Introduction
Glioblastoma is the most frequently occurring
primary malignant brain tumor in adults and exhibits a spectrum of
aberrantly aggressive phenotypes in the USA (1,2). The
median survival time is generally <1 year from the time of
diagnosis, and even in the most favorable situations, the majority
of patients succumb within 2 years (3–5). At
present, treatment options include surgery, radiation and
chemotherapy with methylating agents and nitrosoureas (6). Unfortunately, the treatment options
often do not result in a cure, subsequently leading to tumor
progression and mortality of the patient. One of the reasons for
failure of treatment is due to de novo or acquired
resistance to chemotherapeutic agents. However, the molecular
mechanisms underlying chemotherapy resistance of glioblastoma cells
remain unknown.
Fibroblast activation protein α (FAP-α) is a member
of the serine integral membrane peptidases (SIMPs) family, which
also includes prolyl endopeptidase, dipeptidyl peptidase IV and
dipeptidyl peptidase IIX. These peptidases are inducible, specific
for proline-containing peptides and are active on the cell surface
(7–9).
FAP-α is significantly associated with poor outcome in patients
with breast cancer (10). In
vitro, it may promote proliferation and inhibit migration of
breast cancer cells, potentially by regulating the focal adhesion
kinase pathway, and its overexpression is associated with
neoplastic progression of esophageal lesions (10). FAP-α is highly expressed on the
surface of glioma cells and contributes to diffuse glioma invasion
through extracellular matrix components (11). However, to the best of our knowledge,
its role has not been studied in cancer-initiating cells (CICs) or
chemotherapy resistance of glioblastoma.
MicroRNAs (miRNAs/miRs) are a class of small
noncoding RNAs (~22 nucleotides) and negatively regulate
protein-coding gene expression by targeting mRNA degradation or
translation inhibition (12–14). Deregulation of miRNAs has been
implicated in the development and progression of glioblastoma, and
they serve pivotal roles in development, particularly in modulating
stem cell-specific pathways (15–17).
Previously, it was demonstrated that miR-204 may be a useful drug
target in the treatment and diagnosis of glioblastoma multiforme
(GBM) (9,18–20). In
the present study, it was identified that miR-204 may reverse
temozolomide resistance and inhibit CICs phenotypes by degrading
FAP-α in glioblastoma.
Materials and methods
Patients
Between November 2013 and October 2015, 18 patients
(10 male, 8 female) with glioblastoma were enrolled at the
Department of Neurosurgery, Beijing Chaoyang Hospital. The mean age
was 57 years (range, 35–78 years). All tissues were examined
histologically, and pathologists confirmed the diagnosis. The
present study was approved by the ethics committee of Beijing
Chaoyang Hospital, and each patient signed an informed consent form
at the time of enrollment.
Human glioblastoma cell line
U251MG cells were purchased from the Institute of
Biochemistry and Cell Biology Institute of Shanghai, Chinese
Academy of Sciences (Shanghai, China) within 3 months of
experiments. To obtain temozolomide (TMZ)-resistant U251MG cells
(U251MG-R cells), U251MG cells were treated with increasing
concentrations of TMZ (10−7, 10−6 and
10−5 M). The U251MG-R cells were considered to be
established when colonies grew at similar rate in the presence or
absence of 10−5 M TMZ for 3 days (data not shown). The
half maximal inhibitory concentration (IC50) of U251MG-R
cells increased by 12-fold, as compared with the U251MG cells (data
not shown). Cells were cultured in normal culture medium which was
composed of Dulbecco modified Eagle medium (Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and
antibiotics (100 mg/ml penicillin; 100 U/ml streptomycin) in a 5%
CO2 incubator at 37°C.
FAP-α expressing plasmid/empty vector,
pre-miR-204/control miR and transfection experiments
FAP-α expressing plasmids and empty vectors were
donated by Dr. Chao Wang (Cardiff University-Peking University
Cancer Institute, Cardiff University School of Medicine, Cardiff,
Wales, UK) and produced as described previously (10). Pre-miR-204 (sequence:
5′-UUCCCUUUGUCAUCCUAUGCCU-3′)/control miR (sequence:
5′-UACCGUAUCUCUUCGUAAGCGU-3′) were purchased from Ambion (Ambion;
Thermo Fisher Scientific, Inc.). For transfection experiments,
U251MG-R cells were cultured in serum-free medium (DMEM) without
antibiotics at 60% confluence for 24 h in a 5% CO2
incubator at 37°C, and then transfected with 50 nM pre-miR-204 or
50 nM control miR using a transfection reagent (Lipofectamine 2000;
Thermo Fisher Scientific, Inc.) at room temperature according to
the manufacturer's protocol. Following incubation for 6 h at 37°C,
the medium was removed and replaced with normal culture medium for
48 h, unless otherwise specified. Subsequent experiments were
performed 48 h after transfection.
Western blotting
U251MG and U251MG-R cells and glioblastoma tissues
were washed once with PBS and lysed using a
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) supplemented with
phenylmethanesulfonyl fluoride (Beyotime Institute of
Biotechnology) for 30 min at 0°C. Protein samples were measured
using Bradford Protein Assay kit (Beyotime Institute of
Biotechnology) for 10 min at room temperature and boiled for 10 min
in SDS sample buffer (Beyotime Institute of Biotechnology). Then
protein extracts (10 µg per lane) were resolved through 8%
SDS-PAGE, transferred to polyvinylidene difluoride membranes
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) and blocked for 60
min at room temperature in 5% skim milk powder (w/v) in
NaCl/Pi at room temperature. Afterwards, the blots were
incubated with antibodies including rabbit anti-FAP-α (cat no.
ab173904; 1:500; Abcam, Cambridge, MA, USA), rabbit anti-signal
transducer and activator of transcription 3 (STAT3; cat no.
ab68153; 1:500; Abcam), rabbit anti-mouse double minute 2 homolog
(MDM2; cat no. ab38618; 1:500; Abcam), rabbit anti-prominin-1
(CD133; cat no. ab19898; 1:500; Abcam), rabbit
anti-tyrosine-protein kinase met (MET; cat no. ab51067; 1:500;
Abcam), rabbit anti-ras-related protein rab-22A (RAB22A; cat no.
ab137093; 1:500; Abcam), rabbit anti-EZRIN (cat no. ab4069; 1:500;
Abcam), rabbit anti-SRY-box 4 (SOX4; cat no. ab80261; 1:500;
Abcam), rabbit anti-activating transcription factor 2 (ATF2; cat
no. ab47476; 1:500; Abcam), rabbit anti-erythropoietin-producing
human hepatocellular receptors (EphB; cat no. ab196793; 1:500;
Abcam) or β-actin (cat no. ab5694; 1:500; Abcam) at 4°C, and then
with IRDye™-800 conjugated goat anti-rabbit secondary
antibody (cat no. ab191866; 1:10,000; Abcam) were used for 30 min
at room temperature. The specific proteins were visualized by
Odyssey™ Infrared Imaging system using Odyssey Image
Studio Software (Version 2009; Gene Company, Ltd., Hong Kong,
China).
