer
containing 350 µg protein, and were then subjected to isoelectric
focusing (IEF) using an Ettan IPGphor 3 IEF System (GE Healthcare,
Chicago, IL, USA). The focus procedure was as follows: Step 1, 100
V slow ramp for 30 min; step 2, 200 V slow ramp for 30 min; step 3,
500 V slow ramp for 30 min; step 4, 1,000 V rapid ramp for 1 h;
step 5, 8,000 V slow ramp for 4 h; step 6, 8,000 V rapid ramp for
40,000 V/h; step 7, a hold at 500 V. Following IEF, IPG strips were
equilibrated in 6 M urea, 2% SDS, 50 mM Tris-HCl, pH 8.8, 20% v/v
glycerol, and 2% DTT for 15 min and then, for an additional 15 min,
placed in the same buffer but with DTT replaced by 2.5%
iodoacetamide. The strips were then placed on 10% SDS-PAGE gels.
Electrophoresis was performed at 10 mA/gel at 4°C overnight,
followed by silver staining as previously described (25).
Gels were scanning using the GE ImageScanner III (GE
Healthcare) and analyzed using PDQuest 8.0 software (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). When spots were detected in
PDQuest, the original gel image was filtered and smoothed to
clarify the spots, then three-dimensional Gaussian spots were
created from the clarified spots. Spot quantity was defined as the
total intensity of a defined spot in a gel image. The formula for
calculating the quantity of Gaussian spots in PDQuest was as
follows: Spot quantity=spot height × π × σx × σy. Spot height is
the peak intensity of the Gaussian representation of the spot,
measured in terms of optical density (OD). σx and σy are the
standard deviation of the Gaussian distribution of the spot in the
direction of the x- and y-axis, respectively, measured in IU (1
IU=100 µM). Protein spots with a fold change >2 were defined as
differential protein spots and subjected to matrix assisted laser
desorption ionization-time of flight mass spectrometry
(MALDI-TOF-MS) analysis.
Gel particles of differential protein spots were
excised and subsequently destained with a solution containing 50%
acetonitrile and 50 mM ammonium bicarbonate at room temperature
several times, until the gel particles were colorless. The gel
particles were dehydrated in 100% acetonitrile, followed with
centrifugation under vacuum at 400 × g for 30 min at 42°C.
Dehydrated particles were digested with trypsin (Promega
Corporation, Madison, WI, USA) at 37°C for 16 h. The peptide
fragments were extracted using 50% acetonitrile containing 5%
trifluoroacetic acid (TFA). Samples were then dehydrated by
centrifugation under vacuum at 400 × g for 30 min at 42°C.
Extracted peptide fragments were dissolved in 1 µl
0.1% TFA and then blended with an equal volume of
a-cyano-4-hydroxycinnamic acid (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). Samples were then spotted on a target plate.
Following air-drying at room temperature, samples were analyzed
using MALDI-TOF-MS (Reflex III MALDI-TOF; Bruker Daltonics,
Leipzig, Germany). A 337 nm nitrogen laser was used to irradiate
and ionize the samples with the pulse width set at 3 ns. The source
1 and source 2 accelerating voltage were set at 20 and 23 kV,
respectively. The delay extraction time was 2,000 ns. The
mass-to-charge ratios (m/z) were determined by the time of ions
flying in the flight tube, and the peptide mass fingerprinting
(PMF) was obtained.
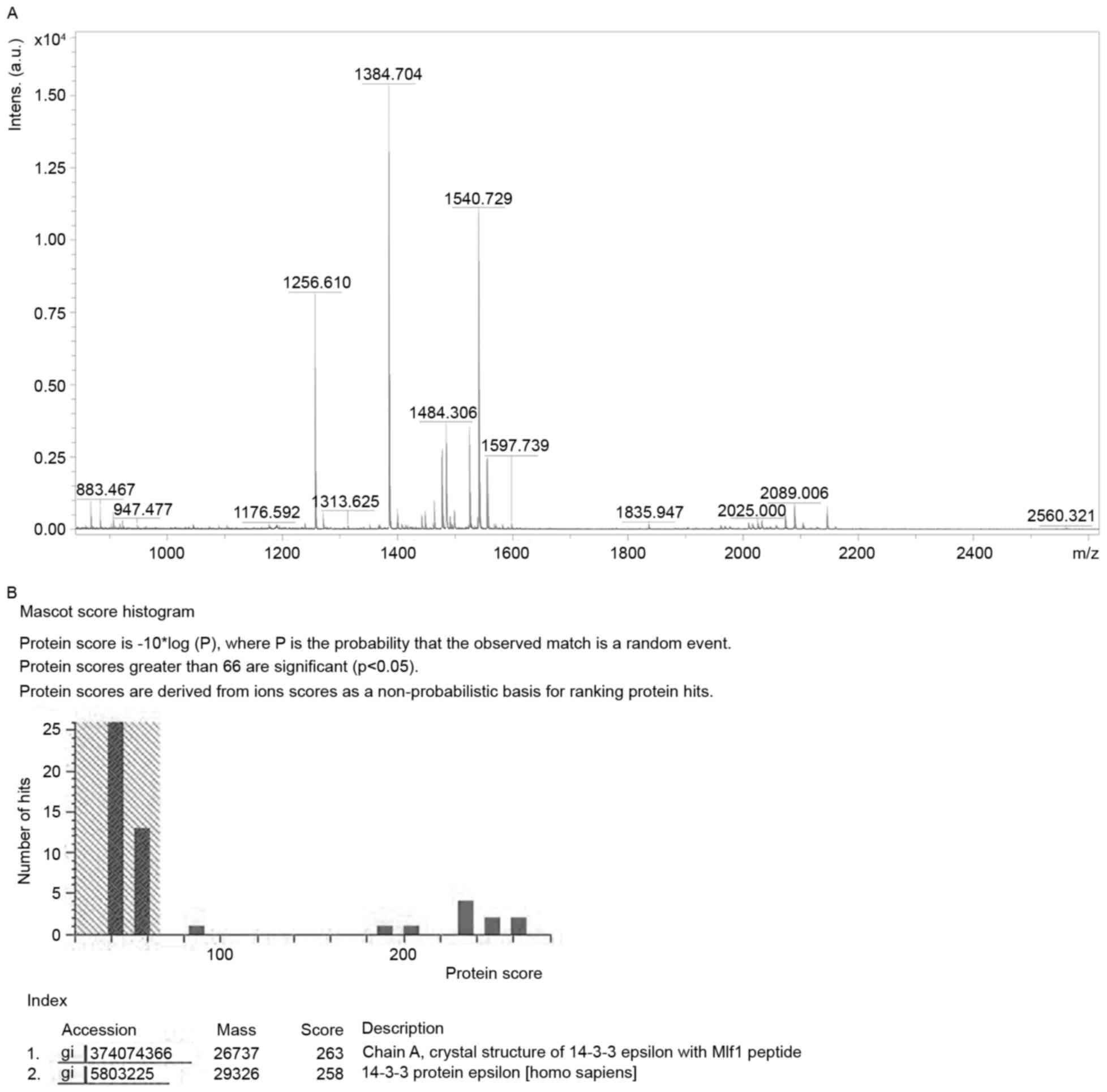
PMF was interpreted and quantified by FlexAnalysis
software 2.0 (Bruker Corporation, Ettlinger, Germany) following
internal calibration with the trypsin autolysis products [842.51
(M+H) and 2211.11 (M+H)]. The data were mapped to corresponding
proteins by searching against the SWISS-PROT (26) and National Center for Biotechnology
Information databases (27) using the
MASCOT search engine (www.matrixscience.com). Up to one missed cleavage was
allowed for each peptide. Fixed modification was carbamidomethyl.
