Introduction
Lung cancer is the leading cause of
cancer-associated mortality in numerous countries, and the
incidences of morbidity and mortality associated with the disease
are increasing; lung cancer was responsible for >160,000
mortalities in the United States in 2010 (1). Among all the types of lung cancer,
non-small cell lung cancer (NSCLC) is the most common histological
subtype, accounting for 80–90% of all cases (2). Although multiple effective treatments,
including radiotherapy, chemotherapy and immunotherapy, have
recently become available for the management of locally confined
NSCLC, these treatments have been unable to reduce the high
mortality rate among patients with advanced-stage NSCLC (1). Thus, it has become increasingly
important to identify methods to diagnose early-stage lung cancer
with high sensitivity and predict clinical outcome.
In recent years, numerous studies have focused on
the association between carcinogenesis and small non-coding
regulatory RNAs, including microRNAs (miRNAs), small interfering
RNAs (siRNAs) and piwi-interacting RNAs (piRNAs) (3–5). Small
non-coding regulatory RNAs may have key roles in carcinogenesis.
The most extensively researched small non-coding regulatory RNAs
are miRNAs, which can act as either oncogenes or tumor suppressors,
according to the function of their target genes (6–8). Previous
studies have confirmed that the up- or downregulation of certain
miRNAs may contribute to human carcinogenesis and cancer
progression (9,10). However, research regarding piRNAs,
which are a novel type of small non-coding RNA with lengths of
26–31 nucleotides, remains insufficient (11,12).
piRNAs serve biological roles through their specific associations
with the piwi proteins (13,14). piRNAs aid the maintenance of DNA
integrity, epigenetic regulation, germ line stem cell
differentiation, embryonic development and disease occurrence and
development. Previous studies have confirmed that piRNAs can also
serve similar roles to miRNAs, acting as oncogenes or tumor
suppressors in several types of cancer (15–17),
including those of the cervix (18),
bladder (19), lung (20), gastrointestinal tract (21), breast (22,23) and
liver (24). It was previously
reported that the piRNA piR651 is overexpressed in several types of
human cancer tissue, including gastric, lung, colon, breast, and
multiple myeloma cancer tissues, compared with paired adjacent
normal tissues (21,22). Furthermore, piR651 expression levels
in gastric cancer tissues are associated with tumor-node-metastasis
(TNM) stage (25,26). The overexpression of piR651 has been
demonstrated in several cancer cell lines, including those of the
lung, gastric, mesothelium, cervix, breast and liver (21). These findings indicate that piR651 may
serve an oncogenic role in carcinogenesis. However, the mechanism
by which piR651 regulates carcinogenesis is unclear. Therefore, the
present study aimed to elucidate the mechanisms of action of piR651
in NSCLC.
Materials and methods
Cell culture
The human NSCLC A549 and HCC827 cell lines were
purchased from the Fourth Affiliated Hospital of Harbin Medical
University (Harbin, China) and the Shanghai Institute of
Biochemistry and Cell Biology, Chinese Academy of Science
(Shanghai, China), respectively. HCC827 and A549 cells were
cultured in RPMI-1640 (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and high-glucose Dulbecco's modified Eagle's
medium (DMEM; Invitrogen; Thermo Fisher Scientific, Inc.),
respectively, supplemented with 100 U/ml penicillin, 100 µg/ml
streptomycin, and 10% fetal bovine serum (FBS; Tianhang
Biotechnology Co., Hangzhou, China) in a humidified atmosphere with
5% CO2 at 37°C.
Cell transfection and detection of
transfection rate
To investigate the possible effects of piRNAs on
tumorigenesis, an siRNA inhibitor of piR651 was transfected into
NSCLC cells. The piR651 inhibitor and negative control (NC) siRNA
were synthesized by Shanghai GenePharma Co. (Shanghai, China). The
sequence of the piR651 inhibitor was
5′-GACGCUUUCCAAGGCACGGGCCCCUCUCU-3′. The sequence of the NC was
5′-CAGUACUUUUGUGUAGUACAA-3′. The inhibitor and NC were transfected
into A549 and HCC827 cells using Lipofectamine 2000 transfection
reagent (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Finally, fluorescence microscopy and flow
cytometry were utilized to determine the transfection rate. Cells
were seeded into 6-well plates 1 day before transfection. When
cells reached between 50 and 60% confluence, 100 pmol piR651
inhibitor with fluorescent marker [6-carboxyfluorescein (FAM)], or
100 pmol NC with FAM was transfected using Lipofectamine 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). The
transfection rate was determined using a fluorescence microscope
(magnification, ×200), by observing the number of cells, and a flow
cytometer (BD LSRII; BD Biosciences, Franklin Lakes, NJ, USA), by
detecting the degree of fluorescence in the piRNA inhibitor and the
NC.
