Introduction
To date, lung cancer has the highest cancer-related
mortality rates in females and male worldwide (1), and non-small cell lung cancer (NSCLC)
accounts for between 80 and 85% of all cases of lung cancer
(2). While a number of
chemotherapeutic agents mediate their effects by inducing the
apoptosis of cancer cells, metastasis remains a main factor
contributing to patient mortality. Previous studies have suggested
that metastasis may occur earlier than the formation of a primary
tumor (3,4). Many patients with NSCLC at the time of
initial diagnosis have aggressive and early metastasis, thus the
5-year survival rate of NSCLC is as low as 11% (5). Therefore, inhibiting the invasion and
migration of lung cancer should be considered as an efficient
therapeutic method for treating lung cancer, particularly prior to
detectable metastatic disease.
Metastasis is a multiple-steps process (6). Previous studies investigating its
underlying molecular mechanisms may provide specific drug targets
for controlling the spread of cancer. Vav guanine nucleotide
exchange factor 1 (Vav1), is a guanine nucleotide exchange factor
(GEF), serves a pivotal role in hematopoietic cells and has been
revealed to be ectopically expressed in numerous types of cancer,
including neuroblastoma (7), ovarian
cancer (8), breast cancer (9), pancreatic ductal adenocarcinoma (PDA)
(10), melanoma (11) and lung cancer (12). Conversely, Cell division cycle 42
(Cdc42), which is regularly activated by GEFs, is a family member
of small RhoGTPases and has also been revealed to serve a role in
cancer (13–16). In PDA, Vav1 GEF activity may
efficiently promote the formation of invadopodia and matrix
degradation by regulating the activation of Cdc42 (17). Also, it has been reported that Cdc42
is overexpressed in lung adenocarcinoma (18), and that the knockdown of Cdc42 was
associated with suppression of gastric cancer cell migration
(19). The present study investigated
whether Vav1 or Cdc42 may be an ideal drug target for the treatment
of lung cancer metastasis.
In the clinic, platinum-based drugs are the main
first-line treatment for lung cancer metastasis (20). However, toxicity and side effects,
including gastrointestinal and myeloid disorders, are inevitable.
Therefore, novel improved drugs targeting specific cancer
metastatic factors are urgently required. Of note, traditional
Chinese herbal medicines have received increasing interest
worldwide as anticancer agents (21).
There are studies suggesting that agents derived from natural plant
species possess the ability to inhibit cancer cell survival
independently (22). Certain natural
agents are able to enhance the anticancer effect of first-line
drugs (23,24).
Total alkaloids of Corydalis saxicola bunting
(TAOCSB) is a traditional Chinese herbal medicine and possesses
numerous biological activities, including anti-inflammatory,
antimicrobial, anticancer, immune-regulatory and central nervous
sedative effects (25–27). Previous studies have suggested that
TAOCSB has been used as an effective auxiliary curable method for
hepatoma (28). TAOCSB acquires the
ability to inhibit proliferation and induce apoptosis in a number
of cancer cell lines, including tongue squamous carcinoma, liver
cancer and ovarian cancer cells (29–31). The
present study demonstrated that TAOCSB facilitated apoptosis and
reversed the process of epithelial to mesenchymal transition (EMT)
in A549 lung cancer cells. In conclusion, TAOCSB is hypothesized to
suppress the metastasis of A549 cells. Further confirmatory studies
should investigate whether TAOCSB exerts its functions as suggested
in the present study and the possible underlying mechanisms
involved.
Materials and methods
Cell culture
A549 human NSCLC cells were purchased from Kunming
Cell Bank of the Chinese Academy of Sciences (Shanghai, China) and
were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% (v/v) fetal bovine serum (FBS; Gemini Bio Products, West
Sacramento, CA, USA) and 1% (v/v) penicillin/streptomycin
(Invitrogen; Thermo Fisher Scientific, Inc.) in a 5% CO2
humid atmosphere and saturated humidity at 37°C. Cells in the
logarithmic growth phase were used for subsequent experiments.
Reagents
MTT was purchased from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany). Rabbit anti-human Cdc42 monoclonal antibody
(ab187643) and mouse anti-human Vav1 monoclonal antibody (ab187643)
were obtained from Abcam (Cambridge, UK). Rabbit anti-human matrix
metalloproteinase (MMP)-9 monoclonal antibody (WL01580) and rabbit
anti-human MMP-2 monoclonal antibody (WL1579) supplied by Wanleibo
Co., Ltd. (Shanghai, China). ML141 (Cdc42 inhibitor; S7686) was
purchased from Selleck Chemicals (Houston, TX, USA). Transwell
chambers were obtained from Corning Incorporated (NY, USA). All the
primer sequences used for reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) were purchased from Generay
Biotech Co, Ltd. (Shanghai, China).
Cell viability assay
For all studies, A549 cells were incubated at 37°C
with 5% CO2 in a humidified atmosphere. TAOCSB was
dissolved in dimethylsulfoxide (DMSO), with DMSO-treatment serving
as a control. In this assay, cells were seeded into 96-well plates
at a density of ~2.5×104 cells/ml overnight.
Subsequently, each group of wells was treated with 0–40 µg/ml
TAOCSB at 37°C. After 24 h, 5.0 mg/ml MTT was added to each well.
After a further incubation for 4 h at 37°C, 150 µl DMSO was loaded
into each well and agitated to dissolve the crystals in the viable
cells. The optical density (OD) value was read at 490 nm using an
automated microplate reader. The results are presented as the mean
of all readings. Cell viability rate=OD of experimental group/OD of
control group ×100%. A total of 7 sets of experiments were
performed, each containing 6 wells per treatment. All experiments
were performed in triplicate.
Wound healing assay
A549 cells (15×104 per well) were plated
in 6-well plates. Once the cells in the individual wells had
achieved 80–90% confluence after ~16 h, a wound was created by
gently scratching the surface using a sterile pipette tip.
Subsequently, the cells were treated with various doses of TAOCSB
(0, 5, 7.5 or 10 µg/ml) at 37°C for 24 h. Cells were imaged at 0
and 24 h using a phase-contrast microscope (magnification, ×100).
The motility ratio of cells was evaluated by measuring the width of
the wound at an identical position. All experiments were performed
in triplicate.
Migration and invasion assays
Using Matrigel-coated chamber invasion plates, A549
cells (2×104 cells per well) were added to the upper
chamber in serum-free medium containing various doses of TAOCSB (0,
5, 7.5 or 10 µg/ml) and the lower chamber containing DMEM
supplemented with 10% FBS and TAOCSB (0, 5, 7.5 or 10 µg/ml). The
plates were incubated at 37°C for 24 h. The cells that invaded
through the Matrigel were fixed with pre-cooled methanol (100%),
washed with PBS at 37°C and stained with 0.1% crystal violet for 30
min at room temperature. The cells were subsequently counted under
a light microscope at a magnification of ×200. All experiments were
performed in triplicate.
Western blot analysis
A549 cells were seeded into 10 cm2
culture dishes, when cells were grown to a confluence of 70–80%.
