Introduction
Human cytomegalovirus (HCMV), a member of the
betaherpesvirinae (HHV) sub-family, is a widespread viral agent
that infects 50–90% of people worldwide (1). Following primary infection, in which the
production of infectious viruses is undetectable and viral gene
expression is highly restricted (2),
HCMV establishes a lifelong latency in cells of the myeloid lineage
(3). It may also transmit from mother
to fetus and results in a disease burden that is substantial and
severe, including sensorineural hearing loss (4–6). In
healthy individuals, infection of HCMV is usually asymptomatic, but
it may lead to life-threatening diseases, with multi-organ
involvement and frequent fatalities in patients with deficient
immune systems (7,8). HCMV glycoprotein (gB) is the major
component of the virus envelope and the most highly conserved
glycoprotein of the human HHV family (9). It serves an essential role in the HCMV
entry process by binding to membranous Integrin β-1 via its HCMV gB
disintegrin-like domain, which mediates virus entry to the host
cell and cell-cell virus transmission (10).
Breast cancer is considered a leading cause of
cancer-associated mortality and the most common malignancy among US
women in 2013 (11). Although it is
clear that age, estrogen level, family history and factors
associated with lifestyle and diet are prominent risk factors for
the onset of breast cancer (12),
studies suggest that viruses maybe an additional high-risk factor
closely associated with human breast cancer (13). For example, infection with human
papillomavirus, Epstein-Barr virus, HCMV and HHV-8 has been
suggested as risk factors or associated with the development of
breast cancer (14). Among these
viruses, HCMV was identified in patients with newly-diagnosed
(15) and metastatic breast cancer
(16). In addition, HCMV proteins and
DNA have been identified in breast ductal carcinoma in situ
and infiltrating ductal carcinoma tissues, suggesting that HCMV
infection may be associated with breast carcinogenesis (17). HCMV gB, the most abundant and highly
antigenic viral envelope protein (9),
serves an important role in host cell entry, cell-cell virus
transmission and fusion of infected cells (18). Previous data have demonstrated that
HCMV gB may promote the growth, migration and infiltration of
glioma by binding and activating platelet-derived growth factor
receptor alpha (PDGFRα) and its downstream signaling pathways;
therefore, targeting HCMV gB may have therapeutic benefits for
patients with HCMV-positive tumors (19). However, it is not clear whether HCMV
gB serves roles in the development of breast cancer.
Transforming growth factor-β (TGF-β) is secreted
abundantly by tumors cells, and serves roles within complex
bidirectional interactions in epithelial carcinogenesis (20,21).
During breast cancer progression, TGF-β inhibits cell proliferation
in well-differentiated, early stage breast tumors through the
induction of cell cycle arrest and apoptosis (22–24), while
in poorly differentiated advanced-stage breast tumors, these
functions were replaced by tumor-promoting and pro-metastatic
responses (25). The binding of TGF-β
to its receptors leads to the recruitment and phosphorylation of
Mothers against decapentaplegic homologs 2/3 (Smad2/3), and the
majority of the pro-metastatic functions of TGF-β are attributed to
the TGF-β/Smad signaling pathway (26,27). As
Smad proteins are not ideal drug targets due to their roles as
transcription factors, anti-TGF-β therapies have demonstrated
potential in preventing the development of breast cancer and other
types of cancer, including melanoma (28–30).
In the present study, to clarify the role of HCMV
glycoprotein B (gB) in breast cancer cells, stable HCMV
gB-transfected MDA-MB-231 cells were established, and it was
identified that although HCMV Gb exhibited no effect on MDA-MB-231
cell proliferation and apoptosis, it led to the suppression of cell
migration. In addition, it was demonstrated that HCMV gB inhibited
TGF-β protein expression and Smad2/3 activation in MDA-MB-231
cells. To the best of our knowledge, these data are the first to
demonstrate that HCMV gB suppresses breast cancer cell migration
and inhibits TGF-β/Smad signaling; this may assist in developing
novel anticancer agents that contribute to tumor suppression.
Materials and methods
Materials
The human MDA-MB-231 cell line was purchased from
the Cell Bank of Type Culture Collection of Chinese Academy of
Sciences (Shanghai, China) and was cultured in Dulbecco's modified
Eagle's medium (DMEM; Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum
(Invitrogen; Thermo Fisher Scientific, Inc.) and maintained in a
humidified atmosphere containing 5% CO2-humidified
atmosphere at 37°C. Western blot assays were performed using the
following primary antibodies: Rabbit anti-human caspase-3 (cat. no.
14220; dilution, 1:1,000); rabbit anti-human caspase-9 (cat. no.
9508; dilution, 1:1,000); rabbit anti-human B-cell lymphoma 2
(Bcl-2)-associated X protein (Bax; cat. no. 2772; dilution,
1:1,000); rabbit anti-human TGF-β (cat. no. 3711; dilution,
1:1,000); rabbit anti-human Smad2/3 (Smad2: cat. no. ab1305; Smad3:
ab28379; dilution, 1:1,000); rabbit anti-human Bcl-2 (cat. no.
3498; dilution, 1:1,000); and mouse anti-actin (all from EMD
Millipore, Billerica, MA, USA; cat. no. MAB1501; dilution
1:10,000). The secondary antibody HRP-conjugated antibodies
[HRP-anti-rabbit antibody (cat. no. 7074; dilution, 1:10,000) and
HRP-anti-mouse antibody (cat. no. 7076; dilution, 1:10,000)] were
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
The BCA protein assay kit was sourced from Beyotime Institute of
Biotechnology (Haimen, China), the Cell Counting Kit-8 (CCK-8) from
Dojindo Molecular Technologies, Inc. (Kumamoto, Japan) and the
apoptosis detection kit from BD Pharmingen (BD Biosciences,
Franklin Lakes, NJ, USA). The 0.25% Trypsin-EDTA (1X) and fetal
bovine serum (FBS) were purchased from Gibco (Thermo Fisher
Scientific, Inc.), and the Transwell inserts were purchased from
Corning Incorporated (Corning, NY, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.). An equal amount of
total RNA was used for first-strand cDNA synthesis using the
oligo-dT primer and M-myeloblastosis virus reverse transcriptase XL
(Promega Corporation, Madison WI, USA). The synthesized
first-strand of cDNA (2 µl) was used for each PCR. The target
fragment and primer (UL55 forward, CCGCTCGAGGCCACCATGGAAT and
reverse, CGGGATCCGACGTTCTCTTCTTC) were designed and synthesized in
Sangon Biotech Co., Ltd. (Shanghai, China), and the qPCR analysis
was performed using the GoTaq qPCR Master Mix (Promega
Corporation), and GAPDH was used as an internal reference control.
PCR was performed at 95°C for 10 min, followed by 40 cycles of 95°C
for 15 sec and 55°C for 1 min. Each reaction was performed in
triplicate. Then, the product was ligated into the lentiviral
vector PHY-022. The fast growing bacteria DH5α were treated with
0.1 M CaCl2 solution at 0°C for 20 min to obtain the
receptive bacteria. The ligation product was transferred into the
prepared receptive bacteria DH5α, and the positive clones were
identified by qPCR. The correct clones were those containing the
lentiviral vector required (Fig.
1A).
