Introduction
Lung cancer is one of the most common types of
cancer and remains the leading cause of cancer-associated mortality
in the world (1). Non-small cell lung
carcinoma (NSCLC) accounts for ~85% of all lung cancer cases
(2) and the majority of patients with
NSCLC are diagnosed at an advanced stage (3). Despite improvements in the diagnosis and
treatment of NSCLC, the 5-year survival rate for NSCLC remains low,
at between 10 and 20% (4,5). Therefore, it is important to identify
key risk factors and to design novel therapeutic strategies to
prevent or treat NSCLC.
Previous studies have suggested that ~20% of all
types of cancer are due to obesity (6,7). Obesity
is strongly associated with the development of type II diabetes
mellitus, which is accompanied by elevated insulin levels (8). Epidemiological studies have demonstrated
that increased insulin levels are associated with an increased risk
of developing cancer, including breast, pancreatic, colon and
bladder cancer (9–12). Insulin acts as a powerful mitogen and
has been implicated in the onset and progression of tumors
(13–15). High levels of insulin accelerated the
proliferation of pancreatic ductal cells and increased migration in
breast cancer and colon cancer cells (13–15).
However, the effect of insulin on NSCLC has not yet been
evaluated.
Activation of the phosphoinositide 3-kinase/protein
kinase B (PI3K/Akt) signaling pathway indicates poor patient
prognosis and is associated with different types of cancer,
including NSCLC, prostate and breast cancer (16–18).
Furthermore, activation of the PI3K/Akt signaling pathway may
promote tumor cell proliferation, migration and drug resistance
(19). It has been demonstrated that
insulin stimulates the PI3K/Akt signaling pathway to increase
carcinogenesis in breast cancer and colon cancer cells (14). However, it remains unknown whether
insulin is able to regulate the development of NSCLC by activating
the PI3K/Akt signaling pathway.
The results of the present study indicated that
insulin enhanced the proliferation, migration and drug resistance
of NSCLC cells. In addition, LY294002, a specific inhibitor of the
PI3K/Akt signaling pathway, reversed the oncogenic effects of
insulin on protein expression. The results of the present study may
therefore improve understanding of the effect of insulin on
NSCLC.
Materials and methods
Reagents
Insulin, LY294002, diamminedichloroplatinum (DDP),
RNase A, propidium iodide (PI) and RIPA buffer were purchased from
Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). BCA Protein Assay
kit and enhanced chemiluminescence (ECL) reagent were purchased
from Thermo Fisher Scientific, Inc., Waltham, MA, USA. RPMI-1640,
high-glucose Dulbecco's modified Eagle's medium (DMEM) and fetal
bovine serum (FBS) were purchased from Gibco; Thermo Fisher
Scientific, Inc. Transwell chambers and Matrigel Invasion Chambers
were purchased from BD Biosciences, Inc., Rockville, MD, USA.
Antibodies against β-actin (cat. no. 3700), phospho-Akt (p-Akt,
cat. no. D25E6) and Akt (cat. no. 11E7) were purchased from Cell
Signaling Technologies, Inc., Danvers, MA, USA. Antibodies against
insulin receptor substrate 1 (IRS1, cat. no. ab52167) and
phospho-IRS1 (p-IRS1, cat. no. ab1194) were purchased from Abcam,
Cambridge, UK). Antibodies against proliferating cell nuclear
antigen (PCNA, cat. no. sc-25280), cyclin A (cat. no. sc-751), p27
(cat. no. sc-528), P-glycoprotein (P-gp, cat. no. sc-55510) and
matrix metalloproteinase 3 (MMP3, cat. no. sc-21732) were all
purchased from Santa Cruz Biotechnology, Dallas, TX, USA. Goat
anti-mouse IgG (cat. no. 31430) and goat anti-rabbit IgG (cat. no.
31460) secondary antibodies conjugated to horseradish peroxidase
(HRP) were purchased from Invitrogen; Thermo Fisher Scientific,
Inc.
Cell culture
The human lung cancer cell lines A549, PC-9 and
NCI-H1975 were provided by Dr Yan-Jun Mi (The First Affiliated
Hospital of Xiamen University, Xiamen, China). Cells were
maintained in RPMI-1640 medium (NCI-H1975 cells) or DMEM (A549 and
PC-9 cells) supplemented with 10% FBS and 100 U/ml penicillin at
37°C in a humidified atmosphere of 5% CO2.
Cell proliferation assay
Cell proliferation was determined using MTT, which
was dissolved in dimethyl sulfoxide. Cells were suspended at a
concentration of 5×103/well, seeded into a 96-well plate
and incubated overnight at 37°C. Following treatment with different
concentration of insulin (0, 400, 800, 1600 nM), 20 µl MTT (5
mg/ml) was added to each well and the plate was incubated under the
same conditions for 4 h. Absorbance was measured at 490 nm using an
auto microplate reader. All experiments were performed in
triplicate.
