Introduction
Breast cancer is the most common malignancy and the
leading cause of cancer-associated mortality in women worldwide
(1,2).
Each year, >1.5 million new cases of breast cancer are reported
around the world (3), and it has been
estimated that 10–12% of women will develop breast cancer over the
course of their lives (4). Although
relapse-free survival of patients with breast cancer has been
improved by modern chemo-, endocrine and targeted therapies,
improved therapeutic options for advanced disease are urgently
required (5).
Oncolytic herpes simplex virus-1 (oHSV-1) is a
potential therapeutic vector for human cancer (6,7). Several
oncolytic HSV mutants have already been used in clinical trials for
various solid tumours (7–9). Talimogene laherparepvec (T-VEC), an oHSV
expressing granulocyte-macrophage colony-stimulating factor, was
approved by the U.S. Food and Drug Administration for the treatment
of melanoma in 2015 (10). G47Δ is a
third-generation oHSV-1 that lacks the γ34.5 neurovirulence gene
and possesses an additional deletion of the α47 gene (11,12).
Previous studies have demonstrated that G47Δ can effectively target
breast cancer cells in vitro and in vivo (13–16).
However, certain breast cancer cell lines remain insensitive to
G47Δ (data not shown). Infected cell protein 34.5 (ICP34.5),
encoded by the γ34.5 gene, has diverse activities: It is critical
for the replication and neurovirulence of oHSV-1 in humans and in
animal models (17). oHSV-1 mutants
without γ34.5 were highly attenuated for neurovirulence, although
deletion of γ34.5 also compromises the replication of the virus
(18). An early report found that a
stretch of 64 amino acids at the C-terminus of the γ34.5 gene was
homologous to the C termini of murine protein phosphatase 1
regulatory subunit 15A (also known as MyD116/GADD34) (19). In addition, the C terminus of MyD116
can substitute for the corresponding domain of the γ34.5 gene of
oHSV-1 to preclude the premature shutoff of total protein synthesis
in infected human cells (19).
Therefore, we hypothesised that the insertion of the MyD116
C-terminus might enhance the replication and virulence of G47Δ.
To test this hypothesis, a bacterial artificial
chromosome (BAC) and two recombinase systems (Cre/loxP and
FLPE/FRT) were used to reconstruct a novel oHSV-1 mutant, GD116, in
which a chimeric gene (γ34.5-MyD116) consisting of the N-terminus
of the γ34.5 gene and the C-terminus of the MyD116 gene was
inserted (19). In this study, the
novel vector, GD116, exhibited enhanced anti-tumour effects on
breast cancer in vitro.
Materials and methods
Cells
The human breast cancer cell lines MCF-7, SK-BR-3
and MDA-MB-231 were provided by Dr Musheng Zeng (State Key
Laboratory of Oncology in Southern China, Sun Yat-sen University
Cancer Center, Guangzhou, China), and the MDA-MB-468 cell line was
obtained from Dr Xiaoming Xie (Sun Yat-sen University Cancer
Center, Guangzhou, China). The Vero cell line (Cercopithecus
aethiops kidney cells) was purchased from the Type Culture
Collection of the Chinese Academy of Sciences (Shanghai, China).
All cell types were cultured in Dulbecco's modified Eagle's medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
which was supplemented with 4.5 g/l glucose and 10% foetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) at 37°C in an
atmosphere containing 5% CO2.
Plasmids
The plasmid pRB4871, containing the γ34.5-MyD116
chimaera was kindly provided by Professor Bernard Roizman (Marjorie
B. Kovler Viral Oncology Laboratories, University of Chicago,
Chicago, IL, USA) (19). The
pG47Δ-BAC backbone plasmid and the pVec91 shuttle plasmid were
obtained from Professor Samuel D. Rabkin (Molecular Neurosurgery
Laboratory, Massachusetts General Hospital, Boston, MA, USA), and
the pG47Δ-BAC backbone plasmid was created by a two-step
replacement procedure, in which G47Δ DNA was cloned into a BAC
vector for propagation in Escherichia coli, as described
previously (20). The γ34.5-MyD116
chimaera fragment from pRB4871 was ligated into the
BamHI-StuI site of pVec91 [containing LacZ, loxP, and
FRT sites; multiple cloning sites; lambda stuffer sequences; and a
cytomegalovirus (CMV) promoter] to generate pVec91-116.
Virus construction
The recombinant oHSV-1 vectors were constructed
using a BAC and Cre/loxP and FLP/FRT recombinase systems, as
reported previously (20,21). Mixtures of the pG47Δ-BAC plasmid (2
µg) and pVec91-116 or pVec91 (200 ng) were incubated with Cre
recombinase (New England BioLabs, Ipswich, MA, USA) at 37°C for 30
min and then electroporated into E. coli DH10B using a Gene
Pulser Xcell™ (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
bacteria were streaked onto LB plates containing chloramphenicol
(15 µg/ml; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and
kanamycin (30 µg/ml; Sigma-Aldrich; Merck KGaA), and positive
clones containing the pG47Δ-BAC-116 recombinant and pG47Δ-BAC-empty
plasmids were selected. Next, the recombinant plasmids were
digested with HindIII and separated by electrophoresis on
0.75% agarose gels in TBE buffer (Tris-base, boric acid and EDTA)
at 2.5 cm/V for 12 h with a 1 kb DNA extension ladder (Invitrogen;
Thermo Fisher Scientific, Inc.). Co-transfection of pG47Δ-BAC-116
or pG47Δ-BAC-empty and a FLP recombinase expression plasmid, pOG44
(Invitrogen; Thermo Fisher Scientific, Inc.), into Vero cells was
performed with Lipofectamine 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Following this, the removal of the FRT-flanked
BAC sequences resulted in the generation of recombinant viruses.
