Introduction
Primary liver cancer is one of the leading causes of
lethal malignancy worldwide, with an increasing rate of incidence
(1). In the last few decades, a
number of medical approaches, including improvements in patient
stratification and introduction of novel therapies such as
sorafenib (2), were applied in
clinical management; however, liver cancer remains a leading
life-threatening disease worldwide (3). Therefore, it is necessary to discover
effective molecule-targeting drugs against liver cancer.
DEP domain containing 1 (DEPDC1), a highly conserved
protein normally expressed only in the testes, was first reported
as being upregulated in bladder cancer cells (4). Subsequently, the ectopic expression of
DEPDC1 has also been detected in other cancer types, including
breast cancer (5), lung
adenocarcinoma (6) and hepatocellular
carcinomas (7). A study of the
bladder cancer UM-UC-3 cell line elucidated the mechanism
underlying carcinogenesis caused by DEPDC1; it interacts with zinc
finger protein 224 (ZNF224) to form a complex, which acts as a
transcription inhibitor of A20, a negative regulator of the nuclear
factor (NF)-κB pathway (8). Notably,
a peptide from the DEPDC1 protein sequence 611–628 (11R-DEP:
611–628) has the ability to disrupt the DEPDC1-ZNF224 complex
efficiently in the bladder cancer UM-UC-3 cell line and thereby
induces apoptosis in vitro and in vivo, inhibits cell
proliferation and suppresses tumor growth (8). These data indicated that this peptide is
a potential therapeutic candidate for different types of cancer
with DEPDC1 expression. A recent study determined that microRNA
(miR)-130a acts as a tumor suppressor in prostate cancer by
targeting DEPDC1 and SEC23B (9);
however, the role of miR-130a in liver cancer remains unknown.
In the present study, the role of DEPDC1 in liver
cancer was investigated and the efficacy of peptide 11R-DEP:
611–628 and miR-130a in HepG2 cells was evaluated.
Materials and methods
Antibodies and peptides
The anti-DEPDC1 antibody (cat. no. LS-C167364;
dilution 1:500) was purchased from LifeSpan BioSciences, Inc.
(Seattle, WA, USA.). The caspase-3 (cat. no. 9665S; dilution
1:1,000), cleaved-caspase-3 (cat. no. 9661; dilution 1:300) and
NF-κB antibodies (cat. no. 8242S; dilution 1:75) were purchased
from Cell Signaling Technology, Inc. (Danvers, MA, USA). The
β-actin antibody (cat. no. TA-09; dilution 1:1,000) was purchased
from OriGene Technologies, Inc. (Beijing, China). The Goat
Anti-Rabbit IgG DyLight® 488 antibody (cat. no. A32731;
dilution 1:500) was purchased from Thermo Fisher Scientific, Inc.
(Waltham, MA, USA). 11R-DEP: 611–628 peptide and the scramble
peptide were synthesized by Shanghai Top-Peptide Biotechnology Co.,
Ltd., (Shanghai, China) according to the sequences previously
reported (8).
Cell culture and transfection
HepG2 (cat. no. KCB 200507YJ) and A549 (cat. no. KCB
200434YJ) cells were purchased from the Kunming Cell Bank of the
Chinese Academy of Sciences (Kunming, Yunnan, People's Republic of
China. http://www.kmcellbank.com/). Bel-7402
(cat. no. TcHu10), SK-Hep-1 (cat. no. TcHu109) and SMMC-7721 (cat.
no. TcHu52) cells were obtained from the Cell Resource Center,
Shanghai Institute of Life Sciences, Chinese Academy of Sciences
(Shanghai, China). The cells were maintained in Dulbecco's modified
Eagle's medium (cat. no. 11965-092; Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS;
cat. no. 10437-028; Gibco; Thermo Fisher Scientific, Inc.) in a
humidified atmosphere at 37°C containing 5% CO2 and 95%
air. HepG2 cells were transiently transfected miR-130a (1.5 µg)
plasmid and empty vector GV514 (1.5 µg) (purchased from Shanghai
GenePharma Co., Ltd., Shanghai, China) using the
Lipofectamine® 3000 reagent (cat. no. L3000015; Thermo
Fisher Scientific, Inc.), according to the manufacturer's
protocols. After 72 h, the cells were harvested for cell counting
and an examination of the cell morphology and apoptosis was
conducted.
Western blot analysis
HepG2, A549, Bel-7402, SK-Hep-1 and SMMC-7721 cells
were lysed with RIPA lysis buffer (150 mM NaCl; 50 M Tris-HCl; pH,
7.5; 1% Triton X-100; 0.1% sodium deoxycholate; and 0.1% SDS)
containing 0.1% phenylmethane sulfonyl fluoride. Protein
concentration of cell lysates was measured by BCA kit (Beyotime
Institute of Biotechnology, Shanghai, China). Equal protein lysate
samples (30 µg) were separated by 10% SDS-PAGE and then transferred
onto polyvinylidene fluoride membrane, which was blocked in 5%
nonfat milk at room temperature for 2 h, followed by incubation
with primary antibodies at 4°C overnight. The membrane was then
incubated with secondary antibody (horseradish
peroxidase-conjugated goat anti-rabbit immunoglobulin G (IgG;
catalog no. G-21234, dilution 1:5,000) or goat anti-mouse IgG
(catalog no. G-21040, dilution 1:5,000) antibodies (Pierce; Thermo
Fisher Scientific, Inc.) at room temperature for 1 h. Blots were
developed using an enhanced chemiluminescence kit (Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China). The
densitometric analysis of bands was performed using FluorChem SA
(6.0.0) (Informer Technologies, Inc, Chicago, IL, USA).
