Introduction
Colorectal cancer is one of the most common types of
cancer and is the third leading cause of malignancy-associated
mortality worldwide (1). Although
screening methodologies have decreased the incidence of patients
with advanced stage, treating late-stage colorectal cancer
continues to challenge physicians. Understanding the molecular
footprint of colorectal cancer is critically important for the
development of novel treatments for this disease.
MicroRNAs (miRNAs/miRs) are small non-coding
single-stranded RNAs that regulate gene expression by inhibiting
mRNA translation by inducing mRNA degradation (2,3). The
miRNAs target ~33% of all protein-coding genes, and each miRNA is
able to potentially repress hundreds of target genes (4,5). miRNA is
able to regulate multiple steps of cancer development and
progression by modulating the expression of target genes. Recently,
a large number of aberrantly expressed miRNAs have been identified
in colorectal cancer and have been associated with colorectal
cancer progression (6–9).
miRNA (miR)-544 has been identified to be involved
in the regulation of numerous cellular processes, including polar
over-dominance inheritance (10),
stem cell self-renewal (11) and
osteogenic differentiation (12).
Although a number of previous studies have been reported that
miR-544 serves an important role in various types of cancer,
including breast cancer (13,14), lung cancer (15), cervical cancer (16), gastric cancer (17,18),
osteosarcoma (19,20) and glioma (21), the role of miR-544 in colorectal
cancer remains unknown. Recently, the role of miRNAs in the
regulation of expression of cell cycle-associated genes has been
investigated (22). For example,
miR-196a promotes cervical cancer proliferation by regulating
forkhead box O1 (FOXO1) and p27 (23). miR-142-3p has been suggested to act as
a tumor suppressor that inhibits tumor proliferation by targeting
cell division cycle 25C (24).
However, there have been few detailed studies investigating the
role of miR-544 in cell cycle-associated cancer cell proliferation
(16,17).
The results of the present study identified miR-544
as a candidate oncogenic miRNA in colorectal cancer. The results of
the present study revealed that miR-544 is upregulated in
colorectal tissues. In vitro investigation demonstrated that
miR-544 promotes colorectal cancer cell progression by binding to
the 3′-untranslated region (UTR) of FOXO1 mRNA. Thus, miR-544 may
serve an important role in the development and progression of
colorectal cancer.
Materials and methods
Cell culture and specimens
HT29, SW480, LOVO and HCT116 colorectal cancer cell
lines were obtained from Shanghai Institute of Biochemistry and
Cell Biology, Chinese Academy of Sciences (Shanghai, China). Cells
were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (FBS; Invitrogen; Thermo Fisher Scientific,
Inc.). The cells were incubated at 37°C in a humidified atmosphere
containing 5% CO2.
A total of 20 fresh colorectal cancer tissues and
matched adjacent non-tumorous tissues were obtained from 9 males
and 11 females with ages ranging between 40 and 65 years from
January 2012 to December 2013. All tissues were stored at −80°C
immediately following removal until use. None of the patients had
received any treatment prior to surgery. The present study was
approved by the Institutional Review Board of the Tianjin Medical
University Cancer Institute and Hospital (Tianjin, China) and
written informed consent was obtained from all patients prior to
enrollment in the present study.
Plasmid, short interfering RNA and
transfection
To construct the FOXO1 expression vector, the entire
coding sequence of the FOXO1 was amplified using the polymerase
chain reaction (PCR) using Platinum SuperFi DNA (Thermo Fisher
Scientific, Inc.) in the HT29 cell line according to the
manufacturer's protocol using the following thermocycling
conditions: 95°C for 3 min, then 95°C for 30 sec, 60°C for 30 sec
and 72°C for 2 min for a total of 40 cycles. The primers were as
following: Forward, 5′-GCGGATCCATGGCCGAGGCGCCTCAGGT-3′ and reverse,
5′-GCCTCGAGTCAGCCTGACACCCAGCTAT-3′. The PCR product was cloned into
the pcDNA3 vector (Thermo Fisher Scientific, Inc.). The mutated
miR-544-binding site sequence on the FOXO1 3′-UTR was cloned into
the psiCHECK2 vector (Promega Corporation, Madison, WI, USA)
[UTR-FOXO1-1/2wild-type (wt) and UTR-FOXO1-1/2mutant (mu)]. The
construct was confirmed by sequencing. The miR-544 mimic, inhibitor
and the negative control were purchased from Guangzhou RiboBio Co.,
Ltd. (Guangzhou, China). miRNAs or plasmids were transfected with
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Western blot analysis
Cells were collected in lysis buffer (60 mM
Tris/HCl, 2% SDS and 10% glycerol) with 0.1% protease inhibitor
cocktail III (EMD Millipore, Billerica, CA, USA). The protein
concentration was determined using the Bradford assay using a com
mercial kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA). A
total of 50 µg protein was separated by SDS-PAGE (10% gel).