The total protein in each group was collected after 24 h using
radioimmunoprecipitation assay lysis buffer (P0013K; Beyotime
Institute of Biotechnology, Haimen, China) on ice. Protein
concentration was determined using the bicinchoninic acid protein
assay kit (orb219872; Biorbyt Ltd., Cambridge, UK), according to
the manufacturer's protocol. For each sample, 30 µg protein was
separated by SDS-PAGE (10% gel) and transferred onto a
polyvinylidene fluoride membrane. The membrane was blocked in
Blocking buffer (Beyotime Institute of Biotechnology) at room
temperature for 1 h, then incubated with Cdc42 (1:1,000), MMP-2
(1:1,000), MMP-9 (1:2,000) and β-actin (1:2,000; Origene
Technologies, Inc., Beijing, China; cat. no. TA-09) primary
antibodies at 4°C overnight. Following washing with Tris-buffered
saline and Tris-buffered saline containing Tween-20 buffers,
antibody binding was detected using horseradish
peroxidase-conjugated goat anti-mouse IgG secondary antibody
(AP124P, dilution 1:5,000) and goat anti-rabbit IgG secondary
antibody (A0545, dilution 1:5,000) at room temperature for 1 h. The
binding bands were visualized using enhanced chemiluminescence
(orb90504; Biorbyt Ltd.) and autoradiography. Protein bands were
analyzed using Quantity One 1-D analysis software (Version 4.6.2,
Bio-Rad Laboratories, Inc., Hercules, CA, USA) and normalized to
the β-actin signal.
RT-qPCR analysis
Total mRNA was isolated from A549 cells in the
exponential growth phase using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. RNA (1 µg per reaction) was
reverse-transcribed to yield first-strand cDNA using transcriptase
(DBI Bioscience, Shanghai, China). Diluted cDNA was then mixed with
pairs of specific primers and SYBR Green PCR Master Mix (DBI
Bioscience) in a total volume of 15 µl. The PCR cycling conditions
were as follows: 2 min at 95°C for 1 cycle, 10 sec at 95°C, 34 sec
at 55°C and 30 sec at 72°C for 40 cycles. Relative mRNA levels were
calculated based on Cq values and corrected for the 18S expression
according to the equation 2−ΔΔCq (32). The sequences of the primers were as
follows: Cdc42 sense, 5′-TTTCTTGCTTGTTGGGACT-3′ and antisense,
5′-GGCTTCTGTTTGTTCTTGG-3′; Vav1 sense, 5′-AAGCGAGACAACGAGACA−3′ and
antisense, 5′-GCCATAGTGAGCCAGAGA-3′; β-actin sense,
5′-GACATCCGCAAAGACCTG-3′ and antisense, 5′-GGAAGGTGGACAGCGAG-3′.
The analysis was performed using the ABI Prism 7500 Real-Time PCR
technology (Applied Biosystems; Thermo Fisher Scientific, Inc.).
Each cDNA sample was run three times independently. The
quantification of the expression of the detectable gene was
normalized to that of β-actin in corresponding samples.
Statistical analysis
SPSS (version 18.0; SPSS, Inc., Chicago, IL, USA)
for Windows was used for all analyses. All quantitative data are
presented as the mean value ± standard deviation of three
independent experiments. The differences between the groups were
analyzed by one-way analysis of variance followed by Tukey's post
hoc test. P<0.05 or P<0.01 were considered to indicate
statistically significant differences.
Results
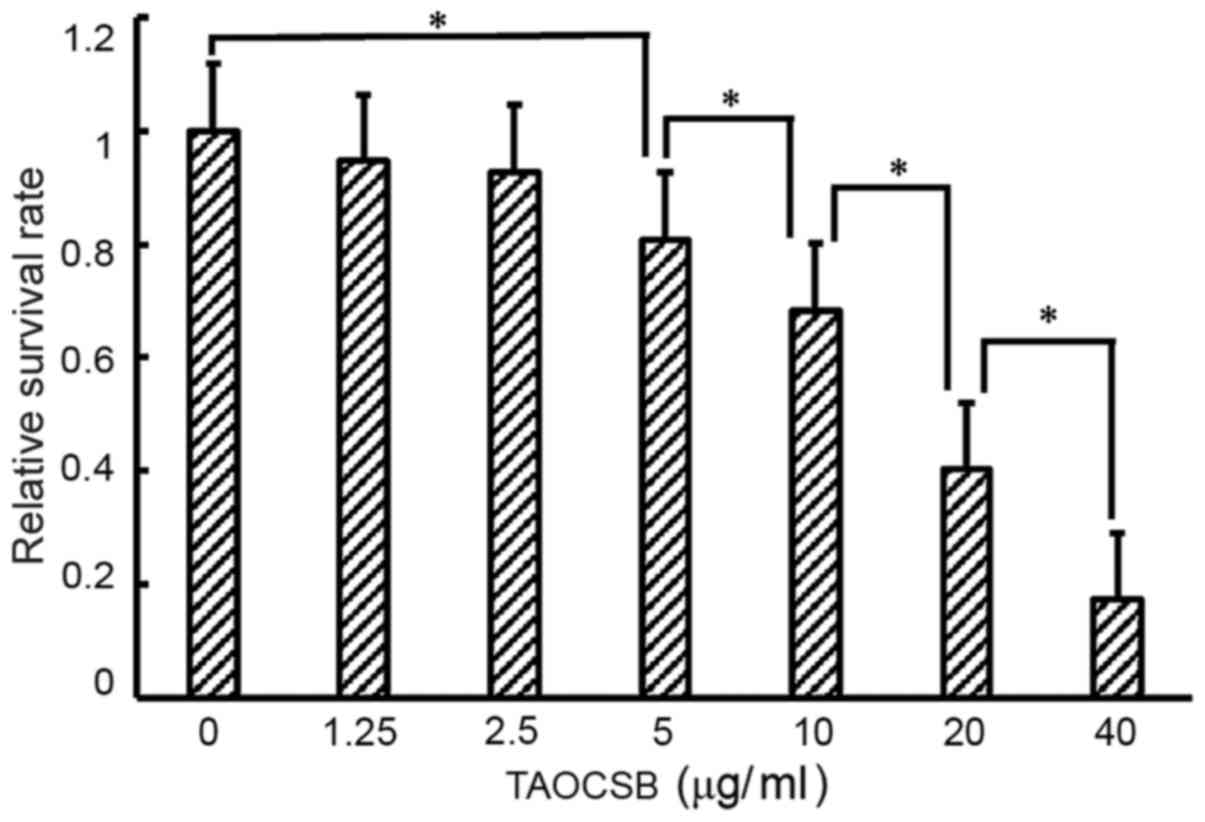
Effects of various concentrations of
TAOCSB on the viability of A549 cells
Treatment of A549 cells with various concentration
of TAOCSB for 24 h resulted in dose-dependent growth inhibition
(Fig. 1). For treatment with 10 µg/ml
TAOCSB, the relative cell survival rate of A549 cells was ~40%.
Therefore, using a concentration below 10 µg/ml may inhibit cell
migration or invasion in a pathological state.
