Introduction
Neutrophils serve crucial roles in the
anti-bacterial and anti-fungal innate immune response and produce
cytokines to initiate inflammatory responses; however, their
inappropriate or excessive activation contributes to tissue damage
during autoimmune and inflammatory diseases (1). Furthermore, the activity of neutrophils
is associated with tumor progression, as demonstrated in
colitis-associated cancer and glioblastoma (2,3).
Previous studies have revealed that in order to
protect the human body from invading pathogens and endogenous
damage-associated molecules, neutrophils sense these threats
through innate immune receptors, such as toll-like receptors (TLRs)
and other pattern recognition receptors, to upregulate the
secretion of inflammatory cytokines (4,5). The
recognition of lipopolysaccharide (LPS) from gram-negative bacteria
by TLR4 is a well-characterized example with the initiation of
proinflammatory gene transcription through multiple signaling
pathways, including nuclear factor (NF)-κB) (6,7). The
exposure of neutrophils to bacterial LPS may contribute to tissue
damage, under certain conditions, including endotoxic shock and
infection-induced adult respiratory distress syndrome (8).
Neddylation is a novel-type protein
post-translational modification, a multistep enzymatic process that
adds the ubiquitin-like molecule neural precursor cell expressed
developmentally downregulated protein 8 (Nedd8) to target proteins
in an ATP-dependent manner. Neddylation is vital for a range of
processes, including cell viability, growth and development
(9–12). To the best of our knowledge, the only
known E1 for neddylation is a heterodimer comprised of the amyloid
precursor protein-binding protein and ubiquitin-like modifier
activating enzyme 3 (9–12). A previous study identified the
ubiquitin-conjugating enzymes Ube2M and Ube2F as Nedd8 E2s
(13). Several targets for Nedd8 have
been identified, however the well-characterized substrates of the
neddylation system are Cullins and key components of Cullin-RING E3
ligases (CRLs) (14,15). Neddylation activates CRLs, which
mediate the ubiquitination and subsequent degradation of various
cellular proteins, including inhibitor of κB (IκB), that sequester
NF-κB to the cytoplasm (16–18). Based on previous studies demonstrating
that it suppressed the growth of diverse cancer cell lines,
MLN4924, a small-molecule inhibitor of Nedd8 E1, may be a novel
anticancer therapeutic agent (16–18).
Additionally, previous studies also revealed that MLN4924
suppresses proinflammatory cytokine production in macrophages and
dendritic cells by preventing the degradation of IκB proteins.
Consequently, MLN4924 treatment led to alleviated mucosal
inflammation in a colitis mouse model, indicating that neddylation
may be involved in immune regulation (6,19,20).
Under certain conditions, neutrophils behave
distinctly to macrophages in response to LPS (21). Nevertheless, the association between
neddylation and LPS-induced proinflammatory cytokine production in
neutrophils remains unclear. The results of the present study
revealed that neddylation inhibition by MLN4924 led to the
suppression of LPS-induced proinflammatory cytokine production in
neutrophils in a dose-dependent manner by disrupting the NF-κB
signaling pathway; this suggests that MLN4924 may serve a role as a
potential chemotherapeutic agent for neutrophil-mediated autoimmune
or inflammatory diseases.
Materials and methods
Mice
Female C57BL/6 mice at the age of 6–8 weeks were
purchased from Beijing Vital River Laboratory Animal, Inc.
(Beijing, China). All mice were maintained under specific
pathogen-free conditions. The care, use and treatment of mice in
the present study strictly followed the guidelines set by the
Institute of Basic Medical Sciences. The Institute of Basic Medical
Sciences (Beijing, China), from whom ethical approval was granted,
approved the present study.
Reagents
Antibodies against phosphorylated (p)-IκBα (catalog
no. #2859), p-IκB kinase (IKK)α/β (catalog no. #2697) and Nedd8
(catalog no. #2745) were purchased from Cell Signaling Technology,
Inc. (Danvers, MA, USA). Antibodies against IκBα (catalog no.
sc-7877), IKKα/β (catalog no. sc-7606) and β-actin (catalog no.
sc-58673) were purchased from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). MLN4924, dissolved in dimethyl sulfoxide, was
purchased from Active Biochemku Ltd. (Maplewood, NJ, USA). The
ATPlite 1 Step kit was purchased from Perkin Elmer, Inc. (Waltham,
MA, USA). LPS (serotype, Escherichia coli) and thioglycolate
were purchased from Sigma-Aldrich (Merck KGaA, Darmstadt,
Germany).
Flow cytometry analysis
Mice received 2 ml 3% thioglycolate by
intraperitoneal injection (Sigma-Aldrich; Merck KGaA). After 4 h,
the mice were anaesthetized by administrating pentobarbital (50
mg/kg) and were subjected to euthanasia by cervical dislocation.
The peritoneal cells were washed in 6 ml of 3 mM EDTA in PBS. The
cells were washed with fluorescence-activated cell sorting (FACS)
washing buffer [2% fetal bovine serum (Amresco, LLC, Solon, OH,
USA) 0.1% NaN3 in PBS] twice prior to incubation with
fluorescein isothiocyanate-conjugated antibodies [anti-F4/80
(catalog no. 565409), anti-γδ T cell receptor (TCR; catalog no.
561486) or anti-cluster of differentiation (CD)19 (catalog no.
561738)], phycoerythrin (PE)-conjugated antibodies [anti-Gr1
(catalog no. 560601), anti-natural killer cell (NK)1.1 (catalog no.
553164)], or allophycocyanin-conjugated anti-CD3 (catalog no.