Drug resistance assay
Cells were seeded into a 96-well plate at a
concentration of 5×103/well. Following treatment with
1,600 nM insulin in the presence of various concentrations of DDP
(0, 5, 10, 20 and 40 µmol) at 37°C for 48 h, 20 µl MTT (5 mg/ml)
was added to each well and the subsequent steps were performed
according to the aforementioned cell proliferation assay protocol.
The group treated with PBS acted as a control.
Flow cytometry
A549 cells were synchronized through treatment with
serum-free high-glucose DMEM for 24 h. Following the addition of
serum-free high-glucose DMEM with or without insulin (1,600 nM) for
3 days, A549 cells were harvested, washed twice with ice cold PBS
and fixed with ice cold 70% ethanol at 4°C overnight. Cells were
washed with cold PBS and incubated with RNase A (100 µg/ml) at 37°C
for 30 min. Cells were subsequently stained with PI (50 µg/ml) at
4°C for 30 min and analyzed using a BD FACSCalibur Flow Cytometer
(BD Biosciences, San Jose, CA, USA). The data were analyzed with
the ModFit 3.3 (Verity Software House, Topsham, ME, USA) software.
All experiments were performed in triplicate.
Cell migration and invasion assay
Cell migration was detected by wound healing and
transwell assays, which were performed following a previously
described protocol (20). A549 cells
were used to investigate cell migration and invasion assay due to
the characteristic of easy culture ability. For the wound healing
assay, scratches were made with 10-µl micropipette tip. Then,
serum-free high-glucose DMEM was added with or without insulin
(1,600 nM). For the transwell assay, A549 cells were suspended at a
concentration of 1×105/well, seeded into the upper
chamber of a 24-well plate in serum-free high-glucose DMEM with or
without insulin (1,600 nM) for 24 h. A total of 500 µl DMEM was
added to the lower chamber. The cells that migrated to the lower
chamber were stained with 0.05% crystal violet at room temperature
for 10 min and counted using an inverted microscope. Cell invasion
assays were performed using transwell invasion chambers with
Matrigel. A549 cells were seeded at a density of
1×105/well into the upper chamber of a 24-well plate in
serum-free high-glucose DMEM with or without insulin (1,600 nM),
and incubated for 24 h. The subsequent steps were performed
according to the aforementioned transwell assay protocol. All
experiments were performed in triplicate.
Western blotting
Western blotting was performed as previously
described (20). Total protein was
extracted using RIPA buffer and its concentration was evaluated
using the BCA Protein Assay kit. Proteins were separated using 10%
SDS-PAGE and transferred onto a polyvinylidene difluoride membrane.
Following blocking with 5% milk in PBST at room temperature for 1
h, the membrane was incubated with primary antibodies (β-actin,
1:1,000; p-Akt, 1:1,000; Akt, 1:1,000; IRS1, 1:1,000; p-IRS1,
1:1,000; PCNA, 1:500; cyclin A, 1:500; p27, 1:500; P-gp, 1:500;
MMP3, 1:500) at 4°C overnight followed by incubation with the
appropriate HRP-conjugated secondary antibody (1:10,000) at room
temperature for 1 h. The signal was subsequently visualized using
an ECL reporter system (Thermo Fisher Scientific, Inc.) followed by
exposure to X-ray film. All experiments were performed in
triplicate.
Statistical analysis
Data are presented as the mean ± standard error of
the mean. Unpaired Student's t test or one-way analysis of variance
were used for comparison of two or more datasets, respectively.
Multiple comparison between the groups was performed using
Student-Newman-Keuls method. P<0.05 was considered to indicate a
statistically significant difference. The data were analyzed with
SPSS version 16.0 statistical software package (SPSS Inc., Chicago,
IL, USA). All groups contained three replicates and all experiments
were repeated three times.
Results
Insulin promotes the proliferation of
NSCLC cells
An MTT assay was performed to determine the role of
insulin in the proliferation of NSCLC cells. The proliferation of
A549, PC-9 and NCI-H1975 cells were enhanced by insulin in a dose-
and time-dependent manner (Fig. 1A-C;
0 nM insulin acted as a control; *P<0.05 vs. 0 nM insulin at 2
or 3 d in A549, PC-9 or NCI-H1975 cells; **P<0.01 vs. 0 nM
insulin at 3 d in PC-9 cells). The enhancing effect of insulin on
the proliferation of the aforementioned cells, were more obvious by
day 3. In addition, the results of flow cytometry demonstrated that
insulin significantly decreased the proportion of cells in the G1
phase (P<0.05) and significantly increased the proportion of
cells in the S-phase compared with control (P<0.05; Fig. 1D and E). Western blotting was
performed to measure the expression of cell cycle and
growth-related proteins in A549 cells. The expression of cyclin A
and PCNA were increased and of p27 protein expression was decreased
in a dose-dependent manner following treatment with insulin
(Fig. 1F). These results indicate
that insulin promotes the proliferation of the NSCLC cells.