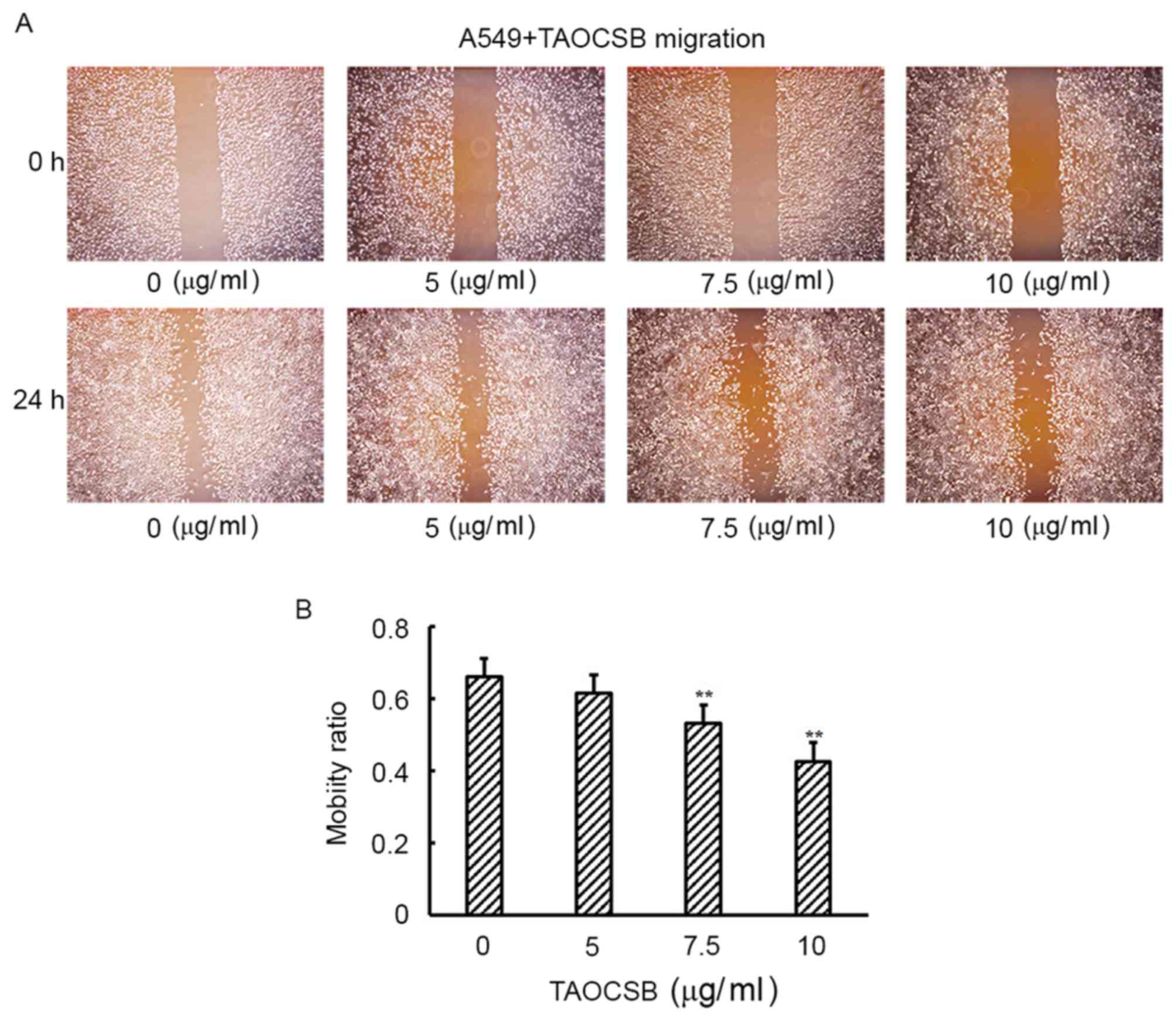
TAOCSB inhibits the migration and
invasion of A549 cells in vitro
Increased cell motility and invasion of carcinoma
cells are key steps in the process of metastasis. The present study
subsequently examined the effect of TAOCSB on the ability of A549
cells to migrate and invade by wound healing and Transwell assays,
respectively. Wound healing experiment results revealed that
TAOCSB-treated A549 cells exhibited lower mobility rates compared
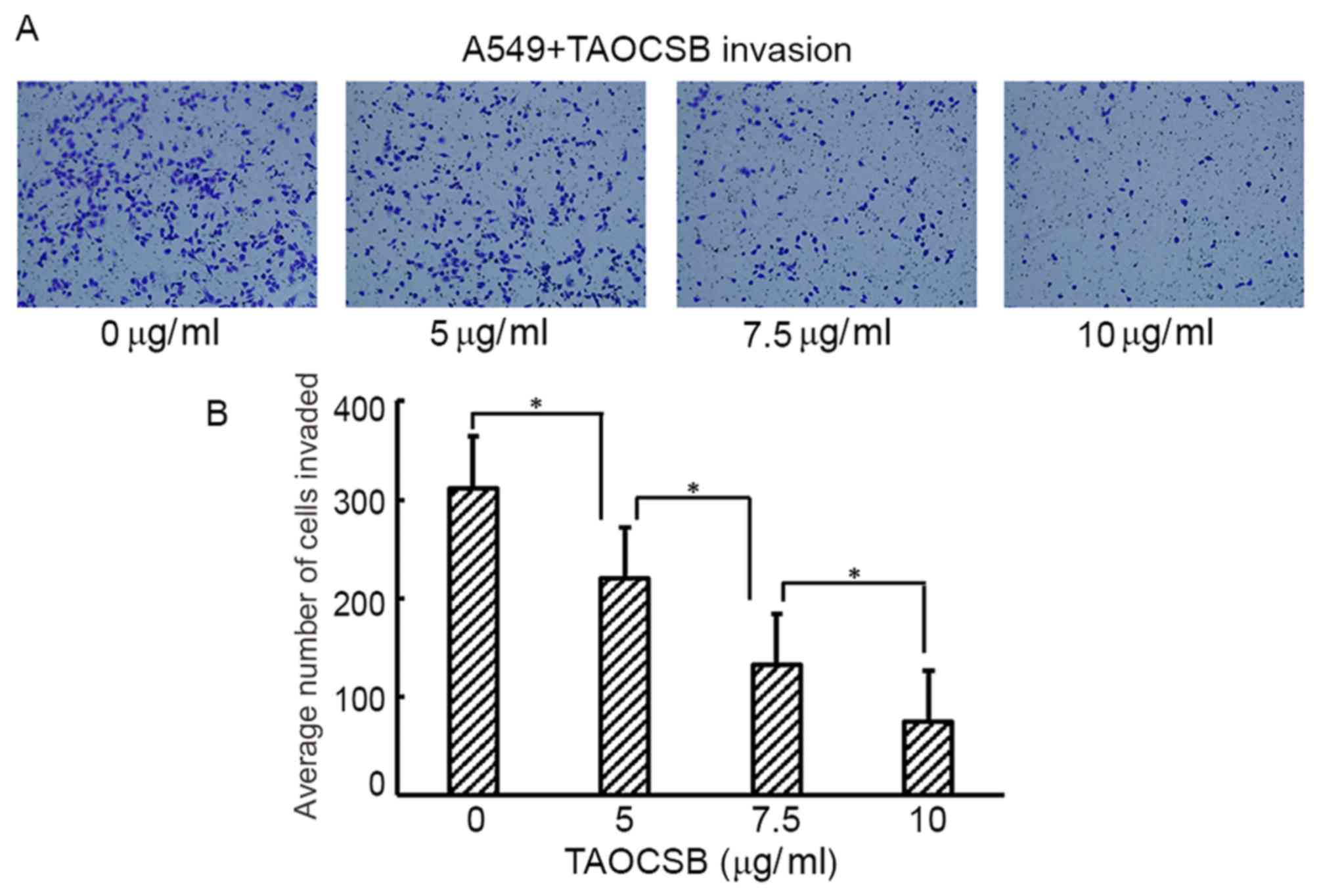
with the control in a dose-dependent manner (Fig. 2A and B). Consistent with these
results, the number of TAOCSB-treated A549 cells that invaded was
also significantly lower compared with the control (Fig. 3A), and the quantification of cells in
the lower chamber demonstrated statistical significance (Fig. 3B). These results demonstrated the
inhibitory effect of TAOCSB on the rates of migration and invasion
in A549 lung cancer cells.