Variable modification was oxidation. The max peptide tolerance was
±200 ppm. MH+ and monoisotopic were selected for mass
values. Protein scores >66 were deemed significant
(P<0.05).
Western blot analysis
Cells were lysed using radioimmunoprecipitation
assay buffer containing 50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1%
Triton X-100, 0.5% SDS, 0.5% sodium deoxycholate, 1 mM PMSF and 1X
protease inhibitor cocktail (S8830; Sigma-Aldrich; Merck KGaA) at
4°C for 30 min. Subsequently, the lysate was centrifuged at 12,000
× g for 15 min at 4°C. The Bradford assay was used to determine
protein concentrations and, subsequently, the proteins (20 µg) were
separated using 12% SDS-PAGE, and transferred onto a polyvinylidene
difluoride (PVDF) membrane. The membrane was then blocked in 1X TBS
containing 5% non-fat milk at room temperature for 1 h. The PVDF
membrane was then incubated with primary antibodies against 14-3-3ε
(cat. no. 11648-2-AP; ProteinTech Group, Inc., Chicago, IL, USA;
cat. no. sc-393177; Santa Cruz Biotechnology, Inc., Dallas, TX,
USA) or β-actin (cat. no. 60008-1-Ig; ProteinTech Group, Inc.) at a
dilution of 1:1,000 in 1X TBST (20 mM Tris-HCl, pH 7.4, 150 mM
NaCl, 0.2% Tween20) at 4°C overnight. Following washing with 1X
TBST 3 times (7 min per wash), the membrane was incubated with
horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG (cat.
no. Pierce-31460; Thermo Fisher Scientific Inc.) or HRP-conjugated
goat anti-mouse IgG (Pierce-31430; Thermo Fisher Scientific Inc.;
diluted in 1X TBST at a dilution of 1:5,000) at room temperature
for 1 h. The signals were detected using WesternBright™ enhanced
chemiluminescence kit (cat. no. K-12045-D50; Advansta, Inc, Menlo
Park, CA, USA). The results were analyzed using Quantity One
software (version 4.6.2, Bio-Rad Laboratories, Inc.).
Selective extraction of nuclear matrix
intermediate fiber system and sample preparation
MG-63 cells were seeded on a 6-cm culture dish with
a 13-mm coverslip in it. For the control group, ~4×103
cells were seeded. For the curcumin-treated group,
~1×104 cells were seeded, as cell growth was suppressed
and certain cells died or detached during curcumin treatment.
Following treatment for 36 h, coverslips with seeded MG-63 cells
were washed twice with warm PBS (37°C). Subsequently, the cells
were selectively extracted as previously described (28). Following selective extraction, the
samples were fixed in 4% paraformaldehyde at 4°C for 20 min. For
observation using light microscopy, samples were stained with
Coomassie brilliant blue at room temperature for 30 min, followed
by washing with double-distilled water several times. The samples
were then observed by light microscopy under ×400 magnification.
For observation using fluorescence microscopy, samples were
subjected to immunofluorescent staining as subsequently
described.
Immunofluorescent staining (IF)
MG-63 cells were seeded on a 6-well culture plate
with a 13-mm coverslip in each well. For the control group,
~2×103 cells were seeded. For the curcumin treated
group, ~5×103 cells were seeded. Cells seeded on
coverslips were rinsed in warm PBS three times, permeabilized in
PBS (containing 0.5% Triton X-100) at 4°C for 20 min and fixed in
4% paraformaldehyde at 4°C for 10 min. After 1 h of blocking in 5%
bovine serum album (A8020; Beijing Solarbio Science &
Technology Co., Ltd., Beijing, China) at room temperature, cells
were incubated with specific primary antibodies at room temperature
for 30 min and then at 4°C for overnight. The primary antibodies
were as follows: Rabbit anti-14-3-3ε (1:50) (cat. no. 11648-2-AP;
ProteinTech Group, Inc., Chicago, IL, USA)/mouse
anti-Bcl-2-associated X protein (Bax; 1:100; cat. no. 60267-1-Ig;
ProteinTech Group, Inc.), rabbit anti-14-3-3ε (1:50)/mouse
anti-B-cell lymphoma 2 (Bcl-2; 1:100; cat. no. 60178-1-Ig;
ProteinTech Group, Inc.), rabbit anti-14-3-3ε (1:50)/mouse anti-P53
(1:100; cat. no. 60283-1-Ig; ProteinTech Group, Inc.), muse
anti-14-3-3ε (1:50; cat. no. sc-393177; Santa Cruz Biotechnology,
Inc.)/rabbit anti-c-Fos (1:50; cat. no. 26192-1-AP; ProteinTech
Group, Inc.), mouse anti-14-3-3ε (1:50)/rabbit anti-retinoblastoma
(RB; 1:50; cat. no. 25628-1-AP; ProteinTech Group, Inc.). Following
3 washes (5 min per wash) at room temperature with PBS, cells were
incubated with secondary antibodies mixture containing goat
anti-mouse IgG labeled with fluorescein isothiocyanate (1:200;
FITC; cat. no. P0196; Beyotime Institute of Biotechnology, Haimen,
China) and goat anti-rabbit IgG labeled with cyanine (Cy)3 (1:200;
cat. no. P0183; Beyotime Institute of Biotechnology) at room
temperature for 3 h. Hoechst 33258 (cat. no. C1017; Beyotime
Institute of Biotechnology) was used to stain the nucleus at room
temperature for 5 min. Following 3 washes (5 min/wash) at room
temperature with PBS, the slides were then mounted with antifade
mounting medium (cat. no. P0126; Beyotime Institute of
Biotechnology), sealed at the edges with nail enamel and naturally
dried at room temperature for 1 h. The whole process was performed
in darkness. The prepared samples were preserved in a light-tight
box at −20°C prior to observation. Samples were observed by laser
scanning confocal microscope (LSCM; Olympus FV1000; Tokyo, Japan).
Images were captured under ×400 or ×600 magnification.
Hematoxylin and eosin staining
(HE)
Cells seeded on coverslips were fixed in 95% ethanol
(pre-cooling) at 4°C for 20 min. All steps below were then
performed at room temperature. Following washing with tap water for
40 sec, the coverslips were dipped into hematoxylin solution
containing 0.5% (m/v) hematoxylin, 5% ethanol, 10% (m/v)
KAl(SO4)2, 0.25% (m/v) HgO for 5 min, washed
with tap water for 5 sec, differentiated in 75% ethanol containing
0.5% HCl for 5–10 sec, and then washed with tap water again for 15
min. The coverslips were then stained with eosin Y solution
containing 0.5% (m/v) eosin Y and 20% ethanol for 1 min, followed
by dehydrating with two changes of 95% ethanol and two changes of
100% ethanol for 5 min each. Samples were then cleared with two
changes of xylene for 5 min each, and mounted with neutral balsam.