The progeny viruses were further selected by limiting dilution and
were finally designated ‘GD116’ and ‘GD-empty’.
Analyses of viral DNA
Viral DNA was isolated using the Hirt Supernatant
DNA Purification kit (GenMed Scientifics Inc., Shanghai, China),
and polymerase chain reaction (PCR) was performed using recombinant
Taq DNA polymerase (Takara Bio, Inc., Otsu, Japan). Primers
(CMV forward 5′-ACTGCTTACTGGCTTATCG-3′ and reverse,
5′-GTCTAACTCGCTCGTCTC-3′) were used to amplify a specific 113 bp
fragment for detection of the CMV sequence. Primers (γ34.5-MyD116
forward, 5′-GCGGCTCAGATTGTTCAA-3′ and γ34.5-MyD116 reverse,
5′-CGGACTGTGGAAGAGATG-3′) were used to amplify a 284 bp product
that was specific for the γ34.5-MyD116 chimaera. The samples were
initially denatured for 5 min at 94°C; followed by 35 cycles of
denaturation at 94°C for 30 sec, re-naturation at 52°C for 30 sec
and extension at 72°C for 20 sec; with a final extension at 72°C
for 10 min. pVec91 or pRB4871 plasmids and PBS were used as the
positive and blank controls, respectively. The PCR products were
separated by size in 1% agarose gels.
Western blotting
A total of 5×106 Vero cells were mock
treated or treated with viruses at an MOI (multiplicity of
infection) of 2 for 24 h, and then cells were lysed using a Whole
Cell Lysis kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China).
A total of 30 µg protein (quantified using the BCA protein
quantification kit; Nanjing KeyGen Biotech Co., Ltd.) was separated
by 15% SDS-PAGE and transferred to a 0.22-µm thick nitrocellulose
membrane (EMD Millipore, Billerica, MA, USA). Following blocking
with 5% non-fat dry milk in TBS-Tween-20 (TBS-T, 0.1% Tween) for 1
h at room temperature, the membrane was incubated overnight at 4°C
with primary antibodies against ICP34.5 (amino terminus) (diluted
1:500, rabbit polyclonal; cat no. HX5191; TSZ Biosciences, San
Fransisco, CA, USA; http://www.tsz-biological.com/) or β-actin (diluted
1:1,000; rabbit monoclonal; cat no. 4970; Cell Signaling
Technology, Inc., Danvers, MA, USA). The membrane was then washed
and blotted with a horseradish peroxidase (HRP)-linked anti-rabbit
secondary antibody (diluted 1:6,000; cat no. 7074; Cell Signaling
Technology, USA) for 1 h at room temperature. The Protein antibody
complexes were visualized with the chemiluminescent HRP substrate
(EMD Millipore) using a chemiluminescence imaging system
(Tanon-5200; Tanon, Guangzhou, China).
Virus replication assay
Cells were seeded in 12-well plates at
1×105 cells per well and infected at 70–80% confluence
with GD116 or GD-empty at a MOI of 0.1 (Vero, MCF-7, SK-BR-3 and
MDA-MB-468) or 0.3 (MDA-MB-231). The inoculum was removed after 2 h
and replaced with DMEM supplemented with 1% heat-inactivated FBS
(iFBS). The cells and the supernatant were harvested at the
indicated times (24 and 48 h) post-infection, processed with three
freeze/thaw cycles, and virus titres were determined by plaque
assays on Vero cells. In brief, Vero cells were seeded in 12-well
plates at 1×105 cells/well and infected with 500 µl
virus dilute (10−4/ml, 10−5/ml,
10−6/ml or 10−7/ml) at 100% confluence. Then,
the infected Vero cells were cultured at 37°C in 5% CO2
for an additional 48 h prior to subsequent X-gal staining for
plaque counting. The X-gal staining was performed using the in
situ β-galactosidase staining kit (Beyotime Institute of
Biotechnology, Haimen, China) according to the manufacturer's
protocol. The virus titres were calculated using the following
equation: Virus titres=plaque numbers/0.5 × dilution ratio.
Experiments were repeated at least three times.
Cell susceptibility assay
Cells were seeded in 48-well plates at 8,000 cells
per well. After 16 h, the cells were infected with either GD116 or
GD-empty at MOIs of 0.01 and 0.1 (MCF-7, SK-BR-3 and MDA-MB-468) or
at MOIs of 0.1 and 0.3 (MDA-MB-231). Cells were incubated for up to
5 days in DMEM medium supplemented with 1% iFBS at 37°C. The cell
viability was assessed daily with an MTT assay and expressed as a
percentage of the mock-infected control. Formazan crystals were
dissolved using 400 µl DMSO and the visualization wavelength was
490 nm. Meanwhile, infection was monitored using β-galactosidase
activity via X-gal staining at the indicated times (1, 2, 3, 4 and
5 days post-infection). Experiments were repeated at least three
times for each condition in quadruplicate.
Statistical analysis
Student's t-test was used for statistical analyses.
P<0.05 were considered to be statistically significant. Data are
presented as the mean ± standard deviation. SPSS 17.0 software
(SPSS, Inc., Chicago, IL, USA) was used for all of the statistical
analyses.
Results
Cloning of the γ34.5-MyD116 chimaera
into the shuttle plasmid
The genomic structure of the mutants is depicted in
Fig. 1A. The pRB4871 plasmid was
cleaved using the BamHI and StuI restriction
endonucleases, and then a 1.51 kb fragment containing the
γ34.5-MyD116 chimaera was purified from an agarose gel and
directionally cloned into the pVec91 vector to generate pVec91-116.