Cell proliferation assay
The MTT dissolved in 0.1 M PBS at a concentration of
5 mg/ml was purchased from Beijing Solarbio Science &
Technology Co., Ltd. The HepG2 cells were seeded onto 96-well
plates in medium containing 2% FBS in the presence of 11R-DEP:
611–628 peptide or scramble peptide at 0, 0.5, 1, 2 and 3 µM. The
medium was replaced with medium containing same concentration of
peptides every 24 h for 4 days, and then the proliferation of the
cells was detected. A total of 10 µl MTT was added into each well
and then incubated at 37°C for 4 h. Finally, the DMEM culture
medium supplemented with 2% FBS was discarded and 150 µl dimethyl
sulfoxide was added into each well to dissolve the crystals
completely. Absorbance was measured with a microplate reader at 490
nm.
Immunofluorescence staining
HepG2 cells were fixed with 4% paraformaldehyde for
30 min at room temperature following treatment with 11R-DEP:
611–628 peptide or scramble peptide at 1, 6 and 12 h. Following
this, the cells were permeabilized with a 5% blocking solution (0.1
M PBS; pH, 7.4 containing 5% bovine serum albumin; and 0.3% Triton
X-100) for 2 h at room temperature. The sections were subsequently
incubated with the rabbit anti-NF-κB antibody overnight at 4°C in a
humid chamber. Following three washes with PBS, the cells were
incubated with Goat Anti-Rabbit IgG DyLight 488 antibody for 2 h at
room temperature. The cell nuclei were counterstained with DAPI for
10 min at room temperature. Images of the cells were captured with
a fluorescence microscope (IX71; Olympus Corporation, Tokyo, Japan)
at a magnification of ×200.
Terminal
deoxynucleotidyl-transferase-mediated dUTP nick end labeling
(TUNEL) staining
The HepG2 cells (8×104/well) were seeded
into wells of the chamber slides one day prior to the experiments.
Cells were treated with 3 µM 11R-DEP: 611–628 peptide for 3 h at
37°C. Cells were fixed in freshly diluted 1% paraformaldehyde in
PBS, pH 7.4 for 10 min at room temperature. TUNEL staining was
performed with the Apoptosis Detection kit (cat. no. S7100; EMD
Millipore, Billerica, MA, USA) according to the manufacturer's
protocols. The nuclei were counterstained in 0.1% hematoxylin for 1
min at room temperature, and then rinsed with running tap water for
5 min. The slides were mounted with mounting medium Permount (cat.
no. SP15-500, Thermo Fisher Scientific, Inc.), and a total of 10
randomly selected fields of view were observed under a fluorescent
microscope (magnification, ×200).
Flow cytometric analysis
The HepG2 Cells (2.5×105) were
transfected (the transfection protocol was performed as previously
described) with the miR-130a expression plasmid or a control
plasmid. After 72 h, the cells were harvested and washed with PBS
twice. The cells then were stained with the FITC Annexin V/Dead
Cell kit (Merck KGaA, Darmstadt, Germany) for 20 min at room
temperature in the dark, according to the manufacturer's protocols.
Finally, cells were analyzed using a flow cytometer
(Muse® Cell Analyzer, MuseSoft_V1.5.0.0; Merck
Millipore, Darmstadt, Germany).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from the HepG2 cells using
the TRIzol® reagent (Takara Bio Inc., Otsu, Japan). For
RT-qPCR, the first-strand cDNA was reverse transcribed from 2 µg
total RNA using the Reverse Transcription kit (Tiangen Biotech Co.,
Ltd., Beijing, China) according to the manufacturer's protocol. The
following primers were used: A20 forward,
5′-CGTCCAGGTTCCAGAACACCATTC-3′ and reverse,
5′-TGCGCTGGCTCGATCTCAGTTG-3′; DEPDC1 forward,
5′-GAGGTCACTGATGATACATAC-3′ and reverse,
5′-TGCAGTCTGTAAGTAAGAGG-3′; and GAPDH forward,
5′-CAGGAGGCATTGCTGATGAT-3′ and reverse, 5′-GAAGGCTGGGGCTCATTT-3′.
To analyze miR-130a expression, the cDNA was amplified according to
the manufacturer's protocol using the Mir-X miRNA First Strand
Synthesis kit (#638313, Takara Bio Inc.). qRT-PCR was conducted
according to the manufacturer's instructions of the Mir-X miRNA
qRT-PCR SYBR kit (#638314, Takara Bio Inc.). The U6 (forward,
5′-GGAACGATACAGAGAAGATTAGC-3′ and reverse,
5′-TGGAACGCTTCACGAATTTGCG-3′) and miR-130a special primer (forward,
5′-GGCCAGAGCTCTTTTCACAT-3′ and reverse,
5′-CGGCCAATGCCCTTTTAACAT-3′) were purchased from Takara Bio Inc.
The relative level of gene expression was quantified using the
2−∆∆Cq method (10),
normalized to GAPDH or U6, and expressed as the fold induction of
the control. PCR conditions were as followed: 95°C for 10 sec; 40
cycles: 95°C for 5 sec, and 60°C for 20 sec.
Statistical analysis
The values are expressed as the mean ± standard
deviation. Comparisons between two groups were conducted using the
Student's t-test and comparisons among multiple groups were
performed using one-way analysis of variance, followed by the
Student-Newman-Keuls post hoc test. SPSS 13.0 software (SPSS,
Chicago, IL, USA) and GraphPad Prism 5.0 (GraphPad Software, Inc.,
La Jolla, CA, USA) software were utilized for statistical analyses.
All experiments were performed at least three independent times,
and representative data are presented. P<0.05 was considered to
indicate a statistically significant difference.