Once the mounting medium dried, samples were observed under a light
microscope. Images were captured under ×400 magnification.
Statistical analysis
Data reported in the graphs represent the averages
of three independent experiments, and are presented as the mean ±
standard deviation. Data were analyzed by GraphPad Prism 6 software
(GraphPad Software, Inc., La Jolla, CA, USA) using two-way analysis
of variance followed by Tukeys multiple comparison test.
Results
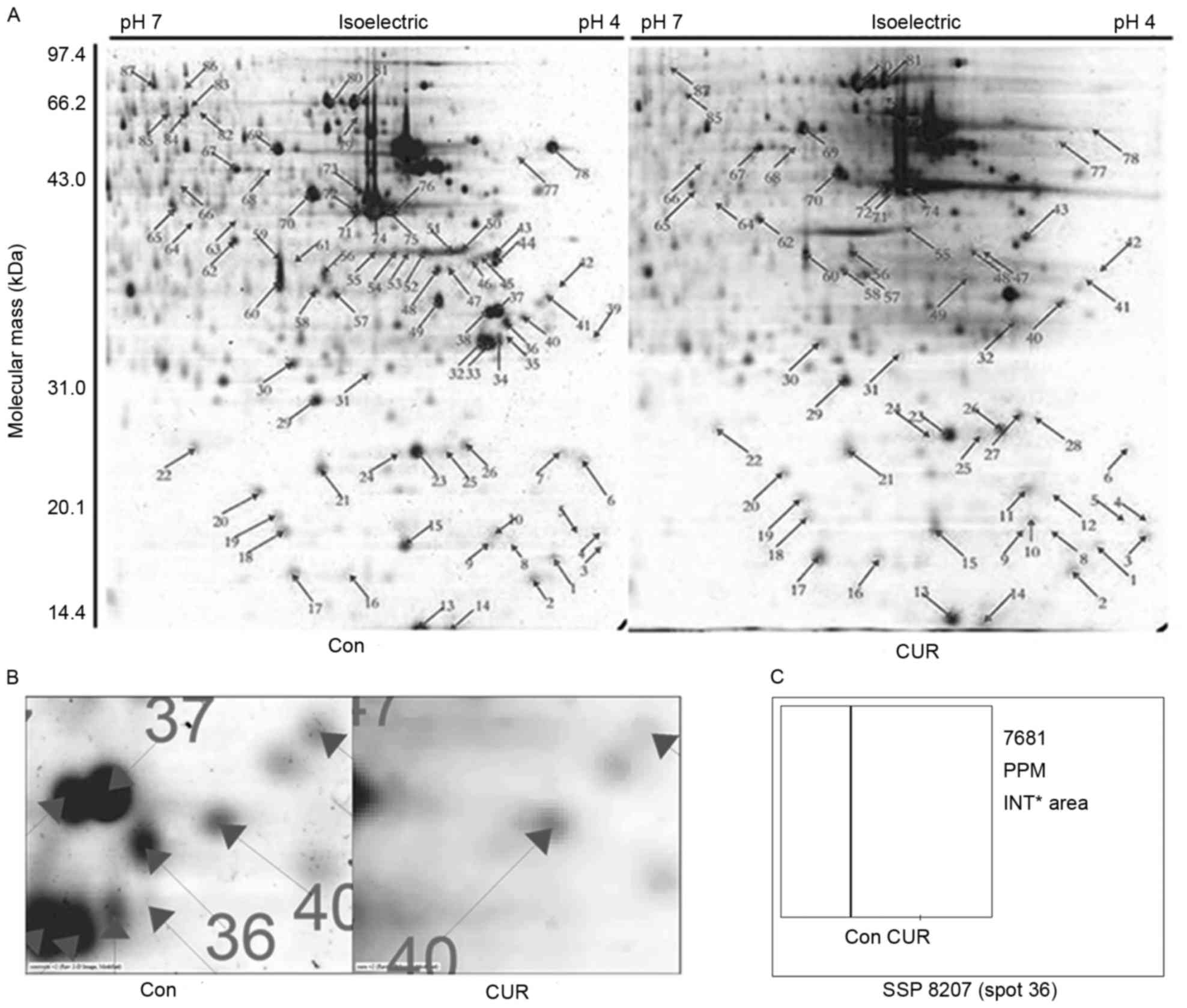
Differentially expressed NMPs were
identified using 2D gel electrophoresis
In order to systematically analyze the alteration in
the protein expression profile in the nuclear matrix during
curcumin-induced apoptosis, selectively extracted NMPs were
separated using 2D gel electrophoresis. The concentrations of
protein spots were analyzed using PDQuest 8.0 software. Protein
spots with a fold change >2 were subjected to MALDI-TOF-MS
analysis (Fig. 1A). The protein spot
36 was identified as protein 14-3-3ε. As presented in Fig. 1B and C, the quantity of spot 36 was
decreased markedly following treatment with curcumin, compared with
the control group. The identification is presented in Fig. 2 and Table
I. Protein score was 258 (a score ≥66 indicates successful
identification).
 | Table I.Identification of 14-3-3ε. |
Table I.
Identification of 14-3-3ε.