MTT assay
To monitor resistance to TMZ, U251MG and U251MG-R
cells were treated with 10 µM TMZ for 24 h. An MTT assay was
performed as described previously (19). Data were analyzed with Origin 7.5
software (OriginLab, Northampton, MA, USA) to fit a sigmoidal
curve. IC50 is the TMZ concentration that reduces
proliferating cells by 50%.
Sphere formation assay
U251MG and U251MG-R cells (103/ml) in
serum-free RPMI-1640/1 mM Na-pyruvate (Hyclone; GE Healthcare Life
Sciences, Logan, UT, USA) were seeded on 0.5% agar pre-coated
6-well plates. After 10 days, half the medium was exchanged every
third day. Single spheres were selected and measured using a phase
contrast microscope (magnification, ×100) with scales (Eclipse
TS-100; Nikon Corporation, Tokyo, Japan).
Immunofluorescence staining
This protocol was performed as described previously
(9). Following transfection, the U251
MG-R cells were fixed in 4% paraformaldehyde for 15 min at room
temperature, and then blocked with goat serum blocking solution
(Tiangen Biotech Co., Ltd., Beijing, China) for 20 min at room
temperature. Then, a rabbit anti-FAP-α antibody (1:200 dilution;
Abcam) was added, and the mixtures were incubated in a humid
chamber overnight in a 5% CO2 incubator at 37°C.
Subsequent to washing 3 times with NaCl/Pi, cells were incubated
with goat anti-rabbit secondary antibody (conjugated with Alexa
Fluor® 488; cat no. ab150077; 1:10,000; Abcam) for 30
min at 37°C. Following washing with NaCl/Pi, the samples were
observed under a laser scanning confocal microscope (magnification,
×100; Olympus Corporation, Tokyo, Japan). The cells nuclei were
stained with DAPI (1 µg/ml) for 20 min at 37°C.
Bioinformatics
The analysis of potential microRNA target sites was
performed using the prediction algorithm miRanda (http://www.microrna.org/). Target mRNA Search was used
(date of access: 2010-11-01) using the following search terms:
Target mRNA=FAP (FAP-α) and species=Homo sapiens.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was performed to determine the expression of
miR-204 and FAP-α. Total RNA was extracted from glioblastoma
tissues and U251 MG-R cells using TRIzol® (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. Total RNA (500 ng) was quantitated at 260 nm and
reverse-transcribed into cDNA using the PrimeScript RT reagent kit
(Takara Biotechnology, Co., Ltd., Dalian, China) according to the
manufacturer's protocol, at 37°C for 15 min and 85°C for 30 sec.
qPCR was performed using the SYBR Premix Ex Taq™ kit (Takara
Biotechnology, Co., Ltd.) according to the manufacturer's protocol
in the ABI PRISM 7900HT system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The thermocycling conditions were as follows:
50°C for 2 min, 95°C for 10 min followed by 40 cycles with each
cycle consisting of 30 sec at 95°C, and 1 min at 60°C. Cycle
threshold (Cq) values were determined using SDS version 2.4
software (Applied Biosystems; Thermo Fisher Scientific, Inc.).
FAP-α expression levels were normalized to GAPDH expression using
the 2−ΔΔCq method (21).
The primer sequences were as follows: FAP-α forward,
5′-TTAGTCTGACAAAGAGAAACACTG-3′ and reverse,
5′-ATGAAGACTTGGGTAAAAATCG-3′; GAPDH forward,
5′-CGAAGTCAACGGATTTGGTCGTAT-3′ and reverse
5′-AGCCTTCTCGGTGGTGAAGAC-3′.
For miR-204 detection, 1 µg total RNA extracted from
clinical samples was converted to cDNA using the TaqMan MicroRNA
Reverse Transcription kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
resulting cDNA was diluted at a ratio of 1:40 and mixed with 1 µl
miR-204 or U6 TaqMan primers in triplicate wells using TaqMan
Universal Master Mix II without Uracil DNA glycosylase (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The primer sequences
were as follows: miR-204 forward, 5′-CTGTCACTCGAGCTGCTGGAATG-3′ and
reverse, 5′-ACCGTGTCGTGGAGTCGGCAATT-3′; U6 forward,
5′-GCTTCGGCAGCACATATACTAAAAT-3′, reverse
5′-CGCTTCACGAATTTGCGTGTCAT-3′. The thermocycling conditions were as
follows: Denaturation at 95°C for 30 sec, followed by 40 cycles at
95°C for 5 sec and 60°C for 30 sec, and extension at 95°C for 15
sec. The plates were read using the ABI PRISM 7900HT system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). Cq values
were calculated using SDS version 2.4 software (Applied Biosystems;
Thermo Fisher Scientifc, Inc.). miR-204 expression level was
normalized to that of U6 using the 2−ΔΔCq method
(21). The TaqMan probes for miR-204
and U6 were purchased from Applied Biosystems (Thermo Fisher
Scientific, Inc.). The assay was performed in triplicate.
Statistical analysis
Statistical analysis was performed using SAS
statistical software (version 9.4; SAS Institute, Cary, NC). Data
are presented as the mean ± the standard error of the mean.
Student's t test (two-tailed) was used to compare between two
groups. The correlation between the expression levels of miR-204
and FAP-α were analyzed using Spearman correlation analysis
(22). P<0.05 was considered to
indicate a statistically significant difference.
Results
TMZ resistance promotes formation of
CICs and upregulates FAP-α expression in glioblastoma
To identify U251MG-R cells that were resistant to
TMZ, an MTT assay using TMZ was performed using the U251MG and
U251MG-R cells. The results indicated that the U251MG-R cells were
resistant to TMZ (Fig. 1A). In order
to identify whether TMZ resistance affected CICs traits in U251MG-R
cells, sphere forming assays were performed to assess the capacity
of CIC or CIC-like cell self-renewal in U251MG, and U251MG-R cells.
The sphere forming assay indicated that U251MG-R cells formed
larger spheres after 14 days of culture compared with U251MG cells
(Fig. 1B). STAT3 and MDM2 may promote
TMZ-resistance (22–24). CD133 and MET is essential for
glioblastoma stem cell maintenance (25,26).
Western blotting identified that STAT3, MDM2, CD133 and MET were
markedly upregulated in U251MG-R cells compared with U251MG cells
(Fig. 1C). In order to detect whether
TMZ-resistance is associated with FAP-α protein expression, FAP-α
protein expression was analyzed in U251MG and U251MG-R cells. The
results demonstrated that the level of FAP-α protein was markedly
upregulated in U251MG-R cells (Fig.
1D).