Results
DEPDC1 is expressed in HepG2
cells
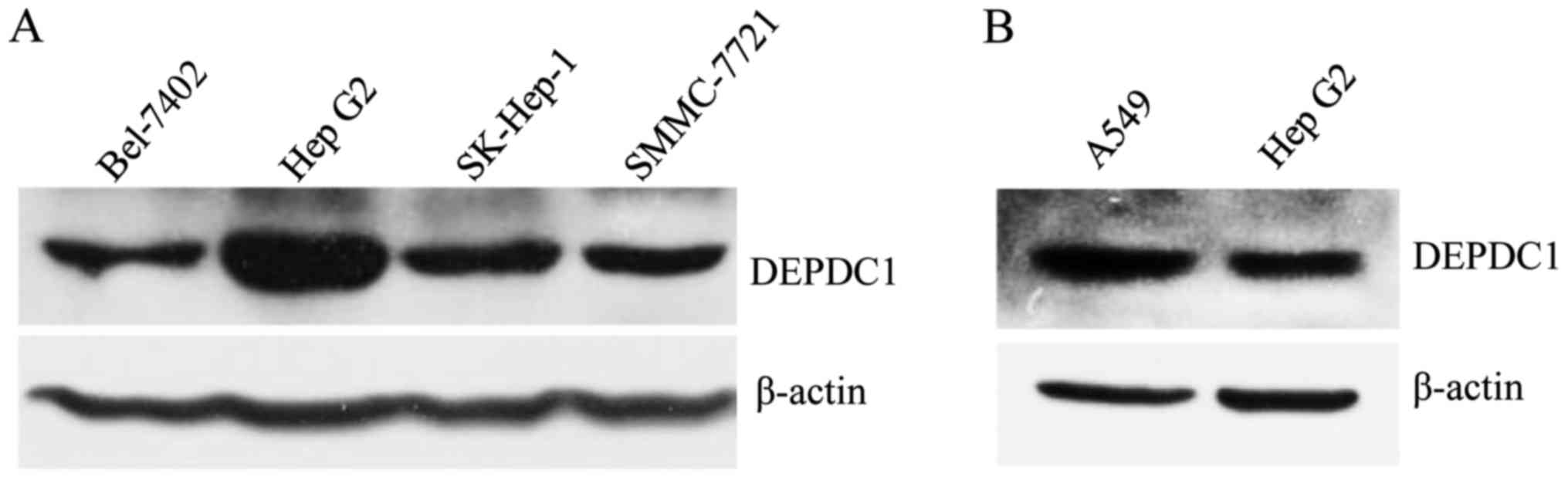
To understand the role of DEPDC1 in liver cancer,
DEPDC1 expression at a protein level in HepG2 cells and other liver
cancer cell lines was examined using a western blotting assay. To
the best of our knowledge, DEPDC1 expression has not been detected
in liver cancer cell lines in any published report to date;
therefore, the A549 cell line was used as a positive control due to
the role of DEPDC1 in A549 having been recently reported and due to
the fact that samples were available (11). The results demonstrated that DEPDC1 is
expressed in all of the liver cancer cell lines used, and its
protein levels in HepG2 cells are the highest among the examined
liver cancer cell lines (Fig. 1A),
but lower than the A549 lung cancer cells (Fig. 1B); therefore, the HepG2 cell line was
used to conduct further experiments.
11R-DEP: 611–628 treatment inhibits
HepG2 cell proliferation
A study reported that DEPDC1 has the ability to
promote cell proliferation by forming a complex with ZNF224, which
inhibits associated gene expression in the bladder cancer UM-UC-3
cell line (8). It was hypothesized
that DEPDC1 functions in the same manner in HepG2 cells, and this
hypothesis was investigated by treating HepG2 cells with 11R-DEP:
611–628, which is capable of disrupting the DEPDC1-ZNF224 complex
in the bladder cancer UM-UC-3 cell line (8), and in the lung cancer A549 cell line
(11). Cell proliferation was then
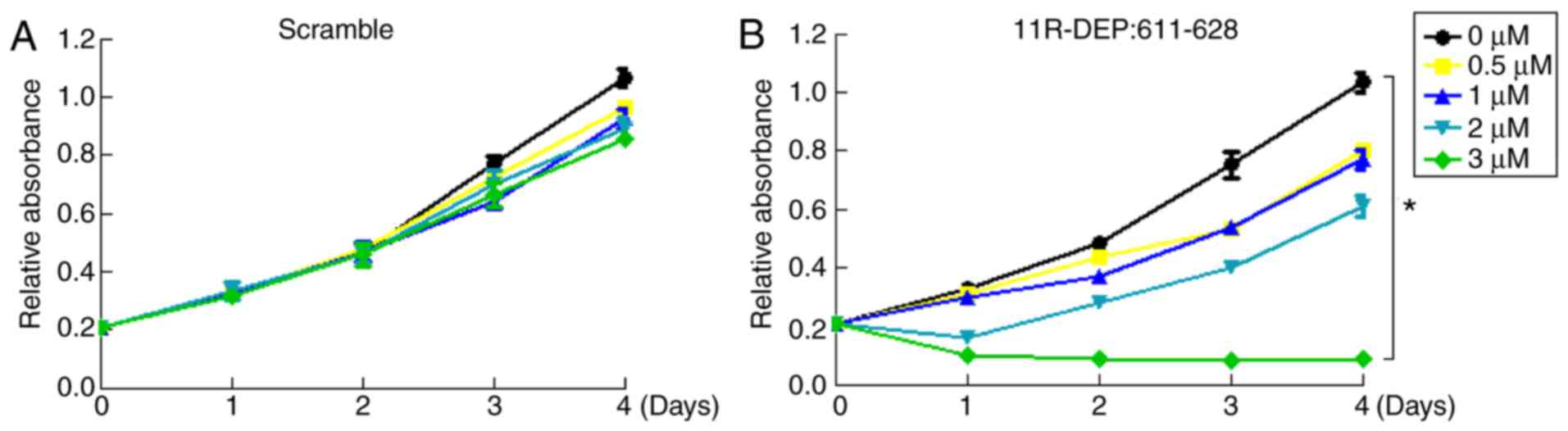
analyzed using an MTT assay. Compared with the scramble peptide
(Fig. 2A), 11R-DEP: 611–628 (3 µM)
significantly inhibited HepG2 cell proliferation from day 1–4
(P<0.05) (Fig. 2B); therefore, 3
µM was selected as the optimal concentration for further
experiments. Since the 11R-DEP: 611–628 peptide was designed to
disrupt the DEPDC1-ZNF224 complex (8), this peptide inhibited the proliferation
of HepG2 cells, indicating that the DEPDC1-ZNF224 complex serves a
key role in promoting HepG2 cell proliferation. To summarize,
DEPDC1 promotes HepG2 cell proliferation via interacting with
ZNF224.