| Parameter | Value |
|---|
| Spot no. | 36 |
| Identified
protein | 14-3-3ε |
| Gene symbol | YWHAE |
| NCBI accession
number | NP_006752 |
| Mass, Da | 29,326 |
| pI | 4.63 |
| Coverage, % | 47 |
| Matching
peptides | 16 |
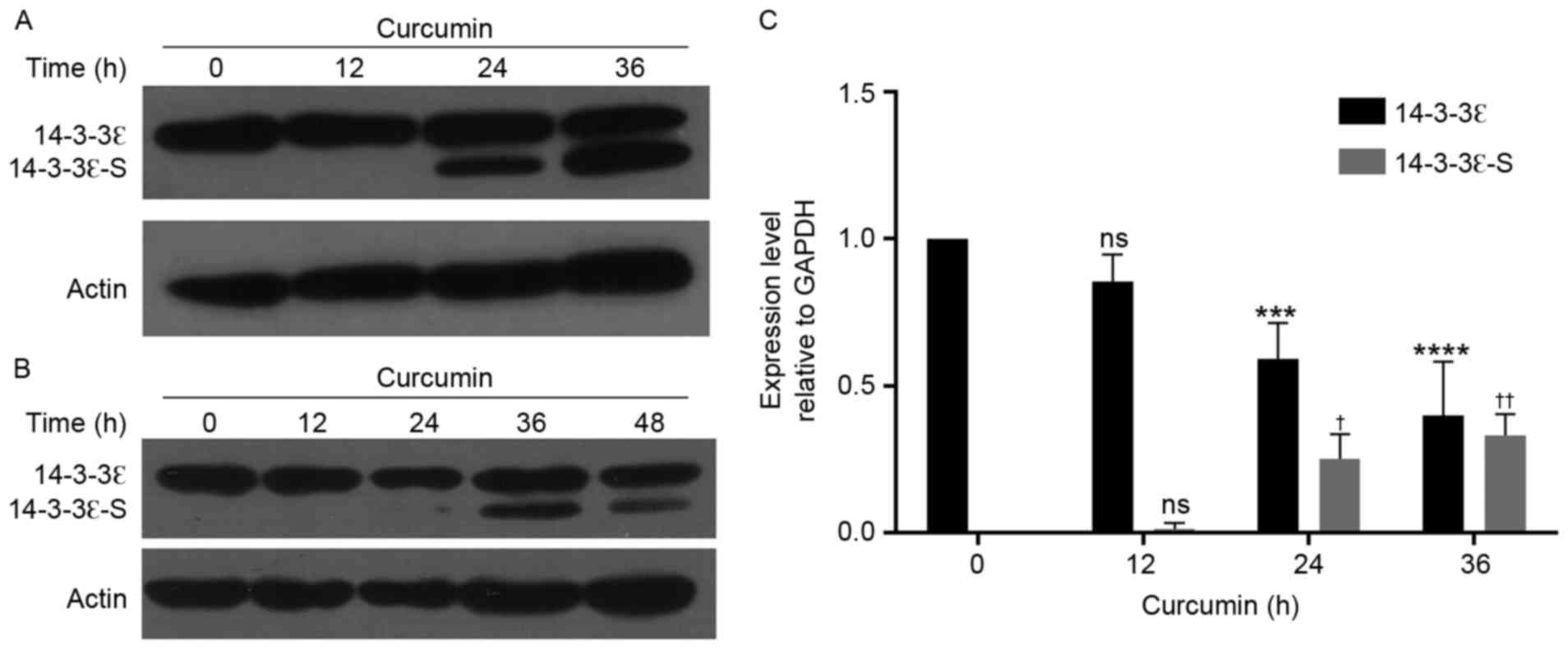
Expression of 14-3-3ε is downregulated
following curcumin treatment
To determine whether curcumin may regulate the
expression of 14-3-3ε, western blot analysis was performed to
detect the expression level of 14-3-3ε. As presented in Fig. 3, following treatment with curcumin for
24 h, the expression level of 14-3-3ε was significantly decreased
in MG-63 cells (P<0.001), as compared with that in control
cells. Notably, a truncated fragment was identified with the
downregulation of 14-3-3ε. To validate whether the detected
fragment was the 14-3-3ε protein, a rabbit antibody (Fig. 3A) and a mouse monoclonal antibody
(Fig. 3B) were used for an immunoblot
assay, and the truncated fragment was detected using the two
antibodies. Thus, it was deduced that this fragment was a specific
short term of 14-3-3ε (14-3-3ε-S).
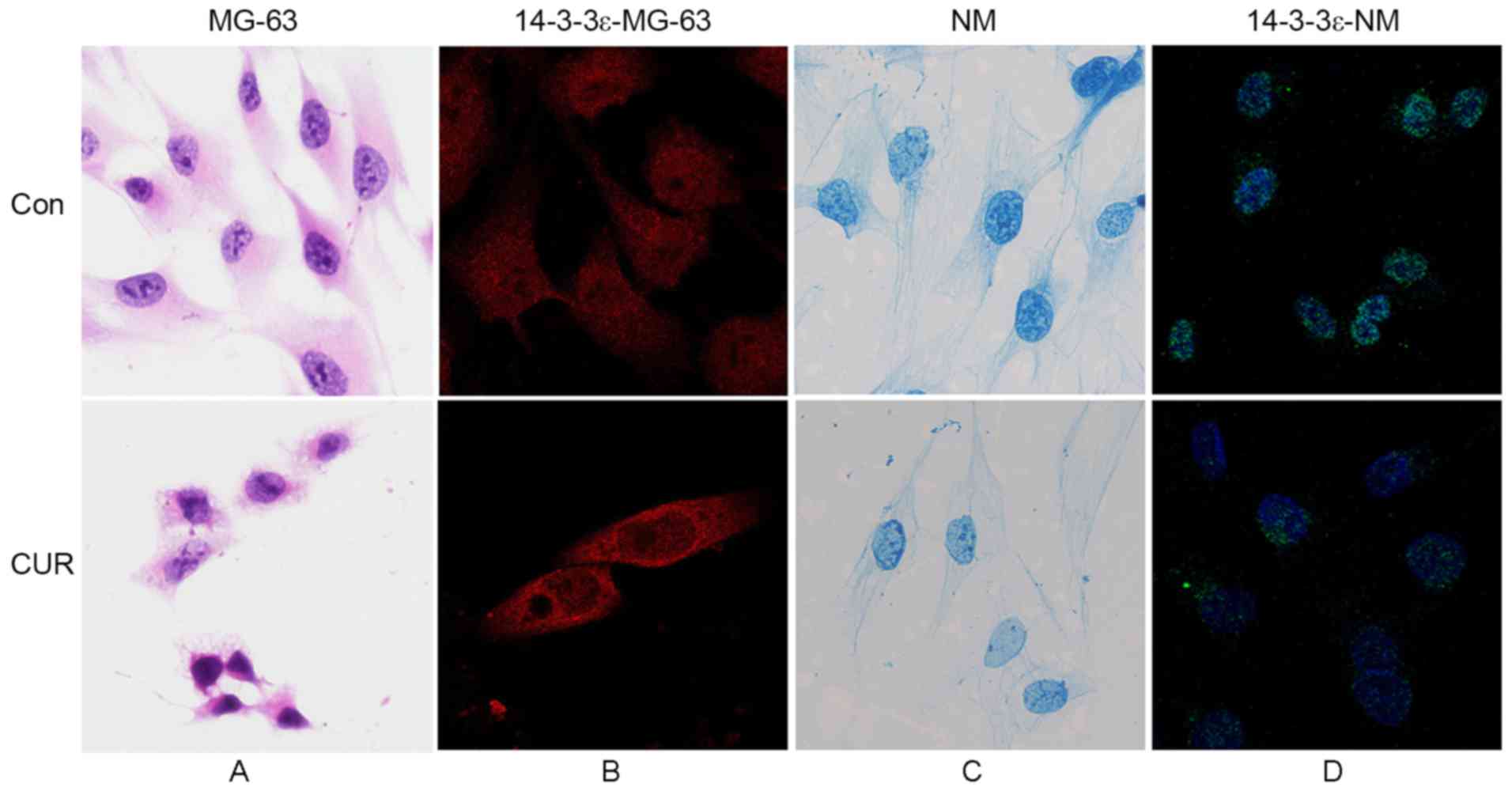
Fluorescence intensity of 14-3-3ε in
the nuclear matrix of apoptotic cells is decreased following
treatment with curcumin
Commonly, the cellular localization of proteins
decides their functions (29–31). Using immunofluorescence labeling
technology enabled the identified of the localization of 14-3-3ε in
MG-63 cells and the nuclear matrix-intermediate filament system,
prior to and following curcumin-induced apoptosis. As presented in
Fig. 4A, MG-63 cells exhibited
irregular morphology, the majority of which were long spindle- or
multiple angle-shaped with sheet or horn-like processes on the cell
surface, and large off-center nuclei. Results from the
immunofluorescence assay (Fig. 4B)
demonstrated that 14-3-3ε was distributed evenly throughout MG-63
cells. Following treatment with curcumin, the cells shrunk and
became round in shape, and the connections between cells and the
processes on the cell surface disappeared. Nucleus staining
revealed karyopyknosis and more intense staining, as compared with
the control cells (Fig. 4A).