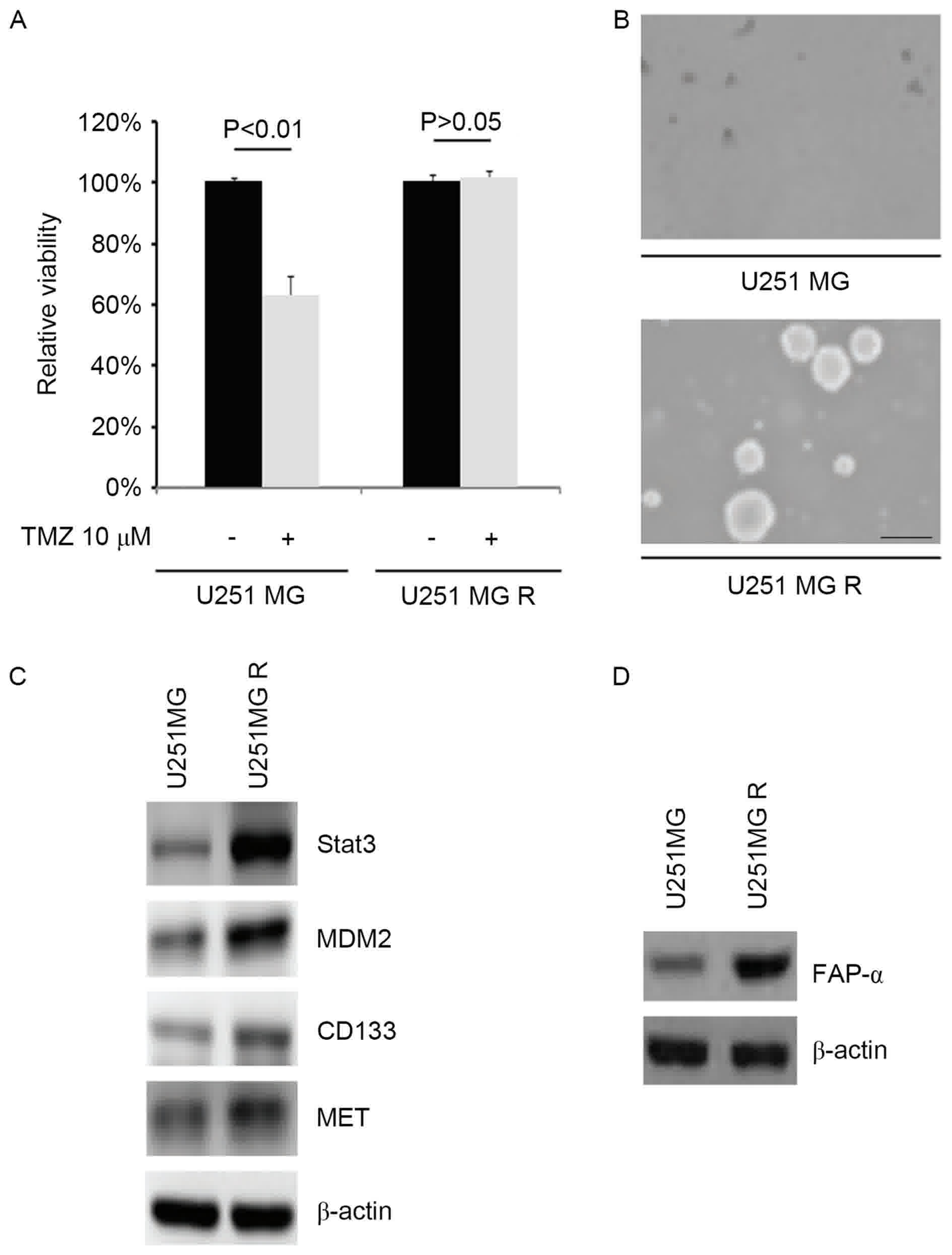 | Figure 1.TMZ resistance promotes the formation
of cancer-initiating cells and upregulates FAP-α expression in
glioblastoma (n=3). (A) MTT for cell viability in U251MG and
U251MG-R cells. Cells were untreated or treated with TMZ. (B)
Sphere growth for U251MG and U251MG-R cells. Scale bar, 200 µm. (C)
Western blotting for STAT3, MDM2, CD133 and MET in U251MG and
U251MG-R cells. β-actin was the loading control. (D) Western
blotting for FAP-α in U251MG and U251MG-R cells. β-actin was the
loading control. R, resistant; TMZ, temozolomide; FAP-α, Fibroblast
activation protein α; STAT3, Signal transducer and activator of
transcription 3; MDM2, Mouse double minute 2 homolog; CD133,
Prominin-1; MET, Tyrosine-protein kinase Met. |
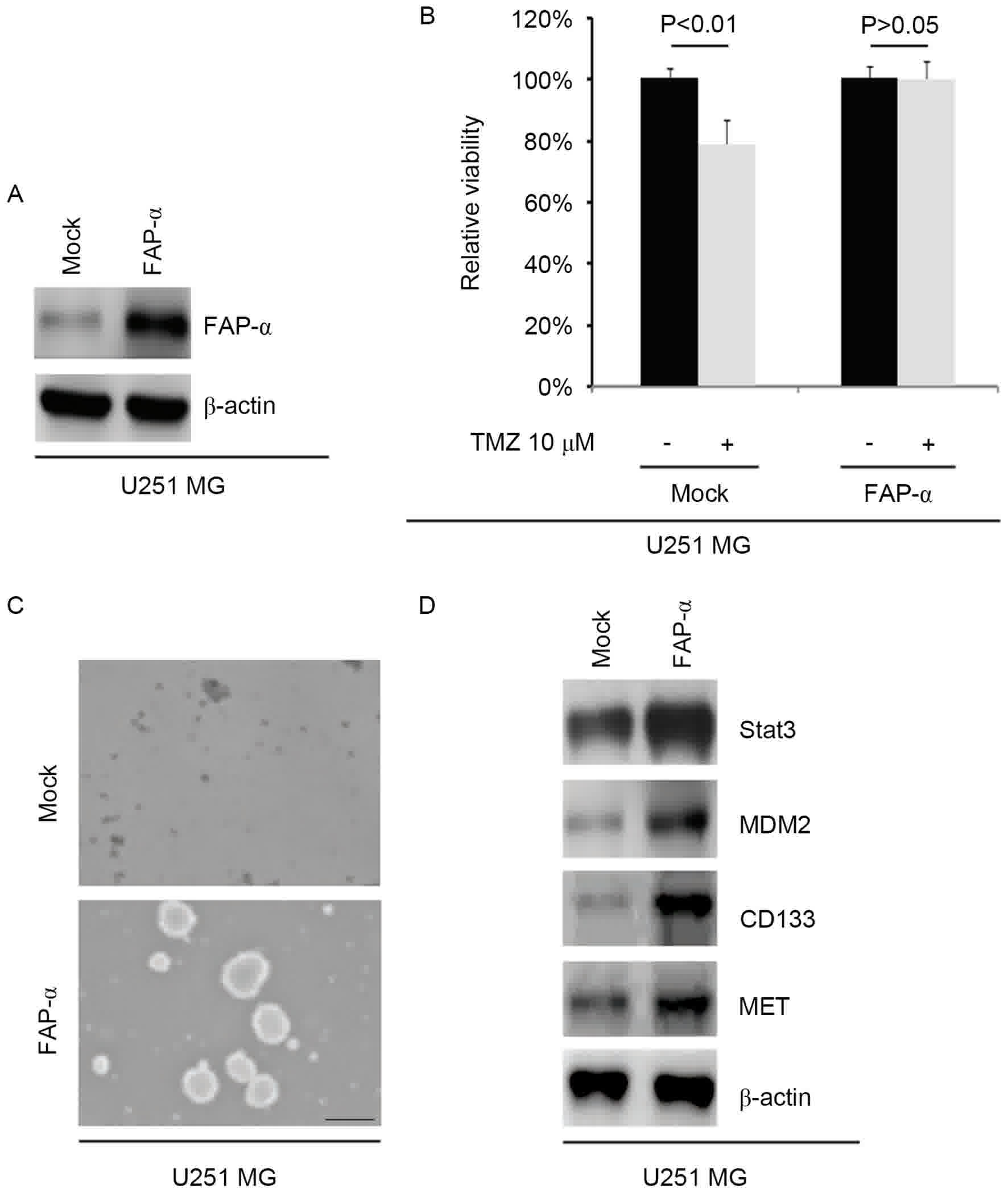
FAP-α promotes TMZ resistance and
induces formation of CICs in U251MG cells
To identify the role of FAP-α, whether FAP-α
expressing plasmids were able to stably express FAP-α protein in
U251MG cells was tested. The results indicated that the levels of
FAP-α protein were markedly increased by FAP-α-expressing plasmids
in the cells (Fig. 2A). To
additionally identify whether FAP-α affected TMZ efficacy in
glioblastoma cells, U251MG cells were transfected with
FAP-α-expressing plasmids. Then an MTT assay was performed in the
cells transfected with FAP-α expressing plasmids. The results
indicated that overexpressing FAP-α transformed U251MG cells into
U251MG-R cells (Fig. 2B), suggesting
that its overexpression promotes TMZ resistance.
In order to identify if FAP-α affected the CIC
traits of U251MG cells, a sphere forming assay was performed to
assess the capacity of CIC or CIC-like cell self-renewal in U251MG
cells. The sphere forming assay demonstrated that FAP-α