Subsequently, the proteins were transferred onto polyvinylidene
membranes (EMD Millipore). Following blocking with 5% skimmed milk
in TBST at room temperature for 1 h, the membranes were probed with
anti-FOXO1 (cat. no. sc-11350; dilution, 1:1,000; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), anti-cyclin D1 (cat. no.
sc-70899; dilution, 1:2,000; Santa Cruz Biotechnology, Inc.),
anti-phosphorylated retinoblastoma protein (pRb; cat. no. 8516;
dilution, 1:2,000; Cell Signaling Technology, Inc., Danvers, MA,
USA) and anti-p27 antibodies (cat. no. 3686; dilution, 1:2,000;
Cell Signaling Technology, Inc.) at 4°C overnight. The following
day, the membranes were incubated with horseradish
peroxidase-conjugated secondary antibody (cat. nos. 7076 and 7074;
dilution, 1:2,500) at 4°C for 1 h, and the bands were visualized
using enhanced chemiluminescence detection reagents (EMD
Millipore). GAPDH (cat. no. sc-365062; dilution, 1:3,000; Santa
Cruz Biotechnology, Inc.) served as a loading control.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
miRNA from tissue samples and cultured cells was
extracted using the mirVana miRNA Isolation kit (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. Total
RNA was extracted using TRIzol® (Thermo Scientific)
reagent, according to the manufacturer's protocol. A 5 µg amount of
total RNA was converted into first-strand cDNA using the TaqMan
miRNA reverse transcription kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) or SuperScript II Reverse Transcriptase kit
(Invitrogen; Thermo Fisher Scientific, Inc.). The expression level
of miR-544 was quantified using the miRNA-specific TaqMan miRNA
assay kit (Applied Biosystems; Thermo Fisher Scientific, Inc.),
which is specific for hsa-miR-544. The RT-qPCR analysis of FOXO1
expression levels were performed using the Fast SYBR-Green Master
Mix System (Invitrogen; Thermo Fisher Scientific, Inc.), according
to the manufacturer's protocol using the following thermocycling
conditions: 95°C for 3 min, then 95°C for 15 sec and 60°C for 1 min
for a total of 40 cycles. The following primers were used: GAPDH
forward, 5′-GAAGGTGAAGGTCGGAGTC-3 and reverse,
5′-GAAGATGGTGATGGGATTTC-3; and FOXO1 forward,
5′-AAGAGCGTGCCCTACTTCAA-3′ and reverse, 5′-CTGTTGTTGTCCATGGATGC-3′.
The targeted gene relative quantification was provided by the Cq
values and the relative mRNA or miRNA expression levels of targeted
genes were determined as 2−[(Cq of FOXO1 or miR-544)-(Cq of
GAPDH or U6)] (25). The
experiment was performed in triplicate.
Colony formation assay
A total of 1×103 cells were seeded in a
60-mm dish following transfection and cultured at 37°C in 5%
CO2. Following a 2-week incubation, the cells were
fixation with 10% formaldehyde for 30 min at room temperature and
stained with 1% crystal violet for another 30 min at room
temperature.
MTT assay
A total of 2×103 cells were seeded in
96-well plates per well after transfection. The cells were
incubated in 10 µl MTT (0.5 mg/ml; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) at the indicated time points for 4 h at 37°C.
The medium was then removed and 100 µl dimethyl sulfoxide was
added. The absorbance at 570 nm was detected using a microplate
auto-reader (Bio-Rad Laboratories, Inc.).
Invasion assay
The in vitro invasion assay was performed
using an 8 µm pore size 24-well Transwell chamber coated with 150
µg Matrigel (BD Biosciences, San Jose, CA, USA). Cells
(1×105) in serum-free DMEM were added to the inserts and
600 µl DMEM supplemented with 10% FBS was added to the lower wells
to act as the nutritional attractant. Following incubation for 20 h
at 37°C, the cells on the upper surface of the membrane were
removed and the migratory cells that had attached to the lower
surface were fixed with 100% methanol for 5 min and stained with 1%
crystal violet for 30 min at room temperature. Cells in 3 selected
microscopic fields (magnification, ×400) fields of view were
counted and expressed as the average number of cells per field of
view using a light microscope.
Bioinformatic prediction of miR-544
potential targets
The microRNA.org
targets and expression (www.microrna.org) database was used to predict
potential targets for miR-544, as previously described (26).
Flow cytometric analysis
Cells were collected by trypsinization and fixed in
70% ethanol in PBS at 4°C overnight. Cells were centrifuged at 500
× g for 1 min at room temperature, and then washed with cold PBS
and re-centrifuged as before. The cells were incubated with 20
mg/ml propidium iodide (Sigma-Aldrich; Merck KGaA) supplemented
with 2 mg/ml RNase A for 20 min at room temperature. The cells were
then analyzed using a flow cytometer (FACSCalibur II; BD
Biosciences). The data were processed using the ModFit LT 3.2
software (Verity Software House, Inc., Topsham, ME, USA).
Luciferase reporter assay
Cells (2×104) were seeded in triplicate
in 24-well plates. A total of 200 ng UTR-FOXO1-1/2wt or
UTR-FOXO1-1/2 mu was co-transfected with miR-544 mimic or control
into the cells using Lipofectamine 3000 (Thermo Fisher Scientific,
Inc.), according to the manufacturer's protocol. Luciferase and
Renilla luciferase activities were evaluated 48 h after
transfection using the Dual Luciferase Reporter Assay kit (Promega
Corporation), according to the manufacturer's protocol. The results
are expressed as a normalized ratio of Renilla to firefly
luciferase.
Statistical analysis
The results are representative of three independent
experiments and presented as the mean ± standard deviation.
Student's t-test was used to perform comparisons between two groups
of data. Multiple comparisons between data were performed using
one-way analysis of variance, followed by Dunnett's test. P<0.05
was considered to indicate a statistically significant difference.
Statistical analysis was performed using SPSS software (version
22.0; IBM Corp., Armonk, NY, USA).