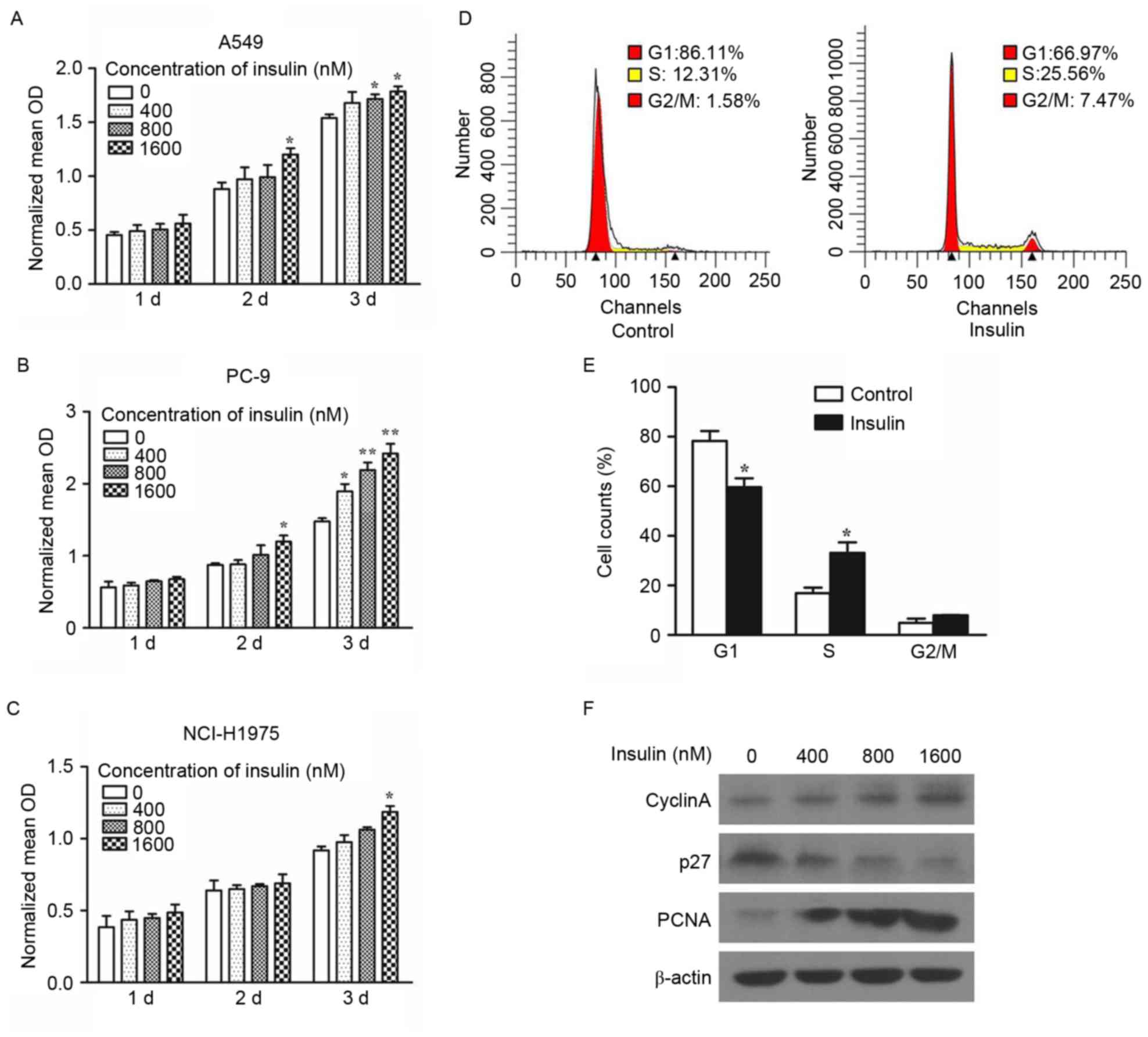 | Figure 1.The proliferation of NSCLC cells
following treatment with insulin. The proliferation of (A) A549,
(B) PC-9 and (C) NCI-H1975 cells was measured using an MTT assay.