Fluorescence microscopy demonstrated that the fluorescence
intensity in the nucleus was weaker (Fig.
4B), compared with that in the cytoplasm, indicating a
decreased level of 14-3-3ε in the nucleus.
As presented Fig. 4C,
the nuclear matrix-intermediate filament (NM-IF) system in MG-63
cells underwent significant alteration during curcumin-induced
apoptosis. Nuclear matrix connected with intermediate filaments
through the nuclear lamina to form an integrated system. In cells
of the control group, intermediate filament was extended between
the edge of the nuclear lamina and the periphery of the cell to
form an intact network structure. In addition, the filaments were
arranged regularly, and gradually became sparse from the
perinuclear region to the edge of the cell. The nuclear lamina was
thick and intensely stained. Fiber hierarchy of nuclear matrix was
compact with deep staining. Following treatment with curcumin, the
nuclear matrix-intermediate filament system tended to be fractured,
disordered and lightly stained. The intermediate fiber was sparse
and discontinuous. The nuclear lamina was thin and the fiber
hierarchy of the nuclear matrix was sparse. The cytoskeleton was no
longer intact. Immunofluorescence staining results (Fig. 4D) revealed that the fluorescence
intensity of 14-3-3ε in the nuclear matrix of apoptotic cells was
decreased, compared with that in normal cells.
Apoptosis-associated proteins are
co-localized with 14-3-3ε in MG-63 cells
The expression level and localization of 14-3-3ε in
MG-63 cells were significantly altered following treatment with
curcumin. To validate that 14-3-3ε was associated with apoptosis, a
confocal laser-scanning double immunofluorescence staining method
was used to observe the localization association between 14-3-3ε
and apoptosis-associated proteins, including Bcl-2, Bax, p53, RB
and c-FOS. In Fig. 5A-C, 14-3-3ε was
labeled with red fluorescent dye Cy3, whereas Bax, Bcl-2 and p53
were labeled with green fluorescent FITC. In Fig. 5D and E, 14-3-3ε was labeled with FITC,
and RB and c-FOS protein were labeled with Cy3. Orange fluorescence
indicated a co-localization association between two proteins. The
results of the present study demonstrated that 14-3-3ε was
primarily distributed in the cytoplasm with a centralized
distribution in the area adjacent to the nucleus. The distribution
of the protein in the nucleus was decreased relative to that in the
cytoplasm. Following treatment with curcumin, 14-3-3ε distribution
remained primarily in the cytoplasm, but in the nucleus the
distribution was weakened.
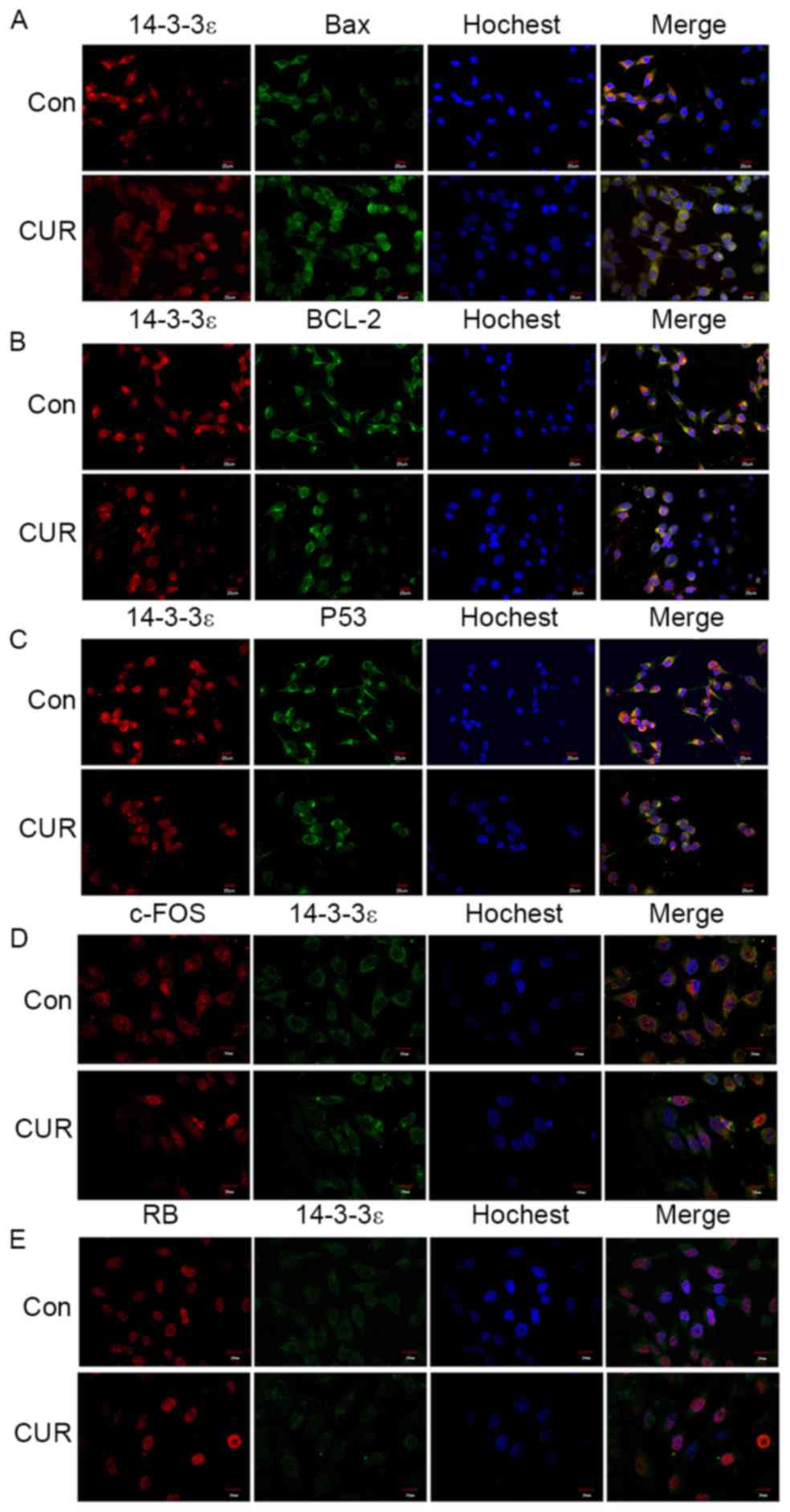 | Figure 5.Laser-scanning confocal microscopy
revealed the co-localization of 14-3-3ε with apoptosis-associated
proteins. (A) Co-localization of 14-3-3ε with Bax. (B)
Co-localization of 14-3-3ε with Bcl-2. (C) Co-localization of
14-3-3ε with P53. Magnification, ×400, with 14-3-3ε labeled with
red Cy3 and Bax, Bcl-2 and p53 labeled with green FITC. (D)
Co-localization of 14-3-3ε with c-FOS. (E) Co-localization of
14-3-3ε with RB. Magnification, ×600, with 14-3-3ε labeled with
green FITC and c-FOS and RB labeled with red Cy3. Bcl-2, B-cell
lymphoma 2; Bax, Bcl-2-associated-X protein; RB, retinoblastoma;
FITC, fluorescein isothiocyanate; Cy, cyanine; Con, control; CUR,
curcumin-treated. |
As presented in Fig.