MTT assay
MTT assays were utilized to ascertain the cell
growth rate following transfection with the piR651 inhibitor. Five
replicate wells were seeded for each group in this assay. A549 and
HCC827 cells were cultured in 96-well plates for 24 h at 37°C, and
transfected with either the piR651 inhibitor or the NC. After 12,
24, 48 and 72 h, 20 µl MTT (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) solution was added to each well. Cells were incubated at
37°C for 4 h in the dark and subsequently the supernatant was
discarded and 150 µl dimethyl sulfoxide (Thermo Fisher Scientific,
Inc.) was added to dissolve the crystallized formazan product. The
plate was agitated mildly for 10 min. The optical density (OD)
value was detected for each well at 490 nm using a Wellscan reader
(Labsystems, Santa Fe, NM, USA). Growth proliferation was
calculated as a percentage using the following formula: (OD
experiment/OD control) ×100. Independent MTT experiments were
performed three times.
Apoptosis analysis
To evaluate the association between piR651 and
apoptosis, HCC827 and A549 cells were collected and washed with PBS
24 h after transfection. Subsequently, 106 cells/ml were
stained using an Annexin V-fluorescein isothiocyanate apoptosis
detection kit (cat. no. CX1001-2; Beijing Biosea Biotechnology Co.,
Ltd., Beijing, China), according to the manufacturer's protocol.
The cells were subsequently suspended in binding buffer (Beijing
Biosea Biotechnology Co., Ltd.) and analyzed using a flow cytometer
(BD FACSCalibur; BD Biosciences). All tests were repeated three
times.
Western blot analysis
Western blot analysis was performed to detect
changes in apoptosis-associated proteins following transfection of
NSCLC cells with the piR651 inhibitor. In brief, HCC827 and A549
cells transfected with the piR651 inhibitor or the NC were lysed
with radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Haimen, China) after 46 h. The lysates were
separated by centrifugation at 12,000 × g for 15 min at 4°C. The
supernatants were boiled (10 min) and quantified using the
bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology), according to the manufacturer's protocol.
Subsequently, protein was separated using SDS-PAGE (12 and 6% gels;
Beyotime Institute of Biotechnology) and transferred onto
polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA,
USA). The membrane was subsequently blocked in Tris-buffered
saline-Tween-20 (TBST), containing 5% skimmed milk powder (TBSTM),
for 1 h at room temperature with gentle agitation. Membranes were
incubated at 4°C overnight with the following primary antibodies
against: Antibodies B-cell lymphoma 2 (Bcl-2; cat. no. ZS-492;
dilution, 1:100), Bcl-2-associated X protein (Bax; cat. no. ZS-526;
dilution, 1:100), β-actin (cat. no. TA-09; dilution, 1:1,000; all
from OriGene Technologies, Beijing, China), caspase-3-p17 (cat. no.
25546-1-AP; dilution, 1:200; ProteinTech Group, Inc., Wuhan, China)
and poly(ADP-ribose) polymerase 1 (PARP1; cat. no. 13371-1-AP;
dilution, 1:200; ProteinTech Group, Inc.). Subsequently, membranes
were washed with TBST (four times for 15 min each) and incubated at
room temperature with horseradish peroxidase (HRP) conjugated
affinipure goat anti-mouse immunoglobulin G (IgG) (cat. no.
ZB-2305; dilution, 1:5,000; OriGene Technologies, Inc., Beijing,
China) or HRP conjugated affinipure goat anti-rabbit IgG (cat. no.
ZB2301; dilution, 1:5,000; OriGene Technologies, Inc.) secondary
antibodies for 1 h. Following incubation, members were washed with
TBST three times for 15 min each. Subsequently, membranes were
incubated in Pierce ECL Western Blotting Substrate (cat. no. 32106;
Thermo Fisher Scientific, Inc.) for 5 min at room temperature.
Visualization was achieved using the X-OMAT BT Film (cat. no.
03690336810396; Carestream Health, Inc., Rochester, NY, USA) and
the Medical Intensifying Screen (cat. no. 2013004; Jianmin Medical
Instrument Co., Nanchang, China). All experiments were performed
three times.
Transwell migration and invasion
assays
Cell migration and invasion assays were performed
using Transwell chambers with 8-mm membrane pores (Corning
Incorporated, Corning, NY, USA). For the migration assay,
7–8×104 transfected cells per well were seeded in
serum-free medium in the upper chamber with a non-coated membrane.
In the invasion assays, 7–8×104 cells per well were
seeded in serum-free medium in the upper chamber with a membrane
that was pre-coated with Matrigel (BD Biosciences). For each assay,
the contents of the lower chambers were the same as those of the
upper chamber. After incubation for 48 h, the membranes were
removed from the plate, and the non-migrated cells were removed
from the upper surface with a cotton swab. The cells on the
underside of the membrane were fixed in 100% methanol and stained
with 0.5% crystal violet (diluted in 100% methanol). The cells on
the lower surface were counted under an inverted light microscope
and images were captured. Cells were counted in six randomly
selected fields and the average number was recorded. The relative
migration/invasion (%) was calculated as follows: (Average
number/100) ×100. Three independent experiments were performed.