11R-DEP: 611–628 treatment induces the
apoptosis of HepG2 cells
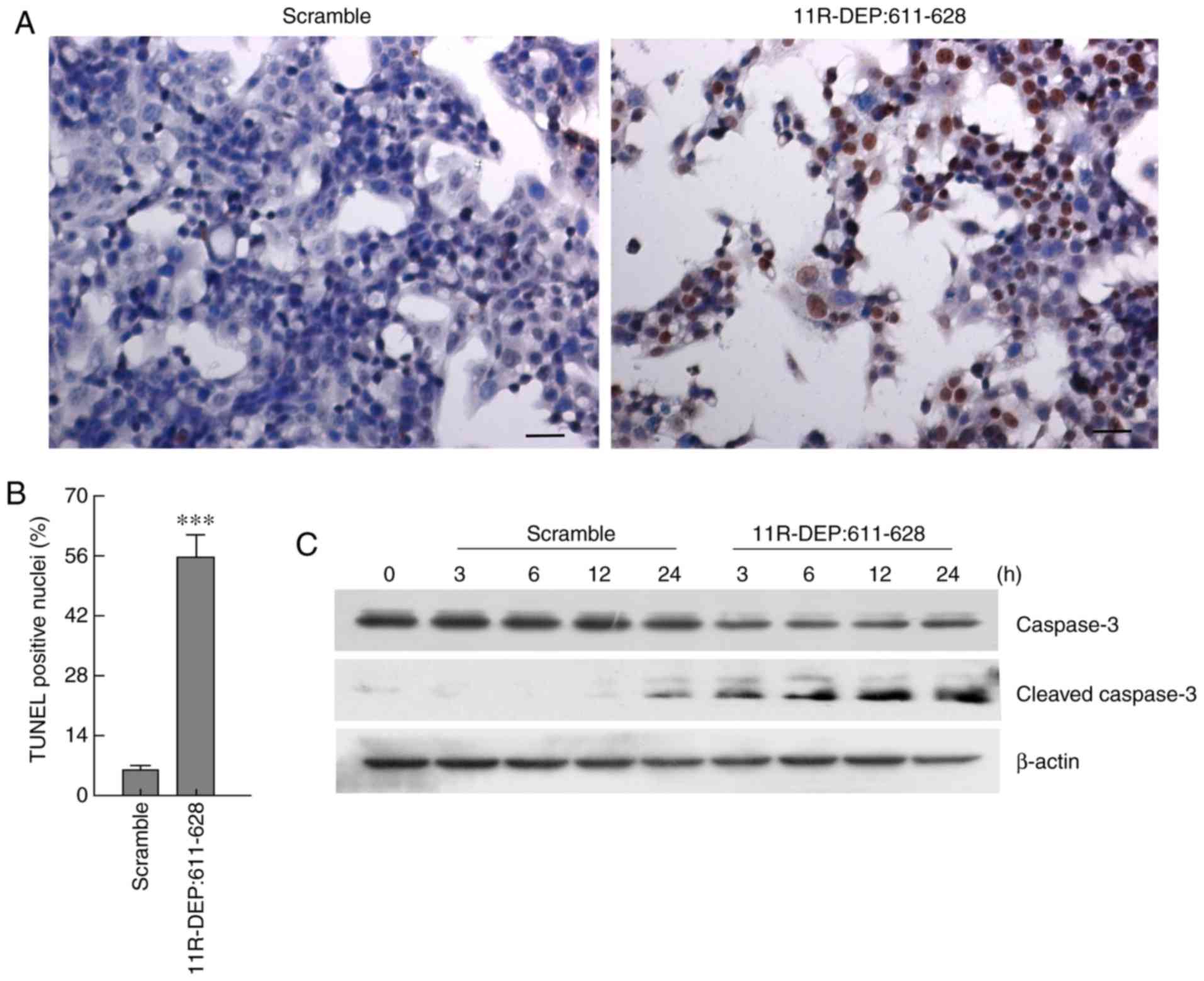
To further reveal how DEPDC1 affects cell
proliferation, HepG2 cells were treated with 3 µM 11R-DEP: 611–628
peptide for 3, 6, 12, and 24 h; following this, apoptosis was
analyzed by TUNEL staining, and the apoptotic marker caspase-3
protein, which is cleaved when apoptosis occurs, was detected via
western blotting (12). The results
demonstrated that treatment with 11R-DEP: 611–628 causes a
significant increase (P<0.001) in TUNEL positive cells (Fig. 3A and B), a decrease in caspase-3
protein levels and an increase in the cleaved caspase-3 protein
level, compared with the scramble peptide (Fig. 3C). These results demonstrated that
11R-DEP: 611–628 treatment enhances apoptosis, indicating that
DEPDC1 inhibits apoptosis in HepG2 cells.