563123) (1 µg/1×106 cells; BD Pharmingen; BD
Biosciences, Franklin Lakes, NJ, USA) for 30 min on ice in the
presence of 2.4G2 monoclonal antibody (catalog no. 562896; 1
µg/1×106 cells; BD Pharmingen; BD Biosciences, Franklin
Lakes, NJ, USA) to block Fcγ receptor binding. Isotype antibodies
[FITC-anti-Mouse IgG1 (catalog no. 550616), PE-anti-Mouse IgG1
(catalog no. 562027), APC-anti-Mouse IgG1 (catalog no. 550874)]
were included as the negative control. Following washing with FACS
buffer, the cells were fixed with 1% (w/v) paraformaldehyde in PBS
and preserved at 4°C. Flow cytometry 6 was carried out with a
Becton Dickinson FACSCalibur™ (BD Biosciences) and analyzed using
FlowJo software (version 10; FlowJo LLC, Ashland, OR, USA).
Purification of neutrophils
Peritoneal cells were stained with PE-conjugated
anti-Gr1 antibody as above. Other cells were then magnetically
separated using anti-PE antibody-coated mircobeads (Miltenyi
Biotec, Inc., Auburn, CA, USA), according to the manufacturer's
protocol. Cell purity was verified to be >90% by flow
cytometry.
Western blot
The purified neutrophils were seeded into 24-well
plates at a density of 1×106 cells/well. Neutrophils
were treated by MLN4924 with various concentrations (0, 0.1, 0.5
and 2.5 µM) for 30 min at 37°C and then washed with PBS and
harvested using ice-cold lysis buffer (0.5% Tert-NP-40, 20 mM
Tris-Cl, pH 7.6, 250 mM NaCl, 3 mM EDTA, 3 mM EGTA, 1 mM sodium
orthovanadate, 1 mM dithiothreitol, 10 mM PNPP, 10 mg/ml
aprotinin). Protein concentration was determined by bicinchoninic
acid assay (cat. no. SK3021; Bio Basic Inc., Ontario, Canada)
according to the manufacturer's protocol. Cell lysates were
resolved by SDS-PAGE (10% gel; 20 µg protein/lane) prior to being
transferred to nitrocellulose membranes. Nitrocellulose membranes
were then incubated with 5% (w/v) nonfat dry milk in washing buffer
(20 mM Tris-Cl, pH 7.6, 150 mM NaCl, and 0.1% Tween 20) for 1 h at
37°C to block nonspecific protein binding. Primary antibodies
against Nedd8, (p)-IκBα, IκBα, IKKα/β, (p)-IKKα/β and β-actin were
diluted in PBS (1:1,000) containing 5% bovine serum albumin
(Sigma-Aldrich; Merck KGaA) and applied to the membranes overnight
at 4°C. Following extensive washing, the membranes were incubated
with goat anti-rabbit IgG-horseradish peroxidase (HRP) or
anti-mouse IgG-HRP (1:2,500 in washing buffer containing 5% (w/v)
nonfat dry milk) antibodies for 1 h at room temperature. Following
washing, immunoreactive bands were visualized using an
ECL™ Western Blotting Detection Reagents (GE Healthcare,
Chicago, IL, USA).
ELISA
The purified neutrophils were seeded into 24-well
plates at a density of 1×106 cells/well. The
supernatants were collected at 0 and 4 h after LPS treatment. The
expression levels of TNF-α (catalog no. 70-EK2822/2), IL-6 (catalog
no. 70-EK2062/2), IL-10 (catalog no. 70-EK2102/2) and IL-1β
(catalog no. 70-EK201B2/2) were measured using ELISA kits purchased
from eBioscience (Thermo Fisher Scientific, Inc.) following the
manufacturer's protocols.
ATPlite assays
The purified cells were seeded into 96-well plates
(2.5×105 cells/well). Following pretreatment with
distinct doses (0, 0.1, 0.5 and 2.5 µM) of MLN4924 for 30 min at
37°C, cells were stimulated with 100 ng/ml LPS for 4 h. Then, cell
viability was assessed using an ATPlite 1 step Single Addition
Luminescence ATP Detection Assay System, according to the
manufacturer's protocol (Perkin Elmer, Waltham, MA, USA).
Statistical analysis
The data are presented as the mean ± standard
deviation using SPSS software (version 19.0; IBM Corp., Armonk, NY,
USA). The data are presented as the mean ± standard deviation. The
Student's t-test was employed to determine significant differences
between two groups (paired or unpaired), and one-way analysis of
variance (Please refer to cover letter) was used to determine
significant differences among multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
MLN4924 suppresses the neddylation of
Cullins in neutrophils in a dose-dependent manner
To examine the possible role of neddylation in
neutrophils, peritoneal neutrophils were induced by a 3%
thioglycolate intraperitoneal injection. Flow cytometry revealed
that the percentage of peritoneal neutrophils (Gr1+) in the treated
mice markedly increased after 4 h, whereas the percentages of
macrophages (F4/80+), γδT cells (CD3+γδTCR+), T cells (CD3+NK1.1-),
NK cells (CD3-NK1.1+), NKT cells (CD3+NK1.1+) and B cells (CD19+)
decreased in the treated mice, to varying extents, as compared with
in the non-treated mice (Fig. 1A-E).
Peritoneal neutrophils were then purified with magnetic microbeads
(Fig. 1F).