*P<0.05 and **P<0.01 vs. 0 nM insulin. (D and E) A549 cells
were treated with 1,600 nM insulin for 3 days and subsequently
analyzed by flow cytometry. *P<0.05 vs. the control. (F) Cyclin
A, p27 and PCNA expression were assessed in A549 cells by western
blotting following treatment with 400, 800 and 1,600 nM insulin for
3 days. Data are presented as the mean ± standard error of the
mean. *P<0.05, **P<0.01. PCNA, proliferating cell nuclear
antigen; NSCLC, non-small cell lung cancer; d, days; OD, optical
density. |
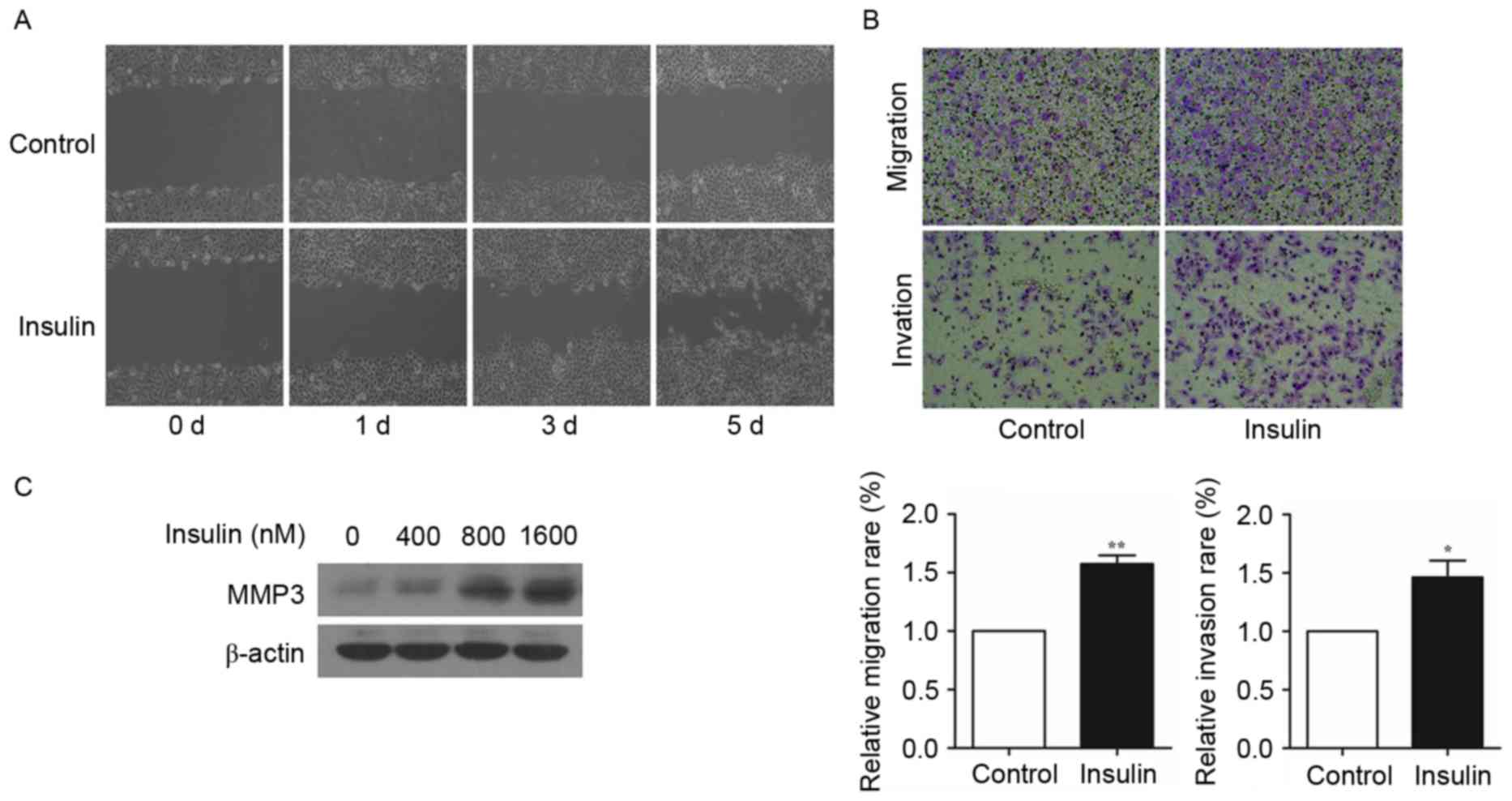
Insulin promotes the migration and
invasion of NSCLC cells
Subsequently, the effect of insulin on NSCLC
migration and invasion was detected by wound healing and Transwell
assays. The ability of migration and invasion were markedly
elevated by insulin in A549 cells (Fig.
2A and B). Furthermore, the number of migrating and invading
A549 cells were significantly increased following treatment with
insulin compared with untreated control cells (Fig. 2B; P<0.05). In addition, insulin
enhanced the expression of MMP3 in a dose-dependent manner in A549
cells (Fig. 2C). Taken together,
these results suggest that insulin may promote the migration and
invasion of NSCLC cells by regulating MMP3 expression.