11R-DEP: 611–628 treatment results in
the blocking of NF-κB nuclear translocation in HepG2 cells
A previous study indicated that DEPDC1 functions
through regulating A20 expression to inhibit NF-κB function in
bladder cancer cell lines (8), and
thus the A20 mRNA level should be increased following the
disruption of the DEPDC1-ZNF224 complex with 11R-DEP: 611–628
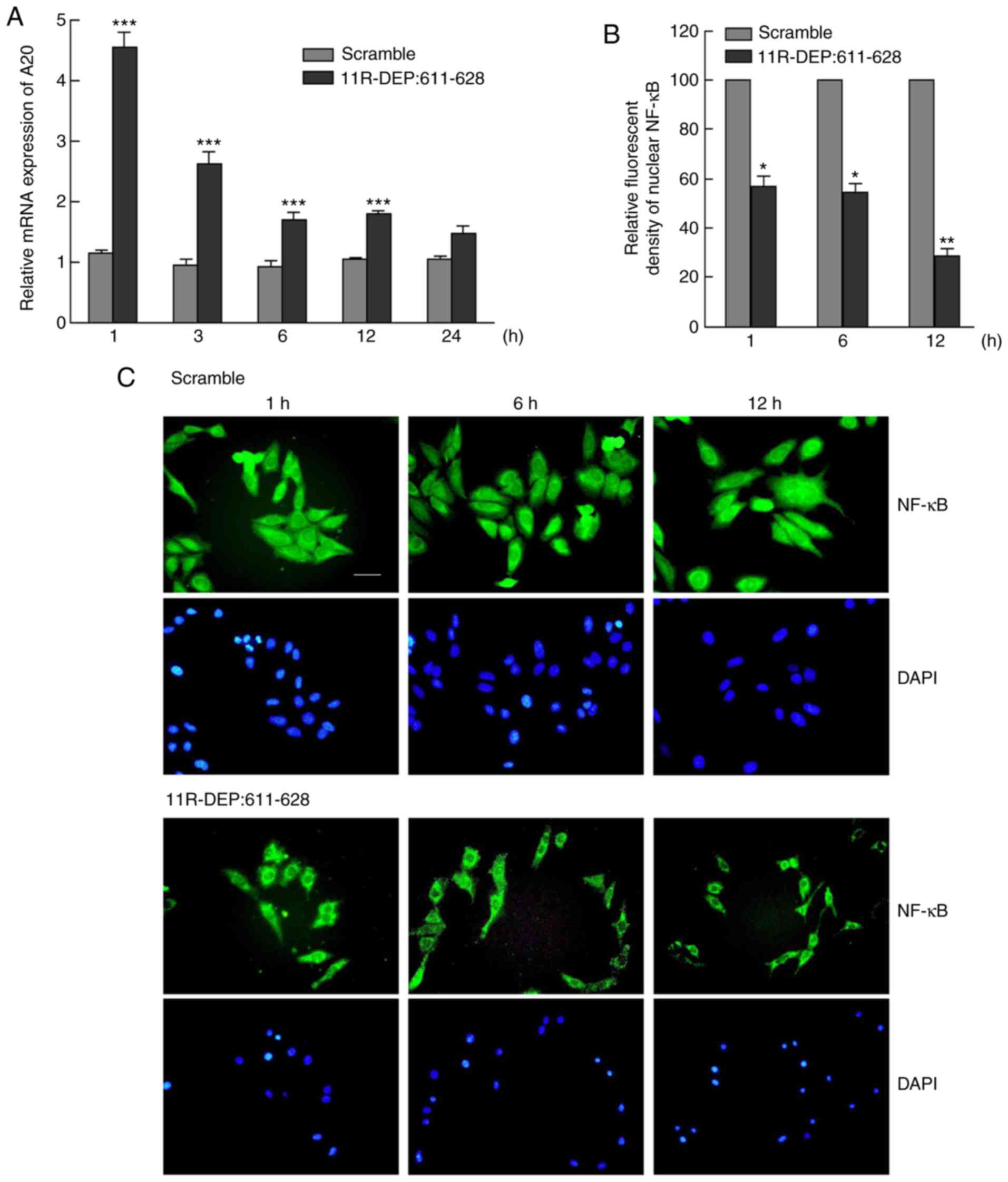
(8). Therefore, it was investigated
whether DEPDC1 serves the same role in HepG2 cells. The A20 mRNA
expression levels were examined by RT-qPCR and the results
demonstrated a significant increase (P<0.05) (Fig. 4A) between 1 and 24 h following
treatment with the 11R-DEP: 611–628 peptide, compared with the
scramble peptide. A20 protein expression levels were detected with
commercial antibodies, but the protein level was not obtained due
to poor antibody quality (data no shown). However, A20 mRNA
increase following 11R-DEP: 611–628 treatment should be sufficient
to support the theory that A20 expression is regulated by the
DEPDC1-ZNF224 complex. In addition, the nuclear NF-κB expression
was significantly decreased (P<0.05) in 11R-DEP: 611–628
peptide-treated cells, compared with scramble peptide-treated cells
(Fig. 4B and C). These data indicated
that the 11R-DEP: 611–628 peptide treatment induces A20 expression,
which blocks NF-κB (p65) nuclear translocation.