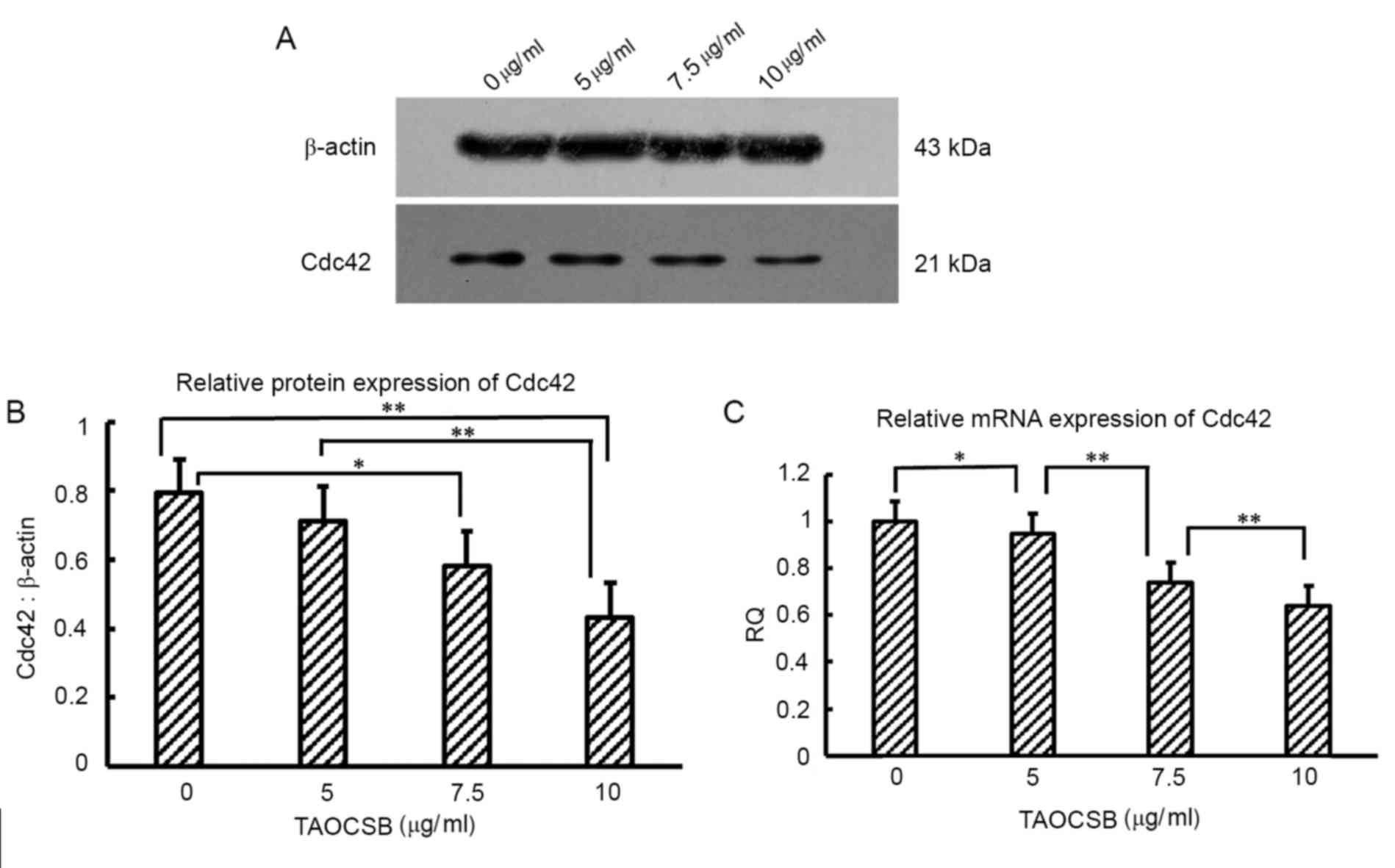
TAOCSB inhibits Cdc42 expression in
A549 cells
For Cdc42, promoting the formation of invadopodia
structures contributes to enhanced metastasis. The present study
therefore investigated whether the effects reported in A549 cells
were associated with variations in the level of Cdc42 expression by
performing RT-qPCR at the mRNA level and western blot analysis at
the protein level. In untreated cells, the expression levels of
Cdc42 protein and mRNA were high (Fig.
4). The mRNA and protein expression level of Cdc42 was
investigated following the treatment with 5–10 µg/ml TAOCSB. These
results revealed a decrease in the expression of Cdc42 at the
protein and mRNA levels in a dose-dependent manner.