5A-C, Bax, Bcl-2 and p53 were primarily distributed in the
cytoplasm, and these proteins were at low levels in the nucleus.
Among these proteins, Bcl-2 revealed an intense fluorescent stain
in a concentrated area. The results from the merged images
demonstrated orange fluorescence in the cytoplasm for the three
proteins, indicating that 14-3-3ε was co-localized with Bax, Bcl-2
and p53 in the cytoplasm. Following treatment with curcumin, Bax
and Bcl-2 remained in the cytoplasm, the fluorescence intensity of
Bax was increased compared with that in control cells and the
concentrated distribution of Bcl-2 fluorescence was no longer
visible. The distribution of p53 remained primarily in the
cytoplasm, but there was an increase of fluorescence in the
nucleus, as compared with that in control cells. No orange
fluorescence was observed in the merged images, which indicated
that co-localization of 14-3-3ε with Bax, Bcl-2 and p53 was
decreased or non-existent following curcumin treatment.
The co-localization association between 14-3-3ε and
c-FOS and RB is presented in Fig. 5D and
E. c-FOS was primarily distributed in the cytoplasm, with a
limited amount in the nucleus. RB was restricted to the nucleus,
with the exception of the nucleolus. The superimposition
demonstrated that 14-3-3ε was co-localized with c-FOS in the
cytoplasm. However, for RB, there was no apparent co-localization
association between 14-3-3ε and RB. Compared with the control
group, after treatment with curcumin, the fluorescence intensity of
c-FOS was identified to be significantly decreased in the
cytoplasm, but in the nucleus the fluorescence intensity of c-FOS
was increased. The fluorescence intensity of RB was increased with
no localization alteration. The superimposition demonstrated that
there was no co-localization between 14-3-3ε and RB, and the
co-localization association between 14-3-3ε and c-FOS was
significantly weakened.
Discussion
NMPs are involved with a number of different
activities in the nucleus including DNA replication and synthesis,
RNA transcription and processing, and gene expression regulation;
thus, NMPs serve an important function in cells (22). Identifying the nuclear localization of
14-3-3ε may demonstrate that it exhibits a novel regulatory role in
MG-63 cells. In the present study, 14-3-3ε was identified to be a
nuclear matrix-associated protein using 2D gel electrophoresis. In
addition, western blot analysis validated the existence of 14-3-3ε
in the nuclear matrix, and the expression and localization of
14-3-3ε in the nuclear matrix was observed using immunofluorescent
labeling. Liu et al (7)
identified that 14-3-3ε was primarily located in the nucleus of
SW480 colon cancer cells, with a perinuclear translocation induced
by triptolide. There are a limited number of studies which have
focused on the localization of 14-3-3ε. Our early studies focusing
on apoptosis of MG-63 cells had identified that important
functional proteins including nucleophosmin, prohibitin and
heterogeneous nuclear ribonucleoprotein (hnRNP) A2/B1 were located
in the nuclear matrix (32). By
associating with the nuclear matrix, nucleophosmin, prohibitin and
hnRNP A2/B1 served functions in regulating cell carcinogenesis and
apoptosis (32). Thus, the
localization of 14-3-3ε in the nuclear matrix indicated that it
served an unknown function in the regulation of cell processes,
which required further study.
Expression levels of important functional proteins
are typically altered during cell apoptosis (33–35). The
results of the present study, using 2D analysis, revealed a
significant decrease of 14-3-3ε in the nuclear matrix of apoptotic
MG-63 cells. This alteration indicated that 14-3-3ε, as an
important nuclear matrix-associated protein, was associated with
the regulation of apoptosis in MG-63 cells. In addition, western
blot analysis demonstrated that the expression level of 14-3-3ε was
decreased following curcumin treatment, in a time-dependent manner.
However, the expression level of 14-3-3ε-S demonstrated an opposite
trend, as this increased with treatment duration. Results from
immunofluorescence assays demonstrated a decrease in fluorescence
intensity of 14-3-3ε in the nuclear matrix of apoptotic cells.
Therefore, the results of the present study demonstrated a decrease
in the expression level of 14-3-3ε. Nagappan et al (5) validated that downregulation of 14-3-3ε
induced apoptosis in AGS gastric cancer cells. Consistently, a
study by Wu et al (36)
revealed that rosiglitazone-induced upregulation of 14-3-3ε
decreased the apoptosis of neuronal cells. These studies indicated
that the downregulation of 14-3-3ε was an upstream event of
apoptosis, rather than a downstream one. Notably, the results of
the present study revealed that the expression level of 14-3-3ε-S
was increased along with apoptosis, which should be additionally
investigated.
The regulation of cell apoptosis is a complex
process (37). Thus, we hypothesized
that the interactions between 14-3-3ε and a number of additional
apoptosis-associated proteins may be involved in cell apoptosis.
Using LSCM, it was observed in the present study that 14-3-3ε was
co-localized with Bax, Bcl-2, p53 and c-FOS in the cytoplasm of
MG-63 cells. Following treatment with curcumin, their
co-localization was decreased. To the best of our knowledge, only
Bcl-2-associated death promoter has been identified to bind to
14-3-3ε, resulting in its retention in the cytoplasm and
suppression of apoptosis (38). Our
group hypothesized that Bax may bind to 14-3-3ε, thus impairing its
pro-apoptotic ability. Previous studies have demonstrated that
14-3-3σ, τ, ζ and ε may bind to Bax (18,39), but
the interaction between 14-3-3ε and Bax requires validation. The
ratio between Bax and Bcl-2 is a key factor to determine whether
apoptosis occurs or not (40). The
results of the present study demonstrated that 14-3-3ε was
co-localized with Bax and Bcl-2, which indicated that 14-3-3ε may
regulate the progress of apoptosis by influencing the ratio between
Bax and Bcl-2. As an important transcription factor, p53 may
regulate the expression of multiple pro-apoptotic genes including
Bax and Fas, thereby serving a pro-apoptotic function (41,42).