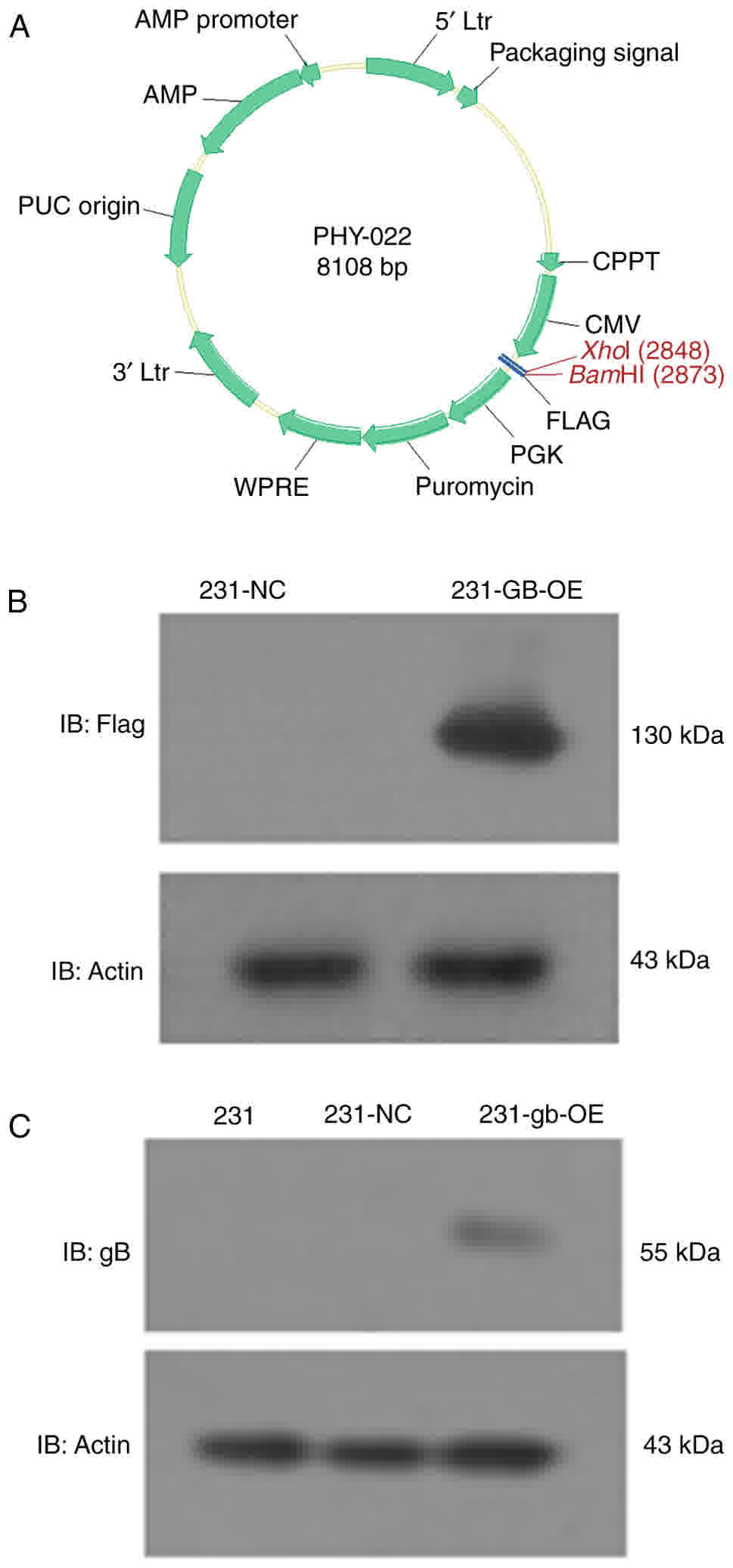 | Figure 1.HCMV gB is successfully transfected
into MDA-MB-231 cells. (A) Lentiviral vector structure. (B) Flag
was expressed in 231-GE-OE cells. (C) Normal MDA-MB-231 cells (231)
and the empty vector transfected cells (231-NC) did not express
HCMV gB, while HCMV gB-transfected MDA-MB-231 cells (231-GB-OE)
demonstrated gB expression; IB, immunoblotting; NC, negative
control; AMP, Adenosine 5′-monophosphate; Ltr, long terminal
repeat; WPRE, Woodchuck Hepatitis Virus posttranscriptional
regulatory element; CPPT, central polypurine tract; CMV,
cytomegalovirus; HCMV, human CMV; PGK, phosphoglycerate kinase
promotor plasmid; PUC, origin of replication; gB, glycoprotein
B. |
Lentiviral vector construction and
stable transfection of MDA-MB-231 cells
Lentiviral vector for gB (UL55) expression sequence
were constructed by Hanyin Co (http://www.hanyinbt.com). (Shanghai, China). Prior to
construction, a flag (GACTACAAGGACGATGACGACAAGTGA) used as a lable
of expression was added to the end of the UL55 sequence for
tracking effect. The recombinant lentivirus and the negative
control (NC) lentivirus (Hanyin Co.) were prepared and titered to
109 TU/ml (transfection unit). To obtain the stable cell
line, 2×104 MDA-MB-231 cells were seeded in 6-well
plates and incubated overnight in DMEM supplemented with 10% fetal
bovine serum (Invitrogen; Thermo Fisher Scientific, Inc.) and
maintained in a humidified atmosphere with 5% carbon
dioxide-humidified atmosphere at 37°C. The cells exhibited 30–50%
confluence at the time of transduction. The recombinant and
negative control lentivirus particles were dissolved on ice, the
DMEM was removed from the cells, and then the recombinant and
negative control lentivirus/polybrene mixture medium [20 µl of
lentiviral particles and 8 µg/ml polybrene (Sigma Aldrich; Merck
KgaA, Darmstadt, Germany)] was added and the two group cells [cells
transfected with the recombinant lentivirus (231-GB-OE) and the
negative control (NC) lentivirus (231-NC)] were incubated for 72 h
in a humidified atmosphere with 5% CO2-humidified
atmosphere at 37°C. The lentivirus/polybrene mixture medium was
exchanged with fresh DMEM, and the 231-GB-OE and 231-NC cells were
incubated for at least 5 days at 37°C. Subsequently, 231-GB-OE and
231-NC cells were collected and lysed after 5 days, and HCMV gB
expression was detected by western blot analysis of the flag added
in the end of UL55 sequence. For the selection of viable cells, 2
µg/ml puromycin (Sigma Aldrich; Merck KGaA) was added to the
culture medium on day 3.
CCK-8
To test cytotoxicity, MDA-MB-231 cells with
recombinant lentivirus (231-GB-OE) and the negative control
(231-NC) cells (3,000 cells/well) suspension was prepared. A total
of 100 µl cell suspension was dispensed into a 96-well plate. The
plate was pre-incubated in the humidified incubator at 37°C and 5%
CO2. After 24, 48, 72, 96 and 120 h, 10 µl CCK-8
solution was added to each well of the plate consecutively, and the
plate was incubated at 37°C and 5% CO2 for an additional
2 h. The absorbance was measured at 450 nm using a microplate
reader.