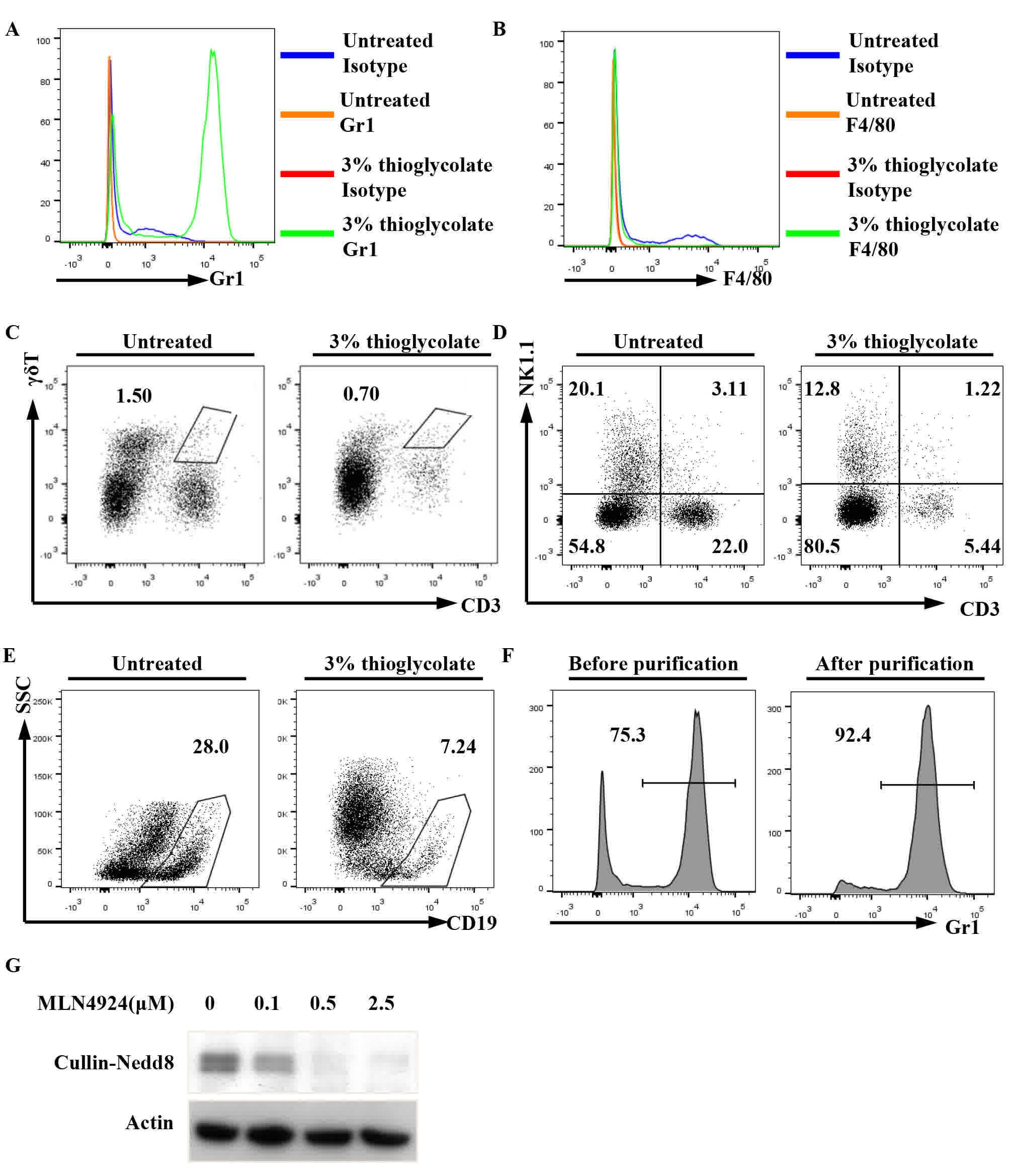 | Figure 1.MLN4924 suppressed the neddylation of
Cullins in neutrophils in a dose-dependent manner. Mice received an
intraperitoneal injection of 3% thioglycolate or were left
untreated. After 4 h, peritoneal cells were subjected to flow
cytometry to examine the percentages of (A) neutrophils, (B)
macrophages, (C) γδT cells, (D) NKT cells and (E) B cells. (F) The
purity of Gr1+ cells was >90% as confirmed by flow cytometry.
(G) The purified peritoneal neutrophils were seeded into 24-well
plates at a density of 1×106 cells/well. Following
treatment with various doses of MLN4924, ranging between 0 and 2.5
µM for 30 min, the cell lysates were prepared and used for western
blot analysis with antibodies against Nedd8 and β-actin
(representative data are presented; n>3). Nedd8, neural
precursor cell expressed developmentally downregulated protein 8;
CD, cluster of differentiation; SSC, side scattering; NK, natural
killer. |
To explore whether MLN4924 suppresses the
neddylation of Cullins in neutrophils, immunoblotting analysis with
an antibody against Nedd8 was used. A major band between 90 and 100
kDA is usually recognized as neddylated Cullins (Nedd8-Cullin) and
was examined in the present study (22–27). As
predicted, the major band was detected in the neutrophils, and
MLN4924 treatment for 30 min led to diminished Nedd8-Cullin in a
dose-dependent manner (Fig. 1G). The
results suggest that neddylation is active in neutrophils and that
MLN4924 inhibits the neddylation of Cullins in neutrophils.