Wound-healing assays
For the wound-healing assays, cells were grown in
6-well plates to near confluence. A linear scratch was carefully
made with a 10-µl pipette tip, and any cell debris was gently
washed away with phosphate-buffered saline (PBS). The HCC827 or
A549 cells were then incubated with serum-free RPMI-1,640 or DMEM,
respectively. Images were captured of the cells 48 h after the
scratch was created using an inverted light microscope (Olympus
Corporation, Tokyo, Japan). The distance between the cells which
migrated the furthest on each side to calculate the relative wound
closure (%).
Statistical analysis
Data were presented as the mean ± standard error of
the mean, of three independent experiments. MTT data was processed
using two-factor analysis of variance followed by Bonferroni's
correction and the remaining tests were analyzed by a two-sample
t-test using SPSS software (version 17.0; SPSS, Inc., Chicago, IL,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
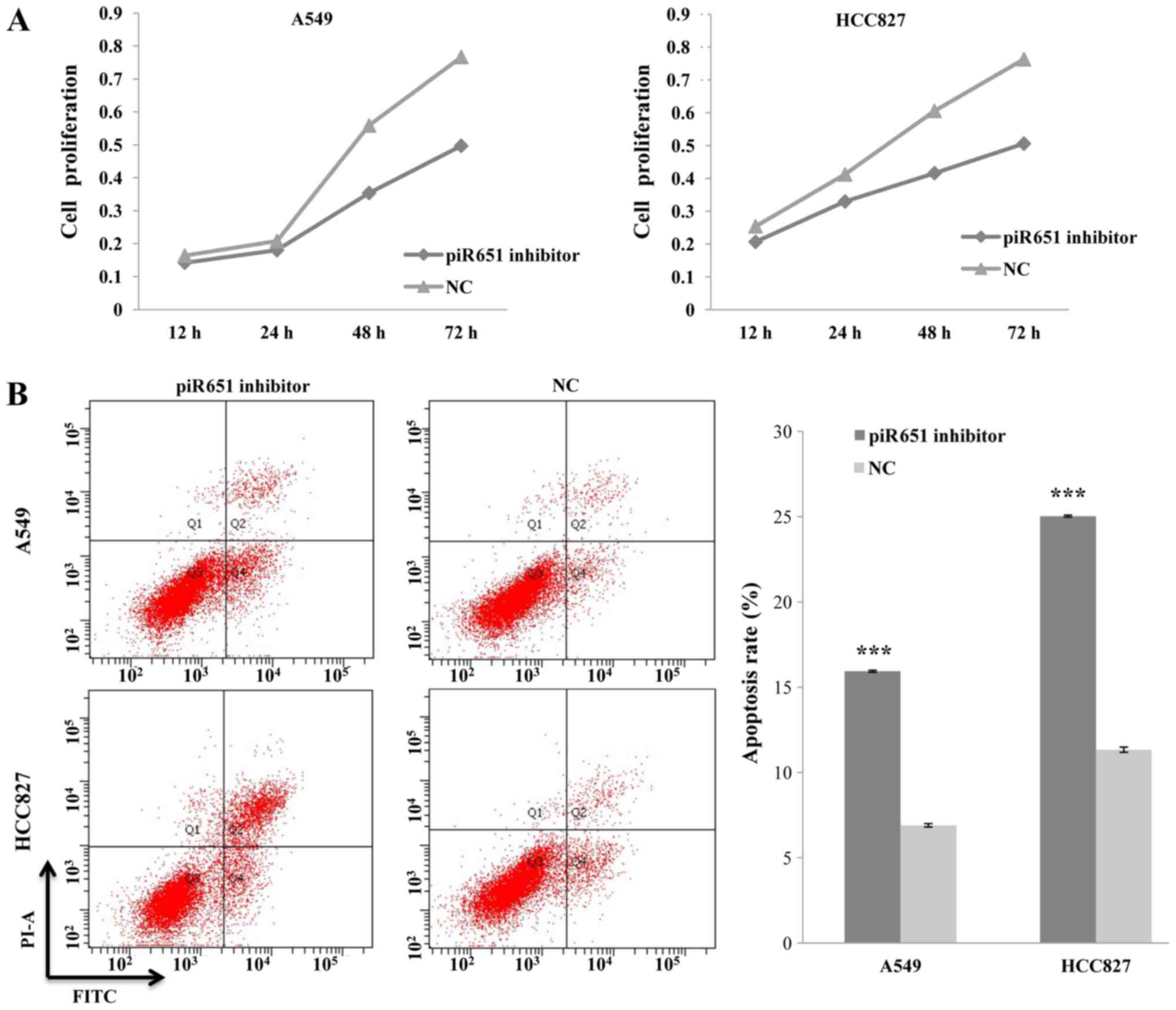
piR651 promotes cell proliferation in
a time-dependent manner and inhibits apoptosis in NSCLC cells
A previous study found piR651 to be overexpressed in
several cancer cell lines, including those of lung cancer (21). Thus, the present study reduced the
endogenous expression of piR651 by transfecting the NSCLC A549 and
HCC827 cell lines with a piR651 inhibitor. The rate of cell
proliferation decreased, compared with the NC, in response to
transfection with the piR651 inhibitor in the two cell lines. In
addition, the piR651 inhibitor regulated the cell proliferation
rate in a time-dependent manner (P<0.001; Fig. 1A).