Transwell migration assay
The 231-GB-OE and 231-NC cells were cultured in
serum-free DMEM in the humidified incubator at 37°C and 5%
CO2 for 12 h. After 12 h serum starvation, the medium
was removed from the serum-free culture and cells were rinsed with
PBS 3 times, harvested and the cell concentration was diluted to
1×105/ml with DMEM. A total of 100 µl cells were seeded
in the upper chamber, and 600 µl medium with 10% FBS was added to
the lower chamber. The cells were incubated at 37°C and 5%
CO2 for 48 h. The medium was removed from the upper
section, and the cells that had migrated through the membrane were
fixed with 4% paraformaldehyde for 15 min and stained with 0.1%
crystal violet for 30 min at room temperature, then rinsed 3 times
with PBS to remove the excess crystal violet. A total of 6 fields
of view were randomly selected under the light microscope (×100
magnification), then the number of cells was counted and the
average calculated.
Cell scratch assay
The HCMV gB-transfected cells (231-GB-OE) and
231-negative control (NC) cells were seeded onto 6-well tissue
culture plates in a humidified atmosphere containing 5%
CO2 at 37°C at a density of 1×105/ml that
after 24 h of growth they reached 70–80% confluence as a monolayer.
The monolayer was gently and slowly scratched with a new 1 ml
pipette tip across the center of each well. While scratching across
the surface of the well, the long axial of the tip was
perpendicular to the bottom of the well; the resulting distance of
the gap was therefore equal to the outer diameter of the end of the
tip. The well was gently washed 3 times with PBS to remove the
detached cells. DMEM containing 2% FBS was added to the wells, and
the cells were grown for an additional 24 h, following which images
were captured by a camera under the light microscope (×10
magnification).
Cell apoptosis analysis
The 231-GB-OE and 231-NC cells were cultured in DMEM
supplemented with 10% FBS. Cells were seeded (2×104)
onto 6-well tissue culture plates and incubated for 24 h at 37°C
and 5% CO2. The culture medium was removed, washed once
with pre-warmed PBS and digested with 0.25% Trypsin-EDTA. The cells
were collected into a 15-ml tube and centrifuged at 1,000 × g for
15 min at room temperature. The cells were then washed once with
500 µl binding buffer (BD Biosciences), the 200 µl binding buffer
containing 5 µl Annexin V-fluorescein isothiocyanate (FITC; BD
Biosciences) was added to the tube and the cells were stained for
10 min in the dark at room temperature. Subsequently, 5 µl
propidium iodide (PI) was added for an additional 5 min at room
temperature. A total of 300 µl binding buffer was added to the
tube, and cell apoptosis was analyzed using Cell Quest softwell on
a BD FACSAria flow cytometer (BD Biosciences).
Western blot analysis
The cells were lysed with 150 ml/well RIPA buffer
(50 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1% Triton X-100, 0.5%
Na-deoxycholate) containing protease inhibitors (Roche, Complete
Mini) (www.roche.com.cn), and the protein
concentration for each cell lysate was determined through BCA
method (BCA kit; Beyotime Institute of Biotechnology). A total of
20–30 µg protein were loaded into the wells of the 10% SDS-PAGE
gel, following which the proteins were transferred from the gel to
the polyvinylidene fluoride membrane and blocked for 1–2 h at room
temperature with 5% skim milk powder in TBS and 0.05% Tween-20
(TBST). The membrane was incubated with primary antibodies (rabbit
anti-human caspase-3, rabbit anti-human caspase-9, rabbit
anti-human Bax, rabbit anti-human TGF-β), rabbit anti-human
Smad2/3, rabbit anti-human Bcl-2 and mouse anti-actin) overnight at
room temperature, washed three times with TBST and incubated with
conjugated secondary antibody (HRP-anti-rabbit antibody and
HRP-anti-mouse antibody) at room temperature for 2 h. The membrane
was then placed in a cartridge and electrochemiluminescence
reaction solution (cat. no. PI32209; Pierce; Thermo Fisher
Scientific, Inc.) was added for 1–3 min, followed by exposure with
X-ray in the darkroom at room temperature for 1 min. Resulting
bands were analyzed with Image J software 1.4.3.67 (National
Institutes of Health, Bethesda, MD, USA).
Statistical analysis
Data were expressed as the mean ± standard deviation
and statistical differences or comparisons between two groups were
assessed by paired Student's t-test using SPSS for Windows v.17.0
(SPSS, Inc., Chicago, IL, USA). Three repeat experiments were
performed for each protocol. P<0.05 was considered to indicate a
statistically significant difference.
Results
HCMV gB is expressed in MDA-MB-231
cells
The constructed gB overexpression vectors were
sequenced prior to transfection into MDA-MB-231 cells, and no
genetic variation was confirmed within the wild gB gene sequence
(data not shown). Then, the whole gene of HCMV gB was transfected
into MDA-MB-231 cells by the lentiviral vector PHY-022 (Fig. 1A), and the levels of HCMV gB protein
were determined by western blot analysis. Fig. 1B indicates that the UL55-flag which
tagged HCMV gB was stably expressed in the 231-GB-OE cells. In
addition, 231-GE-OE demonstrated gB expression while normal
MDA-MB-231 cells (231) and negative control cells transfected with
empty vectors (231-NC) did not (Fig.
1C).
Effect of HCMV gB on cell
apoptosis
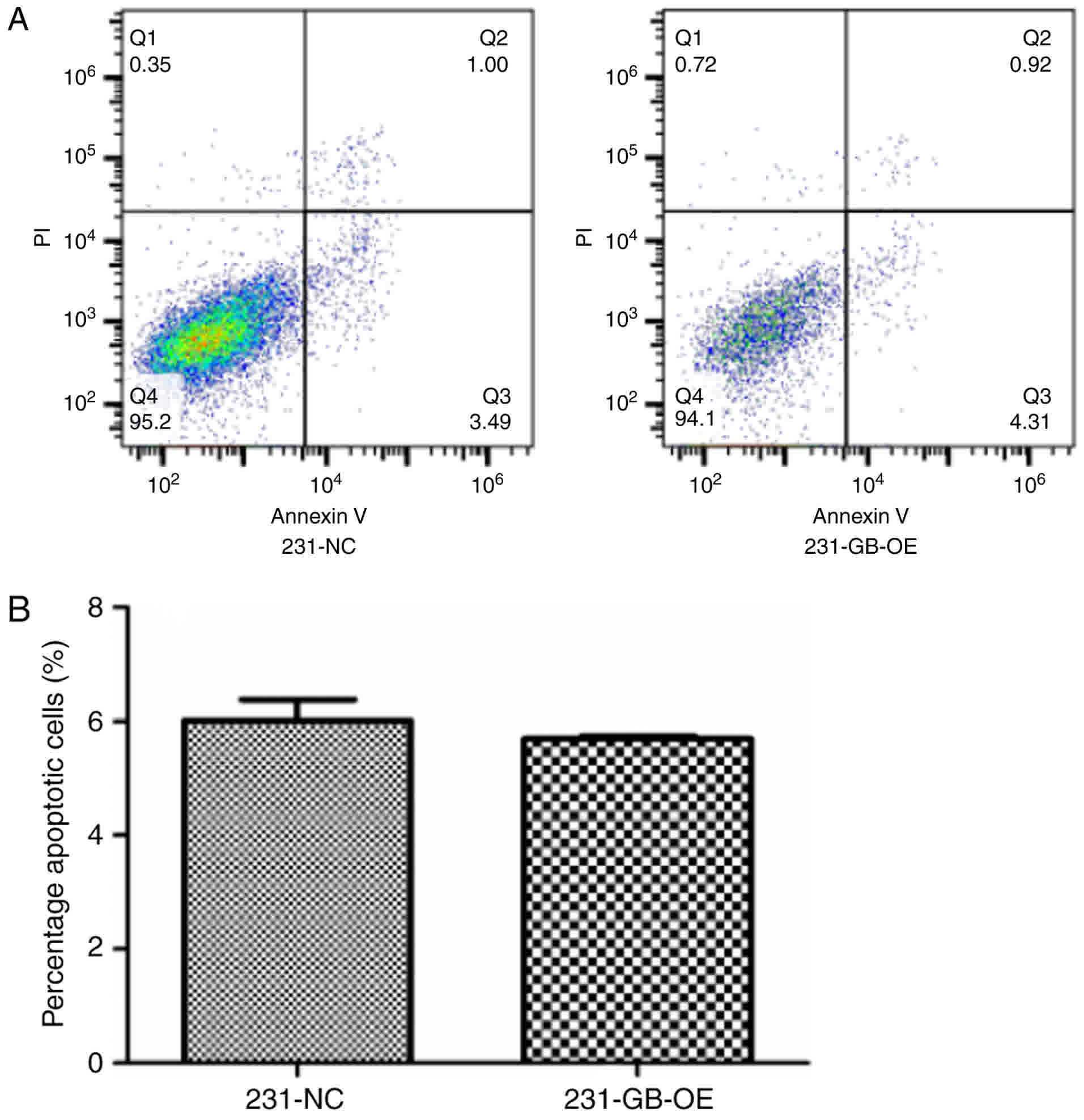
The flow cytometry analysis was used to confirm
whether HCMV gB expression induced apoptosis in MDA-MB-231 cells.