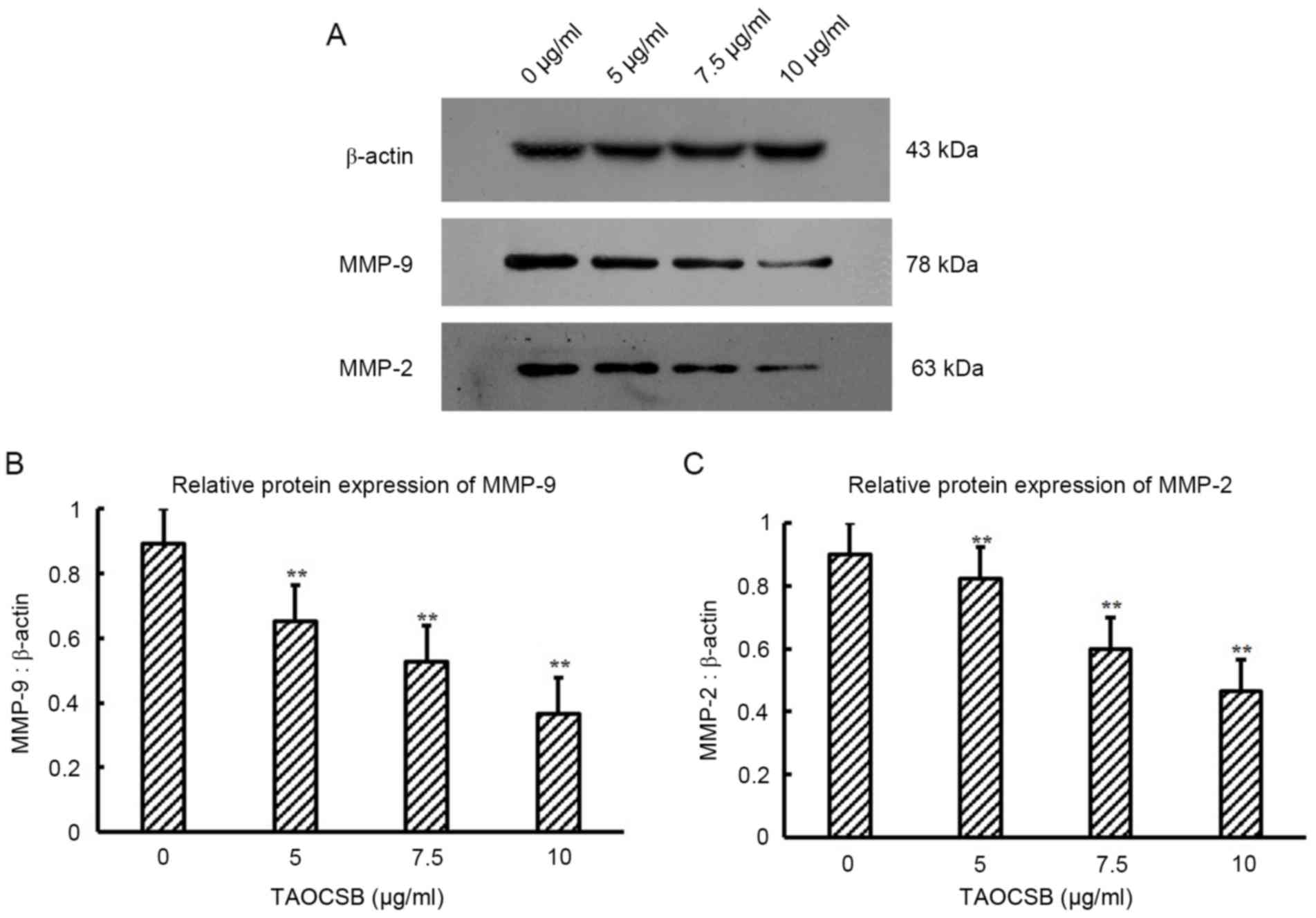
TAOCSB inhibits the expression levels
of MMP-9 and MMP-2 in A549 cells
Cdc42 is involved in the activation of MMPs
responsible for extracellular matrix digestion, which is required
for tumor cell invasion. Therefore, the present study evaluated
whether TAOCSB-mediated decrease in Cdc42 protein expression level
in A549 cells was paralleled by variations in the levels of MMP-2
and MMP-9 protein expression. The results revealed that treatment
with TAOCSB was able to markedly inhibit the protein expression
levels of MMP-2 and MMP-9 in A549 cells (Fig. 5). Furthermore, the decrease in MMP-2
and MMP-9 expression levels mediated by TAOCSB was
dose-dependent.
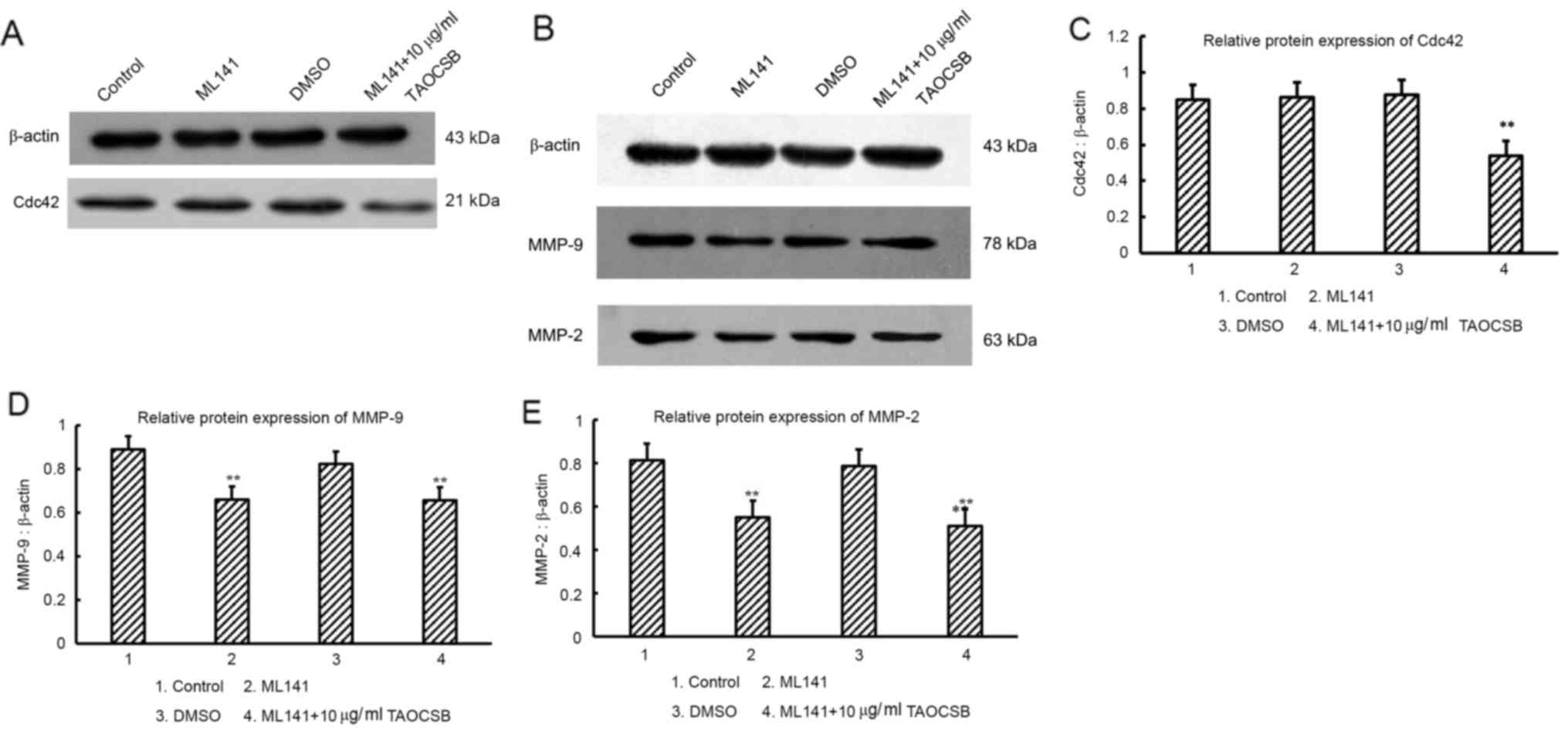
Variant analysis of the levels of
MMP-9 and MMP-2 protein expression following cdc42 inhibitor
(ML141) treatment
Although it has been demonstrated that TAOCSB may
decrease the expression levels of Cdc42, MMP-2 and MMP-9, whether
TAOCSB is able to selectively inhibit the levels of total cdc42
protein, which leads to defective secretion of MMP-9 and MMP-2 in
A549 cells, required further investigation.