MLN4924 inhibits LPS-induced
proinflammatory cytokine production in neutrophils in a
dose-dependent manner
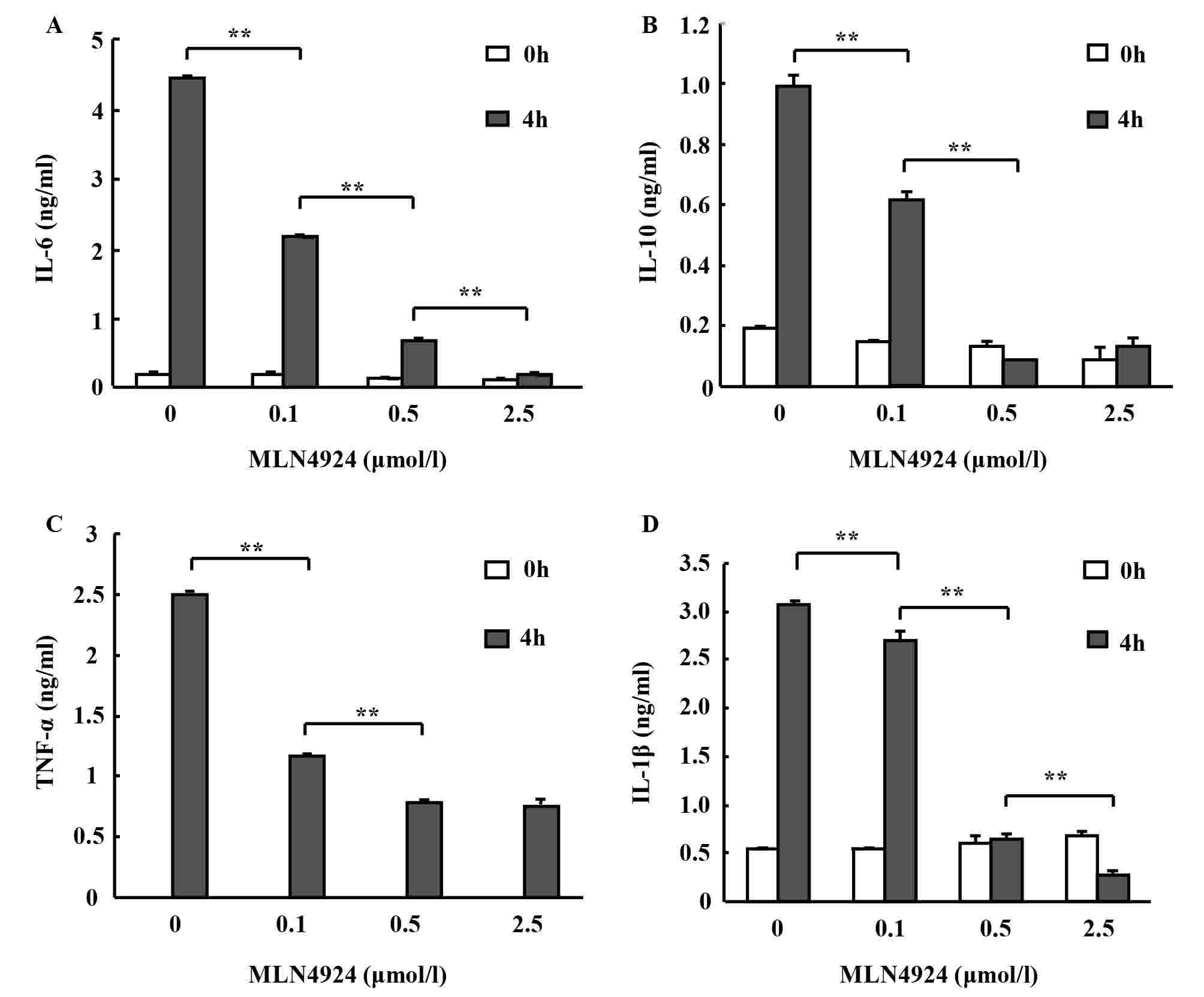
To investigate whether MLN4924 affects inflammatory
cytokine secretion from neutrophils, peritoneal neutrophils were
pretreated with various doses of MLN4924 for 30 min followed by
stimulation with or without LPS for 4 h. Culture supernatants of
the neutrophils were collected and the levels of TNF-α, IL-6 and
IL-10 were determined using an ELISA. MLN4924 inhibited LPS-induced
IL-6, IL-10 (observed in low concentrations in neutrophils) and
TNF-α production in a dose-dependent manner (Fig. 2A-C; *P<0.05, **P<0.01). The
decrease of the inflammatory cytokine secretion from neutrophils is
associated with Cullins neddylation. The results indicate that
MLN4924 may suppress the neutrophil-mediated inflammatory
response.
Nedd8 silencing, or treatment with MLN4924, led to
diminished caspase-1 processing and decreased IL-1β maturation in
macrophages and epithelial cells following inflammasome activation
(28,29). Aberrant IL-1β production may lead to
various diseases, including pneumonia, diabetes, atherosclerosis,
obesity, cancer, Alzheimer's and arthritis (30,31).
Therefore, mature IL-1β was detected in the culture media in the
presence of ATP and it was observed that MLN4924 inhibited
LPS-induced production of mature IL-1β in neutrophils in a
dose-dependent manner, as revealed by ELISA (Fig. 2D).