The 231-NC and 231-GB-OE cells were incubated in 6-well plates for
24 h, and as demonstrated in Fig. 2A,
PI Annexin V-FITC double staining revealed that there were no
significant differences between 231-NC and 231-GB-OE cells in the
percentage of apoptotic cells (Fig.
2B). These results indicated that HCMV gB did not induce
apoptotic in the MDA-MB-231 cells.
Effect of HCMV gB on cell
proliferation
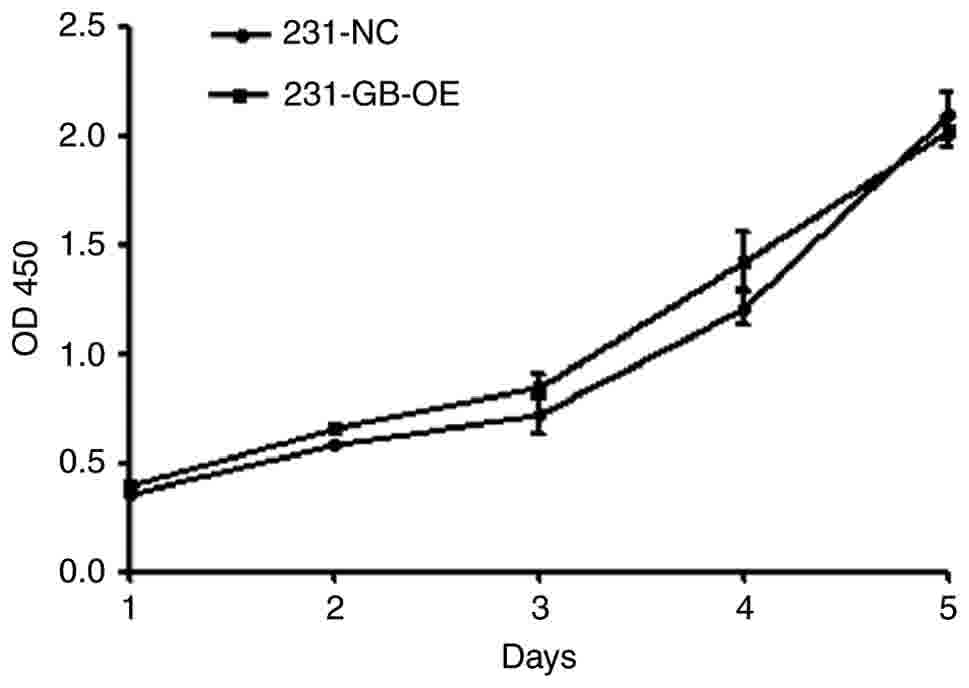
To evaluate whether HCMV gB expression affects
MDA-MB-231 cell proliferation, cell viability was determined by a
CCK-8 cell proliferation assay. The absorbance at 450 nm, which has
been directly associated with cell proliferation (31), indicates that cell proliferation in
the 231-GB-OE cells exhibited no significant difference in
comparison with the 231-NC cells (Fig.
3). This result suggests that HCMV gB has no effect on the
growth of MDA-MB-231 cells.
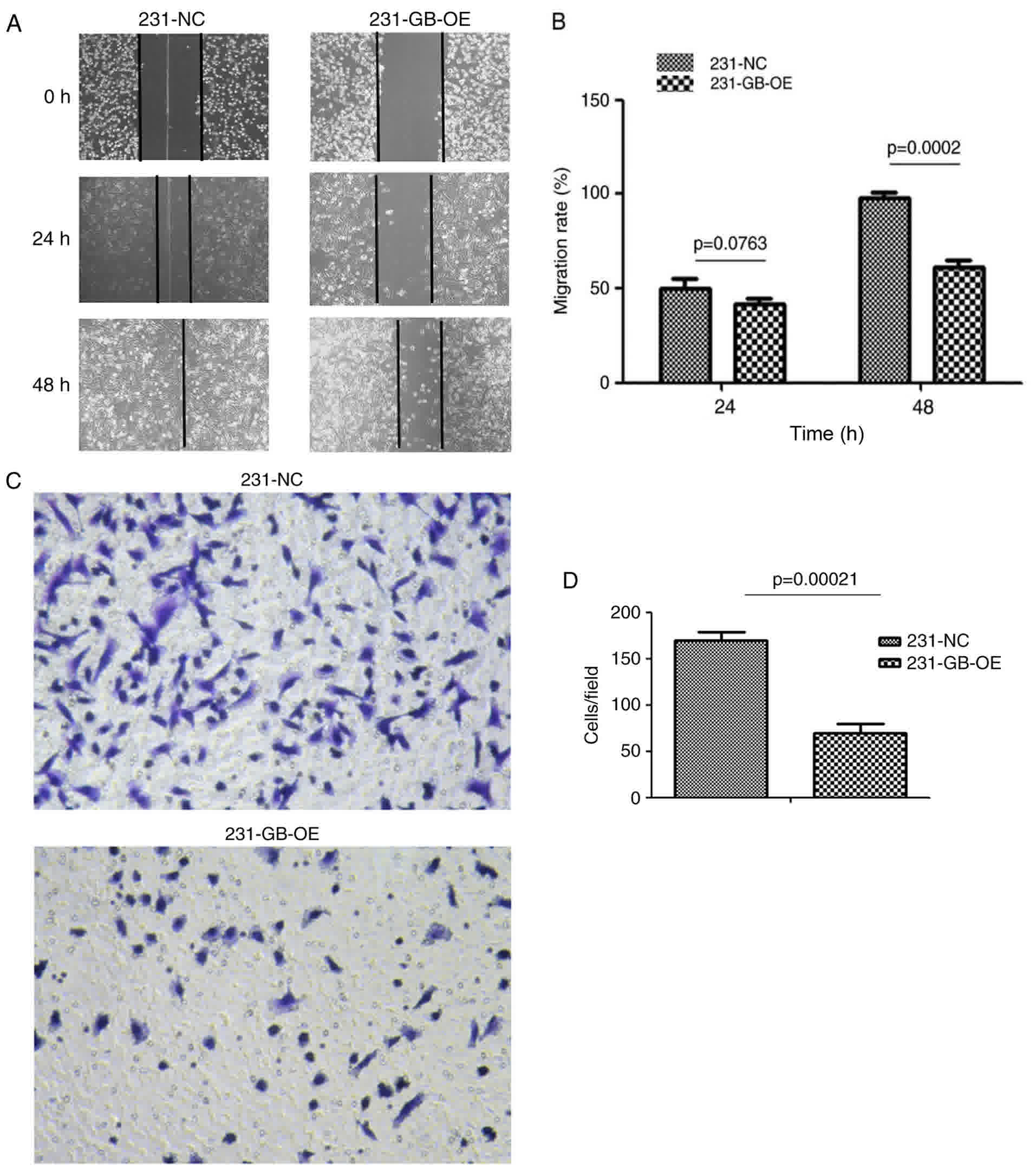
HCMV gB inhibits cell migration
As HCMV gB expression has no effect on cell
apoptosis and proliferation, Transwell and cell scratch assays were
used to determine whether HCMV gB affected tumor migration. The
results indicate that the wound-healing rate in 231-GB-OE cells was
significantly decreased compared with that in the 231-NC cells at
24 and 48 h (Fig. 4A and B). To
additionally validate this observation, cell migration was measured
by a Transwell assay. The migration rates of 231-GB-OE cells to the
lower part of the insert were significantly decreased following 48
h incubation compared with the 231-NC cells (Fig. 4C and D). Collectively, these results
suggest that HCMV gB may inhibit the migration capacity of
MDA-MB-231 cells in vitro.