A previous study indicated that apoptosis serves an
important role in regulating carcinogenesis (27). Therefore, we hypothesized that piR651
regulates tumorigenesis by altering the rate of NSCLC cell
apoptosis. An Annexin V-PI kit was used to detect apoptosis in A549
and HCC827 cells following transfection with the piR651 inhibitor
or the NC. The apoptotic rates of the piR651 inhibitor-transfected
groups were higher than those of the respective the NC-transfected
groups (t=145.31, P=0.001; t=135.5, P=0.001; Fig. 1B). On the basis of these results, it
was concluded that piR651 may affect tumorigenesis by promoting
cell proliferation and inhibiting apoptosis in NSCLC cells.
piR651 regulates apoptosis by altering
the expression of apoptosis-associated proteins in NSCLC cells
The previous experiment demonstrated that piR651
inhibited NSCLC cell apoptosis. Numerous studies have confirmed
that several pathways can regulate tumor cell apoptosis, including
the caspase pathway (28–31). Thus, classical apoptosis-associated
proteins, including caspase-3, Bax, Bcl-2 and PARP1, were
investigated. Using western blotting assays, it was confirmed that,
compared with the NC in A549 and HCC827 cells, caspase-3-p17
(P<0.001 and P=0.022), Bax (P=0.004 and P=0.014) and
cleaved-PARP-1 (P=0.014 and P<0.001) expression levels were
increased, whereas Bcl-2 (P=0.003 and P<0.001) expression was
decreased by the piR651, respectively (Fig. 2). Caspase-3 and Bax/Bcl-2 are proteins
involved in the mitochondrial apoptosis pathway (30–32). In
conclusion, the piR651 inhibitor induced apoptosis in A549 and
HCC827 cells, most likely through a mitochondria-dependent
pathway.
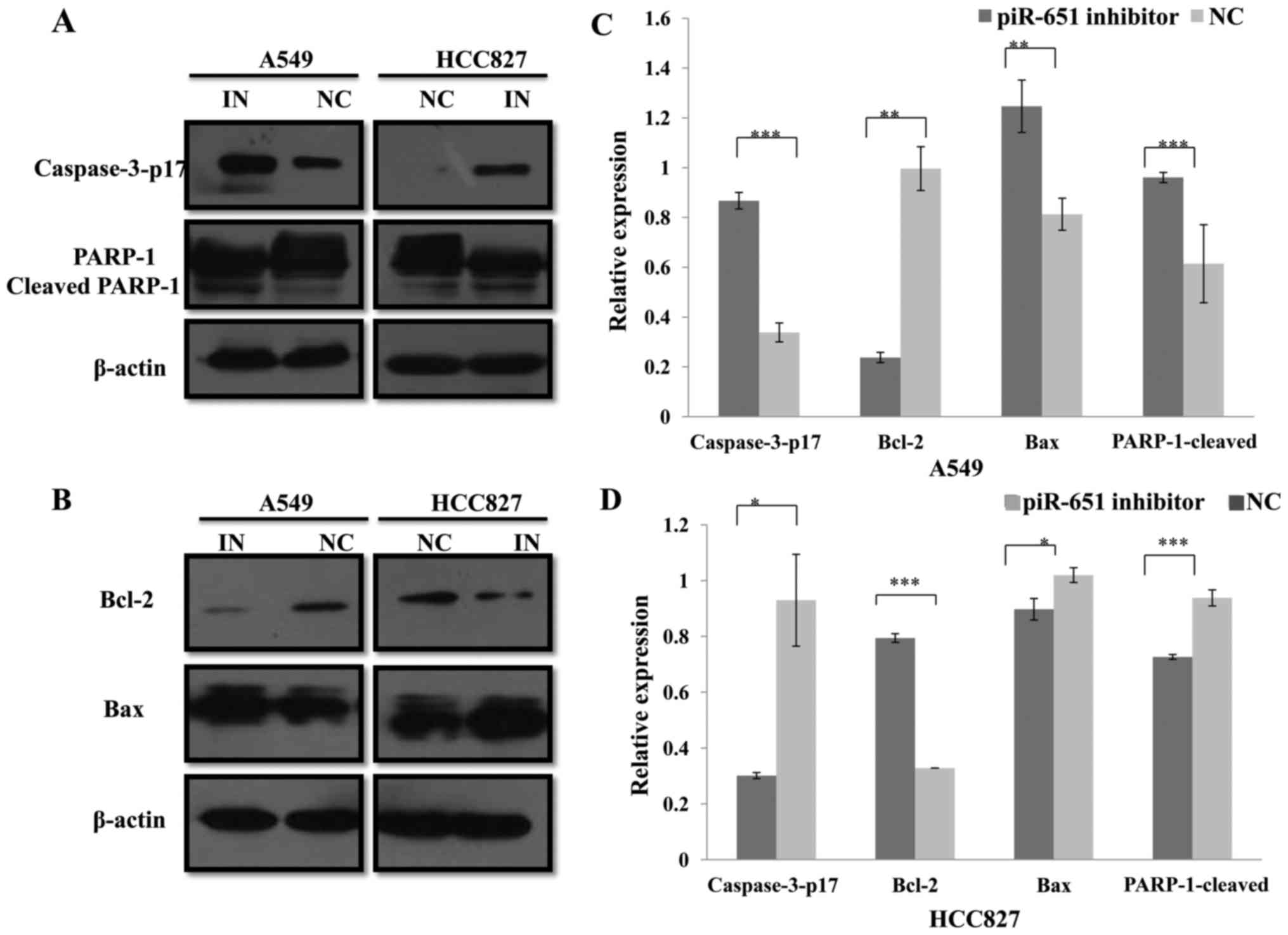 | Figure 2.piR651 regulates the expression of
apoptosis-associated proteins in A549 and HCC827 cells. (A)
Caspase-3-p17, PARP-1 and cleaved PARP1 protein levels in A549 and
HCC827 cells were analyzed using western blot. β-actin was used as
an internal control. (B) Bax and Bcl-2 protein levels in A549 and
HCC827 cells were analyzed using western blot. β-actin was used as
an internal control. (C) Compared with the NC, the expression of
caspase-3-p17 (***P<0.001), Bax (**P=0.004) and cleaved PARP1
(*P=0.014) increased and the expression of Bcl-2 (**P=0.003;)
decreased in response to the piR651 inhibitor in A549 cells. (D) In