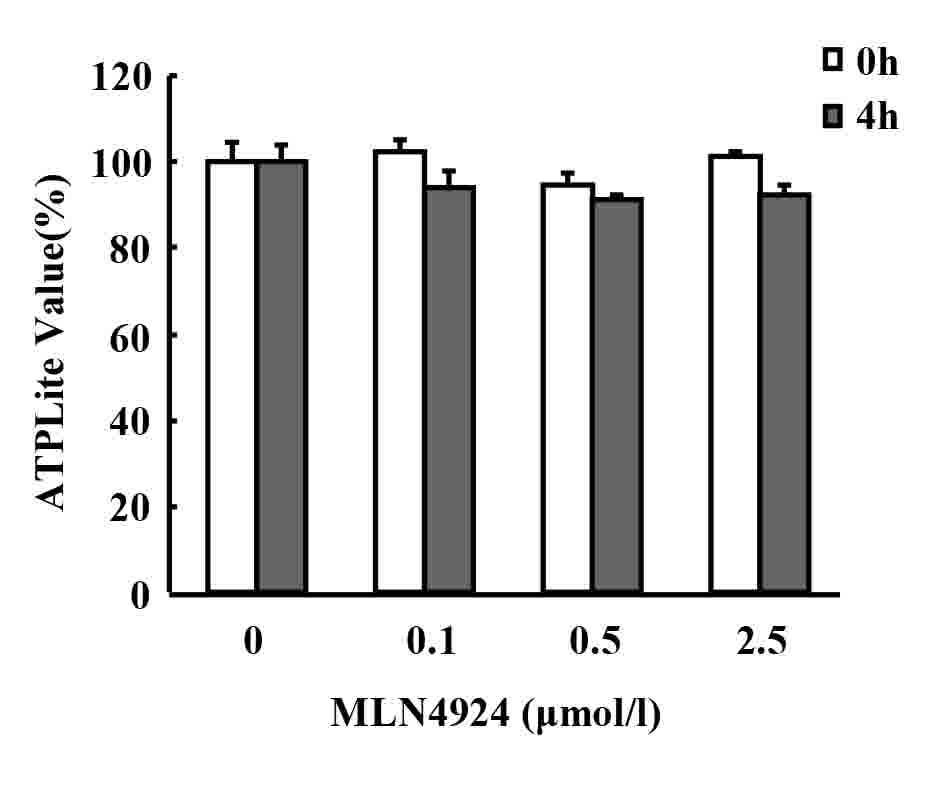
MLN4924 pretreatment followed by LPS
stimulation for 4 h revealed marginal effects on the viability of
neutrophils
The neddylation inhibitor MLN4924 has been
demonstrated to suppress the oncogenic growth of various
hematological malignancies, including myeloma, B-cell lymphoma and
acute myeloid leukemia via cell cycle arrest and apoptosis
(22,32,33).
Furthermore, prolonged and complete (1.0 µM for 24 or 48 h)
inhibition of neddylation led to decreased macrophage viability
(34). To detect whether MLN4924
inhibited LPS-induced inflammatory cytokine production in
neutrophils as a result of neutrophil apoptosis, the viability of
neutrophils was examined using ATPlite assays. As presented in
Fig. 3, MLN4924 pretreatment for 30
min followed by LPS stimulation for 4 h demonstrated only marginal
effects on the viability of neutrophils and no statistically
significant differences were identified.
MLN4924 inhibits LPS-induced
proinflammatory cytokine production in neutrophils by suppressing
NF-κB
LPS can induce proinflammatory cytokine production
in neutrophils through activating the IKK/NF-κB signaling pathway
(27). The NF-κB signaling pathway is
also associated with the production of pro-IL-1β (35). Neddylation has been demonstrated to
serve a key role in the degradation of IκB proteins and subsequent
NF-κB activation (6,27). The present study investigated whether
a similar molecular mechanism occurred in neutrophils; as expected,
MLN4924 suppressed the degradation and resynthesis of IκBα, which
is associated with accumulated P-IκBα. However, MLN4924 treatment
exhibited no effect on the phosphorylation of IKK (Fig. 4A). The results suggest that the
inhibition of NF-κB signaling may be involved in the inhibitory
effects of MLN4924.
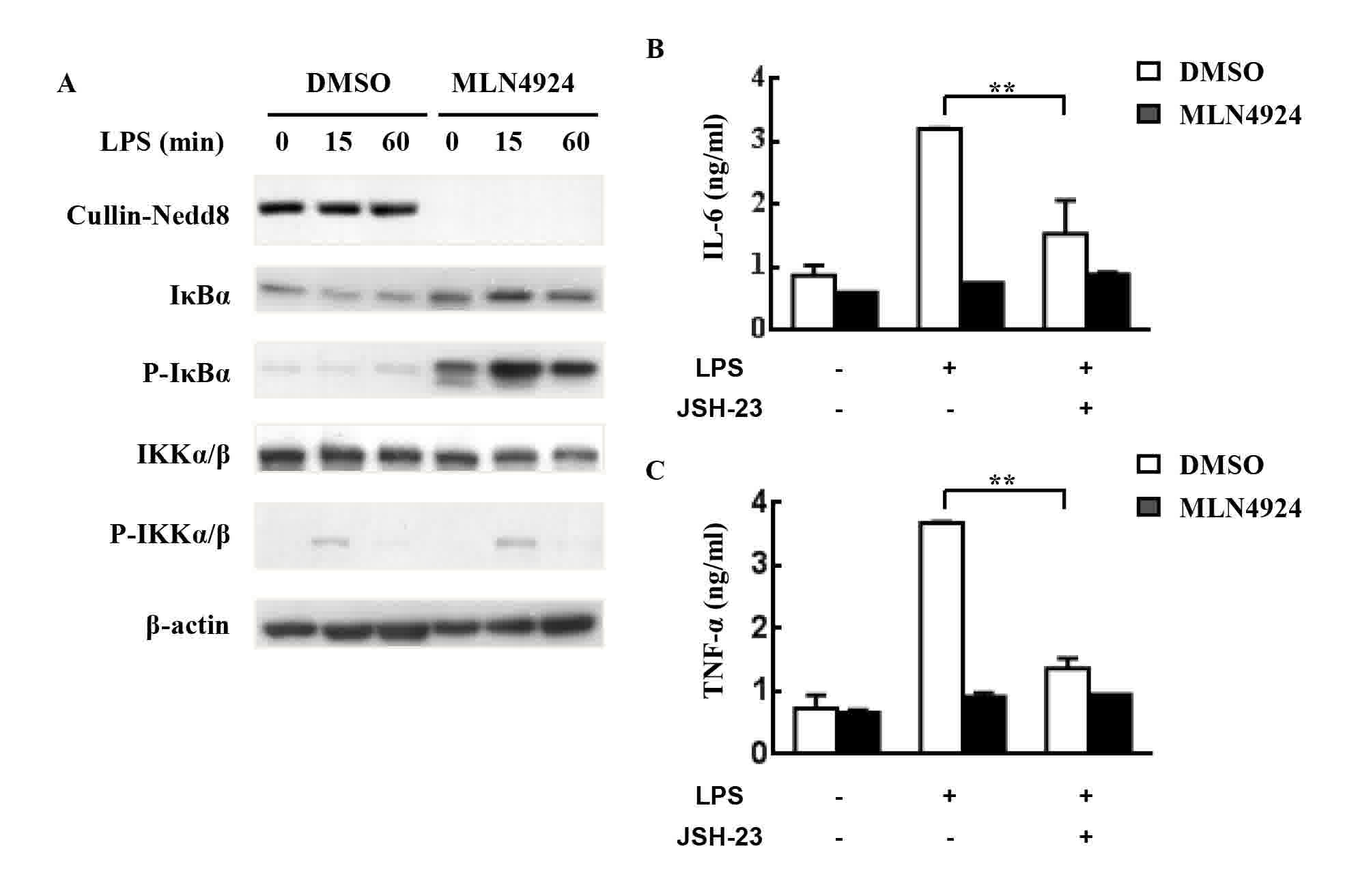 | Figure 4.MLN4924 inhibits LPS-induced
proinflammatory cytokine production from neutrophils by suppressing
NF-κB. (A) The purified peritoneal neutrophils were seeded into
24-well plates at a density of 5×106 cells/well.
Following pretreatment with 0.5 µM MLN4924 for 30 min, cells were
stimulated with 100 ng/ml LPS for 0, 15 or 60 min. Cell lysates
were then prepared to analyze the levels of neddylated Cullins,
P-IKKα/β, P-IκBα, IKKα/β, IκBα and β-actin. Neutrophils were
pretreated with 0.5 µM MLN4924 and/or 80 µM JSH-23 or an equal
volume of DMSO for 30 min. Then neutrophils were treated with LPS
for the indicated periods of time. The concentration of (B) IL-6
and (C) TNF-α in the supernatants was measured with ELISA.
**P<0.01. LPS, lipopolysaccharide; NF-κB, nuclear factor κB;
IκB, inhibitor of κB; P-IKK, phosphorylated IκB kinase; DMSO,
dimethyl sulfoxide; IL, interleukin; TNF, tumor necrosis
factor. |