Effect of HCMV gB on caspase-3,
caspase-9, Bax, Bcl-2 and TGF-β expression
To confirm the aforementioned results that HCMV gB
has no effect on the growth of MDA-MB-231 cells, cell extracts were
obtained from 231-NC and 231-GB-OE cells and the protein levels of
caspase-3, caspase-9, Bax and Bcl-2, which serve important roles in
cell apoptosis, were determined. The results demonstrate that the
levels of these proteins were similar in 231-NC and 231-GB-OE cells
(Fig. 5), which is consistent with
the results from the apoptosis and proliferation assays.
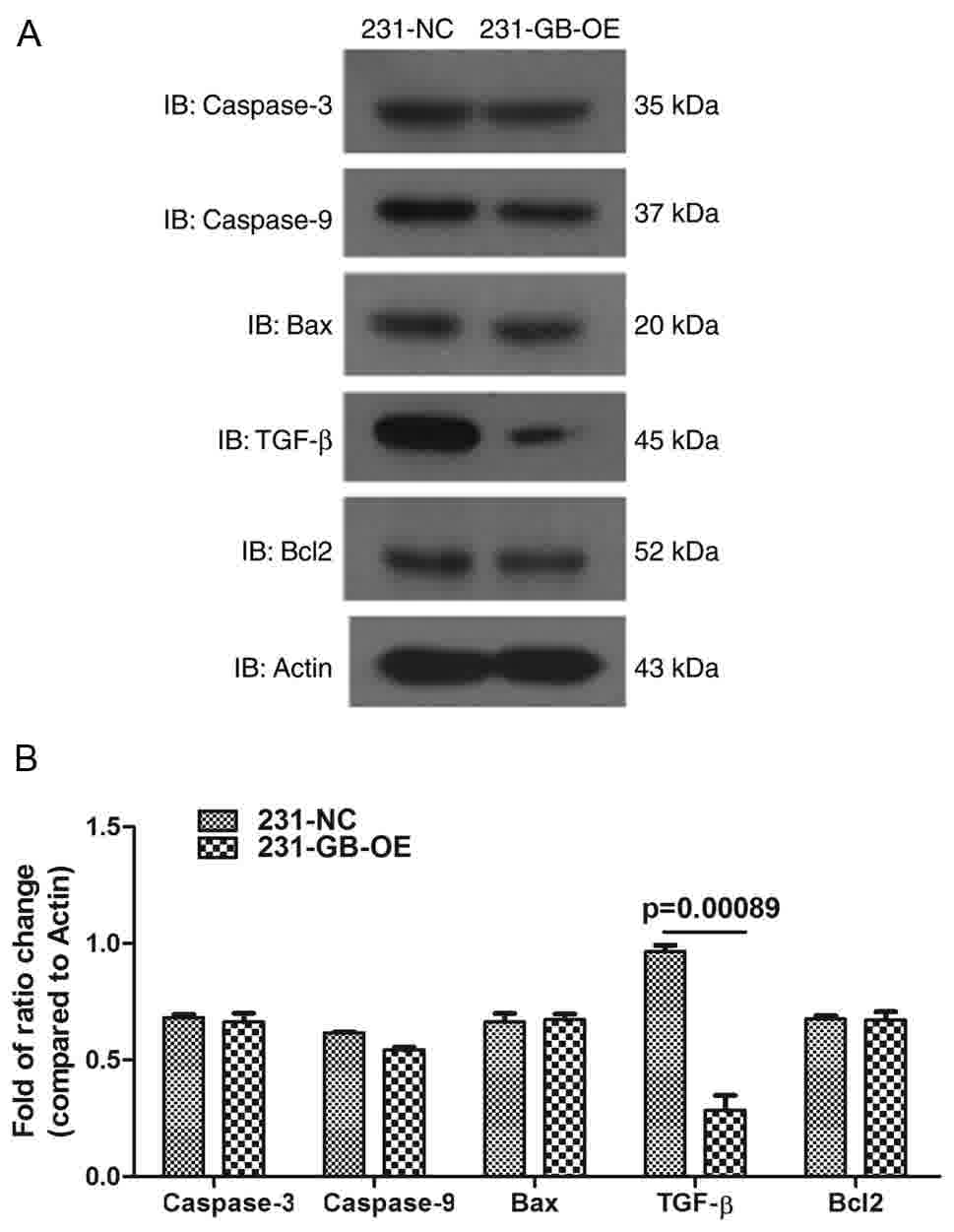 | Figure 5.Effect of HCMV gB on caspase-3,
caspase-9, Bax and TGF-β expression. (A) Apoptosis-associated
protein caspase-3, caspase-9, Bax and Bcl2 were not altered by HCMV
gB transfection, while TGF-β was inhibited by HCMV gB. (B)
Histogram shows the means of three independent experiments (n=3;
mean; SE). Bcl2, B-cell lymphoma 2; Bax, Bcl-2-associated X
protein; TGF-β, transforming growth factor β; HCMV, human
cytomegalovirus; NC, negative control; gB, glycoprotein B: IB,
immunoblotting. |
The molecular mechanisms of how HCMV gB inhibits
MDA-MB-231 migration were then investigated. As TGF-β serves a
prominent role in breast cancer progression and tumor metastasis,
TGF-β protein expression was measured in 231-NC and 231-GB-OE
cells, and the results indicated that TGF-β expression was
significantly inhibited in 231-GB-OE cells compared with the 231-NC
cells (Fig. 5A and B). This suggests
that HCMV gB may suppress TGF-β expression.