HCC827 cells, the expression of caspase-3-p17 (*P=0.022), Bax
(*P=0.014) and cleaved PARP1 (***P<0.001) decreased and the
expression of Bcl-2 (***P<0.001) increased in response to the
piR651 inhibitor compared with the NC. piR651, piwi-interacting
RNA-651; Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X protein;
PARP, poly(ADP-ribose) polymerase; IN, piR651 inhibitor; NC,
negative control. |
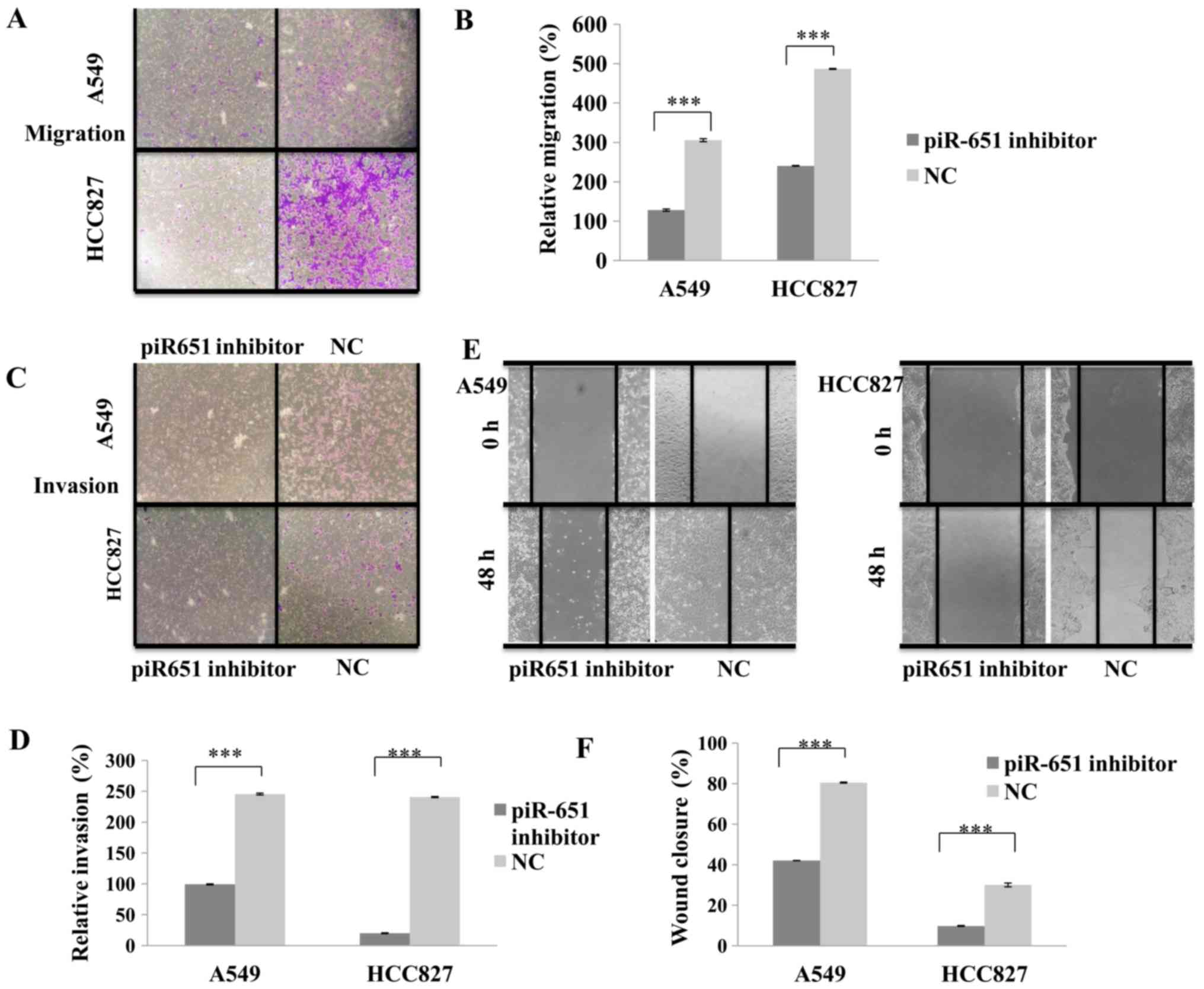
piR651 promotes NSCLC cell migration
and invasion in vitro
To determine whether the knockdown of endogenous
piR651 affects NSCLC cell migration and invasion, the piR651
inhibitor was transfected into NSCLC A549 and HCC827 cell lines to
downregulate the endogenous expression of piR651. As presented in
Fig. 3A and B, compared with the NC,
cells transfected with the piR651 inhibitor exhibited significantly
decreased cell migration (P<0.001). The results of the
wound-healing assay were similar to those of the migration assay;
transfection with the piR651 inhibitor significantly decreased the
wound-closure rate compared with the NC (P<0.001; Fig. 3C and D). The results of the cell
invasion assay were also similar to those of the migration assay,
with significantly fewer cells invading through the membrane in the
piR651 inhibitor-transfected group compared with the NC group in
the two cells lines (P<0.001; Fig. 3E
and F). These data support the hypothesis that a reduction in
endogenous piR651 expression may suppress NSCLC cell migration and
invasion.
Discussion
Accumulating evidence suggests the presence of
aberrant piRNA expression in numerous types of human cancer cells
and tissues, including gastric carcinoma, hepatic carcinoma,
bladder cancer, ovarian carcinoma and NSCLC, in which piRNAs may
act as oncogenes or tumor suppressors (18–24,26).
piR651, piR20365, piR4987, piR20485 and piR20582 are all
overexpressed in breast cancer tissues and cell lines (22,23),
whereas expression of piR823 is downregulated in gastric cancer
tissues and cell lines (20).
Although there are a growing number of studies concerning the
association between piRNAs and carcinogenesis (18–25), the
mechanism by which piRNAs regulate carcinogenesis remains
inadequately understood.
In a previous study, the expression of piR651 in
human gastric cancer tissues was found to be significantly
increased compared with paired normal tissues and was associated