Results
miR-544 is upregulated in colorectal
cancer cell lines
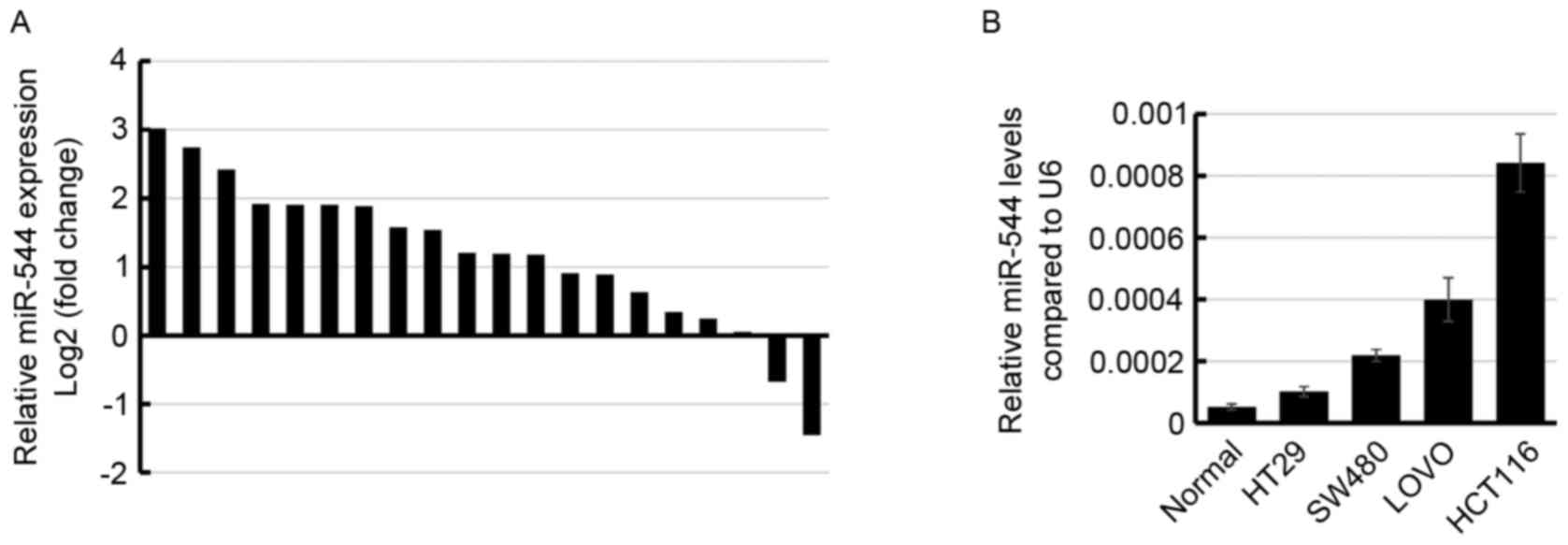
To examine the miR-544 expression level in
colorectal cancer, the miR-544 expression level was determined in
20 pairs of colorectal cancer tissues and their adjacent normal
tissues using RT-qPCR. The results demonstrated that miR-544 was
higher in 60% (12/20) colorectal cancer tissue specimens compared
with in normal tissues (Fig. 1A).
Subsequently, the miR-544 expression level was determined in 4
colorectal cancer cell lines. Consistent with the results observed
in the clinical tissue samples, miR-544 was significantly increased
in the 4 colorectal cancer cell lines compared with the normal
tissue samples (Fig. 1B), suggesting
that miR-544 may be upregulated in human colorectal cancer.
miR-544 promotes colorectal cancer
cell proliferation and invasion
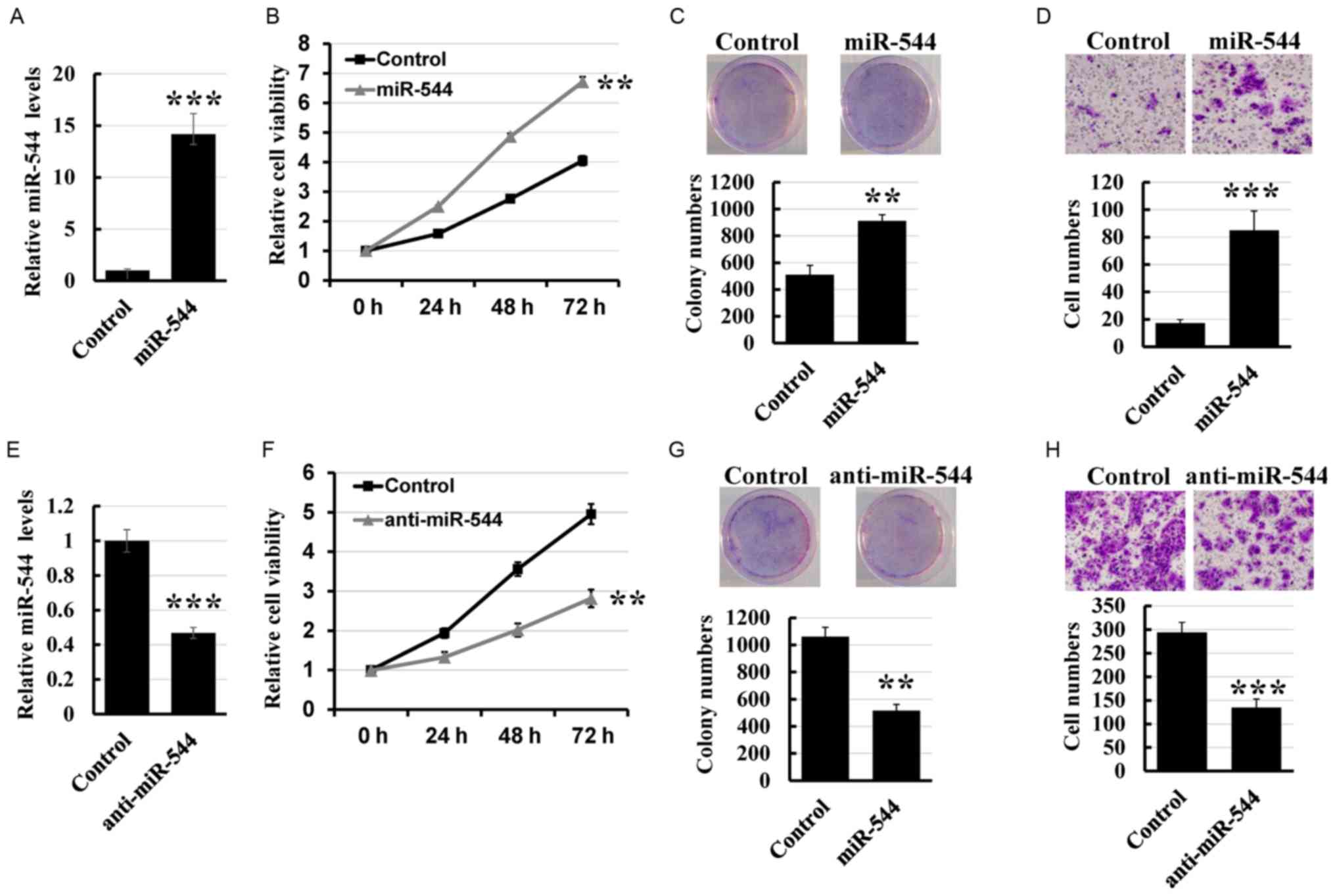
To determine the effect of miR-544 on colorectal
cancer cell proliferation and invasion, miR-544 mimics were
transfected into the HT29 cell line (Fig.