To investigate the role NF-κB has in the inhibitory
effects of MLN4924 on LPS-induced proinflammatory cytokine
production, neutrophils were pretreated with JSH-23, a specific
inhibitor of NF-κB nuclear translocation and transcriptional
activity (36), prior to LPS
treatment for 4 h in the presence or absence of MLN4924. ELISA
revealed that JSH-23 treatment alone resulted in marked inhibition
of LPS-induced IL-6/TNF-α production (Fig. 4B and C). The inhibitory effects of
MLN4924 on LPS-induced proinflammatory cytokine production
diminished in the presence of the NF-κB inhibitor JSH-23 (Fig. 4B and C). Thus, the results of the
present study suggest that neddylation inhibition may suppress
neutrophil functions through, at least partially, suppression of
the NF-κB signaling pathway.
Discussion
The results of the present study demonstrated that
the neddylation inhibitor MLN4924 suppressed the proinflammatory
cytokine production in neutrophils in a dose-dependent manner,
which is consistent with that observed in mononuclear cells
(6,19,20,28,37).
In order to purify peritoneal neutrophils induced by 3%
thioglycolate intraperitoneal injection, peritoneal cells were
stained with PE-conjugated anti-mouse Gr1, followed by isolation
with anti-PE antibody-coated magnetic microbeads (38). The purity of neutrophils was verified
to be >90% by flow cytometry.
Previous studies had demonstrated that MLN4924 may
be used as an anticancer treatment, being tested in Phase I
clinical trials for both hematological and non-hematological tumors
(39,40). Previous studies have also revealed
that MLN4924 markedly suppressed inflammatory responses mediated by
innate immune cells, including macrophages and dendritic cells
(19,20,34). The
results of the present study suggest that the same suppressed
inflammatory response is experienced in neutrophils. As neutrophils
promote tumor progression under certain circumstance (2,3),
neddlylation may contribute to tumor growth partially through
enhancing neutrophil-mediated inflammation.
However, the mechanism by which neddlylation
promotes inflammation remains unclear. NF-κB proteins are a family
of dimeric transcription factors that mediate the expression of
inflammatory cytokines (41);
transcriptional mediators are pre-existing in the cytoplasm and are
able to be rapidly activated by various extracellular stimuli,
including microbial components and inflammatory cytokines, prior to
entry into the nucleus to mediate an inflammatory response
(42). The degradation of IκB
proteins is the key to NF-κB activity (42). Impaired degradation of IκBα upon
neddylation inhibition has been attributed to the inhibitory
effects of MLN4924 on cytokine production in macrophages and
dendritic cells (6,19,20,37).