overexpressing cells formed larger spheres after 14 days of
culture, thus expressing more CIC-like traits compared with the
mock group (Fig. 2C). To identify
whether FAP-α affects STAT3, MDM2, CD133 and MET protein
expression, western blotting was performed to detect their
expression in U251MG cells transfected with FAP-α expressing
plasmids. All four protein levels were markedly increased following
FAP-α overexpression compared with the mock control group (Fig. 2D).
miR-204 inhibits FAP-α protein
expression in U251MG-R cells
Having demonstrated that FAP-α expression was
specifically upregulated in U251MG-R cells and promoted the
formation of CICs in U251MG cells, the mechanisms that induced
FAP-α expression in U251MG-R cells were then investigated. miRNAs
are a class of small (~22 nucleotide) noncoding RNAs that
negatively regulate protein-coding gene expression by targeted mRNA
degradation or translation inhibition (23). The downregulation of specific miRNA
may contribute to the upregulation of oncogenes and TMZ resistance
(24,25). Therefore, the present study
hypothesized whether FAP-α was upregulated by the downregulation of
specific miRNA in U251MG-R cells.
To confirm this hypothesis, a commonly used
prediction algorithm, miRanda (http://www.microrna.org/), was used to analyze the 3′
untranslated region (UTR) of FAP-α. The algorithm predicted that
miR-204 targeted the 3′UTR of FAP-α (Fig.
3A).
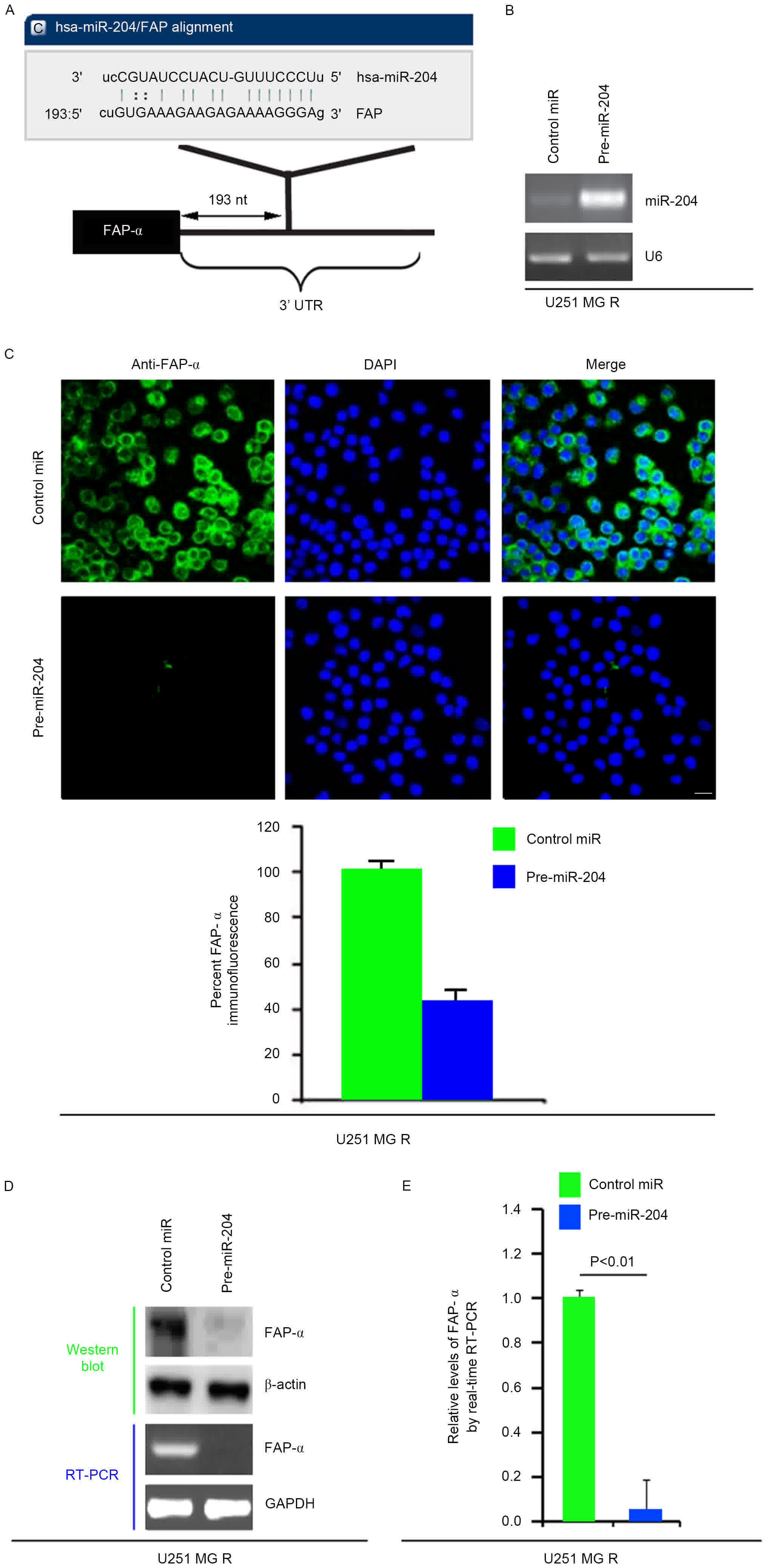 | Figure 3.miR-204 inhibits FAP-α protein
expression in U251MG-R cells. (A) Schematic of predicted miR-204
binding sites in the 3′UTR of FAP-α mRNA by miRanda. (B) qPCR for
miR-204 in U251MG-R cells. U251MG-R cells were infected with
pre-miR-204 or control miR (mock). U6 was the loading control.