2A). MTT and colony formation assays revealed that
overexpression of miR-544 markedly increased the growth rate
compared with that of control cells (Fig.
2B and C). Furthermore, the Transwell assay demonstrated that
overexpression of miR-544 increased the invasion rate of HT29 cells
compared with the control cells (Fig.
2D). To further investigate the role of miR-544 on colorectal
cancer cell proliferation and invasion, the miR-544 inhibitor was
transfected into the HCT116 cell line (Fig. 2E). As presented in Fig. 2F-H, knockdown of miR-544 significantly
decreased the proliferation and invasion of HCT116 cells compared
with the control cells. Thus, these results indicated that miR-544
promoted colorectal cancer cell proliferation and invasion.
Effect of miR-544 on the cell
cycle
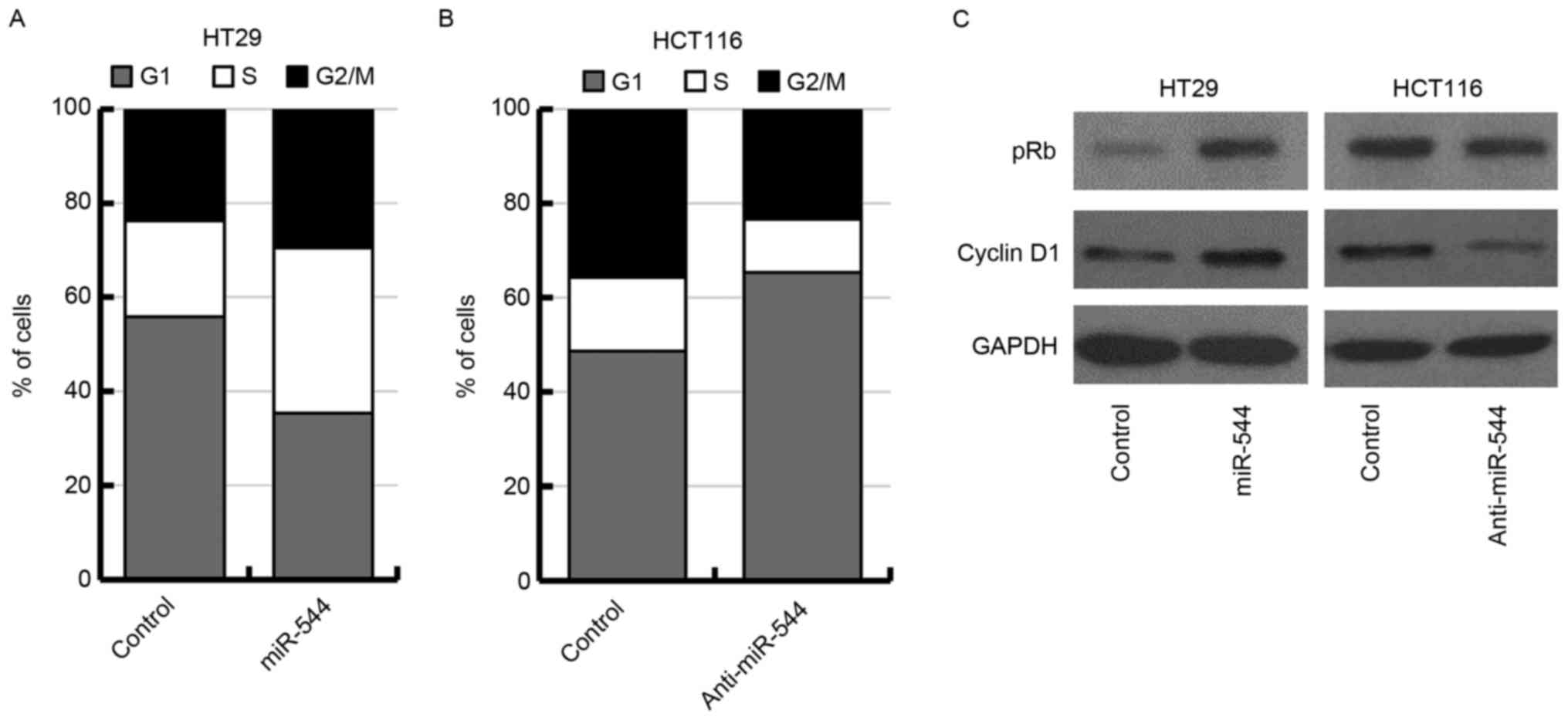
As miR-544 markedly affected cell proliferation in
HT29 and HCT116 cells, the results of the present study suggested
that miR-544 may function by regulating the cell cycle of
colorectal cancer cells. Thus, the effect of miR-544 on cell cycle
distribution was examined in miR-544-overexpressed HT29 cells,
miR-544-depleted HCT116 cells and their corresponding control cells
by flow cytometry. The results revealed that overexpression of
miR-544 increased the percentage of cells in the S phase in HT29
cells (Fig. 3A). Knockdown of miR-544
increased the percentage of cells in G0/G1
phase in HCT116 cells (Fig. 3B). In
the present study, it was investigated further whether the
expression of pRb or cyclin D1 may be regulated by miR-544. The
results indicated that miR-544-overexpressed HT29 cells exhibited a
significant upregulation of pRb and cyclin D1 as determined using
western blotting (Fig. 3C, left
panel). Conversely, the expression levels of pRb and cyclin D1 were
downregulated in miR-544-depleted HCT116 cells (Fig. 3C, right panel). Together, these
results indicated that miR-544 may promote G1 to S phase
transition in colorectal cancer cells.