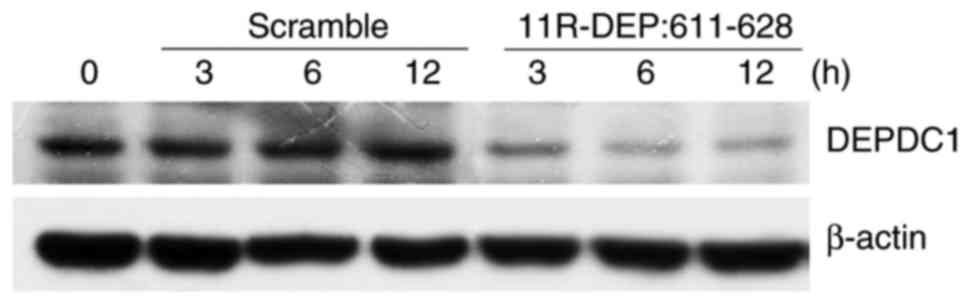
11R-DEP: 611–628 treatment
downregulates DEPDC1 protein expression
DEPDC1 is upregulated in a variety of cancer types
such as breast (5), lung cancer
(6) and bladder cancer (8); however, the mechanism underlying the
regulation of DEPDC1 expression in cancer cells remains unknown. It
was hypothesized that DEPDC1 expression may be regulated by the
DEPDC1-ZNF224 complex. To investigate this theory, HepG2 cells were
treated with 11R-DEP: 611–628 peptide and then the DEPDC1 protein
expression levels were examined. A notable decrease in DEPDC1
protein expression was observed in cells treated with 11R-DEP:
611–628 peptide (Fig. 5) compared
with those in cells treated with the scramble peptide, indicating
that DEPDC1 expression is regulated by the DEPDC1-ZNF224
complex.
miR-130a modulates proliferation and
apoptosis by targeting DEPDC1 in HepG2 cells
It was reported that the overexpression of miR-130a
induces apoptosis and inhibits proliferation in prostate cancer
cells by targeting DEPDC1 and SEC23B, and demonstrated that
miR-130a inhibits DEPDC1 MRE1-directed luciferase activity
(9). Based on these data, miR-130a
was used to investigate the role of DEPDC1 in HepG2 cells. HepG2
cells were transfected with miR-130a expressing plasmids, and the
effects of miR-130a expression were examined. The results
demonstrated that, compared with the control group, the total cell
number was decreased, whilst the number of apoptotic cells was
increased in the miR-130a expression group (Fig. 6A-C). DEPDC1 expression at the protein
and mRNA level was significantly reduced (P<0.0005) (Fig. 6D and E), and A20 expression was
significantly increased (P<0.05) at 72 h post-transfection,
compared with the control group (Fig.
6F). These results indicated that miR-130a may regulate HepG2
cell apoptosis by targeting DEPDC1.
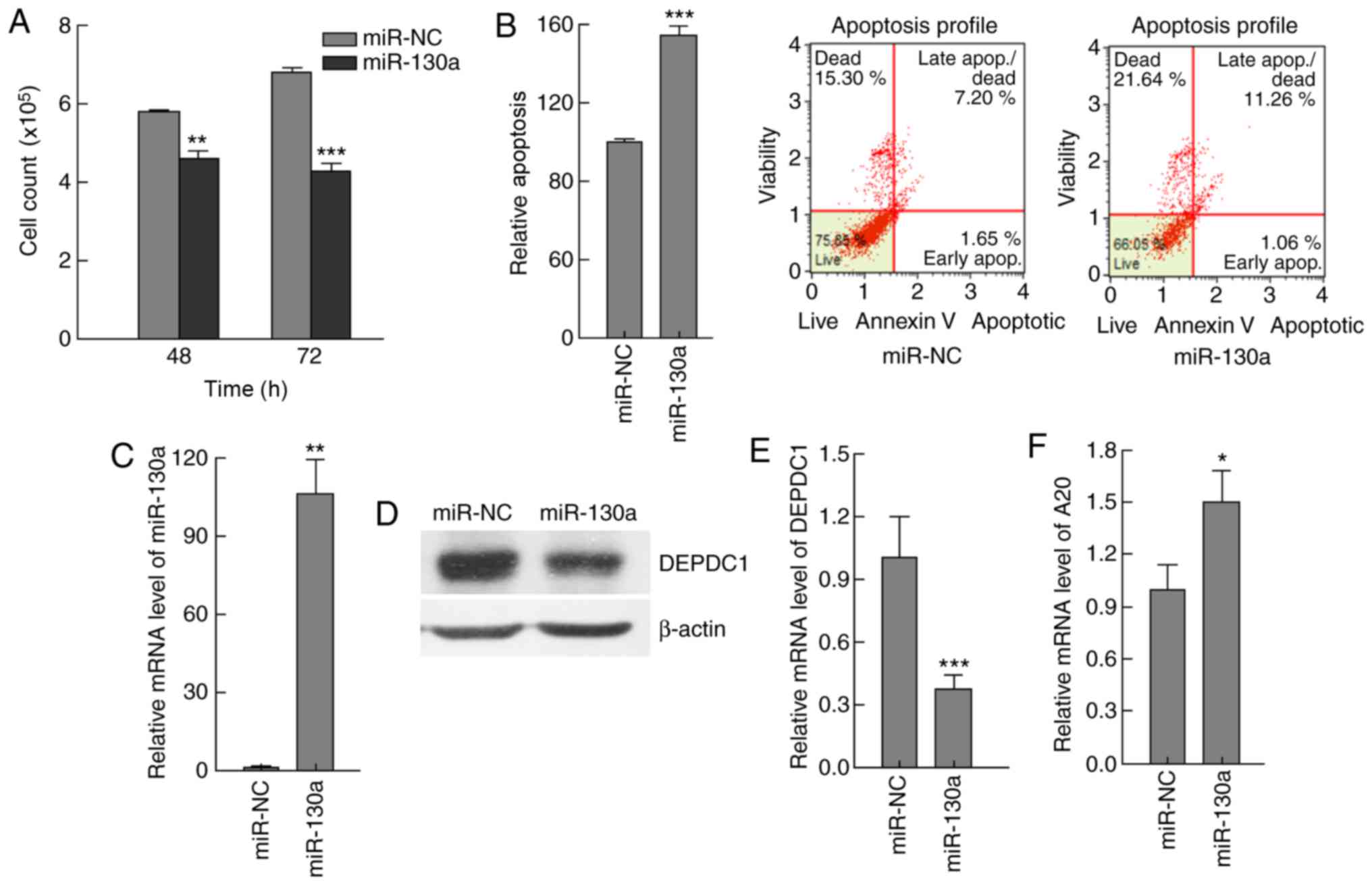 | Figure 6.miR-130a modulated apoptosis by
downregulating DEPDC1 expression in HepG2 cells. HepG2 cells were
transfected with miR-130a expressing plasmids or NC plasmids. (A)
Cells were counted at 48 and 72 h, and the results indicated that
miR-130a expression resulted in a significant decrease in the cell
number, compared with miR-NC. (B) Apoptosis was analyzed by flow
cytometry, and the results demonstrated that miR-130a expression
enhanced apoptosis, compared with miR-NC. (C) miR-130a expression
in cells transfected with miR-130a expressing plasmids or NC
plasmids was examined by RT-qPCR, and the results demonstrated that
miR-130a expression increased ~100-fold in cells transfected with
miR-130a expressing plasmids, compared with miR-NC. (D) The DEPDC1
protein levels were significantly reduced 72 h post-transfection as
demonstrated by western blot analysis data, compared with miR-NC.