(n=3). (C) Immunofluorescence analyses for U251MG-R cells
transfected with pre-miR-204 and control miR (mock). Upper panel
demonstrates microscopy images of immunofluorescence staining of
one representative experiment (magnification, ×100). Bottom panel
indicates graphic representation of mean fluorescence intensities.
(n=3). Scale bar, 20 µm. (D) Western blotting for and RT-PCR for
FAP-α protein and FAP-α mRNA, respectively, in U251MG-R cells
infected as indicated. β-actin and GAPDH were the loading controls
for the western blotting and RT-PCR (n=3). (E) qPCR for FAP-α in
U251MG-R cells transfected with pre-miR-204 or control miR (mock).
GAPDH was the loading control (n=3). UTR, untranslated region; miR,
microRNA; R, resistant; PCR, polymerase chain reaction; RT, reverse
transcription; q, quantitative; FAP-α, Fibroblast activation
protein α. |
Therefore, it was reasoned that miR-204
downregulated FAP-α expression by targeting its 3′UTR in U251MG-R
cells, and that FAP-α was increased in U251MG-R cells due to the
downregulation of miR-204 (26). In
an attempt to identify the role of miR-204 in regulating FAP-α
expression in U251MG-R cells, cells were transfected with
pre-miR-204 and control miR. Following transfection, miR-204
expression was detected by qPCR and the results demonstrated that
miR-204 was markedly increased following pre-miR-204 transfection
in the cells (Fig. 3B). To confirm
this observation, immunofluorescence analyses were performed in
U251MG-R cells transfected with pre-miR-204 or control miR. The
results indicated that FAP-α protein was suppressed in the cells
transfected with pre-miR-204 (Fig.
3C). RT-PCR and western blotting was next performed to detect
FAP-α expression in U251MG-R cells transfected with pre-miR-204 or
control miR. The results demonstrated that FAP-α protein and mRNA
were markedly downregulated in the cells transfected with
pre-miR-204 (Fig. 3D). Consistent
with the results of the RT-PCR, the qPCR data demonstrated that
FAP-α mRNA was significantly reduced in U251MG-R cells transfected
with pre-miR-204 compared with control miR-transfected group
(Fig. 3E). All the data demonstrated
that miR-204 may suppress FAP-α mRNA and protein expression in
U251MG-R cells.
miR-204 inhibits the formation of CICs
and reverses TMZ resistance in U251MG-R cells
In order to detect whether TMZ resistance is
associated with miR-204 expression, miR-204 expression was analyzed
in U251MG and U251MG-R cells. The results suggested that miR-204
expression was markedly downregulated in U251MG-R cells (Fig. 4A). To identify the role of miR-204 in
CIC formation, the ability of pre-miR-204 to stably express miR-204
in U251MG-R cells was examined. The results indicated that the
levels of miR-204 were increased by pre-miR-204 in the cells
(Fig. 4B). To additionally identify
whether miR-204 affected the formation of CICs in U251MG-R cells,
U251MG-R cells were transfected with pre-miR-204 and then a sphere
formation assay was performed. The results demonstrated that
miR-204 inhibited the formation of CICs in U251MG-R cells (Fig. 4C). To additionally identify whether
miR-204 was able to affect TMZ efficacy in U251MG-R cells, U251MG-R
cells were transfected with pre-miR-204. Then, an MTT assay in
U251MG-R cells transfected with pre-miR-204 was performed. The
results demonstrated that miR-204 was able to transform U251MG-R to
U251MG cells (Fig. 4D), suggesting
that miR-204 restoration reversed TMZ resistance. To identify
whether miR-137 affected STAT3, MDM2, CD133 and MET expression,
western blotting was performed to detect their expression in Huh7-R
cells. The results suggested that STAT3, MDM2, CD133 and MET
expression levels were attenuated by miR-204 (Fig. 4E).
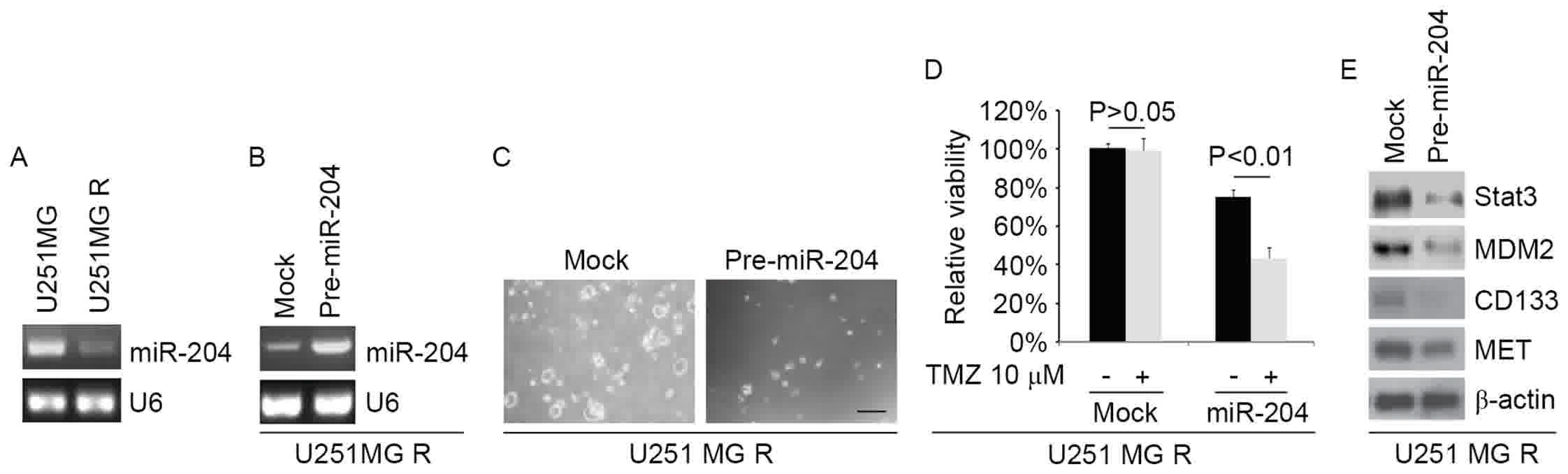 | Figure 4.miR-204 inhibits formation of
cancer-initiating cells and reverses TMZ resistance in U251MG-R
cells. (A) qPCR for miR-204 in U251MG and U251MG-R cells. U6 was
the loading control (n=3). (B) qPCR for miR-204 in U251MG-R cells
transfected with pre-miR-204 and control miR (mock). β-actin was
the loading control (n=3). (C) Sphere growth for U251MG-R cells
transfected with pre-miR-204 and control miR (mock). Scale bar, 200
µm (n=3). (D) MTT assay for cell viability in U251MG-R cells.