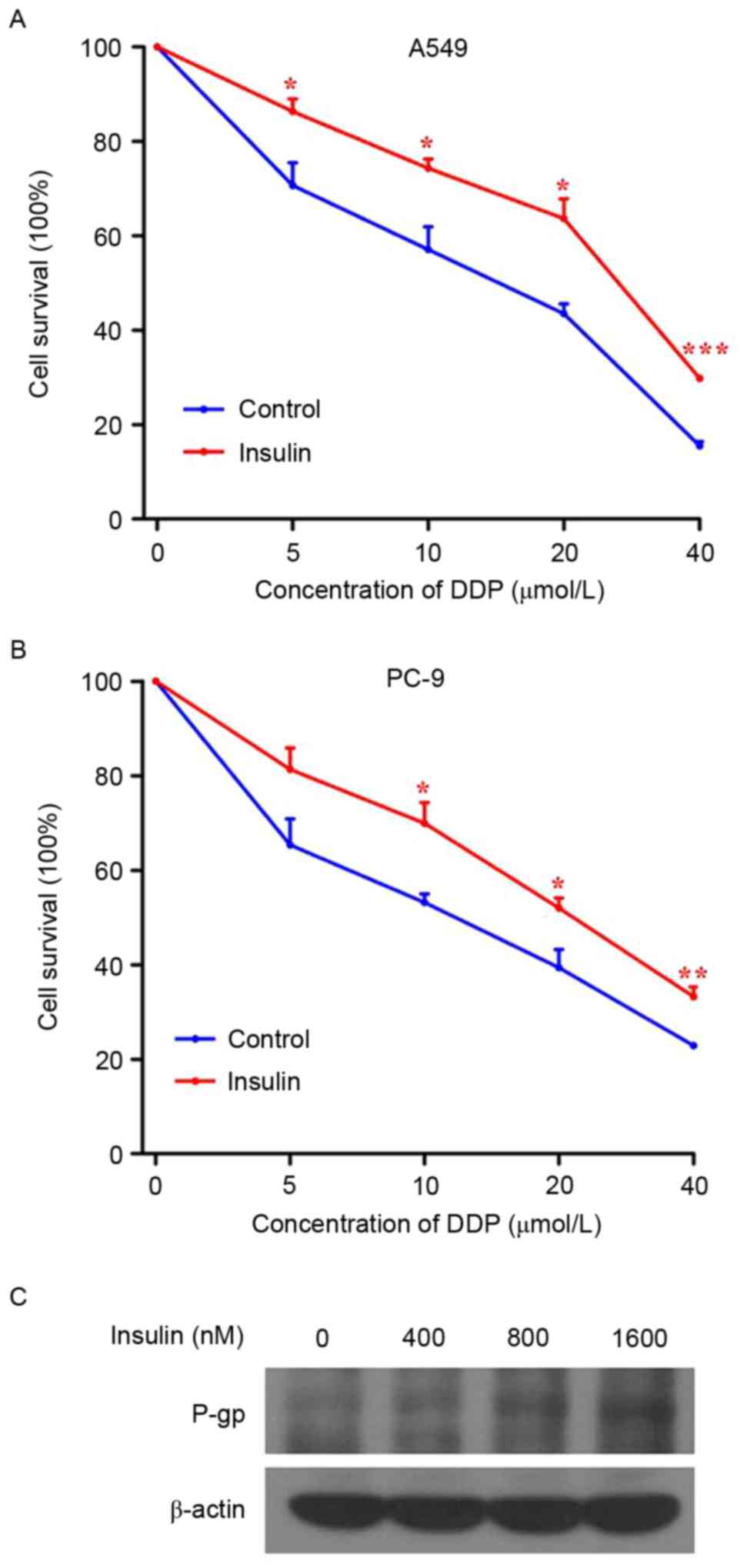
Insulin increases the resistance of
NSCLC cells to DDP
To investigate the effect of insulin on drug
resistance, A549 and PC-9 cells were treated with different
concentrations of DDP (0, 5, 10, 20 and 40 µmol) in the absence or
presence of insulin. The sensitivity of A549 and PC-9 cells to DDP
was significantly reduced by insulin (Fig. 3A and B; P<0.05). Furthermore, the
expression of P-gp in A549 cells was increased by insulin in a
dose-dependent manner (Fig. 3C),
indicating that insulin may reduce the sensitivity of NSCLC cells
to DDP by upregulating P-gp expression.
Insulin exhibits oncogenic activity
via the PI3K/Akt signaling pathway
To further explore the mechanism of insulin on cell
proliferation, migration and drug sensitivity, the effect of
insulin on the IRS and PI3K/Akt signaling pathway in NSCLC, which
has been investigated in other types of cancer (14,21), was
determined by western blot analysis. The results demonstrated that
insulin increased p-Akt and p-IRS1 expression in a dose-dependent
manner (Fig. 4A). Furthermore,
following the addition of LY294002, a PI3K/Akt inhibitor, the
expression of p-Akt and p-IRS1 in A549 cells was reduced compared
with cells treated with insulin alone and was similar to that of
untreated control cells (Fig. 4B).