To identify the recombinant plasmid pVec91-116, pRB4871 and
pVec91-116 were digested with BamHI and StuI, and the
products were separated on a 1% agarose gel. As shown in the map of
double restriction endonuclease digestions (Fig. 1B), the desired 1.51 kb bands on an
agarose gel confirmed the successful construction of the
recombinant pVec91-116 vector.
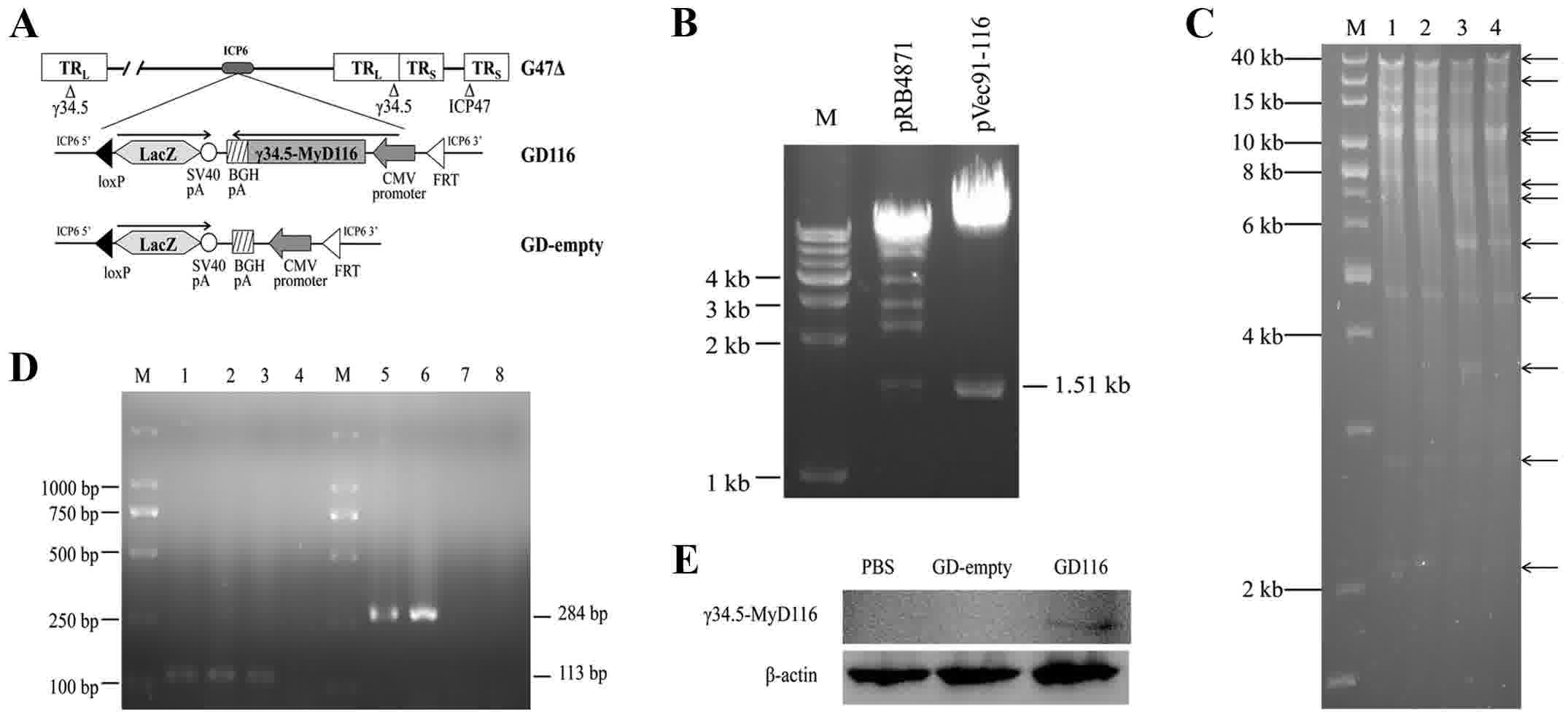 | Figure 1.Construction and characterization of
the GD116 and GD-empty vectors. (A) A diagram depicting the genomic
structure of GD116 and GD-empty. (B) Restriction digestion with
BamHI and StuI revealed the presence of a 1.51-kb
insert from the plasmids pRB4871 and pVec91-116. Lane M, 1 kb DNA
ladder. (C) HindIII restriction digestion confirmed the
structures of the integrated plasmids obtained following the first
step of Cre recombination. Lane M, 1 kb DNA ladder; lanes 1 and 2,
pG47Δ-BAC; lane 3, pG47Δ-BAC-Vec91-empty; lane 4,
pG47Δ-BAC-Vec91-116. (D) Polymerase chain reaction amplification of
the CMV and γ34.5-MyD116 sequences using recombinant virus genomic
DNAs as a template. Lane M, DL2000 ladder; lane 1, pVec91; lanes 2
and 6, GD116; lanes 3 and 7, GD-empty; lanes 4 and 8, PBS; lane 5,
pRB4871. (E) Expression of the γ34.5-MyD116 protein was confirmed
by western blotting. Vero cells were treated with PBS (left),
GD-empty (middle) or GD116 (right). CMV, cytomegalovirus. |
Construction of recombinant oHSVs with
or without the γ34.5-MyD116 chimaera
The generation of two novel oHSV-1 mutants was
performed by a previously described two-step replacement procedure
(22). In the first step, the shuttle
plasmid pVec91-116 or pVec91-empty were integrated into pG47Δ-BAC
using Cre recombinase in a tube and an integrated BAC clone
(pG47Δ-BAC-Vec91-116 or pG47Δ-BAC-empty) was isolated in E.
coli by selection with chloramphenicol and kanamycin. The
integrated plasmids were collected and the DNA structures of the
plasmids were confirmed by gel analyses following Hind III
restriction endonuclease digestion (Fig.