Gbormittah et al (43)
demonstrated a co-expression and co-localization association
between 14-3-3ε and p53, using chromosome gene mapping. To the best
of our knowledge, there are no other studies which demonstrated an
association between 14-3-3ε and p53. In the present study, 14-3-3ε
was identified to be co-localized with p53 in the cytoplasm in
MG-63 cells, which indicated that the interaction between 14-3-3ε
and p53 may serve a function in the apoptosis of MG-63 cells. c-FOS
is a proto-oncogene which has been identified to be associated with
the occurrence, invasion and metastasis of multiple types of tumor
(44). In some studies, c-FOS was
identified to be involved with the death of nerve cells (45–47). In
addition, 14-3-3ε has been demonstrated to serve a function in the
death of nerve cells (48–50). Although, to the best of our knowledge,
no studies have identified an association between 14-3-3ε and
c-FOS, their co-localization suggests that these two may interact.
Co-localization of 14-3-3ε with the aforementioned
apoptosis-associated proteins indicated that 14-3-3ε may
participate in the regulation of apoptosis of MG-63 cells.
The present study demonstrated that 14-3-3ε exists
in the nuclear matrix of MG-63 cells and is downregulated during
curcumin-induced apoptosis. Furthermore, 14-3-3ε is co-localized
with Bax, Bcl-2, p53 and c-FOS in the cytoplasm of MG-63 cells, and
these co-localizations are decreased following curcumin treatment.
These results indicated that 14-3-3ε is a nuclear matrix-associated
protein and it may serve a role in the regulatory events of
apoptosis.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81272921, 81272245,
81272445, 81471970 and 81670542), Joint Programme by Healthy Care
System and Educational Department in Fujian Province (grant no.
WKJ-FJ-16), The Natural Science Foundation of Fujian Province of
China (grant no. 2013D004) and the Fundamental Research Funds for
the Central Universities (grant no. 20720140545).
References
|
1
|
Morrison DK: The 14-3-3 proteins:
Integrators of diverse signaling cues that impact cell fate and
cancer development. Trends Cell Biol. 19:16–23. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
van Hemert MJ, Steensma HY and van Heusden
GP: 14-3-3 proteins: Key regulators of cell division, signalling
and apoptosis. Bioessays. 23:936–946. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kosaka Y, Cieslik KA, Li L, Lezin G,
Maguire CT, Saijoh Y, Toyo-oka K, Gambello MJ, Vatta M,
Wynshaw-Boris A, et al: 14-3-3ε plays a role in cardiac ventricular
compaction by regulating the cardiomyocyte cell cycle. Mol Cell
Biol. 32:5089–5102. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cui C, Ren X, Liu D, Deng X, Qin X, Zhao
X, Wang E and Yu B: 14-3-3 epsilon prevents G2/M transition of
fertilized mouse eggs by binding with CDC25B. BMC Dev Biol.
14:332014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nagappan A, Park HS, Park KI, Hong GE,
Yumnam S, Lee HJ, Kim MK, Kim EH, Lee WS, Lee WJ, et al:
Helicobacter pylori infection combined with DENA revealed altered
expression of p53 and 14-3-3 isoforms in Gulo-/- mice. Chem Biol
Interact. 206:143–152. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhong J, Kong X, Zhang H, Yu C, Xu Y, Kang
J, Yu H, Yi H, Yang X and Sun L: Inhibition of CLIC4 enhances
autophagy and triggers mitochondrial and ER stress-induced
apoptosis in human glioma U251 cells under starvation. PLoS One.
7:e393782012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu Y, Song F, Wu WK, He M, Zhao L, Sun X,
Li H, Jiang Y, Yang Y and Peng K: Triptolide inhibits colon cancer
cell proliferation and induces cleavage and translocation of 14-3-3
epsilon. Cell Biochem Funct. 30:271–278. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fong WH, Tsai HD, Chen YC, Wu JS and Lin
TN: Anti-apoptotic actions of PPAR-gamma against ischemic stroke.
Mol Neurobiol. 41:180–186. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang X, Grammatikakis N, Siganou A and
Calderwood SK: Regulation of molecular chaperone gene transcription
involves the serine phosphorylation, 14-3-3 epsilon binding, and
cytoplasmic sequestration of heat shock factor 1. Mol Cell Biol.
23:6013–6026. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Leal MF, Calcagno DQ, Demachki S,
Assumpcão PP, Chammas R, Burbano RR and Mde A Smith: Clinical
implication of 14-3-3 epsilon expression in gastric cancer. World J
Gastroenterol. 18:1531–1537. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ruiz E, sparza-Garrido R, Velázquez-Flores
MÁ, Diegopérez-Ramirez J, López-Aguilar E, Siordia-Reyes G,
Hernández-Ortiz M, Martinez-Batallar AG, Encarnación-Guevara S,
Salamanca-Gómez F and Arenas-Aranda DJ: A proteomic approach of
pediatric astrocytomas: MiRNAs and network insight. J Proteomics.
94:162–175. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ko BS, Jan YJ, Chang TC, Liang SM, Chen
SC, Liu TA, Wu YM, Wang J and Liou JY: Upregulation of focal
adhesion kinase by 14-3-3ε via NFκB activation in hepatocellular
carcinoma. Anticancer Agents Med Chem. 13:555–562. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ko BS, Chang TC, Hsu C, Chen YC, Shen TL,
Chen SC, Wang J, Wu KK, Jan YJ and Liou JY: Overexpression of
14-3-3ε predicts tumour metastasis and poor survival in
hepatocellular carcinoma. Histopathology. 58:705–711. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liang S, Xu Y, Shen G, Liu Q, Zhao X, Xu
Z, Xie X, Gong F, Li R and Wei Y: Quantitative protein expression
profiling of 14-3-3 isoforms in human renal carcinoma shows 14-3-3
epsilon is involved in limitedly increasing renal cell
proliferation. Electrophoresis. 30:4152–4162. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cimino D, Fuso L, Sfiligoi C, Biglia N,
Ponzone R, Maggiorotto F, Russo G, Cicatiello L, Weisz A, Taverna
D, et al: Identification of new genes associated with breast cancer
progression by gene expression analysis of predefined sets of
neoplastic tissues. Int J Cancer. 123:1327–1338. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Porter GW, Khuri FR and Fu H: Dynamic
14-3-3/client protein interactions integrate survival and apoptotic
pathways. Semin Cancer Biol. 16:193–202. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zha J, Harada H, Yang E, Jockel J and
Korsmeyer SJ: Serine phosphorylation of death agonist BAD in
response to survival factor results in binding to 14-3-3 not
BCL-X(L). Cell. 87:619–628. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nomura M, Shimizu S, Sugiyama T, Narita M,
Ito T, Matsuda H and Tsujimoto Y: 14-3-3 Interacts directly with
and negatively regulates pro-apoptotic Bax. J Biol Chem.