U251MG-R cells transfected with pre-miR-204 and control miR (mock)
were untreated or treated with TMZ. (E) Western blotting for STAT3,
MDM2, CD133 and MET in U251MG-R cells transfected with pre-miR-204
and control miR (mock). β-actin was the loading control (n=3). TMZ,
temozolomide; STAT3, Signal transducer and activator of
transcription 3; MDM2, Mouse double minute 2 homolog; CD133,
Prominin-1; MET, Tyrosine-protein kinase Met; miR, microRNA; R,
resistant; PCR, polymerase chain reaction; RT, reverse
transcription; q, quantitative. |
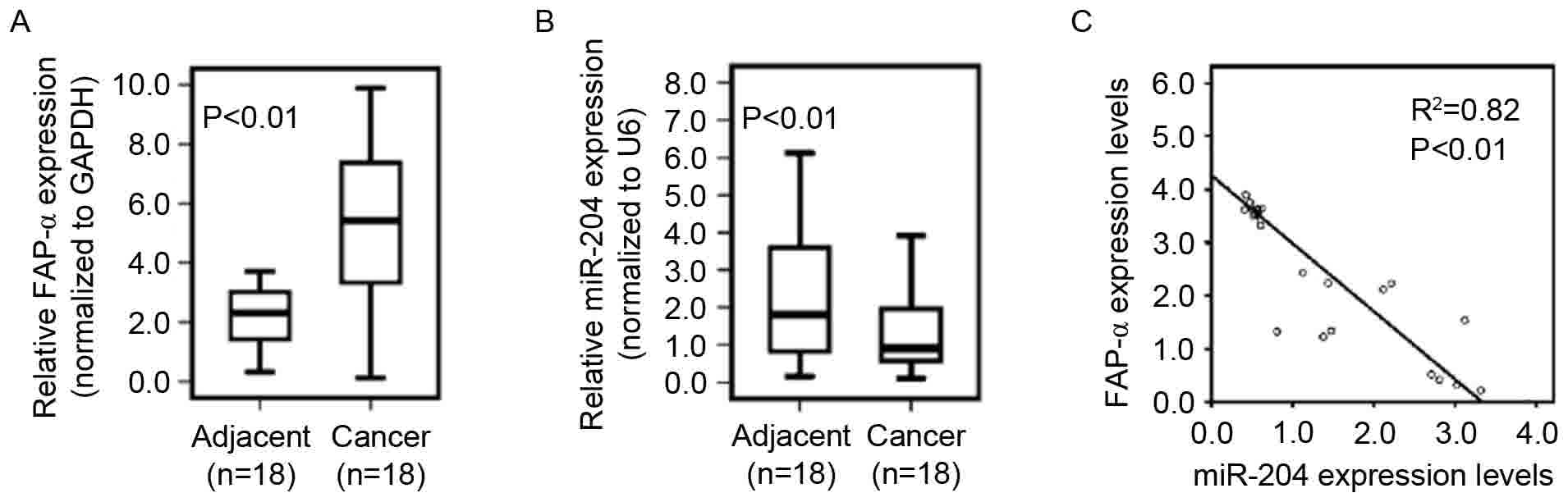
miR-204 expression is negatively
associated with FAP-α levels in human glioblastoma tissues
To determine whether reduced miR-204 expression
correlates with increased levels of FAP-α in glioblastoma tissues,
18 pairs of primary glioblastoma tissues and adjacent normal
tissues were used to determine FAP-α, and miR-204 expression using
qPCR. The results indicated that the FAP-α mRNA level in
glioblastoma tissues was significantly increased compared with that
of adjacent normal tissues (Fig. 5A)
and that the miR-204 level in glioblastoma tissues was
significantly decreased compared with that in adjacent normal
tissues (Fig. 5B). As demonstrated in
Fig. 5C, linear correlation analysis
suggested a significant inverse correlation between miR-204 and
FAP-α mRNA expression in glioblastoma tissues (P<0.01).
miR-204 inhibits RAB22A, EZRIN, SOX4,
ATF2 and EphB protein in U251MG-R cells
In order to detect whether miR-204 inhibits RAB22A,
EZRIN, SOX4, ATF2 and EphB protein expression, U251MG-R cells were
transfected with pre-miR-204. Then, western blotting was performed
to detect the protein levels. The results demonstrated that the
levels of RAB22A, EZRIN, SOX4, ATF2 and EphB proteins were
downregulated in U251MG-R cells transfected with pre-miR-204
(Fig. 6A).