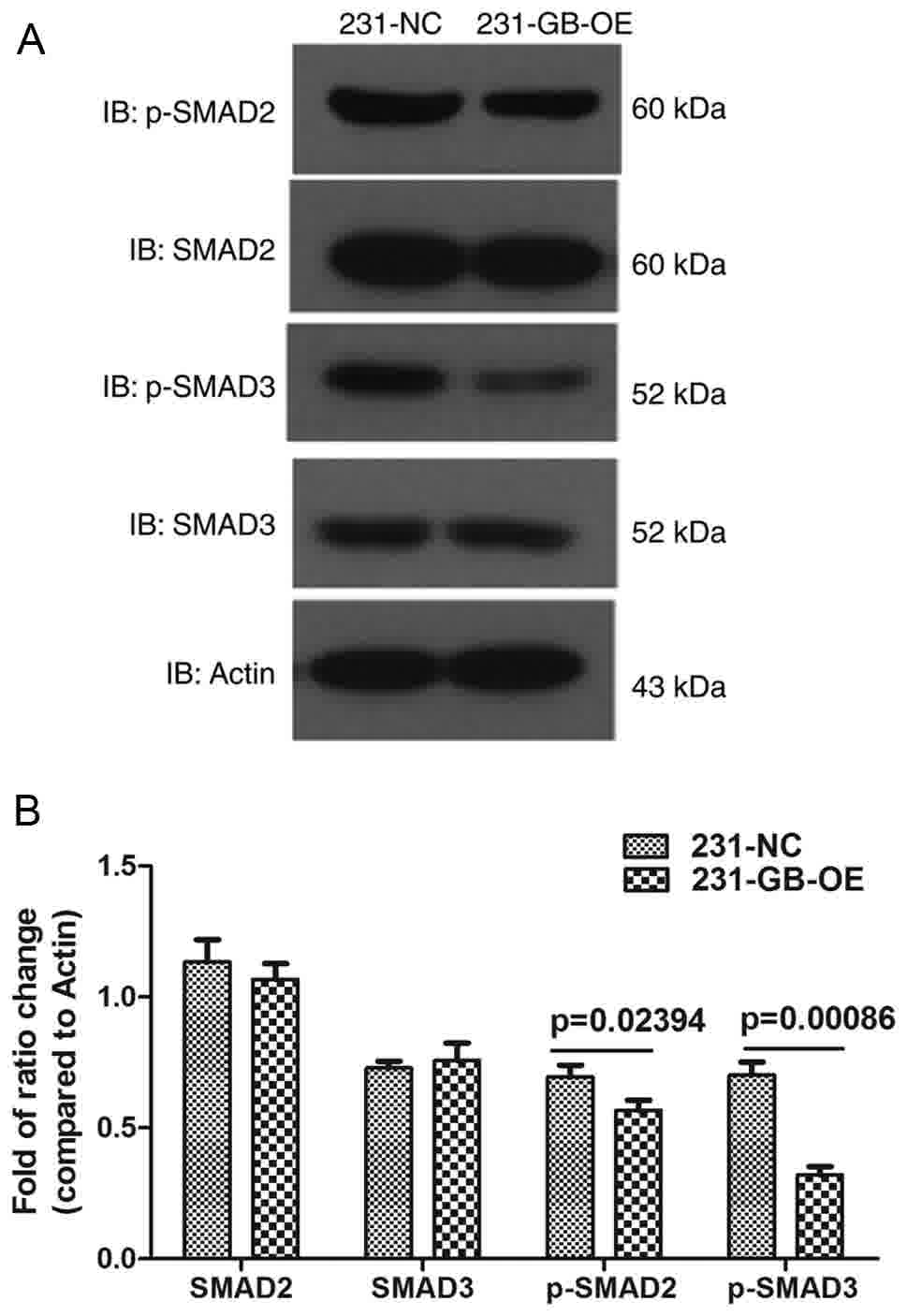
Effect of HCMV gB on Smad2/3
The majority of the biological functions of TGF-β
are attributed to a canonical signaling pathway mediated by Smad
transcription factors (32). To
additionally validate our assumption that the migration may be
inhibited via impaired TGF-β/Smad signaling, the phosphorylation
and total protein levels of Smad2/3 were measured. As expected, the
levels of phosphorylation of Smad2 and Smad3 were decreased in
231-GB-OE cells, compared with the 231-NC cells, while no
difference in total Smad2/3 levels was observed (Fig. 6A and B). These data indicated that
HCMV gB inhibited TGF-β/Smad signaling. Therefore, it was firstly
suggested that the inhibition of migration may be due to impaired
TGF-β/Smad signaling.
Discussion
Although HCMV is not generally regarded as an
oncogenic virus, multiple previous studies have demonstrated that
HCMV infection and expression may be specifically associated with
certain types of human cancer, including glioblastoma, colon cancer
and breast cancer (8,33,34). The
HCMV gB was highly expressed during the entire replication cycle of
the virus, and responsible for activation of HCMV-induced cellular
signaling (35). At present, in order
to investigate the effect of gB in glioma cells, Cobbs et al
(19) introduced HCMV gB into the
retroviral vector and transfected into glioma cells, demonstrating
the functional expression of HCMV gB by gene sequencing and Western
blot analysis: They identified that HCMV gB binds to the
Platelet-derived growth factor subunit A (PDGFa) receptor in glioma
cells, mediates entry of HCMV, induces sustained PDGFRα
phosphorylation and enhances the invasion of primary glioblastoma
cells into Matrigel® and rat brain slices. However, an
additional study indicated that PDGFRα did not serve as a receptor
for HCMV, but increased PDGFRα facilitates virus entry into cells
via a novel entry pathway involving clathrin-independent,
dynamin-dependent endocytosis of HCMV followed by low
pH-independent fusion (36). These
studies were performed in glioma cells, and how HCMV gB entry in
breast cancer cells remains unclear. To identify whether HCMV gB
protein was involved in the effect of HCMV on breast cancer cells,
a HCMV gB-expressing 231-GB-OE cell line was established (Fig. 1). Although HCMV has been detected in
patients with breast cancer (37) and
HCMV gene products was demonstrated to serve in an oncomodulatory
manner through the blockage of cellular differentiation and
induction of chromosomal instability (38), the present study indicated that cell
viability and proliferation were not affected by HCMV gB (Figs. 2 and 3).
Soroceanu and Cobbs (38)
demonstrated that HCMV gene products may induce cell migration and
angiogenesis; however, the present study identified that HCMV gB
inhibited the migration capacity of MDA-MB-231 cells by Transwell
assay and by cell scratch test (Fig.
4).