For further experiments, a new specific inhibitor of
Cdc42 (ML141) was used. ML141 is a Cdc42 selective non-competitive
inhibitor that prevents Cdc42 from bounding to guanosine
triphosphate (GTP), which leads to inactive Cdc42 and reduction in
the secretion of MMPs (33). As a
result, there appeared to be no significant difference in MMP-9 and
MMP-2 protein expression levels among ML141 and ML141 plus
TAOCSB-treated groups (Fig. 6).
Therefore, TAOCSB had no direct inhibitory effect on the expression
levels of MMP-9 and MMP-2 proteins, the decrease of which may
result from inhibition of active Cdc42-GTP by ML141. It was
suggested that Cdc42 or factors upstream of Cdc42 may be the drug
target for TAOCSB in A549 cells, and TAOCSB-mediated inhibition of
total Cdc42 may also limit the Cdc42-GTP bound state (Fig. 6).
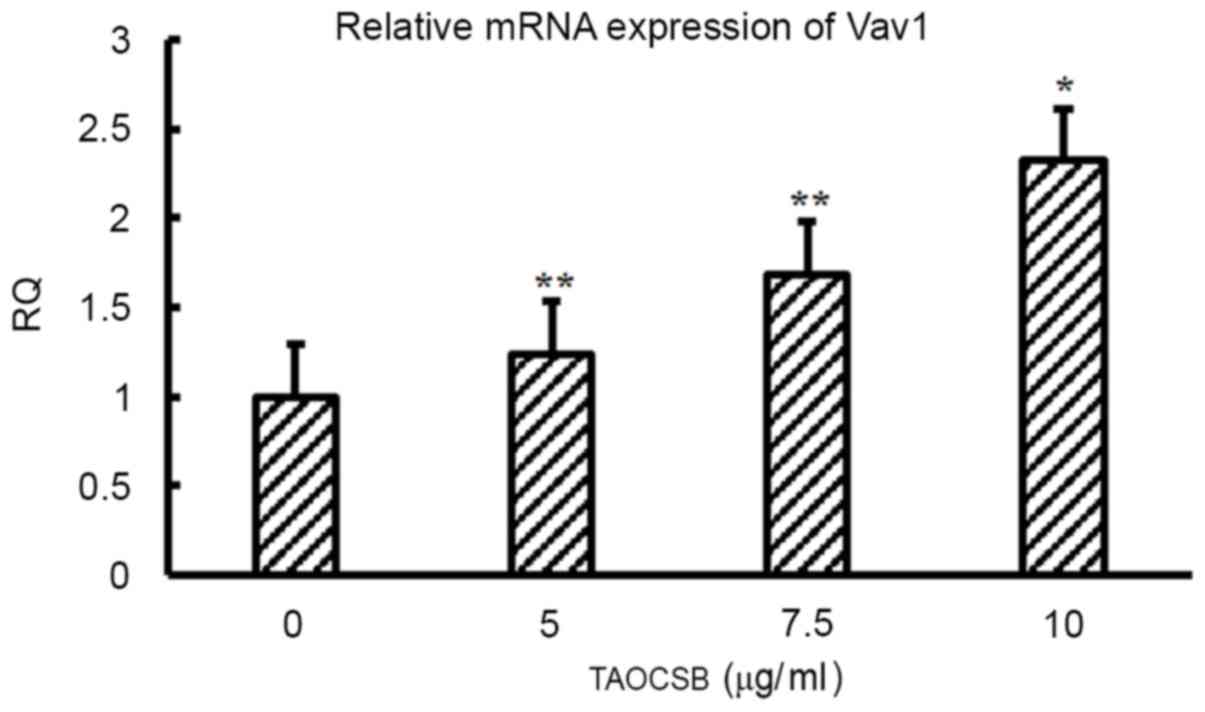
Treatment with TAOCSB affects the
level of Vav1 expression in A549 cells
It was revealed that Vav1 may promote metastasis by
activating Cdc42 in PDA. Therefore, the present study suggested
that the same association between Vav1 and Cdc42 may exist in
NSCLC. Of note, the mRNA expression level of Vav1 was identified,
and treatment with TAOCSB may increase the level of Vav1 expression
(Fig. 7).
Discussion
In a previous study, it was demonstrated that TAOCSB
was able to inhibit the proliferation of A549 lung cancer cells,
and this inhibitory effect was associated with cell cycle arrest at
the G1/S phase, as well as induction of apoptosis via
increasing the expression levels of caspase-3 and decreasing
survivin expression (34).
Subsequently, with suppression of the snail gene and increased
E-cadherin protein expression, TAOCSB may have reversed the effect
of EMT. The present study aimed to investigate the effects of
TAOCSB on the metastasis of NSCLC A549 cells and the associated
underlying mechanisms.
Cdc42 is overexpressed in various types of cancer
(35,36). In lung cancer, Cdc42 is overexpressed
in lung adenocarcinoma (18). A
recent study demonstrated that the knockdown of Cdc42 may markedly
suppress the migration and invasion of gastric cancer cells
(19). Another previous study
revealed that the overexpression of Cdc42 in lung cancer cell line
801D was downregulated by curcumin (24). Additionally, Vav1 as a GEF serves a
pivotal role in the development of cancer, which is particularly
associated with cancer metastasis (17). In PDA, Cdc42 was revealed to be a main
downstream factor for Vav1, rather than other RAS superfamily
members. However, targeting Vav1 may efficiently suppress the
formation of invadopodia and matrix degradation (37).
Degradation of the basement membrane and
extracellular matrix is an essential step for cancer cells to
escape from the primary tumor site. MMP-9 and MMP-2, as a class of
zinc-dependent enzymes, serve a pivotal role in this process
(38,39). Conversely, restricted expression
levels of MMP-2 and MMP-9 in cancer cells leads to an improved
prognosis (40–42). According to a number of studies, Cdc42
is involved in the formation and stabilization of invadopodia
structures in cancer cells (43,44).
Additionally, Cdc42 may mediate the secretion of MMPs to enhance
the function of invadopodia (45).