278:2058–2065. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang L, Chen J and Fu H: Suppression of
apoptosis signal-regulating kinase 1-induced cell death by 14-3-3
proteins. Proc Natl Acad Sci USA. 96:pp. 8511–8515. 1999;
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Brunet A, Kanai F, Stehn J, Xu J,
Sarbassova D, Frangioni JV, Dalal SN, DeCaprio JA, Greenberg ME and
Yaffe MB: 14-3-3 transits to the nucleus and participates in
dynamic nucleocytoplasmic transport. J Cell Biol. 156:817–828.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fey EG, Wan KM and Penman S: Epithelial
cytoskeletal framework and nuclear matrix-intermediate filament
scaffold: Three-dimensional organization and protein composition. J
Cell Biol. 98:1973–1984. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gerner C, Seelos C and Sauermann G:
Alteration of nuclear matrix protein composition during apoptosis
in rat embryo cells. Exp Cell Res. 238:472–480. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Michishita E, Kurahashi T, Suzuki T,
Fukuda M, Fujii M, Hirano H and Ayusawa D: Changes in nuclear
matrix proteins during the senescence-like phenomenon induced by
5-chlorodeoxyuridine in HeLa cells. Exp Gerontol. 37:885–890. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jing GJ, Xu DH, Shi SL, Li QF, Wang SY, Wu
FY and Kong HY: Aberrant expression of nuclear matrix proteins
during HMBA-induced differentiation of gastric cancer cells. World
J Gastroenterol. 16:2176–2182. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shevchenko A, Wilm M, Vorm O and Mann M:
Mass spectrometric sequencing of proteins silver-stained
polyacrylamide gels. Anal Chem. 68:850–858. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Boutet E, Lieberherr D, Tognolli M,
Schneider M, Bansal P, Bridge AJ, Poux S, Bougueleret L and
Xenarios I: UniProtKB/Swiss-Prot, the manually annotated section of
the UniProt KnowledgeBase: How to use the entry view. Methods Mol
Biol. 1374:23–54. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
NCBI Resource Coordinators, . Database
resources of the national center for biotechnology information.
Nucleic Acids Res. 45:D12–D17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li QF: Effect of retinoic acid on the
changes of nuclear matrix in termediate filament system in gastric
carcinoma cells. World J Gastroenterol. 5:417–420. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Poüs C, Klipfel L and Baillet A:
Cancer-related functions and subcellular localizations of septins.
Front Cell Dev Biol. 4:1262016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hsu YT, Wolter KG and Youle RJ:
Cytosol-to-membrane redistribution of Bax and Bcl-X(L) during
apoptosis. Proc Natl Acad Sci USA. 94:pp. 3668–3672. 1997;
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Boonstra J and Verkleij AJ: Regulation of
enzyme activity in vivo is determined by its cellular localization.
Adv Enzyme Regul. 44:61–73. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao ZL, Li QF, Zheng YB, Chen LY, Shi SL
and Jing GJ: The aberrant expressions of nuclear matrix proteins
during the apoptosis of human osteosarcoma cells. Anat Rec
(Hoboken). 293:813–820. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bai Z, Ye Y, Liang B, Xu F, Zhang H, Zhang
Y, Peng J, Shen D, Cui Z, Zhang Z and Wang S: Proteomics-based
identification of a group of apoptosis-related proteins and
biomarkers in gastric cancer. Int J Oncol. 38:375–383.
2011.PubMed/NCBI
|
|
34
|
Wyllie AH: Apoptosis (the 1992 Frank Rose
Memorial Lecture). Br J Cancer. 67:205–208. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bazhanova ED, Sukhanov DS and Teplyi DL:
Pathways of apoptosis regulation in hepatocytes induced by
first-line antitubercular drugs. Bull Exp Biol Med. 158:650–653.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu JS, Cheung WM, Tsai YS, Chen YT, Fong
WH, Tsai HD, Chen YC, Liou JY, Shyue SK, Chen JJ, et al:
Ligand-activated peroxisome proliferator-activated receptor-gamma
protects against ischemic cerebral infarction and neuronal
apoptosis by 14-3-3 epsilon upregulation. Circulation.
119:1124–1134. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wong RS: Apoptosis in cancer: From
pathogenesis to treatment. J Exp Clin Cancer Res. 30:872011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Won J, Kim DY, La M, Kim D, Meadows GG and
Joe CO: Cleavage of 14-3-3 protein by caspase-3 facilitates bad
interaction with Bcl-x(L) during apoptosis. J Biol Chem.
278:19347–19351. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Samuel T, Weber HO, Rauch P, Verdoodt B,
Eppel JT, McShea A, Hermeking H and Funk JO: The G2/M regulator
14-3-3sigma prevents apoptosis through sequestration of Bax. J Biol
Chem. 276:45201–45206. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Selvakumaran M, Lin HK, Miyashita T, Wang
HG, Krajewski S, Reed JC, Hoffman B and Liebermann D: Immediate
early up-regulation of bax expression by p53 but not TGF beta 1: A
paradigm for distinct apoptotic pathways. Oncogene. 9:1791–1798.
1994.PubMed/NCBI
|
|
41
|
Meek DW: The p53 response to DNA damage.
DNA Repair (Amst). 3:1049–1056. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fridman JS and Lowe SW: Control of
apoptosis by p53. Oncogene. 22:9030–9040. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gbormittah FO, Haab BB, Partyka K,
Garcia-Ott C, Hancapie M and Hancock WS: Characterization of
glycoproteins in pancreatic cyst fluid using a high-performance
multiple lectin affinity chromatography platform. J Proteome Res.
13:289–299. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Milde-Langosch K: The Fos family of
transcription factors and their role in tumourigenesis. Eur J
Cancer. 41:2449–2461. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Smeyne RJ, Vendrell M, Hayward M, Baker
SJ, Miao GG, Schilling K, Robertson LM, Curran T and Morgan JI:
Continuous c-fos expression precedes programmed cell death in vivo.
Nature. 363:166–169. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen X, Shen J, Wang Y, Chen X, Yu S, Shi
H and Huo K: Up-regulation of c-Fos associated with neuronal
apoptosis following intracerebral hemorrhage. Cell Mol Neurobiol.
35:363–376. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yu AY, Su QR, Wang L, Zhou J and Liu XH:
Effects of citalopram on the expression of PCNA and C-fos and cell
apoptosis in rat frontal cortical neurons after stress. Zhongguo
Ying Yong Sheng Li Xue Za Zhi. 30:439–442. 2014.(In Chinese).
PubMed/NCBI
|
|
48
|
Morales D, Skoulakis EC and Acevedo SF:
14-3-3s are potential biomarkers for HIV-related neurodegeneration.
J Neurovirol. 18:341–353. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gelman BB and Nguyen TP: Synaptic proteins
linked to HIV-1 infection and immunoproteasome induction: Proteomic
analysis of human synaptosomes. J Neuroimmune Pharmacol. 5:92–102.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Toyo-oka K, Shionoya A, Gambello MJ,
Cardoso C, Leventer R, Ward HL, Ayala R, Tsai LH, Dobyns W,
Ledbetter D, et al: 14-3-3epsilon is important for neuronal
migration by binding to NUDEL: A molecular explanation for
Miller-Dieker syndrome. Nat Genet. 34:274–285. 2003. View Article : Google Scholar : PubMed/NCBI
|