with the TNM stage (20).
Additionally, the expression levels of piR651 were higher in
breast, colon, lung and gastric cancer tissues than in paired
normal tissues (21). The
upregulation of piR651 was also observed in several cancer cell
lines, including those of NSCLC (20,21).
However, the role of piR651 in carcinogenesis remains undefined.
Therefore, in the present study, the potential roles of piR651 in
NSCLC cells were investigated. The results of the present study
revealed that that the knockdown of endogenous piR651 in NSCLC
cells inhibited cell proliferation, induced apoptosis and inhibited
cell migration and invasion. The proliferation rate of A549 and
HCC827 cells decreased in a time-dependent manner following
transfection with the piR651 inhibitor compared with the same cells
transfected with the NC, whereas the apoptotic rate of these cells
was significantly higher. These data suggest that piR651 may serve
an oncogenic role in tumorigenesis.
Previous studies have demonstrated that mitochondria
play an important role in the intrinsic apoptosis pathway, which is
regulated by Bcl-2 family proteins through the pro-apoptotic
protein Bax and the anti-apoptotic protein Bcl-2 (30,31).
Analyses of mitochondria have confirmed their involvement in the
production of reactive oxygen species (ROS), the downregulation of
Bcl-2 family proteins, the upregulation of Bax, the release of
cytochrome c, and the activation of caspase-3 in various
cell lines (33–35). Therefore, we hypothesized that the
knockdown of endogenous piR651 would increase the rate of apoptosis
as well as the expression of cleaved caspase-3 and Bax, and would
downregulate the expression of Bcl-2. Western blot analysis
confirmed this hypothesis (Fig.
2).
It is estimated that ~90% of patients diagnosed with
NSCLC succumb due to metastasis, rather than from the primary
tumor. Owing to the high rates of recurrence and metastasis, NSCLC
is associated with a strong tendency to metastasize to specific
organs (32). Metastasis, which is a
multi-step process involving the proliferation of cancer cells in
the primary tumor, local invasion, invasion of lymphatic vessels
and blood vessels, local engraftment and the formation of a new
tumor (of the same tumor type) at a distal site (36,37), has
been estimated to be associated with >70% of all
cancer-associated mortalities (37).