(E) The DEPDC1 mRNA levels were significantly reduced 72 h
post-transfection as demonstrated by RT-qPCR data, compared with
miR-NC. (F) A20 expression levels were increased significantly 72 h
post-transfection, compared with miR-NC. These results indicated
that miR-130a has the ability to regulate HepG2 apoptosis by
targeting DEPDC1. The data are presented as the mean ± standard
deviation. *P<0.05, **P<0.01, ***P<0.001. RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; NC, control;
miR-130a, microRNA 130a; DEPDC1, DEP domain containing 1. |
Discussion
It was previously reported that DEPDC1 is highly
expressed in bladder cancer cells, and downregulating its
expression with siRNA significantly suppressed the proliferation of
bladder cancer cells (4). DEPDC1
interacts with ZNF224 to form a complex and inhibits the expression
of A20, resulting in an increase in nuclear NF-κB and an inhibition
of apoptosis in bladder cancer cells (8).
In the present study, the role of DEPDC1 was first
investigated in HepG2 cells. It was demonstrated that interfering
with DEPDC1-ZNF224 complex formation using the 11R-DEP: 611–628
peptide resulted in decreased cell proliferation and enhanced
apoptosis. A comprehensive review determined that NF-κB was
constitutively elevated in a variety of tumor types, including
hematological and solid tumors (13).
Continuously activated NF-κB signaling results in the upregulation
of anti-apoptosis and pro-proliferation genes, which is notably
correlated with tumorigenesis and tumor progression (14–16). In
the present study, it was determined that the DEPDC1-ZNF224 complex
suppresses A20 expression and thus activates the NF-κB signaling
pathway, eventually resulting in the inhibition of apoptosis in
HepG2 cells (17). To the best of our
knowledge, the present study was the first to study the role of
DEPDC1 in HepG2 cells, and to demonstrate the efficacy of 11R-DEP:
611–628 and miR-130a in HepG2 cells; these data may ultimately
benefit patients with liver cancer. Performing further experiments
in other liver cancer cell lines will provide further evidence
regarding the role of DEPDC1 in liver cancer and the potential
clinical application of 11R-DEP: 611–628 and miR-130a. Notably, the
results demonstrated that the 11R-DEP: 611–628 peptide treatment
resulted in a decrease in DEPDC1 protein, indicating that the
DEPDC1-ZNF224 complex serves a role in regulating DEPDC1 protein
levels; however, the underlying mechanisms require further
elucidation in the future. The experiments analyzing the
characteristics of the DEPDC1 promoter may be beneficial to
elucidate the mechanisms underlying the regulation of DEPDC1
expression by the DEPDC1-ZNF224 complex.
miRNAs, a class of endogenous 22-nucleotide
non-coding RNAs, inhibit gene expression by imperfectly pairing
with the 3′-untranslated region of the target genes (18). It was reported that miRNAs, including
miR-9, miR-21 and miR-224, regulate NF-κB activity by targeting the
family members of NF-κB or its upstream signaling molecules
(19–22). miR-130a suppresses cell proliferation
and induces apoptosis by targeting DEPDC1 and SEC23B in prostate
cancer cells (9). The present study
indicated that miR-130a regulates apoptosis through the NF-κB
signaling pathway in HepG2 cells by targeting DEPDC1, although
luciferase assay and rescue experiments are required to further
confirm the association between miR-130a and DEPDC1; therefore,
miR-130a may be a potential therapeutic factor for liver cancer
with DEPDC1 expression.
The limitation of the present study is that only the
HepG2 cell line was used to conduct the experiments. The HepG2 cell
line was originally identified as a hepatocellular carcinoma cell
line, but was later confirmed to be a hepatoblastoma-derived cell
line (23); however, the
misidentification of HepG2 does not affect the outcomes and
conclusions of the present study.
In summary, the data demonstrated for the first time
that 11R-DEP: 611–628 and miR-130a are capable of inhibiting
proliferation by inducing apoptosis through the NF-κB signaling
pathway in HepG2 cells; therefore, DEPDC1 could be a therapeutic
target for the treatment of liver cancer with DEPDC1
expression.
Acknowledgements
Not applicable.