Hence, MLN4924 blocks neddylation, thereby suppressing production
of inflammatory cytokines by inhibiting the NF-κB signaling
pathway. However, mechanisms other than the NF-κB signaling pathway
may also be involved; therefore, further studies are required to
investigate this issue.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Futosi K, Fodor S and Mocsai A: Reprint of
Neutrophil cell surface receptors and their intracellular signal
transduction pathways. Int Immunopharmacol. 17:1185–1197. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang Y, Wang K, Han GC, Wang RX, Xiao H,
Hou CM, Guo RF, Dou Y, Shen BF, Li Y and Chen GJ: Neutrophil
infiltration favors colitis-associated tumorigenesis by activating
the interleukin-1 (IL-1)/IL-6 axis. Mucosal Immunol. 7:1106–1115.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rahbar A, Cederarv M, Wolmer-Solberg N,
Tammik C, Stragliotto G, Peredo I, Fornara O, Xu X, Dzabic M, Taher
C, et al: Enhanced neutrophil activity is associated with shorter
time to tumor progression in glioblastoma patients. Oncoimmunology.
5:e10756932015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sabroe I, Prince LR, Jones EC, Horsburgh
MJ, Foster SJ, Vogel SN, Dower SK and Whyte MK: Selective roles for
Toll-like receptor (TLR)2 and TLR4 in the regulation of neutrophil
activation and life span. J Immunol. 170:5268–5275. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takeuchi O and Akira S: Pattern
recognition receptors and inflammation. Cell. 140:805–820. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chang FM, Reyna SM, Granados JC, Wei SJ,
Innis-Whitehouse W, Maffi SK, Rodriguez E, Slaga TJ and Short JD:
Inhibition of neddylation represses lipopolysaccharide-induced
proinflammatory cytokine production in macrophage cells. J Biol
Chem. 287:35756–35767. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sabroe I, Parker LC, Wilson AG, Whyte MK
and Dower SK: Toll-like receptors: Their role in allergy and
non-allergic inflammatory disease. Clin Exp Allergy. 32:984–989.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guthrie LA, McPhail LC, Henson PM and
Johnston RB Jr: Priming of neutrophils for enhanced release of
oxygen metabolites by bacterial lipopolysaccharide. Evidence for
increased activity of the superoxide-producing enzyme. J Exp Med.
160:1656–1671. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Loftus SJ, Liu G, Carr SM, Munro S and La
Thangue NB: NEDDylation regulates E2F-1-dependent transcription.
EMBO Rep. 13:811–818. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xirodimas DP: Novel substrates and
functions for the ubiquitin-like molecule NEDD8. Biochem Soc Trans.
36:802–806. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xirodimas DP, Saville MK, Bourdon JC, Hay
RT and Lane DP: Mdm2-mediated NEDD8 conjugation of p53 inhibits its
transcriptional activity. Cell. 118:83–97. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zuo W, Huang F, Chiang YJ, Li M, Du J,
Ding Y, Zhang T, Lee HW, Jeong LS, Chen Y, et al: C-Cbl-mediated
neddylation antagonizes ubiquitination and degradation of the TGF-β
type II receptor. Mol Cell. 49:499–510. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang DT, Ayrault O, Hunt HW, Taherbhoy
AM, Duda DM, Scott DC, Borg LA, Neale G, Murray PJ, Roussel MF and
Schulman BA: E2-RING expansion of the NEDD8 cascade confers
specificity to cullin modification. Mol Cell. 33:483–495. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liakopoulos D, Doenges G, Matuschewski K
and Jentsch S: A novel protein modification pathway related to the
ubiquitin system. EMBO J. 17:2208–2214. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Osaka F, Kawasaki H, Aida N, Saeki M,
Chiba T, Kawashima S, Tanaka K and Kato S: A new NEDD8-ligating
system for cullin-4A. Genes Dev. 12:2263–2268. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luo Z, Yu G, Lee HW, Li L, Wang L, Yang D,
Pan Y, Ding C, Qian J, Wu L, et al: The Nedd8-activating enzyme
inhibitor MLN4924 induces autophagy and apoptosis to suppress liver
cancer cell growth. Cancer Res. 72:3360–3371. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu T, Wang J, Pei Y, Wang Q, Wu Y, Qiu G,
Zhang D, Lv M, Li W and Zhang J: Neddylation controls basal MKK7
kinase activity in breast cancer cells. Oncogene. 35:2624–2633.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gao F, Cheng J, Shi T and Yeh ET:
Neddylation of a breast cancer-associated protein recruits a class
III histone deacetylase that represses NFkappaB-dependent
transcription. Nat Cell Biol. 8:1171–1177. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mathewson N, Toubai T, Kapeles S, Sun Y,
Oravecz-Wilson K, Tamaki H, Wang Y, Hou G, Sun Y and Reddy P:
Neddylation plays an important role in the regulation of murine and
human dendritic cell function. Blood. 122:2062–2073. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cheng M, Hu S, Wang Z, Pei Y, Fan R, Liu
X, Wang L, Zhou J, Zheng S, Zhang T, et al: Inhibition of
neddylation regulates dendritic cell functions via Deptor
accumulation driven mTOR inactivation. Oncotarget. 7:35643–35654.