Therefore, the identification of molecular factors that predict
tumor metastasis is clinically useful, and the identities of these
factors may suggest effective therapeutic strategies against NSCLC.
In the present study, the relative migration distance following
transfection with a piR651 inhibitor was revealed to be less than
that following transfection with the NC in wound-healing assays.
Furthermore, the numbers of invasive or migratory cells on the
underside of the Transwell membrane in the invasion and migration
assays were lower in the piR651 inhibitor-transfected group
compared with the NC group. These experiments thus confirmed that
piR651 promotes cell migration and invasion.
In conclusion, the results of the present study
results suggest the existence of a novel regulatory mechanism that
drives NSCLC cell proliferation, apoptosis, invasion, and
metastasis via the upregulation or downregulation of piR651
expression. The results of the current study confirm that the
downregulation of piR651 inhibits proliferation, induces apoptosis
and reduces invasion and migration in NSCLC. Therefore, piR651 may
provide an opportunity to improve the clinical diagnosis and
treatment of NSCLC.
Acknowledgements
The authors would like to thank Dr Pan
(Hepatosplenic Surgery Center, The First Affiliated Hospital of
Harbin Medical University, Harbin, Heilongjiang, China) for his
advice with manuscript preparation.
References
|
1
|
Dela Cruz CS, Tanoue LT and Matthay RA:
Lung cancer: Epidemiology, etiology and prevention. Clin Chest Med.
32:605–644. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Govindan R, Page N, Morgensztern D, Read
W, Tierney R, Vlahiotis A, Spitznagel EL and Piccirillo J: Changing
epidemiology of small-cell lung cancer in the United States over
the last 30 years: Analysis of the surveillance, epidemiologic, and
end results database. J Clin Oncol. 24:4539–4544. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Meister G: Argonaute proteins: Functional
insights and emerging roles. Nat Rev Genet. 14:447–459. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Peters L and Meister G: Argonaute
proteins: Mediators of RNA silencing. Mol Cell. 26:611–623. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hutvagner G and Simard MJ: Argonaute
proteins: Key players in RNA silencing. Nat Rev Mol Cell Biol.
9:22–32. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun Y, Ai X, Shen S and Lu S:
NF-κB-mediated miR-124 suppresses metastasis of non-small-cell lung
cancer by targeting MYO10. Oncotarget. 6:8244–8254. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu SL, Chen HY, Chang GC, Chen CY, Chen
HW, Singh S, Cheng CL, Yu CJ, Lee YC, Chen HS, et al: MicroRNA
signature predicts survival and relapse in lung cancer. Cancer
Cell. 13:48–57. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lin P, Yu S and Yang P: MicroRNA in lung
cancer. Br J Cancer. 103:1144–1148. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kourtidis A and Anastasiadis PZ: Bringing
together cell-to-cell adhesion and miRNA biology in cancer
research. Future Oncol. 12:1211–1214. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ling H, Krassnig L, Bullock MD and Pichler
M: MicroRNAs in testicular cancer diagnosis and prognosis. Urol
Clin North Am. 43:127–134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Aravin A, Gaidatzis D, Pfeffer S,
Lagos-Quintana M, Landgraf P, Iovino N, Morris P, Brownstein MJ,
Kuramochi-Miyagawa S, Nakano T, et al: A novel class of small RNAs
bind to MILI protein in mouse testes. Nature. 442:203–207.
2006.PubMed/NCBI
|
|
12
|
Girard A, Sachidanandam R, Hannon GJ and
Carmell MA: A germline-specific class of small RNAs binds mammalian
Piwi proteins. Nature. 442:199–202. 2006.PubMed/NCBI
|
|
13
|
Kwon C, Tak H, Rho M, Chang HR, Kim YH,
Kim KT, Balch C, Lee EK and Nam S: Detection of PIWI and piRNAs in
the mitochondria of mammalian cancer cells. Biochem Biophys Res
Commun. 446:218–223. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Siomi MC, Sato K, Pezic D and Aravin AA:
PIWI-interacting small RNAs: The vanguard of genome defence. Nat
Rev Mol Cell Biol. 12:246–258. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Klattenhoff C and Theurkauf W: Biogenesis
and germline functions of piRNAs. Development. 135:3–9. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suzuki R, Honda S and Kirino Y: PIWI
expression and function in cancer. Front Genet. 3:2042012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Batista PJ, Ruby JG, Claycomb JM, Chiang
R, Fahlgren N, Kasschau KD, Chaves DA, Gu W, Vasale JJ, Duan S, et
al: PRG-1 and 21U-RNAs interact to form the piRNA complex required
for fertility in C. Elegans. Mol Cell. 31:67–78. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu Y, Li C, Zhang K, Sun H, Tao D, Liu Y,