To the best of our knowledge, the present study
revealed for the first time the inhibitory effect of TAOCSB on
proliferation and migration of A549 lung cancer cells. By detecting
variations of treatment conditions of the critical factor, Cdc42,
in TAOCSB-treated A549 cells, the present study demonstrated that a
low dose of TAOCSB was sufficient to reduce the expression level of
Cdc42 at the level of mRNA and protein. In addition, the protein
expression levels of MMP-2 and MMP-9 were also attenuated following
decreased Cdc42 expression. To verify whether TAOCSB directly
reduced MMP expression, the present study used the inhibitor of
Cdc42 (ML141) in further experiments. It was revealed that TAOCSB
did not directly inhibit the expression levels of MMP-9 and MMP-2
proteins. It remains unclear whether TAOCSB targets Cdc42 or its
upstream factors directly to mediate the observed decrease in MMP-9
and MMP-2 expression levels and the reduced invasiveness of A549
cells. The present study did not investigate the protein expression
level of Vav1 in A549 cells. However, it was hypothesized that Vav1
expression may be deregulated rapidly by calcineurin B-like
proteins in A549 cells, which is similar to the effect observed in
AU565 breast cancer cells (46). The
present study determined Vav1 expression at the mRNA level.
Although Vav1 is usually associated with matrix degradation and the
formation of invadopodia, the anticancer effect of Vav1 remains
uncertain; however, in the present study, it was identified that
TAOCSB had a promoting effect on Vav1 mRNA expression. The activity
of Vav may depend on additional genetic aberrations, including the
p53 signaling pathway (46).
Conversely, Grassilli et al (47) demonstrated high nuclear expression
level of Vav1 in breast cancer tissues without lymphoma metastasis
using the immunohistochemical technique. Compared with patients
with high expression levels of Vav1, patients with low expression
levels of Vav1 had a lower risk of distant metastasis. This
observation may indicate that the risk of metastasis was associated
with the distribution patterns of Vav1 protein in cancer cells.
In conclusion, it was identified that TAOCSB
inhibited the proliferation and migration of A549 cells. It was
also revealed that TAOCSB downregulated Cdc42 gene and protein
expression, Cdc42 upstream factor Vav1, and downstream factors MMP2
and MMP9 expression in lung cancer cells. These results suggest
that TAOCSB may inhibit tumor growth and invasion through the Cdc42
signal pathway. Further studies should be performed to uncover the
role of Vav1 in cancer and to resolve the underlying molecular
mechanisms in the pathogenesis of cancer.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81541172), Guangxi
Health Department of Traditional Chinese Medicine (grant no.
GZLC14-37), Traditional Chinese Medicine Science and Technology
Projects of Guangxi Zhuang Autonomous Region Health Department
(grant no. GZPT13-46).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Subramaniam S, Thakur RK, Yadav VK, Nanda
R, Chowdhury S and Agrawal A: Lung cancer biomarkers: State of the
art. J Carcinog. 12:32013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rhim AD, Mirek ET, Aiello NM, Maitra A,
Bailey JM, McAllister F, Reichert M, Beatty GL, Rustgi AK,
Vonderheide RH, et al: EMT and dissemination precede pancreatic
tumor formation. Cell. 148:349–361. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Haeno H, Gonen M, Davis MB, Herman JM,
Iacobuzio-Donahue CA and Michor F: Computational modeling of
pancreatic cancer reveals kinetics of metastasis suggesting optimum
treatment strategies. Cell. 148:362–375. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Verdecchia A, Francisci S, Brenner H,
Gatta G, Micheli A, Mangone L and Kunkler I; EUROCARE-4 Working
Group, : Recent cancer survival in Europe: A 2000-02 period
analysis of EUROCARE-4 data. Lancet Oncol. 8:784–796. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yilmaz M and Christofori G: Mechanisms of
motility in metastasizing cells. Mol Cancer Res. 8:629–642. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hornstein I, Pikarsky E, Groysman M, Amir
G, Peylan-Ramu N and Katzav S: The haematopoietic specific signal
transducer Vav1 is expressed in a subset of human neuroblastomas. J
Pathol. 199:526–533. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wakahashi S, Sudo T, Oka N, Ueno S,
Yamaguchi S, Fujiwara K, Ohbayashi C and Nishimura R: Vav1
represses E-cadherin expression through the transactivation of
Snail and Slug: A potential mechanism for aberrant epithelial to
mesenchymal transition in human epithelial ovarian cancer. Transl
Res. 162:181–190. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Grassilli S, Brugnoli F, Lattanzio R,
Rossi C, Perracchio L, Mottolese M, Marchisio M, Palomba M, Nika E,
Natali PG, et al: High nuclear level of Vav1 is positive prognostic
factor in early invasive breast tumors: A role in modulating genes
related to the efficiency of metastatic process. Oncotarget.
5:4320–4336. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fernandez-Zapico ME, Gonzalez-Paz NC,
Weiss E, Savoy DN, Molina JR, Fonseca R, Smyrk TC, Chari ST,
Urrutia R and Billadeau DD: Ectopic expression of VAV1 reveals an
unexpected role in pancreatic cancer tumorigenesis. Cancer Cell.
7:39–49. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bartolomé RA, Molina-Ortiz I, Samaniego R,
Sánchez-Mateos P, Bustelo XR and Teixidó J: Activation of Vav/Rho
GTPase signaling by CXCL12 controls membrane-type matrix
metalloproteinase-dependent melanoma cell invasion. Cancer Res.
66:248–258. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lazer G, Idelchuk Y, Schapira V, Pikarsky
E and Katzav S: The haematopoietic specific signal transducer Vav1
is aberrantly expressed in lung cancer and plays a role in
tumourigenesis. J Pathol. 219:25–34. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu Y, Wang Y, Zhang Y, Miao Y, Zhao Y,
Zhang PX, Jiang GY, Zhang JY, Han Y, Lin XY, et al: Abnormal
expression of p120-catenin, E-cadherin, and small GTPases is
significantly associated with malignant phenotype of human lung
cancer. Lung Cancer. 63:375–382. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ye H, Zhang Y, Geng L and Li Z: Cdc42
expression in cervical cancer and its effect on cervical tumor
invasion and migration. Int J Oncol. 46:757–763. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kamai T, Yamanishi T, Shirataki H, Takagi
K, Asami H, Ito Y and Yoshida K: Over-expression of RhoA, Rac1, and
Cdc42 GTPases is associated with progression in testicular cancer.
Clin Cancer Res. 10:4799–4805. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kikuchi K, Li X, Zheng Y and Takano Y:
Invasion of breast cancer cells into collage matrix requires TGF-α
and Cdc42 signaling. FEBS Lett. 585:286–290. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Razidlo GL, Schroeder B, Chen J, Billadeau
DD and McNiven MA: Vav1 as a central regulator of invadopodia
assembly. Curr Biol. 24:86–93. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yao R, Wang Y, Lubet RA and You M:
Differentially expressed genes associated with mouse lung tumor
progression. Oncogene. 21:5814–5821. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Du DS, Yang XZ, Wang Q, Dai WJ, Kuai WX,
Liu YL, Chu D and Tang XJ: Effects of CDC42 on the proliferation
and invasion of gastric cancer cells. Mol Med Rep. 13:550–554.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zarogoulidis K, Zarogoulidis P, Darwiche
K, Boutsikou E, Machairiotis N, Tsakiridis K, Katsikogiannis N,
Kougioumtzi I, Karapantzos I, Huang H and Spyratos D: Treatment of
non-small cell lung cancer (NSCLC). J Thorac Dis. 5 Suppl
4:S389–S396. 2013.PubMed/NCBI
|
|
21
|
Ma L, Wang B, Long Y and Li H: Effect of
traditional Chinese medicine combined with Western therapy on
primary hepatic carcinoma: A systematic review with meta-analysis.