2016.PubMed/NCBI
|
|
21
|
Ajibade AA, Wang Q, Cui J, Zou J, Xia X,
Wang M, Tong Y, Hui W, Liu D, Su B, et al: TAK1 negatively
regulates NF-κB and p38 MAP kinase activation in Gr-1+CD11b+
neutrophils. Immunity. 36:43–54. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Swords RT, Kelly KR, Smith PG, Garnsey JJ,
Mahalingam D, Medina E, Oberheu K, Padmanabhan S, O'Dwyer M,
Nawrocki ST, et al: Inhibition of NEDD8-activating enzyme: A novel
approach for the treatment of acute myeloid leukemia. Blood.
115:3796–3800. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hjerpe R, Thomas Y, Chen J, Zemla A,
Curran S, Shpiro N, Dick LR and Kurz T: Changes in the ratio of
free NEDD8 to ubiquitin triggers NEDDylation by ubiquitin enzymes.
Biochem J. 441:927–936. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pelham CJ, Ketsawatsomkron P, Groh S,
Grobe JL, de Lange WJ, Ibeawuchi SR, Keen HL, Weatherford ET,
Faraci FM and Sigmund CD: Cullin-3 regulates vascular smooth muscle
function and arterial blood pressure via PPARgamma and
RhoA/Rho-kinase. Cell Metab. 16:462–472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guihard S, Ramolu L, Macabre C, Wasylyk B,
Noël G, Abecassis J and Jung AC: The NEDD8 conjugation pathway
regulates p53 transcriptional activity and head and neck cancer
cell sensitivity to ionizing radiation. Int J Oncol. 41:1531–1540.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Collier-Hyams LS, Sloane V, Batten BC and
Neish AS: Cutting edge: Bacterial modulation of epithelial
signaling via changes in neddylation of cullin-1. J Immunol.
175:4194–4198. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jubelin G, Taieb F, Duda DM, Hsu Y,
Samba-Louaka A, Nobe R, Penary M, Watrin C, Nougayrède JP, Schulman
BA, et al: Pathogenic bacteria target NEDD8-conjugated cullins to
hijack host-cell signaling pathways. PLoS Pathog. 6:e10011282010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Segovia JA, Tsai SY, Chang TH, Shil NK,
Weintraub ST, Short JD and Bose S: Nedd8 regulates
inflammasome-dependent caspase-1 activation. Mol Cell Biol.
35:582–597. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vince JE and Silke J: The intersection of
cell death and inflammasome activation. Cell Mol Life Sci.
73:2349–2367. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Davis BK, Wen H and Ting JP: The
inflammasome NLRs in immunity, inflammation, and associated
diseases. Annu Rev Immunol. 29:707–735. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Franchi L, Eigenbrod T, Muñoz-Planillo R
and Nuñez G: The inflammasome: A caspase-1-activation platform that
regulates immune responses and disease pathogenesis. Nat Immunol.
10:241–247. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
McMillin DW, Jacobs HM, Delmore JE, Buon
L, Hunter ZR, Monrose V, Yu J, Smith PG, Richardson PG, Anderson
KC, et al: Molecular and cellular effects of NEDD8-activating
enzyme inhibition in myeloma. Mol Cancer Ther. 11:942–951. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Milhollen MA, Traore T, Adams-Duffy J,
Thomas MP, Berger AJ, Dang L, Dick LR, Garnsey JJ, Koenig E,
Langston SP, et al: MLN4924, a NEDD8-activating enzyme inhibitor,
is active in diffuse large B-cell lymphoma models: Rationale for
treatment of NF-{kappa}B-dependent lymphoma. Blood. 116:1515–1523.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li L, Liu B, Dong T, Lee HW, Yu J, Zheng
Y, Gao H, Zhang Y, Chu Y, Liu G, et al: Neddylation pathway
regulates the proliferation and survival of macrophages. Biochem
Biophys Res Commun. 432:494–498. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Futosi K, Fodor S and Mócsai A: Neutrophil
cell surface receptors and their intracellular signal transduction
pathways. Int Immunopharmacol. 17:638–650. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shin HM, Kim MH, Kim BH, Jung SH, Kim YS,
Park HJ, Hong JT, Min KR and Kim Y: Inhibitory action of novel
aromatic diamine compound on lipopolysaccharide-induced nuclear
translocation of NF-kappaB without affecting IkappaB degradation.
FEBS Lett. 571:50–54. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang X, Ye Z, Pei Y, Qiu G, Wang Q, Xu Y,
Shen B and Zhang J: Neddylation is required for herpes simplex
virus type I (HSV-1)-induced early phase interferon-beta
production. Cell Mol Immunol. 13:578–83. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Guma M, Ronacher L, Liu-Bryan R, Takai S,
Karin M and Corr M: Caspase 1-independent activation of
interleukin-1beta in neutrophil-predominant inflammation. Arthritis
Rheum. 60:3642–3650. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang M, Medeiros BC, Erba HP, DeAngelo DJ,
Giles FJ and Swords RT: Targeting protein neddylation: A novel
therapeutic strategy for the treatment of cancer. Expert Opin Ther
Targets. 15:253–264. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Soucy TA, Smith PG, Milhollen MA, Berger
AJ, Gavin JM, Adhikari S, Brownell JE, Burke KE, Cardin DP,
Critchley S, et al: An inhibitor of NEDD8-activating enzyme as a
new approach to treat cancer. Nature. 458:732–736. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Laskin DL: Macrophages and inflammatory
mediators in chemical toxicity: A battle of forces. Chem Res
Toxicol. 22:1376–1385. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mitchell S, Vargas J and Hoffmann A:
Signaling via the NFκB system. Wiley Interdiscip Rev Syst Biol Med.
8:227–241. 2016. View Article : Google Scholar : PubMed/NCBI
|