1C).
In the second step, the integrated plasmid
pG47Δ-BAC-Vec91-116 or pG47Δ-BAC-empty and the FLP expression
plasmid were co-transfected into Vero cells, and the BAC backbone
and lambda stuffer sequences were excised by FLP recombinase. As a
result, the novel oHSV-1 mutants were generated and were designated
GD116 and GD-empty.
To verify the insertion of the target sequences into
the novel mutants, specific PCR analyses were conducted using
pVec91 and pRB4871 plasmids as the positive controls. The PCR
products were checked by electrophoresis on 1% agarose gels. GD116
and GD-empty produced the desired 113 bp band on an agarose gel
when used as templates to amplify the CMV sequence (Fig. 1D). However, only GD116 generated the
expected size band of 284 bp when γ34.5-MyD116-specific PCR
products were detected (Fig. 1D). To
determine the expression of the γ34.5-MyD116 protein, Vero cells
were treated with PBS, GD-empty or GD116. Western blot analysis
revealed that the expression of γ34.5-MyD116 was shown only in the
GD116-treated group (Fig. 1E). In
addition, the two mutants could form blue plaques on Vero cells by
X-gal staining 48 h after infection, which confirmed the insertion
and expression of the LacZ gene (data not shown).
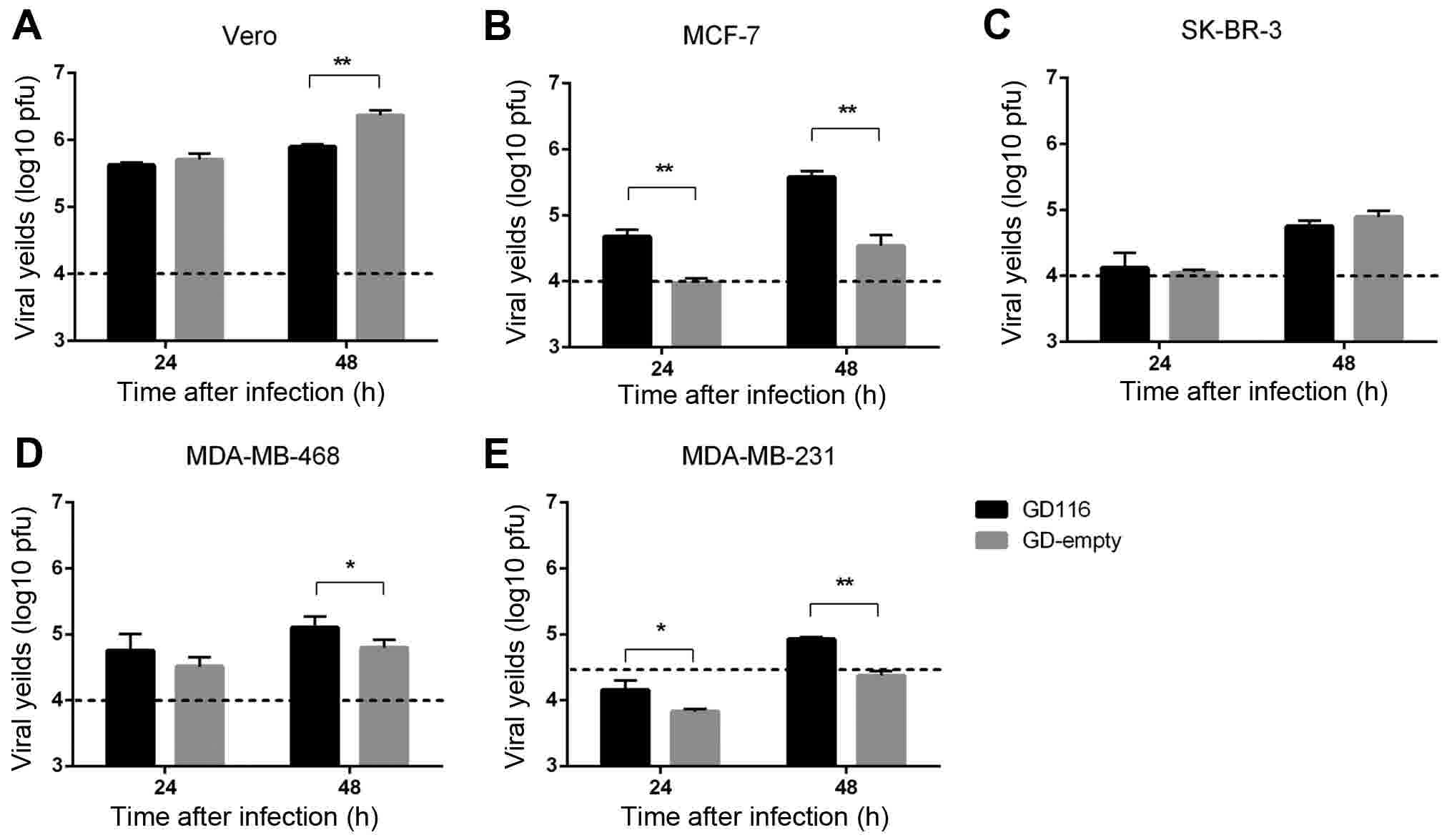
Replication of GD116 and GD-empty in
vitro
Viral replication is one of the key determinants of
oHSV-1 efficacy against tumour cells. The replication of the two
mutants was compared in Vero cells and a panel of human breast
cancer cell lines at 24 and 48 h after infection (Figs. 2A-E). Compared with GD-empty, GD116
had a significantly increased viral yield in breast cancer MCF-7
and MDA-MB-231 cell lines at 24 and 48 h (Fig. 2B and E). Additionally, the viral
yields of GD116 in MDA-MB-468 cells were greater than that of
GD-empty at 48 h (Fig. 2D). However,
the insertion of γ34.5-MyD116 did not significantly increase the
replication rate of oHSV-1 in SK-BR-3 cells (Fig. 2C), and even reduced the viral
replication in Vero cells (Fig.