Zhang S and Ma Y: Identification of piRNAs in Hela cells by massive
parallel sequencing. BMB Rep. 43:635–641. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chu H, Hui G, Yuan L, Shi D, Wang Y, Du M,
Zhong D, Ma L, Tong N, Qin C, et al: Identification of novel piRNAs
in bladder cancer. Cancer Lett. 356:561–567. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cheng J, Guo JM, Xiao BX, Miao Y, Jiang Z,
Zhou H and Li QN: piRNA, the new non-coding RNA, is aberrantly
expressed in human cancer cells. Clin Chim Acta. 412:1621–1625.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheng J, Deng H, Xiao B, Zhou H, Zhou F,
Shen Z and Guo J: PiR-823, a novel non-coding small RNA,
demonstrates in vitro and in vivo tumor suppressive activity in
human gastric cancer cells. Cancer Lett. 315:12–17. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang G, Hu H, Xue X, Shen S, Gao E, Guo
G, Shen X and Zhang X: Altered expression of piRNAs and their
relation with clinicopathologic features of breast cancer. Clin
Transl Oncol. 15:563–568. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang H, Ren Y, Xu H, Pang D, Duan C and
Liu C: The expression of stem cell protein Piwil2 and piR-932 in
breast cancer. Surg Oncol. 22:217–223. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Law PT, Qin H, Ching AK, Lai KP, Co NN, He
M, Lung RW, Chan AW, Chan TF and Wong N: Deep sequencing of small
RNA transcriptome reveals novel non-coding RNAs in hepatocellular
carcinoma. J Hepatol. 58:1165–1173. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cui L, Lou Y, Zhang X, Zhou H, Deng H,
Song H, Yu X, Xiao B, Wang W and Guo J: Detection of circulating
tumor cells in peripheral blood from patients with gastric cancer
using piRNAs as markers. Clin Biochem. 44:1050–1057. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
UyBico SJ, Wu CC, Suh RD, Le NH, Brown K
and Krishnam MS: Lung Cancer Staging Es-sentials: The new TNM
staging system and potential imaging pitfalls. Radiographics.
30:1163–1181. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shin MH, He Y, Marrogi E, Piperdi S, Ren
L, Khanna C, Gorlick R, Liu C and Huang J: A RUNX2-mediated
epigenetic regulation of the survival of p53 defective cancer
cells. PLoS Genet. 12:e10058842016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gherman C, Braicu OL, Zanoaga O, Jurj A,
Pileczki V, Maralani M, Drigla F, Braicu C, Budisan L,
Achimas-Cadariu P and Berindan-Neagoe I: Caffeic acid phenethyl
ester activates pro-apoptotic and epithelial-mesenchymal
transition-related genes in ovarian cancer cells A2780 and
A2780cis. Mol Cell Biochem. 413:189–198. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu Z, Chen J, Shao L, Ma W and Xu D:
Erratum to: Promyelocytic leukemia protein enhances apoptosis of
gastric cancer cells through Yes-associated protein. Tumour Biol.
37:27752016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Malki A, Mohsen M, Aziz H, Rizk O, Shaban
O, El-Sayed M, Sherif ZA and Ashour H: New 3-Cyano-2-substituted
pyridines induce apoptosis in MCF 7 breast cancer cells. Molecules.
21:E2302016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stamelos VA, Fisher N, Bamrah H, Voisey C,
Price JC, Farrell WE, Redman CW and Richardson A: The BH3 mimetic
obatoclax accumulates in lysosomes and causes their alkalinization.
PLoS One. 11:e01506962016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chang JS, Su CY, Yu WH, Lee WJ, Liu YP,
Lai TC, Jan YH, Yang YF, Shen CN, Shew JY, et al: GIT1 promotes
lung cancer cell metastasis through modulating Rac1/Cdc42 activity
and is associated with poor prognosis. Oncotarget. 6:36278–36291.
2015.PubMed/NCBI
|
|
33
|
Cryns V and Yuan J: Proteases to die for.
Genes Dev. 12:1551–1570. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Amar SK, Goyal S, Mujtaba SF, Dwivedi A,
Kushwaha HN, Verma A, Chopra D, Chaturvedi RK and Ray RS: Role of
type I and type II reactions in DNA damage and activation of
caspase3 via mitochondrial pathway induced by photosensitized
benzophenone. Toxicol Lett. 235:84–95. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hamacher-Brady A, Stein HA, Turschner S,
Toegel I, Mora R, Jennewein N, Efferth T, Eils R and Brady NR:
Artesunate activates mitochondrial apoptosis in breast cancer cells
via iron-catalyzed lysosomal reactive oxygen species production. J
Biol Chem. 286:6587–6601. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Germanov E, Berman JN and Guernsey DL:
Current and future approaches for the therapeutic targeting of
metastasis. Int J Mol Med. 18:1025–1036. 2006.PubMed/NCBI
|
|
37
|
You J, He X, Ding H and Zhang T: BRMS1
regulates apoptosis in non-small cell lung cancer cells. Cell
Biochem Biophys. 71:465–472. 2015. View Article : Google Scholar : PubMed/NCBI
|