A previous study indicated that TGF-β was a major
effector of breast tumor metastasis in vivo, and that the
inhibition of TGF-β signaling resulted in decreased metastasis in
mammary tumors (39). Furthermore,
decreased TGF-β responsiveness had no effect on the primary
tumorigenesis but significantly decreased cell metastasis in a
high-grade breast tumor cell line (40). Consistent with these previous data,
the results of the present study suggest that TGF-β downregulation
occurred in 231-GB-OE cells and accordingly, the migration
capability of 231-GB-OE cells was impaired (Fig. 5). Fynan and Reiss (41) demonstrated that the proliferation of
breast tumor cells derived from early stages of the disease was
inhibited by TGF-β, while a malignant breast carcinoma cell line
exhibited resistance to the anti-proliferative effect of TGF-β.
This is consistent with the data from the present study
demonstrating that HCMV gB may inhibit MDA-MB-231 migration and
TGF-β expression, whilst exhibiting no effect on cell
proliferation.
It has been established that the effects of TGF-β
are mediated primarily by the receptor-specific Smad2 and Smad3
proteins (42) and active Smad3
signaling contributes to breast cancer local invasion and distant
metastasis (43). In concordance with
these previous data, the present study identified that the
phosphorylation levels of Smad2 and Smad3 were decreased in
231-GB-OE cells compared with 231-NC cells (Fig. 6), indicating that the impaired
migration capacity in 231-GB-OE cells may be associated with the
inhibited TGF-β/Smad signaling. It was demonstrated previously that
in MCF-10-derived breast tumor cells, blockade of TGF-β signal
transduction resulted in a decreased chance of the development of
metastasis in advanced cancer types (40). Additionally, silencing TGF-β receptor
type-I may lead to a reduction in metastasis and invasiveness
(44). These implicate TGF-β as an
important target for the therapy of advanced cancers and suggest
that HCMV gB, which may inhibit TGF-β and Smad2/3, may serve as a
promising agent.
In the present study, it was initially demonstrated
that although HCMV infection is a risk factor or associated with
the development of breast cancer, HCMV gB may inhibit breast cancer
cell migration. The expression of TGF-β and phosphorylation of Smad
were also downregulated, which is consistent with a number of
studies that have demonstrated that TGF-β serves a crucial role in
breast cancer migration and metastasis (45–47).
However, the present study was not able to clarify how HCMV gB
downregulated the phosphorylation of TGF-β and Smad, and future
studies will be required to determine whether other signaling
pathways are involved and how HCMV gB led to TGF-β/Smad signaling
inhibition. In addition, the absence of a gB glycosylation assay to
detect protein glycosylation is a limitation of the present study.
Burke and Heldwein (48) revealed
that 17/18 N-linked glycosylation sites are completely conserved,
and hypothesized that this high conservation of the glycosylation
pattern in HCMV gB suggests that extensive glycosylation in HCMV gB
may instead protect functionally important regions from immune
recognition, but there are no clear conclusions how glycosylation
affects the protein functionality. Taken together, these studies
suggest that the identification of HCMV gB as a TGF-β/Smad signal
inhibitor may assist in exploitation of the effective therapeutic
strategies of the anti-TGF-β agents that contribute to tumor
suppression.
Acknowledgements
The authors would like to thank Hanyin Biotechnology
for helping with the lentiviral vector construction, and Dr
Yuan-Yuan Li for assistance with the western blot result
analysis.
Funding
The present study was supported by Jinshan Hospital
of Fudan University, Shanghai Jinshan District Health and Family
Planning (grant no. JSKJ-KTQN-2015-01).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article
Authors' contributions
YCL, conceived and designed the experiments; RY,
performed the experiments and analyzed the data; JL, manuscript and
data discussion; GXX, provided technical guidance during
experiments; LMD, HMH, QZS and JY, assisted with performing the
experiments.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Authors' information
Rui Yang, Department of Laboratory Medicine, Jinshan
Hospital, Fudan University, Shanghai, P.R. China. Yun-Chun Li,
corresponding author, Department of Laboratory Medcine, Jinshan
Hospital, Fudan University, Shanghai, P.R. China.
References
|
1
|
Britt V: Manifestations of human
cytomegalovirus infection: Proposed mechanisms of acute and chronic
diseaseHuman Cytomegalovirus. Shenk TE and Stinski MF: Springer
Berlin Heidelberg; Berlin, Heidelberg: pp. 417–470. 2008,
View Article : Google Scholar
|
|
2
|
Colugnati FA, Staras SA, Dollard SC and
Cannon MJ: Incidence of cytomegalovirus infection among the general
population and pregnant women in the United States. BMC Infect Dis.
7:12007. View Article : Google Scholar
|
|
3
|
Poole E, McGregor Dallas SR, Colston J,
Joseph RS and Sinclair J: Virally induced changes in cellular
microRNAs maintain latency of human cytomegalovirus in
CD34+ progenitors. J Gen Virol. 92:1539–1549. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Revello MG, Furione M, Rognoni V, Arossa A
and Gerna G: Cytomegalovirus DNAemia in pregnant women. J Clin
Virol. 61:590–592. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fowler KB and Boppana SB: Congenital
cytomegalovirus (CMV) infection and hearing deficit. J Clin Virol.
35:226–231. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cannon MJ: Congenital cytomegalovirus
(CMV) epidemiology and awareness. J Clin Virol. 46 Suppl 4:S6–S10.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
La Rosa C and Diamond DJ: The immune
response to human CMV. Future Virol. 7:279–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Söderberg-Nauclér C: HCMV microinfections
in inflammatory diseases and cancer. J Clin Virol. 41:218–223.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Varnum SM, Streblow DN, Monroe ME, Smith
P, Auberry KJ, Pasa-Tolic L, Wang D, Camp DG II, Rodland K, Wiley
S, et al: Identification of proteins in human cytomegalovirus
(HCMV) particles: The HCMV proteome. J Virol. 78:10960–10966. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Feire AL, Roy RM, Manley K and Compton T:
The glycoprotein B disintegrin-like domain binds beta 1 integrin to
mediate cytomegalovirus entry. J Virol. 84:10026–10037. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Anders CK and Carey LA: Biology,
metastatic patterns, and treatment of patients with triple-negative
breast cancer. Clin Breast Cancer. 9 Suppl 2:S73–S81. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lawson JS and Heng B: Viruses and breast
cancer. Cancers (Basel). 2:752–772. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Newton R, Ziegler J, Bourboulia D,
Casabonne D, Beral V, Mbidde E, Carpenter L, Reeves G, Parkin DM,
Wabinga H, et al: The sero-epidemiology of Kaposi's
sarcoma-associated herpesvirus (KSHV/HHV-8) in adults with cancer
in Uganda. Int J Cancer. 103:226–232. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fagundes CP, Glaser R, Alfano CM, Bennett
JM, Povoski SP, Lipari AM, Agnese DM, Yee LD, Carson WE III, Farrar
WB, et al: Fatigue and herpesvirus latency in women newly diagnosed
with breast cancer. Brain Behav Immun. 26:394–400. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Breathnach O, Donnellan P, Collins D,
McNicholas W and Crown J: Cytomegalovirus pneumonia in a patient
with breast cancer on chemotherapy: Case report and review of the
literature. Ann Oncol. 10:461–465. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Harkins LE, Matlaf LA, Soroceanu L, Klemm
K, Britt WJ, Wang W, Bland KI and Cobbs CS: Detection of human
cytomegalovirus in normal and neoplastic breast epithelium.