2A).
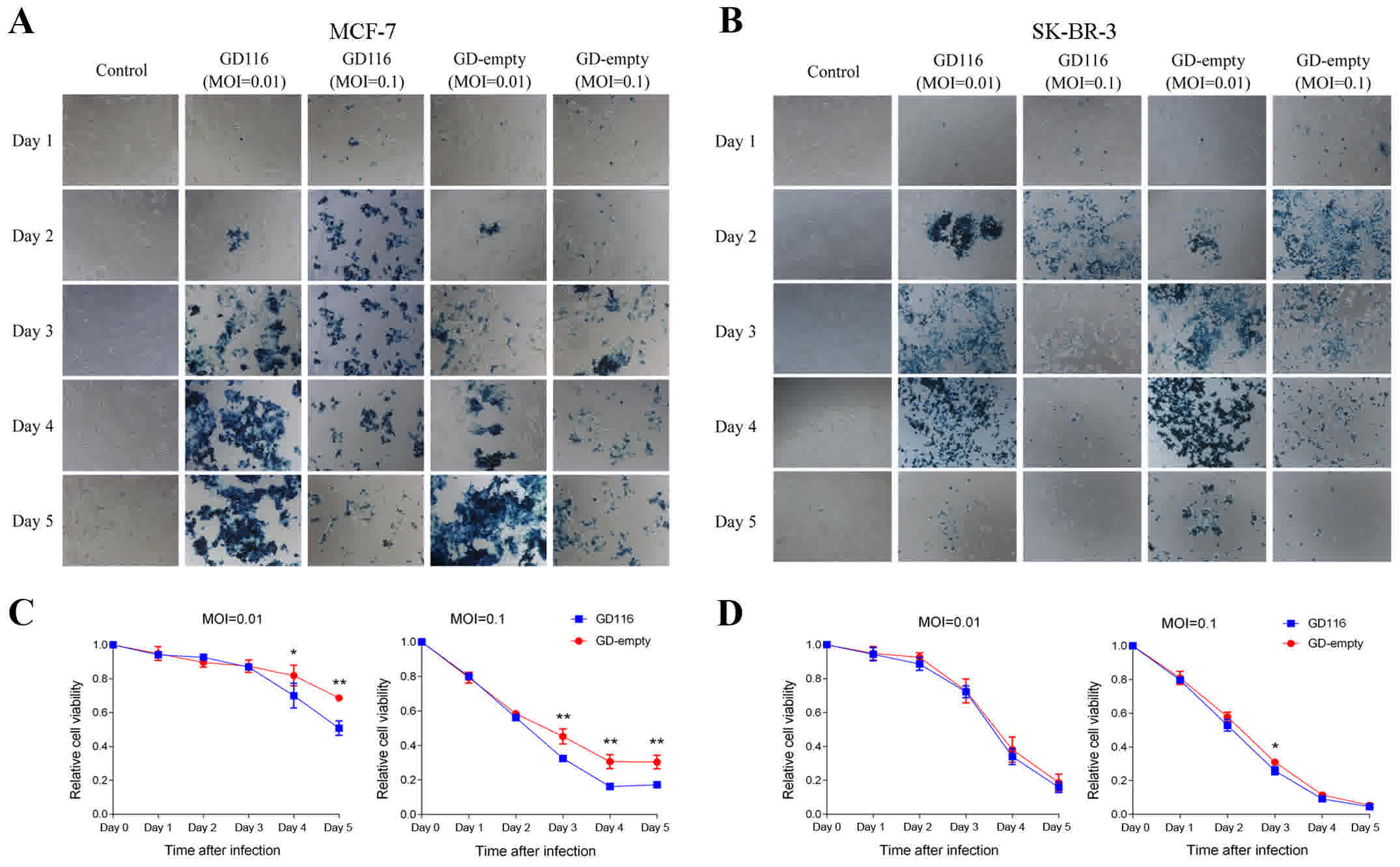
In vitro cytotoxicity of GD116 and
GD-empty in breast cancer cell lines
Next, the oncolytic activity of these two mutants
was examined in human breast cancer cell lines. The breast cancer
cells MCF-7, SK-BR-3 and MDA-MB-468 were infected with GD116 or
GD-empty at MOIs of 0.01 and 0.1, whereas MDA-MB-231 cells were
infected with these two mutants at MOIs of 0.1 and 0.3. In
addition, cells infected with the virus could express LacZ and thus
were stained blue by X-gal; staining of MCF-7 and SK-BR-3 cells
with X-gal was presented in Fig. 3A and
B. Similar to what was observed with virus replication, GD116
was more effective at inhibiting the growth of MCF-7 cells at MOIs
of 0.01 and 0.1 (Fig. 3C). On the
fifth day after infection with GD116 at MOIs of 0.01 and 0.1, 49.2
and 82.8% of MCF-7 cells were killed, respectively, which was
higher than 31.3 and 69.6%, respectively, with GD-empty infection
(Fig. 3C). However, only a slightly
increased efficacy was observed with GD116 than with GD-empty in
SK-BR-3 cells (74.2 vs. 69.0%, respectively) on day 3 at an MOI of
0.1 (Fig. 3D). Aside from that
effect, no significant difference in cytotoxicity was observed
between the two mutants at the other indicated times (Fig. 3D).