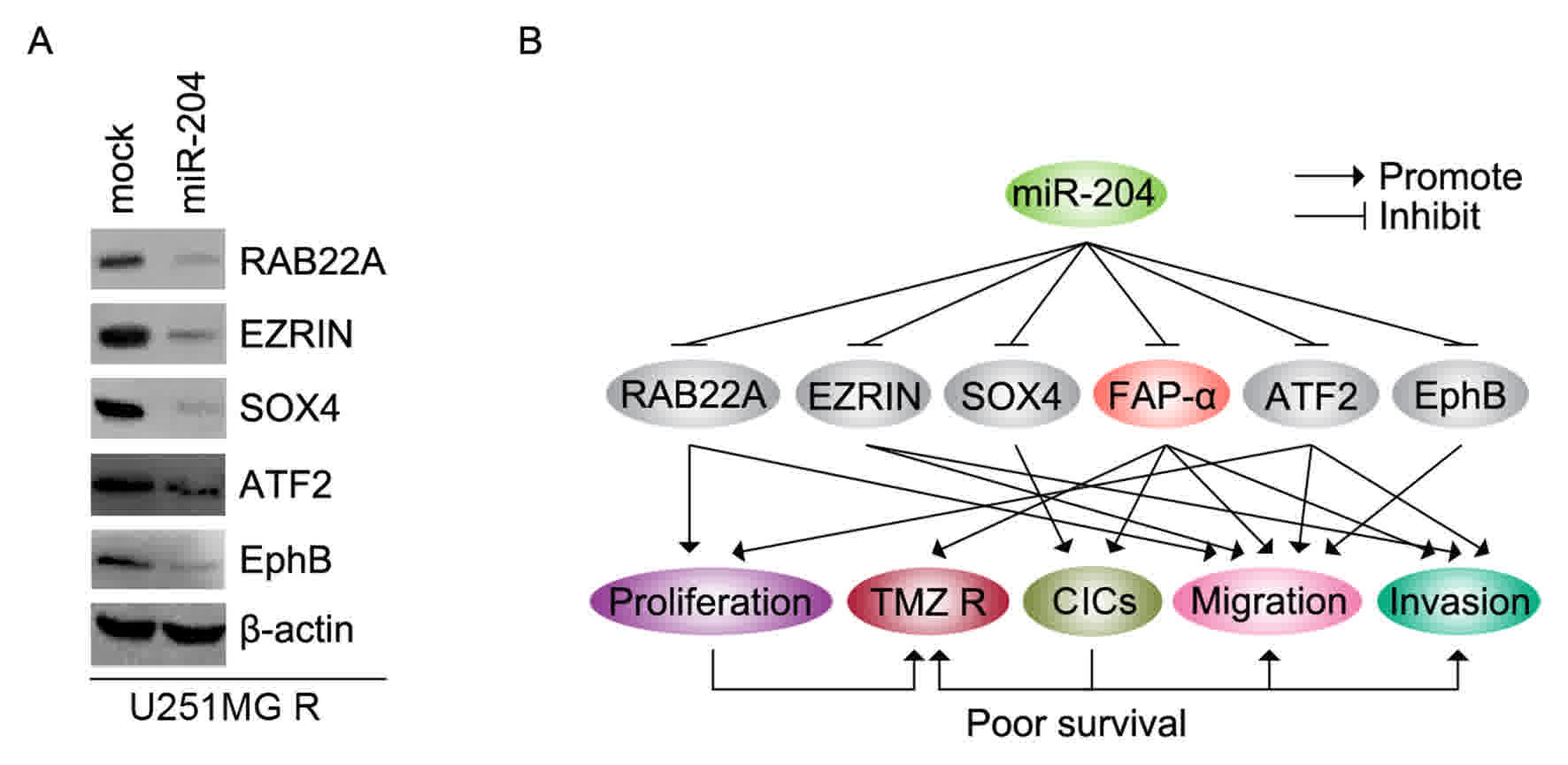 | Figure 6.miR-204 inhibits RAB22A, EZRIN, SOX4,
ATF2 and EphB protein in U251MG-R cells. (A) Western blotting for
RAB22A, EZRIN, SOX4, ATF2 and EphB protein in U251MG-R cells
transfected with pre-miR-204 and control miR (mock). (B) miR-204
regulates proliferation, TMZ resistance, CICs, migration and
invasion by targeting RAB22A, EZRIN, SOX4, ATF2 and EphB protein in
glioblastoma. miR, microRNA; R, resistant; TMZ, temozolomide;
RAB22A, Ras-related protein Rab-22A; SOX4, SRY-box 4; ATF2,
Activating transcription factor 2; EphB, erythropoietin-producing
human hepatocellular receptors. |
Discussion
TMZ has improved the prognosis of patients with GBM
(27,28); however, almost all patients succumb to
recurrence following chemoradiotherapy. miRNAs are involved in the
acquisition of TMZ resistance in GBM (29). The expression level of miR-204 was
significantly reduced in clinical glioma tissues compared with
normal brain tissues (20). The
introduction of miR-204 dramatically suppressed glioma cell growth,
migration and invasion by targeting RAB22A (a member of the RAS
oncogene family) (20), and miR-204
may suppress cell proliferation, migration and invasion through
inhibiting ATF2 and EZRIN expression (18,19).
miR-204 suppresses self-renewal, the stem cell-associated phenotype
of glioma cells by targeting the stemness-governing transcriptional
factor SOX4, and the migration-promoting receptor EphB2 (30). The results of the present study
confirmed that RAB22A, EZRIN, SOX4, ATF2 and EphB were inhibited by
miR-204 (Fig. 6A). Consistent with a
previous study (30), the present
study demonstrated that miR-204 significantly inhibited the
formation of glioma initiating cells (GICs) in glioblastoma, and
that SOX4 and EphB2 may be significantly downregulated by miR-204.
GICs, also termed glioma stem cells, are responsible for tumor
initiation, relapse and therapeutic resistance (26). The results of the present study
demonstrated that the formation of CICs induced by the decrease in
miR-204 levels may serve an important role in promoting TMZ
resistance during the progression of glioblastoma.
FAP-α is highly expressed on the surface of glioma
cells and contributes to diffuse glioma invasion through
extracellular matrix components (11). However, its role in regulating the
formation of GICs and TMZ resistance has not been identified at
present. The present study demonstrated that it serves a crucial
role in regulating TMZ resistance and GIC formation. In addition,
consistent with a previous study, it was identified that FAP-α may
promote proliferation, migration and invasion in glioma cells
(11). The results suggest that FAP-α
may serve an important role in the prognosis of glioblastoma and is
a potential target for therapy.
In conclusion, the present study demonstrated that
miR-204 serves a crucial role in the extensive network of cellular
pathways underlying glioblastoma carcinogenesis, and identified a
novel target gene, FAP-α. Thus, the restoration of miR-204's levels
to physiological levels observed in non-tumor tissues may represent
a novel therapeutic strategy for glioblastoma.
Acknowledgements
Not applicable.
Funding
The present study was supported by Beijing Chaoyang
Hospital, Capital Medical University (Beijing, China).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YNY and ZG performed the majority of the
experimental work, initially conceived the study and wrote a draft
of the manuscript. XHZ, YMW and XZ performed the remainder of the
experimental work and helped during the preparation of the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the ethics
committee of Beijing Chaoyang Hospital, and each patient signed an
informed consent form at the time of enrollment.
Consent for publication
Consent for publication was obtained from each
patient.
Competing interests
All authors declare that there are no competing
interests.