Herpesviridae. 1:82010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Isaacson MK and Compton T: Human
cytomegalovirus glycoprotein B is required for virus entry and
cell-to-cell spread but not for virion attachment, assembly, or
egress. J Virol. 83:3891–3903. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cobbs C, Khan S, Matlaf L, McAllister S,
Zider A, Yount G, Rahlin K, Harkins L, Bezrookove V, Singer E and
Soroceanu L: HCMV glycoprotein B is expressed in primary
glioblastomas and enhances growth and invasiveness via PDGFR-alpha
activation. Oncotarget. 5:1091–1100. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Costanza B, Umelo IA, Bellier J,
Castronovo V and Turtoi A: Stromal Modulators of TGF-β in Cancer. J
Clin Med. 6:72017. View Article : Google Scholar
|
|
21
|
Derynck R, Akhurst RJ and Balmain A:
TGF-beta signaling in tumor suppression and cancer progression. Nat
Genet. 29:117–129. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cao Y, Liu X, Zhang W, Deng X, Zhang H,
Liu Y, Chen L, Thompson EA, Townsend CM Jr and Ko TC: TGF-beta
repression of Id2 induces apoptosis in gut epithelial cells.
Oncogene. 28:1089–1098. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cipriano R, Kan CE, Graham J, Danielpour
D, Stampfer M and Jackson MW: TGF-beta signaling engages an
ATM-CHK2-p53-independent RAS-induced senescence and prevents
malignant transformation in human mammary epithelial cells. Proc
Natl Acad Sci USA. 108:8668–8673. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ewen ME, Sluss HK, Whitehouse LL and
Livingston DM: TGF beta inhibition of Cdk4 synthesis is linked to
cell cycle arrest. Cell. 74:1009–1020. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lebrun JJ: The dual role of TGFβ in human
cancer: From tumor suppression to cancer metastasis. ISRN Mol Biol.
2012:3814282012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Derynck R, Zhang Y and Feng XH:
Transcriptional activators of TGF-beta responses. Cell. 95:737–740.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kang Y: Pro-metastasis function of TGFbeta
mediated by the smad pathway. J Cell Biochem. 98:1380–1390. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Uhl M, Aulwurm S, Wischhusen J, Weiler M,
Ma JY, Almirez R, Mangadu R, Liu YW, Platten M, Herrlinger U, et
al: SD-208, a novel transforming growth factor beta receptor I
kinase inhibitor, inhibits growth and invasiveness and enhances
immunogenicity of murine and human glioma cells in vitro and in
vivo. Cancer Res. 64:7954–7961. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mohammad KS, Javelaud D, Fournier PG,
Niewolna M, McKenna CR, Peng XH, Duong V, Dunn LK, Mauviel A and
Guise TA: TGF-beta-RI kinase inhibitor SD-208 reduces the
development and progression of melanoma bone metastases. Cancer
Res. 71:175–184. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dunn LK, Mohammad KS, Fournier PG, McKenna
CR, Davis HW, Niewolna M, Peng XH, Chirgwin JM and Guise TA:
Hypoxia and TGF-beta drive breast cancer bone metastases through
parallel signaling pathways in tumor cells and the bone
microenvironment. PLoS One. 4:e68962009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ishiyama M, Miyazono Y, Sasamoto K, Ohkura
Y and Ueno K: A highly water-soluble disulfonated tetrazolium salt
as a chromogenic indicator for NADH as well as cell viability.
Talanta. 44:1299–1305. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Massagué J and Gomis RR: The logic of TGFβ
signaling. FEBS Lett. 580:2811–2820. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mitchell DA, Xie W, Schmittling R, Learn
C, Friedman A, McLendon RE and Sampson JH: Sensitive detection of
human cytomegalovirus in tumors and peripheral blood of patients
diagnosed with glioblastoma. Neuro Oncol. 10:10–18. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Scheurer ME, Bondy ML, Aldape KD, Albrecht
T and El Zein R: Detection of human cytomegalovirus in different
histological types of gliomas. Acta Neuropathol. 116:79–86. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Soroceanu L, Akhavan A and Cobbs CS:
Platelet-derived growth factor-alpha receptor activation is
required for human cytomegalovirus infection. Nature. 455:391–395.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vanarsdall AL, Wisner TW, Lei H,
Kazlauskas A and Johnson DC: PDGF receptor-α does not promote HCMV
entry into epithelial and endothelial cells but increased
quantities stimulate entry by an abnormal pathway. PLoS Pathog.
8:e10029052012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Utrera-Barillas D, Valdez-Salazar HA,
Gómez-Rangel D, Alvarado-Cabrero I, Aguilera P, Gómez-Delgado A and
Ruiz-Tachiquin ME: Is human cytomegalovirus associated with breast
cancer progression. Infect Agent Cancer. 8:122013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Soroceanu L and Cobbs CS: Is HCMV a tumor
promoter? Virus Res. 157:193–203. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Muraoka RS, Dumont N, Ritter CA, Dugger
TC, Brantley DM, Chen J, Easterly E, Roebuck LR, Ryan S, Gotwals
PJ, et al: Blockade of TGF-beta inhibits mammary tumor cell
viability, migration, and metastases. J Clin Invest. 109:1551–1559.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tang B, Vu M, Booker T, Santner SJ, Miller
FR, Anver MR and Wakefield LM: TGF-beta switches from tumor
suppressor to prometastatic factor in a model of breast cancer
progression. J Clin Invest. 112:1116–1124. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fynan TM and Reiss M: Resistance to
inhibition of cell growth by transforming growth factor-beta and
its role in oncogenesis. Crit Rev Oncog. 4:493–540. 1993.PubMed/NCBI
|
|
42
|
Liu X, Sun Y, Constantinescu SN, Karam E,
Weinberg RA and Lodish HF: Transforming growth factor beta-induced
phosphorylation of Smad3 is required for growth inhibition and
transcriptional induction in epithelial cells. Proc Natl Acad Sci
USA. 94:10669–10674. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Petersen M, Pardali E, van der Horst G,
Cheung H, van den Hoogen C, van der Pluijm G and Ten Dijke P: Smad2
and Smad3 have opposing roles in breast cancer bone metastasis by
differentially affecting tumor angiogenesis. Oncogene.
29:1351–1361. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Iyer S, Wang ZG, Akhtari M, Zhao W and
Seth P: Targeting TGFbeta signaling for cancer therapy. Cancer Biol
Ther. 4:261–266. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Han G, Lu SL, Li AG, He W, Corless CL,
Kulesz-Martin M and Wang XJ: Distinct mechanisms of
TGF-beta1–mediated epithelial-to-mesenchymal transition and
metastasis during skin carcinogenesis. J Clin Invest.
115:1714–1723. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Korpal M, Yan J, Lu X, Xu S, Lerit DA and
Kang Y: Imaging transforming growth factor-beta signaling dynamics
and therapeutic response in breast cancer bone metastasis. Nat Med.
15:960–966. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jiang HL, Sun HF, Gao SP, Li LD, Hu X, Wu
J and Jin W: Loss of RAB1B promotes triple-negative breast cancer
metastasis by activating TGF-β/SMAD signaling. Oncotarget.
6:16352–16365. 2015.PubMed/NCBI
|
|
48
|
Burke HG and Heldwein EE: Crystal
structure of the human cytomegalovirus glycoprotein B. PLoS Pathog.
11:e10053002015. View Article : Google Scholar : PubMed/NCBI
|