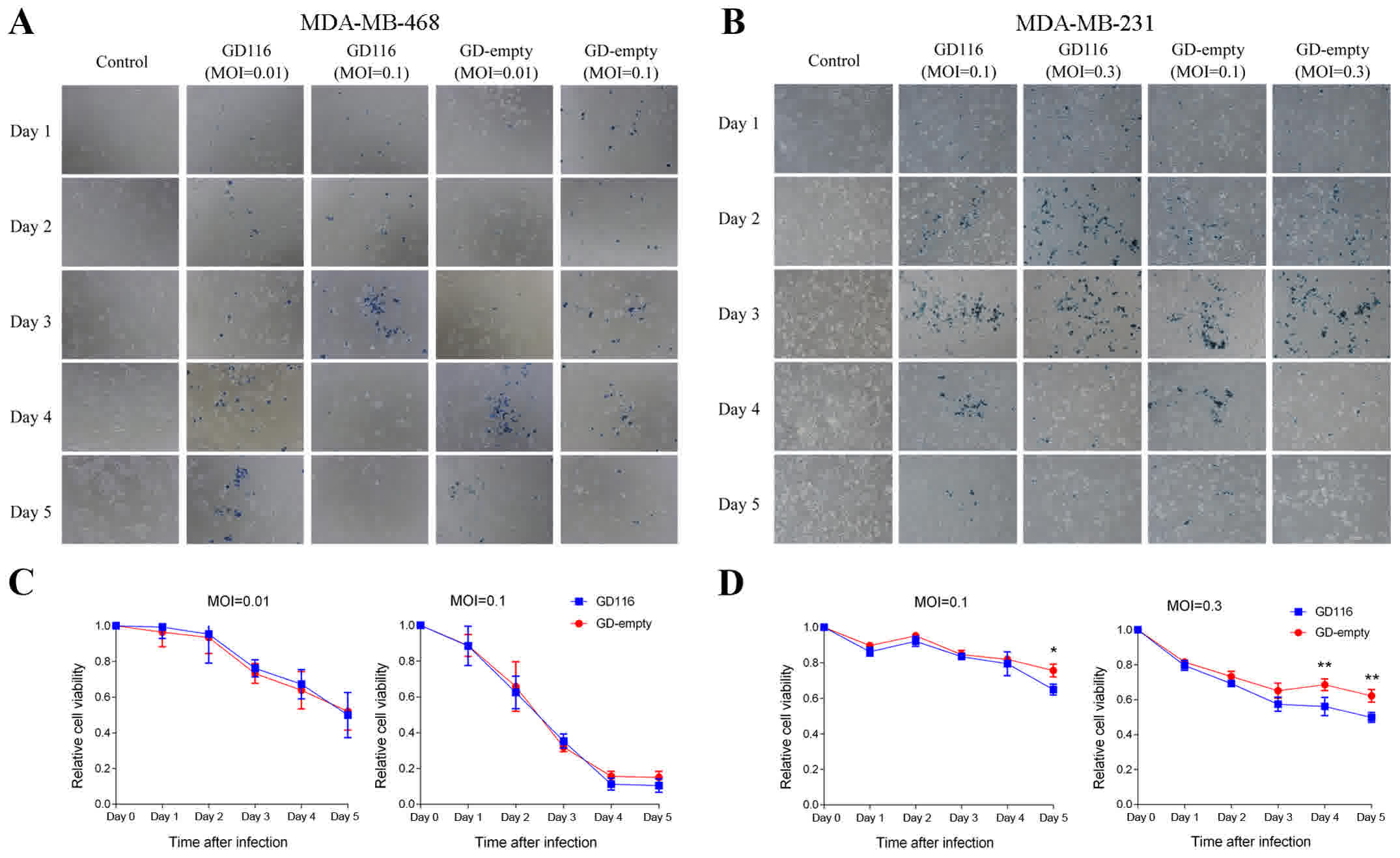
X-gal staining of MDA-MB-468 and MDA-MB-231 cells
was presented in Fig. 4A and B. As
shown in Fig. 4C, GD116 had a similar
cytotoxic effect to GD-empty in MDA-MB-468 cells. However, the
insertion of the γ34.5-MyD116 chimaera could significantly increase
the oncolytic activity of GD116 on MDA-MB-231 cells (Fig. 4D). On day 5 after infection, 35.0 and
50.2% of MDA-MB-231 cells were killed by GD116 at MOIs of 0.1 and
0.3, respectively; however, the killing activity of GD-empty at the
different MOIs was only 24.3 and 37.8%, respectively (Fig. 4D). In vitro, the spread of
oHSV-1 was associated with cytotoxicity; therefore, X-gal staining
visually showed the difference in oncolytic activity between GD116
and GD-empty in different breast cancer cell lines (Figs. 3A and B, and 4A and B).
Discussion
oHSV-1 vectors that specifically replicate in and
kill tumour cells, sparing normal cells, are promising cancer
therapeutic agents. In the oHSV-1 genome, γ34.5 is transcribed as a
leaky-late gene that encodes ICP34.5, whose expression can be
detected as early as 2–3 h post-infection (23). ICP34.5 is involved in multiple aspects
of viral pathogenesis, and one of its key roles is to promote
neurovirulence. oHSV-1 mutants with deletions in ICP34 are unable
to replicate in neuronal cells and are not neurovirulent (24). In this regard, ICP34.5 has currently
been deleted in all oHSV vectors in clinical trials for treating
malignant gliomas, such as 1716 and G207 (18,25).