Funding
The present study was supported in part by the
National Natural Science Foundation of China (grant no. 81560453),
Natural Science Foundation of Guangxi (grant no.
2015GXNSFAA139178), the Guangxi Health and Family Planning
Commission (grant no. S2015-34), the Lijiang Scholar Award and the
‘Sphingolipids and Related Diseases’ Program for Innovative
Research Team of Guilin Medical University. G. H. was supported by
the Hundred Talents Program of Guangxi.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
AL, QW and GFH performed the experiments. AL, JJ and
GJH analyzed and interpreted the data. AL, QW, JJ and GJH wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bruix J, Reig M and Sherman M:
Evidence-based diagnosis, staging, and treatment of patients with
hepatocellular carcinoma. Gastroenterology. 150:835–853. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chacko S and Samanta S: ‘Hepatocellular
carcinoma: A life-threatening disease’. Biomed Pharmacother.
84:1679–1688. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kanehira M, Harada Y, Takata R, Shuin T,
Miki T, Fujioka T, Nakamura Y and Katagiri T: Involvement of
upregulation of DEPDC1 (DEP domain containing 1) in bladder
carcinogenesis. Oncogene. 26:6448–6455. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kretschmer C, Sterner-Kock A, Siedentopf
F, Schoenegg W, Schlag PM and Kemmner W: Identification of early
molecular markers for breast cancer. Mol Cancer. 10:152011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Okayama H, Kohno T, Ishii Y, Shimada Y,
Shiraishi K, Iwakawa R, Furuta K, Tsuta K, Shibata T, Yamamoto S,
et al: Identification of genes upregulated in ALK-positive and
EGFR/KRAS/ALK-negative lung adenocarcinomas. Cancer Res.
72:100–111. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan SG, Liao WJ, Yang JJ, Huang GJ and
Huang ZQ: DEP domain containing 1 is a novel diagnostic marker and
prognostic predictor for hepatocellular carcinoma. Asian Pac J
Cancer Prev. 15:10917–10922. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Harada Y, Kanehira M, Fujisawa Y, Takata
R, Shuin T, Miki T, Fujioka T, Nakamura Y and Katagiri T:
Cell-permeable peptide DEPDC1-ZNF224 interferes with
transcriptional repression and oncogenicity in bladder cancer
cells. Cancer Res. 70:5829–5839. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ramalho-Carvalho J, Martins JB, Cekaite L,
Sveen A, Torres-Ferreira J, Graça I, Costa-Pinheiro P, Eilertsen
IA, et al: Epigenetic disruption of miR-130a promotes prostate
cancer by targeting SEC23B and DEPDC1. Cancer Lett. 385:150–159.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Q, Li A, Jin J and Huang G: Targeted
interfering DEP domain containing 1 protein induces apoptosis in
A549 lung adenocarcinoma cells through the NF-κB signaling pathway.
Onco Targets Ther. 10:4443–4454. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wolf BB, Schuler M, Echeverri F and Green
DR: Caspase-3 is the primary activator of apoptotic DNA
fragmentation via DNA fragmentation factor-45/inhibitor of
caspase-activated DNase inactivation. J Biol Chem. 274:30651–30656.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pacifico F and Leonardi A: NF-kappaB in
solid tumors. Biochem Pharmacol. 72:1142–1152. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Karin M: Nuclear factor-kappaB in cancer
development and progression. Nature. 441:431–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kasibhatla S, Brunner T, Genestier L,
Echeverri F, Mahboubi A and Green DR: DNA damaging agents induce
expression of Fas ligand and subsequent apoptosis in T lymphocytes
via the activation of NF-kappa B and AP-1. Mol Cell. 1:543–551.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Meteoglu I, Erdogdu IH, Meydan N, Erkus M
and Barutca S: NF-KappaB expression correlates with apoptosis and
angiogenesis in clear cell renal cell carcinoma tissues. J Exp Clin
Cancer Res. 27:532008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
López-Terrada D, Cheung SW, Finegold MJ
and Knowles BB: Hep G2 is a hepatoblastoma-derived cell line. Hum
Pathol. 40:1512–1515. 2009. View Article : Google Scholar
|
|
18
|
Gu X, Li A, Liu S, Lin L, Xu S, Zhang P,
Li S, Li X, Tian B, Zhu X and Wang X: MicroRNA124 regulated neurite
elongation by targeting OSBP. Mol Neurobiol. 53:6388–6396. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bazzoni F, Rossato M, Fabbri M, Gaudiosi
D, Mirolo M, Mori L, Tamassia N, Mantovani A, Cassatella MA and
Locati M: Induction and regulatory function of miR-9 in human
monocytes and neutrophils exposed to proinflammatory signals. Proc
Natl Acad Sci USA. 106:5282–5287. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma X, Becker Buscaglia LE, Barker JR and
Li Y: MicroRNAs in NF-kappaB signaling. J Mol Cell Biol. 3:159–166.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Niu J, Shi Y, Tan G, Yang CH, Fan M,
Pfeffer LM and Wu ZH: DNA damage induces NF-κB-dependent
microRNA-21 up-regulation and promotes breast cancer cell invasion.
J Biol Chem. 287:21783–21795. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Scisciani C, Vossio S, Guerrieri F,
Schinzari V, de Iaco R, D'Onorio de Meo P, Cervello M, Montalto G,
Pollicino T, Raimondo G, et al: Transcriptional regulation of
miR-224 upregulated in human HCCs by NFκB inflammatory pathways. J
Hepatol. 56:855–861. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
López-Terrada D, Cheung SW, Finegold MJ
and Knowles BB: Hep G2 is a hepatoblastoma-derived cell line. Hum
Pathol. 40:1512–1515. 2009. View Article : Google Scholar
|