Correspondingly, LY294002 also reversed the effects of insulin on
the expression of cyclin A, p27, PCNA, MMP3 and P-gp in A549 cells
(Fig. 4C). These results indicate
that the PI3K/Akt signaling pathway may be involved in the
oncogenic activity of insulin in NSCLC.
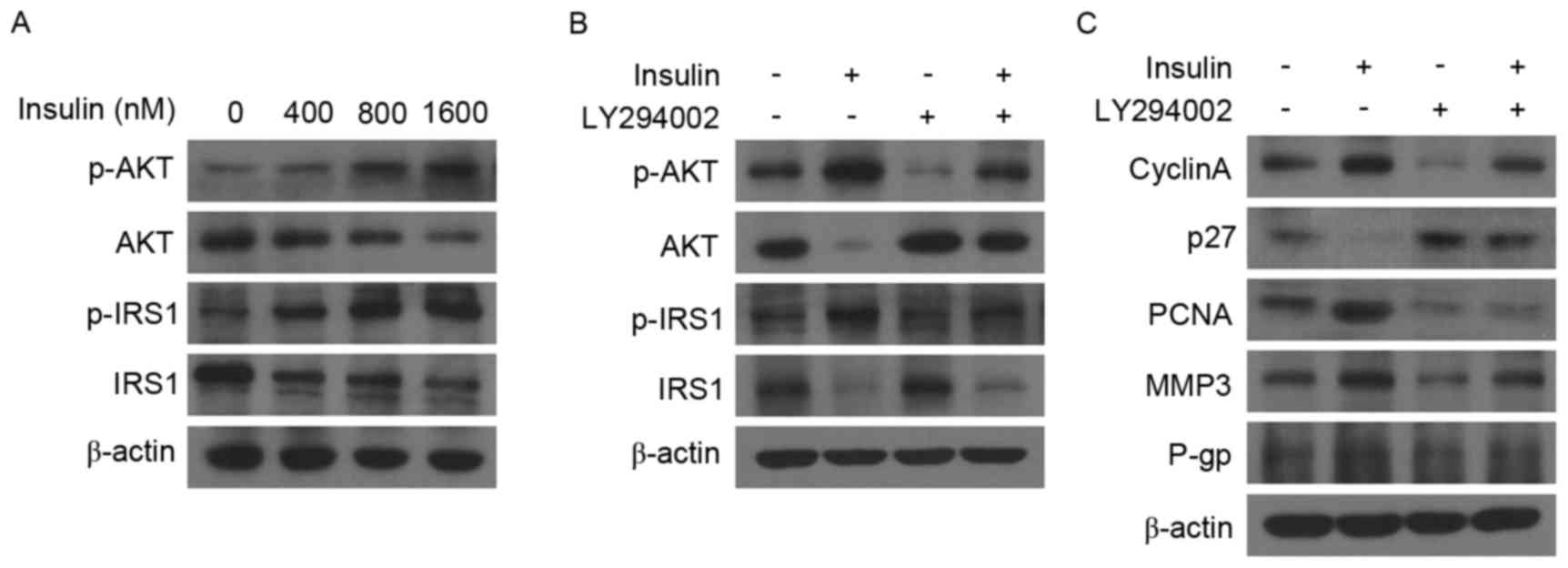 | Figure 4.Insulin regulates the PI3K/Akt
signaling pathway. (A) Expression of p-Akt, Akt, p-IRS1 and IRS1
were assessed by western blotting in A549 cells following treatment
with 0, 400, 800 and 1,600 nM insulin for 3 days. (B and C) Western
blot analysis of A549 cells exposed to 1,600 nM insulin and/or 1 µM
LY294002 for 3 days. PI3K, phosphoinositide 3-kinase; Akt, protein
kinase B; Akt, protein kinase B; p-, phosphorylated; IRS1, insulin
receptor substrate 1; PCNA, proliferating cell nuclear antigen;
MMP-3, matrix metalloproteinase 3; P-gp, P-glycoprotein. |
Discussion
The role of insulin in the pathogenesis of cancer
was first reported in the 1970's (22). Insulin acts as a growth factor and may
stimulate neoplastic development (23). In lung cancer, intracellular insulin
activity is an important factor affecting the progression of
low-grade human lung adenocarcinomas (24). Furthermore, inhaled insulin is
associated with an increased incidence of lung cancer among former
smokers (25).
The results of the present study revealed that
insulin exhibits oncogenic activity during the development of
NSCLC. Insulin markedly increased the proliferation and migration
of NSCLC cells and significantly decreased the sensitivity of NSCLC
cells to DDP. Cell cycle analysis demonstrated that insulin
decreased the proportion of cells in the G1 phase and increased the
proportion of cells in the S-phase. In addition, it was
demonstrated that insulin upregulated cyclin A and PCNA expression
and downregulated p27 expression in a dose-dependent manner.
Furthermore, the expression of MMP3 was markedly increased
following incubation with insulin in A549 cells. Lastly, insulin
enhanced the expression of P-gp in a dose-dependent manner in A549
cells.