However, at the C-terminus of ICP34.5, a region homologous to
murine MyD116 and GADD34 can bridge phosphorylated eukaryotic
initiation factor 2α (eIF2α) and protein phosphatase-1α (PP1α) to
prevent the accumulation of the former and dephosphorylate it
(26,27). As a result, ICP34.5 can counteract
PKR-mediated innate immune responses and allow translation to
proceed (26,28). Therefore, mutants with γ34.5 gene
deletions reveal markedly reduced lethality in the peripheral or
intracerebral routes of infection but replicate poorly in certain
confluent cell types in vitro (23). Thus, it is necessary to reconstruct
novel oHSV-1 mutants that maintain low neurovirulence yet replicate
efficiently in cancer cells.
The present study used a BAC-based recombinase
system to reconstruct novel oHSV-1 mutants. This recombinase system
enabled the rapid generation of recombinant oHSV-1 vectors with the
desired transgene inserted in the place of the deleted ICP6 locus.
The C-terminus of MyD116/GADD34 was used to substitute that of
γ34.5 and to create a novel oHSV-1 mutant, GD116. Compared with
GD-empty, the mutation enhanced the replication and increased the
cytotoxicity of GD116 in breast cancer MCF-7 and MDA-MB-231 cell
lines. Although the insertion of the γ34.5-MyD116 chimaera promoted
the replication of the virus in MDA-MB-468 cells, no increased
cytotoxic effect was obtained. The insertion of γ34.5-MyD116
slightly inhibited viral replication in Vero cells, but the viral
cytotoxicity was not attenuated in cancer cells. Therefore, the
murine MyD116/GADD34 gene could be used to substitute the γ34.5
gene to promote the cytotoxicity of oHSV-1 without increasing the
neurovirulence. However, substitution of the carboxyl-terminus of
the γ34.5 gene with the corresponding domain of murine MyD116 did
not markedly enhance the virulence of G47Δ in breast cancer cell
lines, and even SK-BR-3 cells were not sensitive to the mutation of
G47Δ.
Recently, a region in the N-terminus of γ34.5 [amino
acids (aa) 68 to 87] that interacts with Beclin 1 and inhibits
autophagy, which contributes to neurovirulence in a PKR-dependent
fashion, has been identified (29,30).
Deletion of the Beclin 1-binding domain (BBD) (aa 68–87) resulted
in mutants that could induce autophagy in neurons and was
attenuated for neurovirulence following intracerebral inoculation
(18). In addition, BBD deletion
mutants remain able to dephosphorylate eIF2α and inhibit host
protein shutoff, and thus the replication and cytotoxicity are not
attenuated. Therefore, promoting the anti-tumour efficacy by
expressing BBD-deleted ICP34.5 for oHSV-1 mutants with the deletion
of the two copies of the γ34.5 gene, such as G47Δ may represent a
promising strategy for generation of a therapeutic vector for the
treatment of breast cancer.
Acknowledgements
The authors would like to thank Professor Bernard
Roizman for providing the pRB4871 plasmid and Professor Samuel D.
Rabkin for providing the pG47Δ-BAC plasmid and the shuttle plasmid
pVec91.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81172523 and 81372815;
principle investigator, Renbin Liu).
Availability of data and materials
The datasets generated and analyzed in the present
study are included in this published article.
Authors' contributions
LC, HJ and RL conceived and designed the experiment.
LC, HJ and JF performed the experiments. JW, PH and YR collected
the data and performed the statistical analyses. LC wrote the
manuscript. JW and RL reviewed and revised the manuscript. All
authors read and approved the final manuscript.
Ethics and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huang J, Li H and Ren G:
Epithelial-mesenchymal transition and drug resistance in breast
cancer (Review). Int J Oncol. 47:840–848. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Finn RS, Crown JP, Lang I, Boer K,
Bondarenko IM, Kulyk SO, Ettl J, Patel R, Pinter T9, Schmidt M, et
al: The cyclin-dependent kinase 4/6 inhibitor palbociclib in
combination with letrozole versus letrozole alone as first-line
treatment of oestrogen receptor-positive, HER2-negative, advanced
breast cancer (PALOMA-1/TRIO-18): A randomised phase 2 study.
Lancet Oncol. 16:25–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kleibl Z and Kristensen VN: Women at high
risk of breast cancer: Molecular characteristics, clinical
presentation and management. Breast. 28:136–144. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hartkopf AD, Fehm T, Wallwiener D and
Lauer UM: Oncolytic virotherapy of breast cancer. Gynecol Oncol.
123:164–171. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kuruppu D and Tanabe KK: HSV-1 as a novel
therapy for breast cancer meningeal metastases. Cancer Gene Ther.
22:506–508. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Andtbacka RH, Kaufman HL, Collichio F,
Amatruda T, Senzer N, Chesney J, Delman KA, Spitler LE, Puzanov I,
Agarwala SS, et al: Talimogene laherparepvec improves durable
response rate in patients with advanced melanoma. J Clin Oncol.