FOXO1 is direct target of miR-544 in
colorectal cancer cells
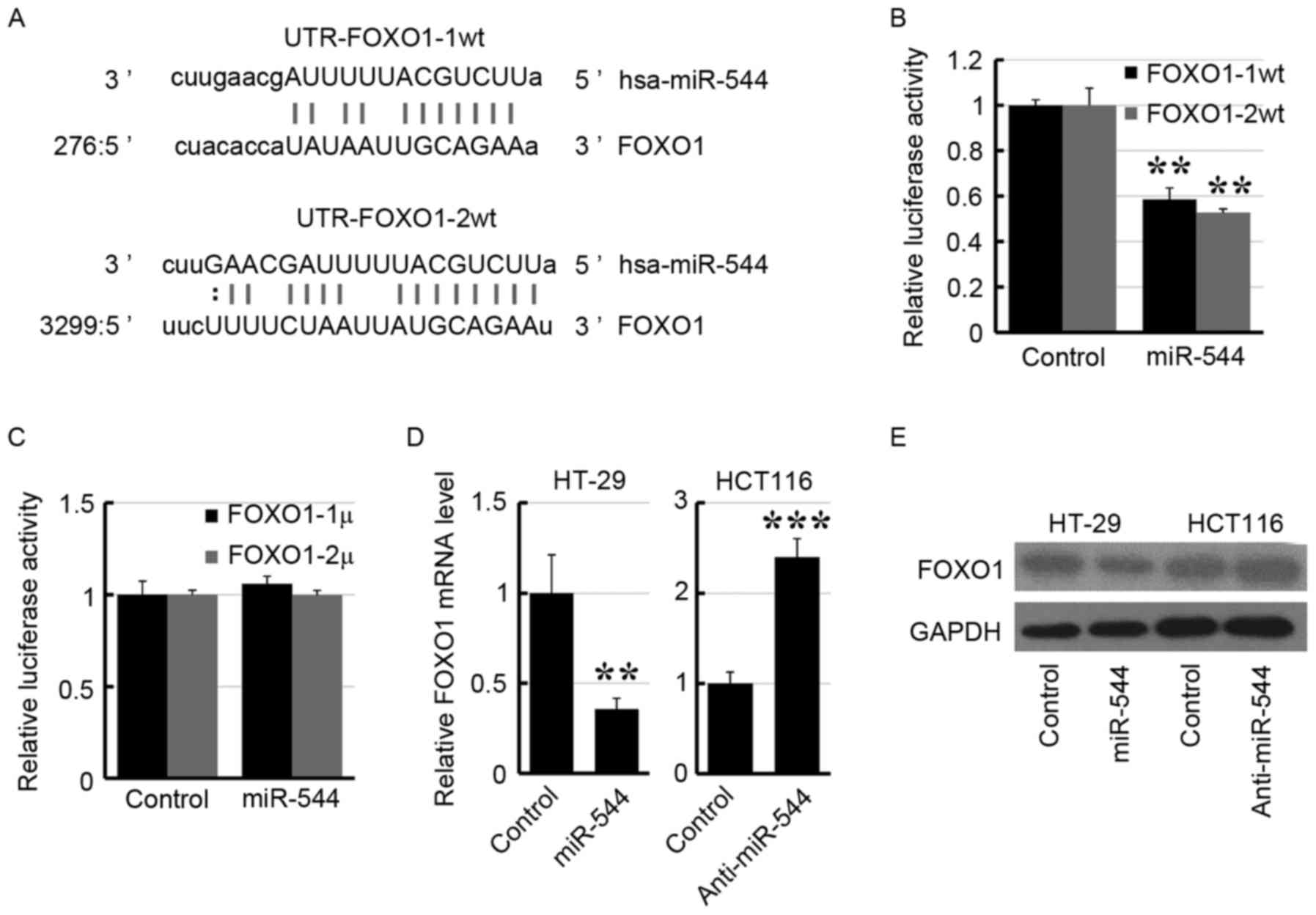
To identify the target gene of miR-544, a publically
available database (http://www.microrna.org) was interrogated and it was
revealed that FOXO1 was a potential target gene of miR-544
(Fig. 4A). Subsequently, the 3′-UTR
of FOXO1 was subcloned into the psiCHECK2 vector. As presented in
Fig. 4B, transfection of the miR-544
mimic suppressed the luciferase activity of UTR-FOXO1-1/2wt in
293FT cells. Furthermore, site mutations in the miR-544-binding
site region of the FOXO1 3′-UTR abrogated the repressive effect of
miR-544 (Fig. 4C). Furthermore,
overexpression of miR-544 decreased the expression level of FOXO1
in HT29 cells, whereas knockdown of miR-544 increased the FOXO1
expression level in HCT116 cells by RT-qPCR (Fig. 4D) and western blotting (Fig. 4E), respectively. These results
suggested that FOXO1 may be the target of miR-544.