The oncogenic activity of insulin has been reported
in other types of cancer tissues. Previous studies have
demonstrated that insulin may contribute to the proliferation and
survival of different types of cancer, including gastric, colon,
pancreatic, breast and bladder cancer (14,15,21,26).
It has also been demonstrated that insulin potentiates the ability
of lysophosphatidic acid to stimulate cell cycle progression and
DNA synthesis in MCF-7 breast cancer cells. The potentiating effect
of insulin appears to increase the expression level of cyclin E and
decrease the expression level of the cyclin-dependent kinase
inhibitor p27Kip1 (27).
Insulin enhances the sensitivity of gastric cancer cells to
5-fluorouracil and stimulates the migration of breast cancer cells
(14,15). Insulin is also able to accelerate the
metabolic capacities of cancer cells by performing dual regulation
of the glycolytic enzyme pyruvate kinase M2 (28).
Insulin binds to the insulin receptor and causes
phosphorylation of insulin receptor (InsR) substrate proteins,
followed by activation of downstream molecules including PI3K/Akt
and MAPK. The activated PI3K/Akt signaling pathway is involved in
various oncogenic functions, including the induction of cell
proliferation, migration and drug resistance (19). Previous studies have demonstrated that
insulin promotes the proliferation of pancreatic, bladder, breast
and colon cancer cells via the PI3K/Akt signaling pathway (13,14,21). In
NSCLC, overexpression of InsR predicts poor patient survival
(29). IRS1 is an adaptor protein for
insulin signaling and it has been suggested that IRS1 loss occurs
early on in the pathogenesis of NSCLC (30). In the present study, it was
demonstrated that insulin enhanced p-Akt and p-IRS1 expression in a
dose-dependent manner. Furthermore, inhibition of PI3K/Akt
activation by LY294002 significantly reversed the insulin-induced
increases in p-Akt, p-IRS1, cyclin A, PCNA, MMP3 and P-gp
expression. At the same time, levels of Akt, IRS1 and p27
expression, which were decreased by insulin, were increased
following treatment with LY294002. These results suggest that the
PI3K/Akt signaling pathway is crucial for the effect of insulin on
NSCLC.
In conclusion, the current study demonstrated that
insulin is an oncogenic factor in the development of NSCLC that may
work by activating the PI3K/Akt signaling pathway. Future studies
should investigate how to counteract the oncogenic activity of
insulin on NSCLC.
Acknowledgements
The current study was supported by the National
Natural Science Foundation of China (grant no. 81572394), the Joint
Research of Critical Diseases Foundation, Xiamen Science and
Technology Bureau, China (grant no. 3502Z20159013), the Natural
Science Foundation of Zhangzhou, Fujian, China (grant no.
ZZ2017J36) and the Youth Nursery Foundation of the Affiliated
Southeast Hospital of Xiamen University, Zhangzhou, Fujian, China
(grant no. 16Y019).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Herbst RS, Heymach JV and Lippman SM: Lung
cancer. N Eng J Med. 359:1367–1380. 2008. View Article : Google Scholar
|
|
3
|
Cappuzzo F, Ciuleanu T, Stelmakh L,
Cicenas S, Szczésna A, Juhász E, Esteban E, Molinier O, Brugger W,
Melezinek I, et al: Erlotinib as maintenance treatment in advanced
non-small-cell lung cancer: A multicentre, randomised,
placebo-controlled phase 3 study. Lancet Oncol. 11:521–529. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Spiro SG and Silvestri GA: One hundred
years of lung cancer. Am J Res Crit Care Med. 172:523–529. 2005.
View Article : Google Scholar
|
|
5
|
van Zandwijk N and Fong KM: Update in lung
cancer: Prologue to a modern review series. Respirology.
20:183–184. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wolin KY, Carson K and Colditz GA: Obesity
and cancer. Oncologist. 15:556–565. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Renehan AG, Tyson M, Egger M, Heller RF
and Zwahlen M: Body-mass index and incidence of cancer: A
systematic review and meta-analysis of prospective observational
studies. Lancet. 371:569–578. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mehran AE, Templeman NM, Brigidi GS, Lim
GE, Chu KY, Hu X, Botezelli JD, Asadi A, Hoffman BG, Kieffer TJ, et
al: Hyperinsulinemia drives diet-induced obesity independently of
brain insulin production. Cell Metab. 16:723–737. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tseng CH: Use of insulin and mortality
from breast cancer among taiwanese women with diabetes. J Diabetes
Res. 2015:6787562015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tseng CH: Diabetes, insulin use, smoking,
and pancreatic cancer mortality in Taiwan. Acta Diabetol.