33:2780–2788. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Campadelli-Fiume G, Petrovic B, Leoni V,
Gianni T, Avitabile E, Casiraghi C and Gatta V: retargeting
strategies for oncolytic herpes simplex viruses. Viruses. 8:632016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang W, Fulci G, Wakimoto H, Cheema TA,
Buhrman JS, Jeyaretna DS, Rachamimov Stemmer AO, Rabkin SD and
Martuza RL: Combination of oncolytic herpes simplex viruses armed
with angiostatin and IL-12 enhances antitumor efficacy in human
glioblastoma models. Neoplasia. 15:591–599. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
U.S. Food and Drug Administration: FDA
approves first-of-its-kind product for the treatment of melanoma.
http://www.webcitation.org/6drvCltG7October
27–2015
|
|
11
|
Todo T: Active immunotherapy: Oncolytic
virus therapy using HSV-1. Adv Exp Med Biol. 746:178–186. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nigim F, Esaki SI, Hood M, Lelic N, James
MF, Ramesh V, Stemmer-Rachamimov A, Cahill DP, Brastianos PK,
Rabkin SD, et al: A new patient-derived orthotopic malignant
meningioma model treated with oncolytic herpes simplex virus. Neuro
Oncoly. 18:1278–1287. 2016. View Article : Google Scholar
|
|
13
|
Fan J, Jiang H, Cheng L and Liu R: The
oncolytic herpes simplex virus vector, G47Δ, effectively targets
tamoxifen-resistant breast cancer cells. Oncol Rep. 35:1741–1749.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zeng WG, Li JJ, Hu P, Lei L, Wang JN and
Liu RB: An oncolytic herpes simplex virus vector, G47Δ, synergizes
with paclitaxel in the treatment of breast cancer. Oncol Rep.
29:2355–2361. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu R, Varghese S and Rabkin SD: Oncolytic
herpes simplex virus vector therapy of breast cancer in C3(1)/SV40
T-antigen transgenic mice. Cancer Res. 65:1532–1540. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang J, Hu P, Zeng M, Rabkin SD and Liu R:
Oncolytic herpes simplex virus treatment of metastatic breast
cancer. Int J Oncol. 40:757–763. 2012.PubMed/NCBI
|
|
17
|
Alexander DE, Ward SL, Mizushima N, Levine
B and Leib DA: Analysis of the role of autophagy in replication of
herpes simplex virus in cell culture. J Virol. 81:12128–12134.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kanai R, Zaupa C, Sgubin D, Antoszczyk SJ,
Martuza RL, Wakimoto H and Rabkin SD: Effect of γ34.5 deletions on
oncolytic herpes simplex virus activity in brain tumors. J Virol.
86:4420–4431. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He B, Chou J, Liebermann DA, Hoffman B and
Roizman B: The carboxyl terminus of the murine MyD116 gene
substitutes for the corresponding domain of the gamma(1)34.5 gene
of herpes simplex virus to preclude the premature shutoff of total
protein synthesis in infected human cells. J Virol. 70:84–90.
1996.PubMed/NCBI
|
|
20
|
Fukuhara H, Ino Y, Kuroda T, Martuza RL
and Todo T: Triple gene-deleted oncolytic herpes simplex virus
vector double-armed with interleukin 18 and soluble B7-1
constructed by bacterial artificial chromosome-mediated system.
Cancer Res. 65:10663–10668. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tsuji T, Nakamori M, Iwahashi M, Nakamura
M, Ojima T, Iida T, Katsuda M, Hayata K, Ino Y, Todo T and Yamaue
H: An armed oncolytic herpes simplex virus expressing
thrombospondin-1 has an enhanced in vivo antitumor effect against
human gastric cancer. Int J Cancer. 132:485–494. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kuroda T, Martuza RL, Todo T and Rabkin
SD: Flip-Flop HSV-BAC: Bacterial artificial chromosome based system
for rapid generation of recombinant herpes simplex virus vectors
using two independent site-specific recombinases. BMC Biotechnol.
6:402006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Korom M, Davis KL and Morrison LA: Up to
four distinct polypeptides are produced from the γ34.5 open reading
frame of herpes simplex virus 2. J Virol. 88:11284–11296. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Brown SM, Harland J, MacLean AR, Podlech J
and Clements JB: Cell type and cell state determine differential in
vitro growth of non-neurovirulent ICP34.5-negative herpes simplex
virus types 1 and 2. J Gen Virol. 75:2367–2377. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zemp FJ, Corredor JC, Lun X, Muruve DA and
Forsyth PA: Oncolytic viruses as experimental treatments for
malignant gliomas: Using a scourge to treat a devil. Cytokine
Growth Factor Rev. 21:103–117. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
He B, Gross M and Roizman B: The
gamma(1)34.5 protein of herpes simplex virus 1 complexes with
protein phosphatase 1alpha to dephosphorylate the alpha subunit of
the eukaryotic translation initiation factor 2 and preclude the
shutoff of protein synthesis by double-stranded RNA-activated
protein kinase. Proc Natl Acad Sci USA. 94:843–848. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li Y, Zhang C, Chen X, Yu J, Wang Y, Yang
Y, Du M, Jin H, Ma Y, He B and Cao Y: ICP34.5 protein of herpes
simplex virus facilitates the initiation of protein translation by
bridging eukaryotic initiation factor 2alpha (eIF2alpha) and
protein phosphatase 1. J Biol Chem. 286:24785–24792. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Davis KL, Korom M and Morrison LA: Herpes
simplex virus 2 ICP34.5 confers neurovirulence by regulating the
type I interferon response. Virology. 468–470:330–339. 2014.
View Article : Google Scholar
|
|
29
|
Tang S, Guo N, Patel A and Krause PR:
Herpes simplex virus 2 expresses a novel form of ICP34.5, a major
viral neurovirulence factor, through regulated alternative
splicing. J Virol. 87:5820–5830. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Orvedahl A, Alexander D, Tallóczy Z, Sun
Q, Wei Y, Zhang W, Burns D, Leib DA and Levine B: HSV-1 ICP34.5
confers neurovirulence by targeting the Beclin 1 autophagy protein.
Cell Host Microbe. 1:23–35. 2007. View Article : Google Scholar : PubMed/NCBI
|