FOXO1 mediates miR-544-induced
colorectal cancer progression
To further demonstrate whether miR-544 induced
colorectal cancer progression via FOXO1 repression, FOXO1 was
transfected into miR-544-overexpressed HT29 cells. As presented in
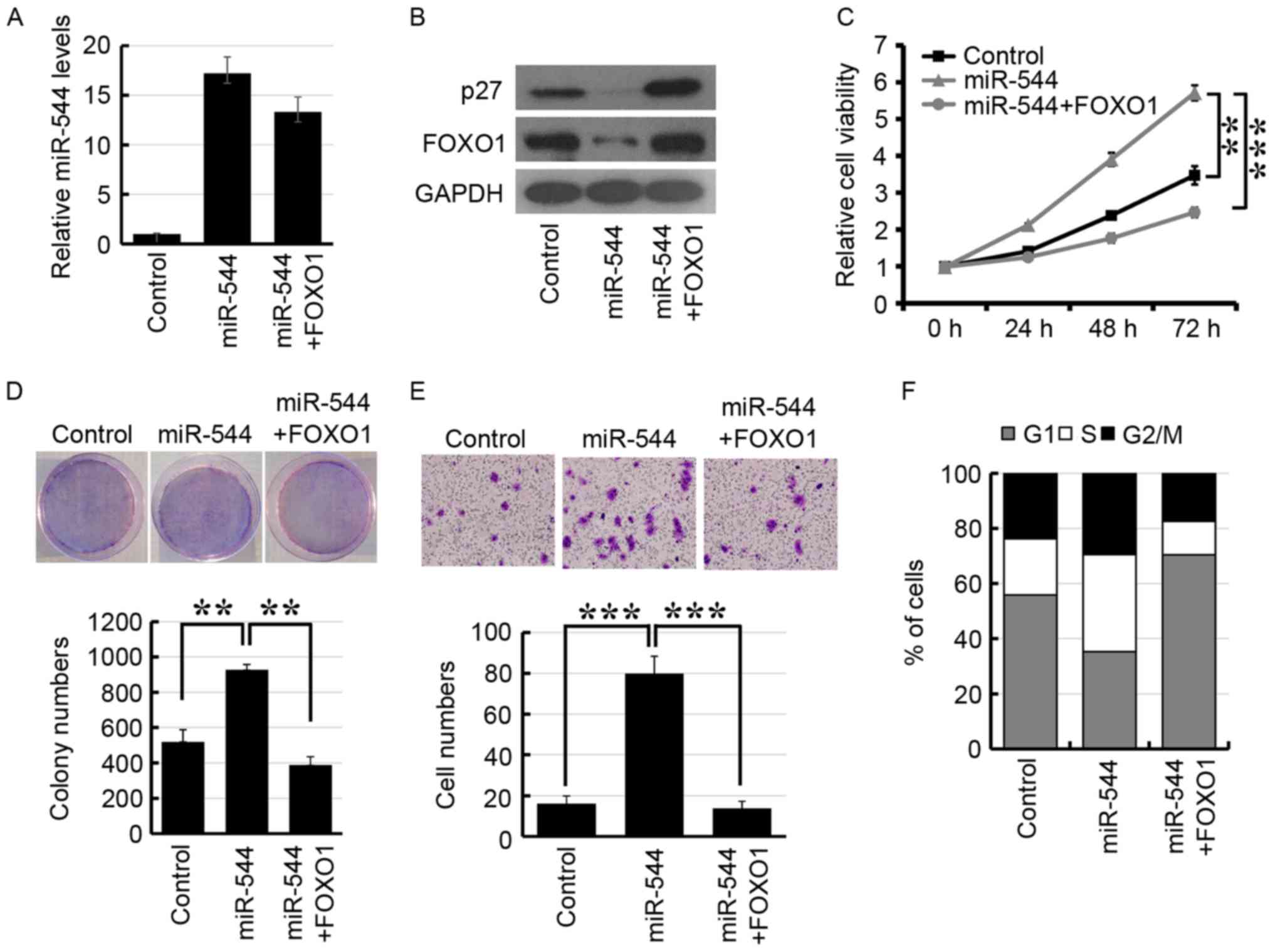
Fig. 5A, RT-qPCR analysis revealed
that the expression level of miR-544 was upregulated in miR-544
mimic-transfected HT29 cells. Furthermore, FOXO1 was significantly
downregulated in miR-544-overexpressed HT29 cells and the
expression of FOXO1 was re-established following transfection of
FOXO1 (Fig. 5B). In addition,
overexpression of miR-544 significantly decreased the expression of
p27, which is the downstream target gene of FOXO1 (Fig. 5B). The MTT, colony formation and
Transwell assays also indicated that co-transfection of miR-544 and
FOXO1 may re-establish miR-544-promoted cell proliferation and
invasion in HT29 (Fig. 5C-E). The
analysis of cell cycle distribution revealed that overexpression of
FOXO1 may re-establish the miR-544-induced increased proportion of
cells in the S phase (Fig. 5F).
Together, these results indicated that miR-544 may promote
colorectal cancer progression by the inhibition of FOXO1.