Front Med. 11:191–202. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen QY, Jiao DM, Yao QH, Yan J, Song J,
Chen FY, Lu GH and Zhou JY: Expression analysis of Cdc42 in lung
cancer and modulation of its expression by curcumin in lung cancer
cell lines. Int J Oncol. 40:1561–1568. 2012.PubMed/NCBI
|
|
23
|
Refaat A, Abdelhamed S, Yagita H, Inoue H,
Yokoyama S, Hayakawa Y and Saiki I: Berberine enhances tumor
necrosis factor-related apoptosis-inducing ligand-mediated
apoptosis in breast cancer. Oncol Lett. 6:840–844. 2013.PubMed/NCBI
|
|
24
|
Ma L, Wang R, Nan Y, Li W, Wang Q and Jin
F: Phloretin exhibits an anticancer effect and enhances the
anticancer ability of cisplatin on non-small cell lung cancer cell
lines by regulating expression of apoptotic pathways and matrix
metalloproteinases. Int J Oncol. 48:843–853. 2016.PubMed/NCBI
|
|
25
|
Li L: The effect of Corydalis saxicola
Bunting on anti-inflammation. Chin J Ethnomed Ethnopharm. 18:20–21.
2009.(In Chinese).
|
|
26
|
Xie PS, Li AY, Zhou F and Zhao Y: The
experimental screening of Chinese herbal medicine in anti-cancer
effect. ShiZhen Chin Med. 19–20. 1996.(In Chinese).
|
|
27
|
Dong K, Wu LZ and Liang YY: The role of
Corydalis saxicola alkaloids in mice immunity. Chin J Immunol.
238–241. 1995.(In Chinese).
|
|
28
|
Xiong LY and Li LL: The clinical trials of
Octreotide combined Corydalis saxicola Bunting injection to treat
advanced liver cancer. Sichuan Journal of Cancer Control. 232–234.
2005.(In Chinese).
|
|
29
|
Wu CF, Liu W, Li FL and Xu YM: The effect
of Corydalis Saxicola bunting total alkaloids (CABTA) on monoamine
neurotransmitters in rat brain. J Shenyang Pharm Univ. 101–104.
1994.(In Chinese).
|
|
30
|
Yin JK and Liao JX: The role of Corydalis
Saxicola bunting total alkaloids on Tca8113 cells proliferation and
apoptosis. Chin J of Oral and Maxillofacial Surg. 4:245–248.
2010.(In Chinese).
|
|
31
|
Tang CL, Zheng H, Wang J, Song H, Lu SY,
Cheng B, Wu F, Zhang HY, Ruan JX, Liang YH and Su ZH: The active
component identification of herba Corydalis saxicolae extract base
on the quantitative composition-activity relationship in inhibition
of human liver cancer cells SMMC-7721. ShiZhen Chin Med.
27:2372–2375. 2016.(In Chinese).
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hong L, Kenney SR, Phillips GK, Simpson D,
Schroeder CE, Nöth J, Romero E, Swanson S, Waller A, Strouse JJ, et
al: Characterization of a Cdc42 protein inhibitor and its use as a
molecular probe. J Biol Chem. 288:8531–8543. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jin Hua, LI Ji, Ying Wang and Rong Zeng
Jing: Total alkaloids of Corydalis saxicola bunting on A549 lung
cancer cells proliferation, apoptosis and its effect on Caspase,
Survivin expression. Chin J Exp Formulas Chin Med Med Exp Formulas
Chin Med. 9:165–169. 2015.(In Chinese).
|
|
35
|
Ellenbroek SI and Collard JG: Rho GTPases:
Functions and association with cancer. Clin Exp Metastasis.
24:657–672. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vega FM and Ridley AJ: Rho GTPases in
cancer cell biology. FEBS Lett. 582:2093–2101. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Razidlo GL, Magnine C, Sletten AC, Hurley
RM, Almada LL, Fernandez-Zapico ME, Ji B and McNiven MA: Targeting
pancreatic cancer metastasis by inhibition of Vav1, a driver of
tumor cell invasion. Cancer Res. 75:2907–2915. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Borkham-Kamphorst E, Alexi P, Tihaa L,
Haas U and Weiskirchen R: Platelet-derived growth factor-D
modulates extracellular matrix homeostasis and remodeling through
TIMP-1 induction and attenuation of MMP-2 and MMP-9 gelatinase
activities. Biochem Biophys Res Commun. 457:307–313. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Davis ME, Gumucio JP, Sugg KB, Bedi A and
Mendias CL: MMP inhibition as a potential method to augment the
healing of skeletal muscle and tendon extracellular matrix. J Appl
Physiol (1985). 115:884–891. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang R, Ke ZF, Wang F, Zhang WH, Wang YF,
Li SH and Wang LT: GOLPH3 overexpression is closely correlated with
poor prognosis in human non-small cell lung cancer and mediates its
metastasis through upregulating MMP-2 and MMP-9. Cell Physiol
Biochem. 35:969–982. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu X, Chen L, Xu B, Xie Q, Sun M, Deng X,
Wu C and Jiang J: Increased MT2-MMP expression in gastric cancer
patients is associated with poor prognosis. Int J Clin Exp Pathol.
8:1985–1990. 2015.PubMed/NCBI
|
|
42
|
El-Badrawy MK, Yousef AM, Shaalan D and
Elsamanoudy AZ: Matrix metalloproteinase-9 expression in lung
cancer patients and its relation to serum MMP-9 activity,
pathologic type, and prognosis. J Bronchology Interv Pulmonol.
21:327–334. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Buccione R, Caldieri G and Ayala I:
Invadopodia: Specialized tumor cell structures for the focal
degradation of the extracellular matrix. Cancer Metastasis Rev.
28:137–149. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yamaguchi H, Lorenz M, Kempiak S,
Sarmiento C, Coniglio S, Symons M, Segall J, Eddy R, Miki H,
Takenawa T and Condeelis J: Molecular mechanisms of invadopodium
formation: The role of the N-WASP-Arp2/3 complex pathway and
cofilin. J Cell Biol. 168:441–452. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Poincloux R, Lizárraga F and Chavrier P:
Matrix invasion by tumour cells: A focus on MT1-MMP trafficking to
invadopodia. J Cell Sci. 122:3015–3024. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sebban S, Farago M, Gashai D, Ilan L,
Pikarsky E, Ben-Porath I and Katzav S: Vav1 fine tunes p53 control
of apoptosis versus proliferation in breast cancer. PLoS One.
8:e543212013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Grassilli S, Brugnoli F, Lattanzio R,
Rossi C, Perracchio L, Mottolese M, Marchisio M, Palomba M, Nika E,
Natali PG, et al: High nuclear level of Vav1 is a positive
prognostic factor in early invasive breast tumors: A role in
modulating genes related to the efficiency of metastatic process.
Oncotarget. 5:4320–4336. 2014. View Article : Google Scholar : PubMed/NCBI
|