References
|
1
|
Schwartzbaum JA, Fisher JL, Aldape KD and
Wrensch M: Epidemiology and molecular pathology of glioma. Nat Clin
Pract Neurol. 2:494–503. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ostrom QT, Gittleman H, Farah P, Ondracek
A, Chen Y, Wolinsky Y, Stroup NE, Kruchko C and Barnholtz-Sloan JS:
CBTRUS statistical report: Primary brain and central nervous system
tumors diagnosed in the United States in 2006–2010. Neuro Oncol. 15
Suppl 2:ii1–ii56. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Buckner JC: Factors influencing survival
in high-grade gliomas. Seminars in oncology Elsevier. 10–14. 2003.
View Article : Google Scholar
|
|
4
|
Curran WJ Jr, Scott CB, Horton J, Nelson
JS, Weinstein AS, Fischbach AJ, Chang CH, Rotman M, Asbell SO,
Krisch RE, et al: Recursive partitioning analysis of prognostic
factors in three Radiation Therapy Oncology Group malignant glioma
trials. J Natl Cancer Inst. 85:704–710. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
DeAngelis LM: Brain tumors. N Engl J Med.
344:114–123. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fine HA: The basis for current treatment
recommendations for malignant gliomas. J Neurooncol. 20:111–120.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen WT and Kelly T: Seprase complexes in
cellular invasiveness. Cancer Metastasis Rev. 22:259–269. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rosenblum JS and Kozarich JW: Prolyl
peptidases: A serine protease subfamily with high potential for
drug discovery. Curr Opin Chem Biol. 7:496–504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liao XH, Lu DL, Wang N, Liu LY, Wang Y, Li
YQ, Yan TB, Sun XG, Hu P and Zhang TC: Estrogen receptor α mediates
proliferation of breast cancer MCF-7 cells via a
p21/PCNA/E2F1-dependent pathway. FEBS J. 281:927–942. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jia J, Martin TA, Ye L and Jiang WG: FAP-α
(Fibroblast activation protein-α) is involved in the control of
human breast cancer cell line growth and motility via the FAK
pathway. BMC Cell Biol. 15:162014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mentlein R, Hattermann K, Hemion C,
Jungbluth AA and Held-Feindt J: Expression and role of the cell
surface protease seprase/fibroblast activation protein-α (FAP-α) in
astroglial tumors. Biol Chem. 392:199–207. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yoshikawa K, Noguchi K, Nakano Y, Yamamura
M, Takaoka K, Hashimoto-Tamaoki T and Kishimoto H: The Hippo
pathway transcriptional co-activator, YAP, confers resistance to
cisplatin in human oral squamous cell carcinoma. Int J Oncol.
46:2364–2370. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee RC, Feinbaum RL and Ambros V: The C.
Elegans heterochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pasquinelli AE, Reinhart BJ, Slack F,
Martindale MQ, Kuroda MI, Maller B, Hayward DC, Ball EE, Degnan B,
Müller P, et al: Conservation of the sequence and temporal
expression of let-7 heterochronic regulatory RNA. Nature.
408:86–89. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Goscinski MA, Suo Z, Flørenes VA,
Vlatkovic L, Nesland JM and Giercksky KE: FAP-alpha and uPA show
different expression patterns in premalignant and malignant
esophageal lesions. Ultrastruct Pathol. 32:89–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang C, Shen F, Du J, Hu Z, Li X, Su J,
Wang X and Huang X: MicroRNA-564 is downregulated in glioblastoma
and inhibited proliferation and invasion of glioblastoma cells by
targeting TGF-β1. Oncotarget. 7:56200–56208. 2016.PubMed/NCBI
|
|
17
|
Gao YT, Chen XB and Liu HL: Up-regulation
of miR-370-3p restores glioblastoma multiforme sensitivity to
temozolomide by influencing MGMT expression. Sci Rep. 6:329722016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Song S, Fajol A, Tu X, Ren B and Shi S:
miR-204 suppresses the development and progression of human
glioblastoma by targeting ATF2. Oncotarget. 7:70058–70065. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mao J, Zhang M, Zhong M, Zhang Y and Lv K:
MicroRNA-204, a direct negative regulator of ezrin gene expression,
inhibits glioma cell migration and invasion. Mol Cell Biochem.
396:117–128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xia Z, Liu F, Zhang J and Liu L: Decreased
expression of MiRNA-204-5p contributes to glioma progression and
promotes glioma cell growth, migration and invasion. PLoS One.
10:e01323992015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lu DL, Sookthai D, Le Cornet C, Katzke VA,
Johnson TS, Kaaks R and Fortner RT: Reproducibility of serum
oxysterols and lanosterol among postmenopausal women: Results from
EPIC-Heidelberg. Clin Biochem. 52:117–122. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sato A, Sunayama J, Matsuda K, Seino S,
Suzuki K, Watanabe E, Tachibana K, Tomiyama A, Kayama T and
Kitanaka C: MEK-ERK Signaling Dictates DNA-Repair Gene MGMT
expression and temozolomide resistance of stem-like glioblastoma
cells via the MDM2-p53 Axis. Stem Cells. 29:1942–1951. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li P, Lu X, Wang Y, Sun L, Qian C, Yan W,
Liu N, You Y and Fu Z: MiR-181b suppresses proliferation of and
reduces chemoresistance to temozolomide in U87 glioma stem cells. J
Biomed Res. 24:436–443. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
She X, Yu Z, Cui Y, Lei Q, Wang Z, Xu G,
Luo Z, Li G and Wu M: miR-181 subunits enhance the chemosensitivity
of temozolomide by Rap1B-mediated cytoskeleton remodeling in
glioblastoma cells. Med Oncol. 31:8922014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Anido J, Sáez-Borderías A, Gonzàlez-Juncà
A, Rodón L, Folch G, Carmona MA, Prieto-Sánchez RM, Barba I,
Martínez-Sáez E, Prudkin L, et al: TGF-β receptor inhibitors target
the CD44(high)/Id1(high) glioma-initiating cell population in human
glioblastoma. Cancer Cell. 18:655–668. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stupp R, Mason WP, Van Den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hegi ME, Diserens AC, Gorlia T, Hamou MF,
de Tribolet N, Weller M, Kros JM, Hainfellner JA, Mason W, Mariani
L, et al: MGMT gene silencing and benefit from temozolomide in
glioblastoma. N Engl J Med. 352:997–1003. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wong ST, Zhang XQ, Zhuang JT, Chan HL, Li
CH and Leung GK: MicroRNA-21 inhibition enhances in vitro
chemosensitivity of temozolomide-resistant glioblastoma cells.
Anticancer Res. 32:2835–2841. 2012.PubMed/NCBI
|
|
30
|
Ying Z, Li Y, Wu J, Zhu X, Yang Y, Tian H,
Li W, Hu B, Cheng SY and Li M: Loss of miR-204 expression enhances
glioma migration and stem cell-like phenotype. Cancer Res.
73:990–999. 2013. View Article : Google Scholar : PubMed/NCBI
|