50:879–886. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang L, Cai S, Teng Z, Zhao X, Chen X and
Bai X: Insulin therapy contributes to the increased risk of
colorectal cancer in diabetes patients: A meta-analysis. Diagn
Pathol. 8:1802013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tseng CH: Insulin use and smoking jointly
increase the risk of bladder cancer mortality in patients with type
2 diabetes. Clin Genitourin Cancer. 11:508–514. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chan MT, Lim GE, Skovsø S, Yang YH,
Albrecht T, Alejandro EU, Hoesli CA, Piret JM, Warnock GL and
Johnson JD: Effects of insulin on human pancreatic cancer
progression modeled in vitro. BMC Cancer. 14:8142014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tomas NM, Masur K, Piecha JC, Niggemann B
and Zänker KS: Akt and phospholipase Cgamma are involved in the
regulation of growth and migration of MDA-MB-468 breast cancer and
SW480 colon cancer cells when cultured with diabetogenic levels of
glucose and insulin. BMC Res Notes. 5:2142012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wei Z, Liang L, Junsong L, Rui C, Shuai C,
Guanglin Q, Shicai H, Zexing W, Jin W, Xiangming C and Shufeng W:
The impact of insulin on chemotherapeutic sensitivity to
5-fluorouracil in gastric cancer cell lines SGC7901, MKN45 and
MKN28. J Exp Clin Cancer Res. 34:642015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang C, Lan T, Hou J, Li J, Fang R, Yang
Z, Zhang M, Liu J and Liu B: NOX4 promotes non-small cell lung
cancer cell proliferation and metastasis through positive feedback
regulation of PI3K/Akt signaling. Oncotarget. 5:4392–4405. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sarker D, Reid AH, Yap TA and de Bono JS:
Targeting the PI3K/AKT pathway for the treatment of prostate
cancer. Clin Cancer Res. 15:4799–4805. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Safdari Y, Khalili M, Ebrahimzadeh MA,
Yazdani Y and Farajnia S: Natural inhibitors of PI3K/AKT signaling
in breast cancer: Emphasis on newly-discovered molecular mechanisms
of action. Pharmacol Res. 93:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Martini M, De Santis MC, Braccini L,
Gulluni F and Hirsch E: PI3K/AKT signaling pathway and cancer: An
updated review. Ann Med. 46:372–383. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang GL, Luo Q, Rui G, Zhang W, Zhang QY,
Chen QX and Shen DY: Oncogenic activity of retinoic acid receptor γ
is exhibited through activation of the Akt/NF-κB and Wnt/β-catenin
pathways in cholangiocarcinoma. Mol Cell Biol. 33:3416–3425. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu S, Li Y, Lin T, Fan X, Liang Y and
Heemann U: High dose human insulin and insulin glargine promote T24
bladder cancer cell proliferation via PI3K-independent activation
of Akt. Diabetes Res Clin Pract. 91:177–182. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kessler II: Cancer and diabetes mellitus.
A review of the literature. J Chronic Dis. 23:579–600. 1971.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Home P: Insulin therapy and cancer.
Diabetes Care. 36 Suppl 2:S240–S244. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mattarocci S, Abbruzzese C, Mileo AM,
Visca P, Antoniani B, Alessandrini G, Facciolo F, Felsani A,
Radulescu RT and Paggi MG: Intracellular presence of insulin and
its phosphorylated receptor in non-small cell lung cancer. J Cell
Physiol. 221:766–770. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kling J: Inhaled insulin's last gasp? Nat
Biotechnol. 26:479–480. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
White PB, True EM, Ziegler KM, Wang SS,
Swartz-Basile DA, Pitt HA and Zyromski NJ: Insulin, leptin, and
tumoral adipocytes promote murine pancreatic cancer growth. J
Gastrointest Surg. 14:1888–1894. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chappell J, Leitner JW, Solomon S,
Golovchenko I, Goalstone ML and Draznin B: Effect of insulin on
cell cycle progression in MCF-7 breast cancer cells. Direct and
potentiating influence. J Biol Chem. 276:38023–38028.
2001.PubMed/NCBI
|
|
28
|
Iqbal MA, Siddiqui FA, Gupta V,
Chattopadhyay S, Gopinath P, Kumar B, Manvati S, Chaman N and
Bamezai RN: Insulin enhances metabolic capacities of cancer cells
by dual regulation of glycolytic enzyme pyruvate kinase M2. Mol
Cancer. 12:722013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim JS, Kim ES, Liu D, Lee JJ, Solis L,
Behrens C, Lippman SM, Hong WK, Wistuba II and Lee HY: Prognostic
impact of insulin receptor expression on survival of patients with
nonsmall cell lung cancer. Cancer. 118:2454–2465. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Han CH, Cho JY, Moon JT, Kim HJ, Kim SK,
Shin DH, Chang J, Ahn CM, Kim SK and Chang YS: Clinical
significance of insulin receptor substrate-I down-regulation in
non-small cell lung cancer. Oncol Rep. 16:1205–1210.
2006.PubMed/NCBI
|