Discussion
miRNAs are post-transcriptional regulators of
oncogenes or tumor suppressors, and their deregulation is
associated with cancer development and progression (27). The results of the present study
indicated that miR-544 was upregulated in colorectal cancer cell
lines and tissues. In addition, overexpression of miR-544 promoted
G1 to S phase transition and cancer progression in
colorectal cancer cell lines, whereas knockdown of miR-544
diminished these effects. Furthermore, FOXO1 is a direct target of
miR-544. The results of the present study suggested that miR-544
may serve a critical role in colorectal cancer progression.
Aberrant expression of miR-544 has been observed in
numerous cancer subtypes, suggesting that miR-544 may serve an
important role in cancer development and progression (10,13–15,17–21).
A previous study demonstrated that miR-544 promoted cell
proliferation by downregulation of the tumor suppressor iroquois
homeobox 1 in gastric cancer (17).
As an oncogenic miRNA, miR-544 has also been revealed to be
involved in the progression of nasopharyngeal carcinoma (28). Previous studies have demonstrated that
miR-544 functions as a tumor suppressor and downregulates 14q32
miRNA (miR-382, miR-134 and miR-544), contributing to the
biological behavior of osteosarcoma (19,20). These
results indicated that dysregulation of miR-544 may occur in a
tissue-specific manner in the various types of cancer; however, the
role of miR-544 in colorectal cancer remains unknown. In the
present study, the expression of miR-544 was upregulated in
colorectal cancer tissues compared with the paired adjacent normal
tissues. Furthermore, miR-544 promoted colorectal cancer cell
proliferation and invasion. These results revealed that miR-544 may
act as an oncogenic miRNA in colorectal cancer development and
progression.
The forkhead box O (FOXO) family proteins, FOXO1,
FOXO3a, FOXO4 and FOXO6, are key effectors of the phosphoinositide
3-kinase/protein kinase B signaling pathway and participate in
numerous biological processes, including cell differentiation,
oxidative stress responses, carcinogenesis and cell cycle
regulation (29–32). Increased FOXO1 expression leads to
G1/S cell cycle arrest, resulting in upregulated
expression of the cyclin-dependent kinase inhibitors p21 and p27,
and downregulation of cyclin D1 (33–35). FOXO1
has been reported to serve a crucial role in cancer development and
progression. Repression of FOXO1 resulted in impaired cell death
control and a dysregulated cell cycle, which may have a central
role in tumorigenesis (23,36). Numerous miRNAs have been reported to
regulate FOXO1 in colorectal cancer development and progression
(37–39). To the best of our knowledge, the
present study is the first to have identified FOXO1 as a target of
miR-544, suggesting a crucial functional role of FOXO1 in
colorectal carcinogenesis.
In conclusion, the results of the present study
revealed that miR-544 promotes colorectal cancer development and
progression by suppression of FOXO1 expression. Understanding the
precise role that miR-544 serves in the induction of cancer cell
proliferation will increase our understanding of the biology of
colorectal cancer, and the inhibition of miR-544 may represent a
novel therapeutic strategy in the treatment of colorectal
cancer.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sakaguchi M, Hisamori S, Oshima N, Sato F,
Shimono Y and Sakai Y: miR-137 regulates the tumorigenicity of
colon cancer stem cells through the inhibition of DCLK1. Mol Cancer
Res. 14:354–362. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pathak S, Meng WJ, Nandy SK, Ping J,
Bisgin A, Helmfors L, Waldmann P and Sun XF: Radiation and SN38
treatments modulate the expression of microRNAs, cytokines and
chemokines in colon cancer cells in a p53-directed manner.
Oncotarget. 6:44758–44780. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sakai H, Sato A, Aihara Y, Ikarashi Y,
Midorikawa Y, Kracht M, Nakagama H and Okamoto K: MKK7 mediates
miR-493-dependent suppression of liver metastasis of colon cancer
cells. Cancer Sci. 105:425–430. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nagaraju GP, Madanraj AS, Aliya S, Rajitha
B, Alese OB, Kariali E, Alam A and El-Rayes BF: MicroRNAs as
biomarkers and prospective therapeutic targets in colon and
pancreatic cancers. Tumour Biol. 37:97–104. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gao YQ, Chen X, Wang P, Lu L, Zhao W, Chen
C, Chen CP, Tao T, Sun J, Zheng YY, et al: Regulation of DLK1 by
the maternally expressed miR-379/miR-544 cluster may underlie
callipyge polar overdominance inheritance. Proc Natl Acad Sci USA.
112:pp. 13627–13632. 2015; View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Song W, Mu H, Wu J, Liao M, Zhu H, Zheng
L, He X, Niu B, Zhai Y, Bai C, et al: miR-544 regulates dairy goat
male germline stem cell self-renewal via targeting PLZF. J Cell
Biochem. 116:2155–2165. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Song WQ, Gu WQ, Qian YB, Ma X, Mao YJ and
Liu WJ: Identification of long non-coding RNA involved in
osteogenic differentiation from mesenchymal stem cells using
RNA-Seq data. Genet Mol Res. 14:18268–18279. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Haga CL, Velagapudi SP, Strivelli JR, Yang
WY, Disney MD and Phinney DG: Small molecule inhibition of miR-544
biogenesis disrupts adaptive responses to hypoxia by modulating
ATM-mTOR signaling. ACS Chem Biol. 10:2267–2276. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu P, Gu Y, Li L, Wang F and Qiu X:
miR-544a promotes breast cancer cell migration and invasion
reducing cadherin 1 expression. Oncol Res. 23:165–170. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mo X, Zhang F, Liang H, Liu M, Li H and
Xia H: miR-544a promotes the invasion of lung cancer cells by
targeting cadherina 1 in vitro. Onco Targets Ther. 7:895–900. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mao L, Zhang Y, Deng X, Mo W, Yu Y and Lu
H: Transcription factor KLF4 regulates microRNA-544 that targets
YWHAZ in cervical cancer. Am J Cancer Res. 5:1939–1953.
2015.PubMed/NCBI
|
|
17
|
Zhi Q, Guo X, Guo L, Zhang R, Jiang J, Ji
J, Zhang J, Zhang J, Chen X, Cai Q, et al: Oncogenic miR-544 is an
important molecular target in gastric cancer. Anticancer Agents Med
Chem. 13:270–275. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yanaka Y, Muramatsu T, Uetake H, Kozaki K
and Inazawa J: miR-544a induces epithelial-mesenchymal transition
through the activation of WNT signaling pathway in gastric cancer.
Carcinogenesis. 36:1363–1371. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sarver AL, Thayanithy V, Scott MC,
Cleton-Jansen AM, Hogendoorn PC, Modiano JF and Subramanian S:
MicroRNAs at the human 14q32 locus have prognostic significance in
osteosarcoma. Orphanet J Rare Dis. 8:72013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Thayanithy V, Sarver AL, Kartha RV, Li L,
Angstadt AY, Breen M, Steer CJ, Modiano JF and Subramanian S:
Perturbation of 14q32 miRNAs-cMYC gene network in osteosarcoma.
Bone. 50:171–181. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ma R, Zhang G, Wang H, Lv H, Fang F and
Kang X: Downregulation of miR-544 in tissue, but not in serum, is a
novel biomarker of malignant transformation in glioma. Oncol Lett.
4:1321–1324. 2012.PubMed/NCBI
|
|
22
|
Bhattacharyya NP, Das E, Bucha S, Das S
and Choudhury A: Regulation of cell cycle associated genes by
microRNA and transcription factor. Microrna. 5:180–200. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hou T, Ou J, Zhao X, Huang X, Huang Y and
Zhang Y: MicroRNA-196a promotes cervical cancer proliferation
through the regulation of FOXO1 and p27Kip1. Br J Cancer.
110:1260–1268. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cao XC, Yu Y, Hou LK, Sun XH, Ge J, Zhang
B and Wang X: miR-142-3p inhibits cancer cell proliferation by
targeting CDC25C. Cell Prolif. 49:58–68. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Betel D, Wilson M, Gabow A, Marks DS and
Sander C: The microRNA.org resource: Targets and expression.
Nucleic Acids Res. 36:D149–D153. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Markopoulos GS, Roupakia E, Tokamani M,
Chavdoula E, Hatziapostolou M, Polytarchou C, Marcu KB,
Papavassiliou AG, Sandaltzopoulos R and Kolettas E: A step-by-step
microRNA guide to cancer development and metastasis. Cell Oncol
(Dordr). 40:303–339. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Luo Z, Zhang L, Li Z, Li X and Li G, Yu H,
Jiang C, Dai Y, Guo X, Xiang J and Li G: An in silico analysis of
dynamic changes in microRNA expression profiles in stepwise
development of nasopharyngeal carcinoma. BMC Med Genomics. 5:32012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shao H, Mohamed EM, Xu GG, Waters M, Jing
K, Ma Y, Zhang Y, Spiegel S, Idowu MO and Fang X: Carnitine
palmitoyltransferase 1A functions to repress FoxO transcription
factors to allow cell cycle progression in ovarian cancer.
Oncotarget. 7:3832–3846. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bicknell KA: Forkhead (FOX) transcription
factors and the cell cycle: Measurement of DNA binding by FoxO and
FoxM transcription factors. Methods Mol Biol. 296:247–262.
2005.PubMed/NCBI
|
|
31
|
Kuscu N and Celik-Ozenci C: FOXO1, FOXO3,
and FOXO4 are differently expressed during mouse oocyte maturation
and preimplantation embryo development. Gene Expr Patterns.
18:16–20. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang Q, Sztukowska M, Ojo A, Scott DA,
Wang H and Lamont RJ: FOXO responses to Porphyromonas gingivalis in
epithelial cells. Cell Microbiol. 17:1605–1617. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schmidt M, Fernandez de Mattos S, van der
Horst A, Klompmaker R, Kops GJ, Lam EW, Burgering BM and Medema RH:
Cell cycle inhibition by FoxO forkhead transcription factors
involves downregulation of cyclin D. Mol Cell Biol. 22:7842–7852.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang H and Tindall DJ: Dynamic FoxO
transcription factors. J Cell Sci. 120:2479–2487. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Y, Gan B, Liu D and Paik JH: FoxO
family members in cancer. Cancer Biol Ther. 12:253–259. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Myatt SS, Wang J, Monteiro LJ, Christian
M, Ho KK, Fusi L, Dina RE, Brosens JJ, Ghaem-Maghami S and Lam EW:
Definition of microRNAs that repress expression of the tumor
suppressor gene FOXO1 in endometrial cancer. Cancer Res.
70:367–377. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gao F and Wang W: MicroRNA-96 promotes the
proliferation of colorectal cancer cells and targets tumor protein
p53 inducible nuclear protein 1, forkhead box protein O1 (FOXO1)
and FOXO3a. Mol Med Rep. 11:1200–1206. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wu L, Li H, Jia CY, Cheng W, Yu M, Peng M,
Zhu Y, Zhao Q, Dong YW, Shao K, et al: MicroRNA-223 regulates FOXO1
expression and cell proliferation. FEBS Lett. 586:1038–1043. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Roy UK, Henkhaus RS, Ignatenko NA, Mora J,
Fultz KE and Gerner EW: Wild-type APC regulates caveolin-1
expression in human colon adenocarcinoma cell lines via FOXO1a and
C-myc. Mol Carcinog. 47:947–955. 2008. View Article : Google Scholar : PubMed/NCBI
|